
Characterization of l-Arabinose Isomerase from Klebsiella pneumoniae and Its Application in the Production of d-Tagatose from d-Galactose


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism, Plasmid, and Gene Cloning
2.2. Culture Conditions and Enzyme Purification
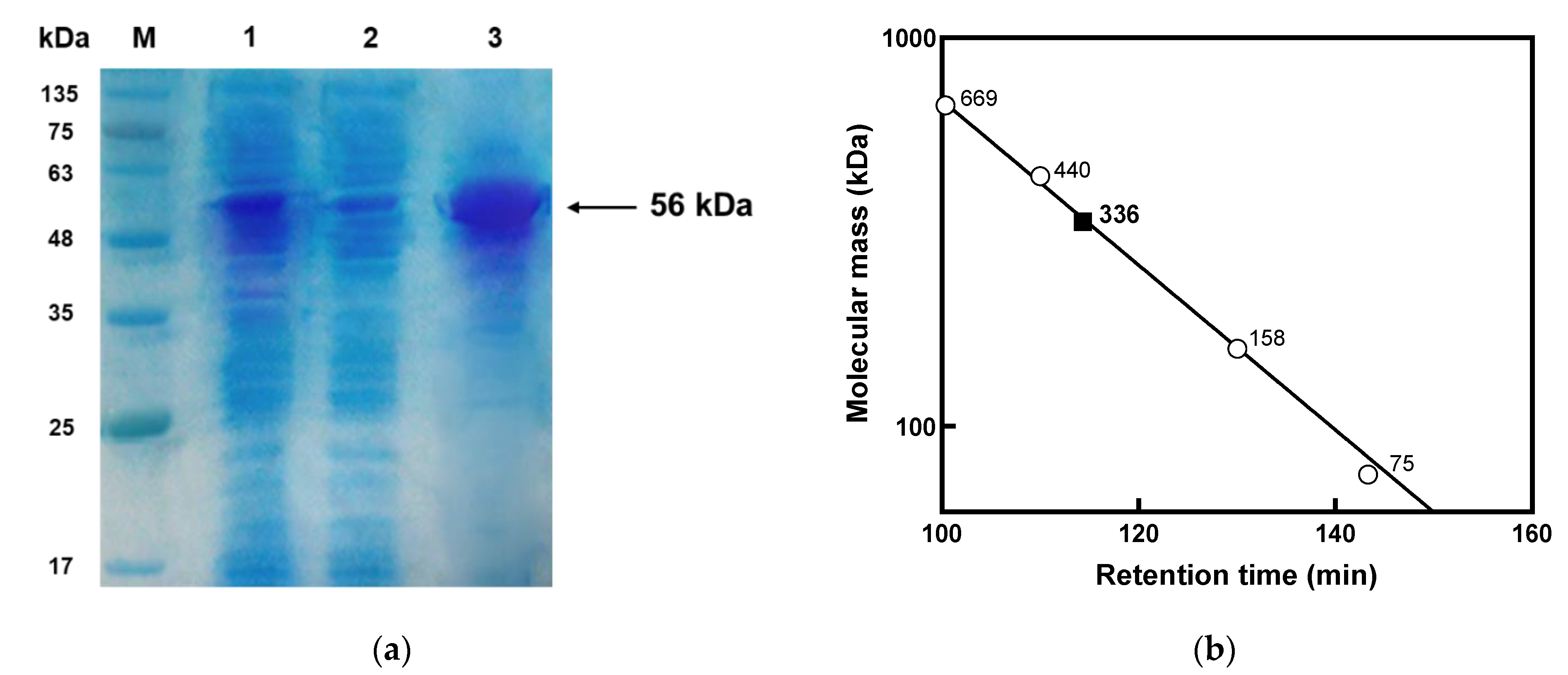
2.3. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE) Analysis and Gel Filtration
2.4. Enzyme Assay
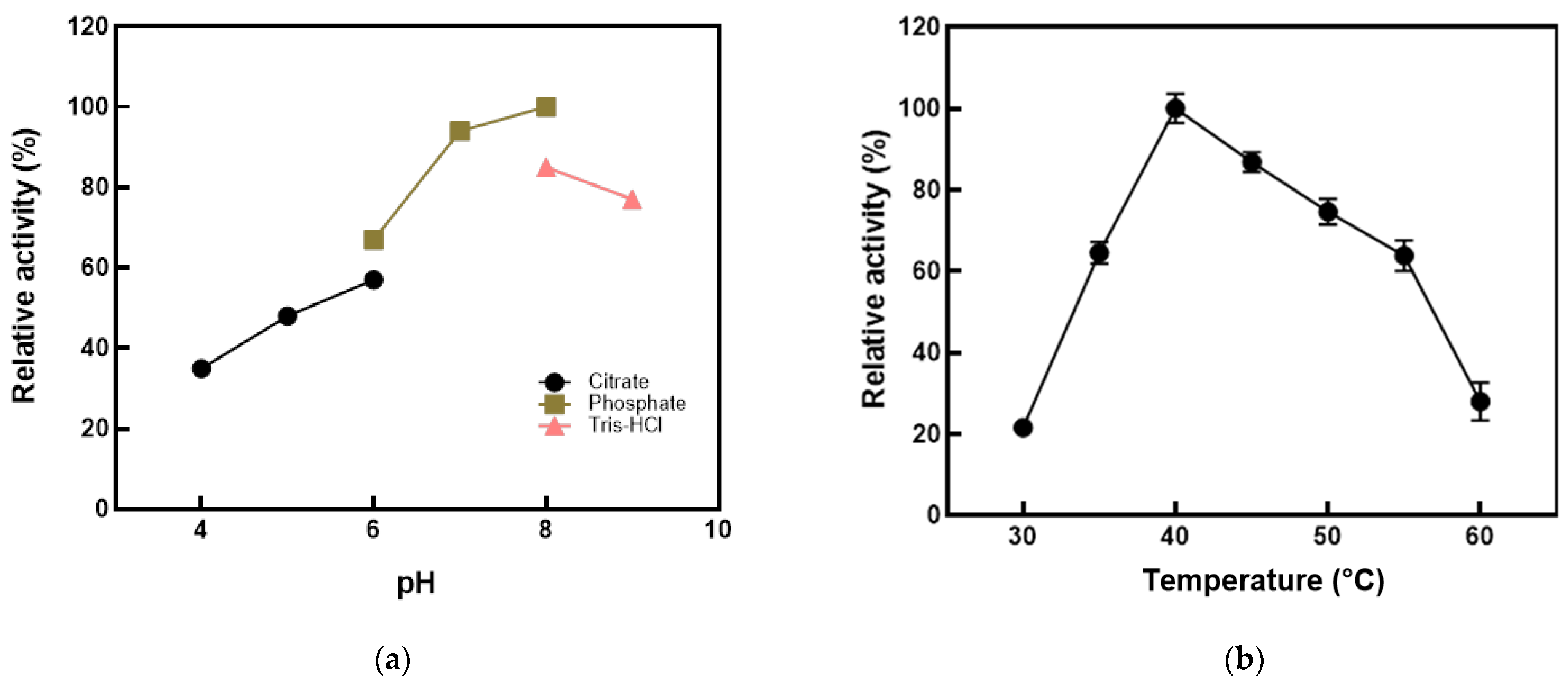
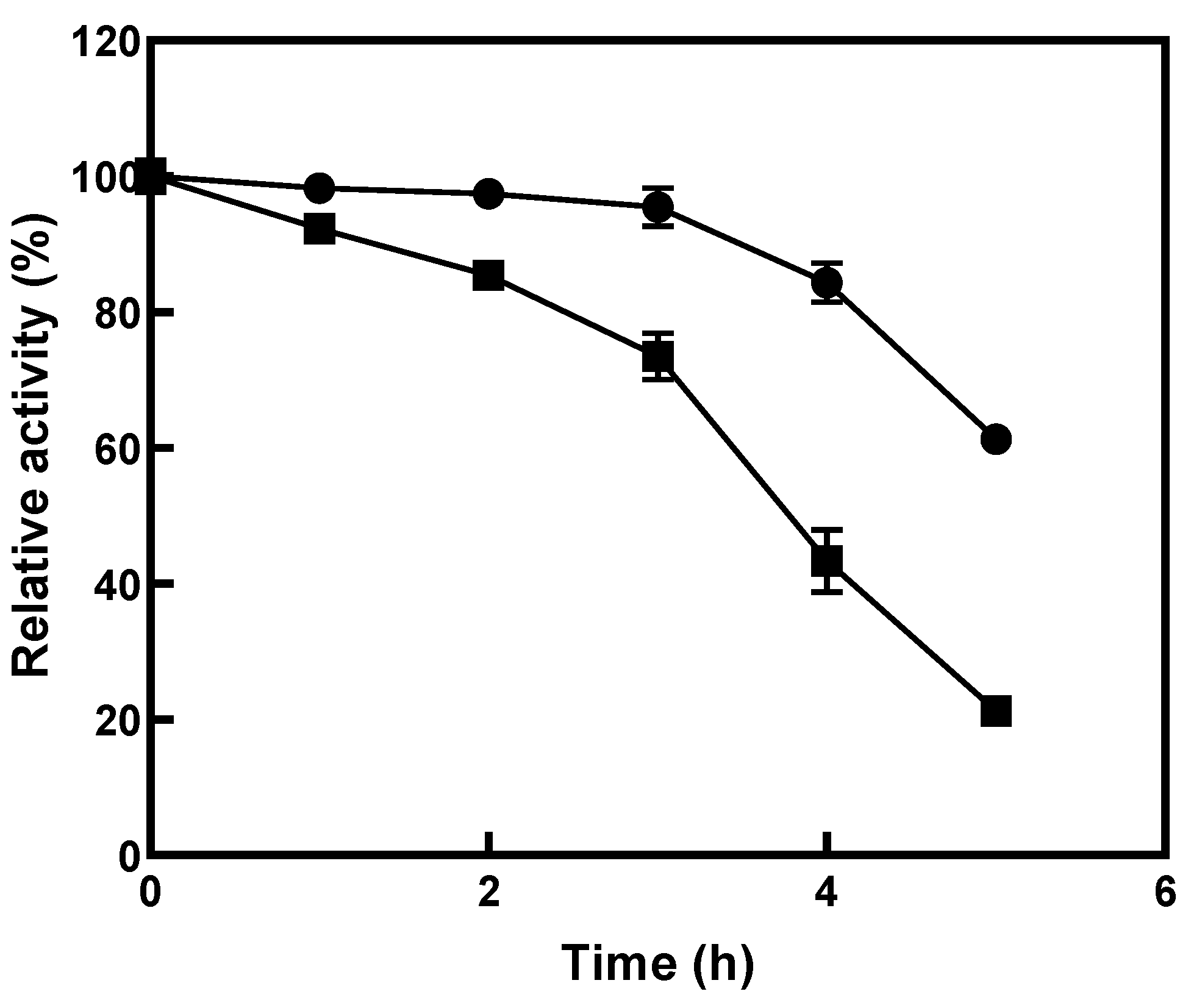
2.5. Effects of pH, Temperature, and Metal Ions
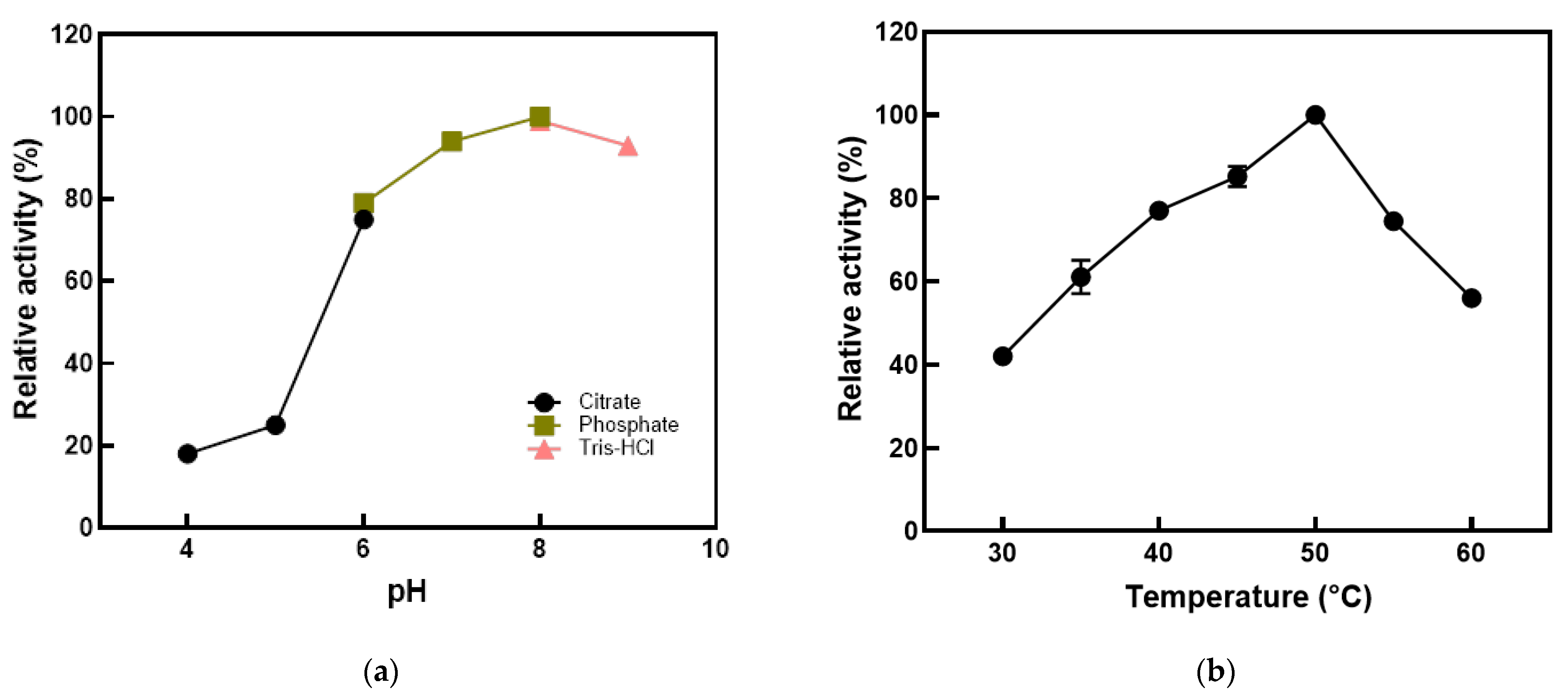
2.6. Optimization of Reaction Conditions Using E. coli Cells Expressing l-Arabinose Isomerase
2.7. Analytical Methods
3. Results and Discussion
3.1. Gene Cloning, Purification, and Molecular Mass Determination of l-Arabinose Isomerase from K. pneumoniae
3.2. Effects of Environmental Factors on the Activity of l-Arabinose Isomerase from K. pneumoniae
3.3. Substrate Specificity of l-Arabinose Isomerase from K. pneumoniae
3.4. Optimization of Reaction Conditions Using E. coli Cells Expressing l-Arabinose Isomerase from K. pneumoniae
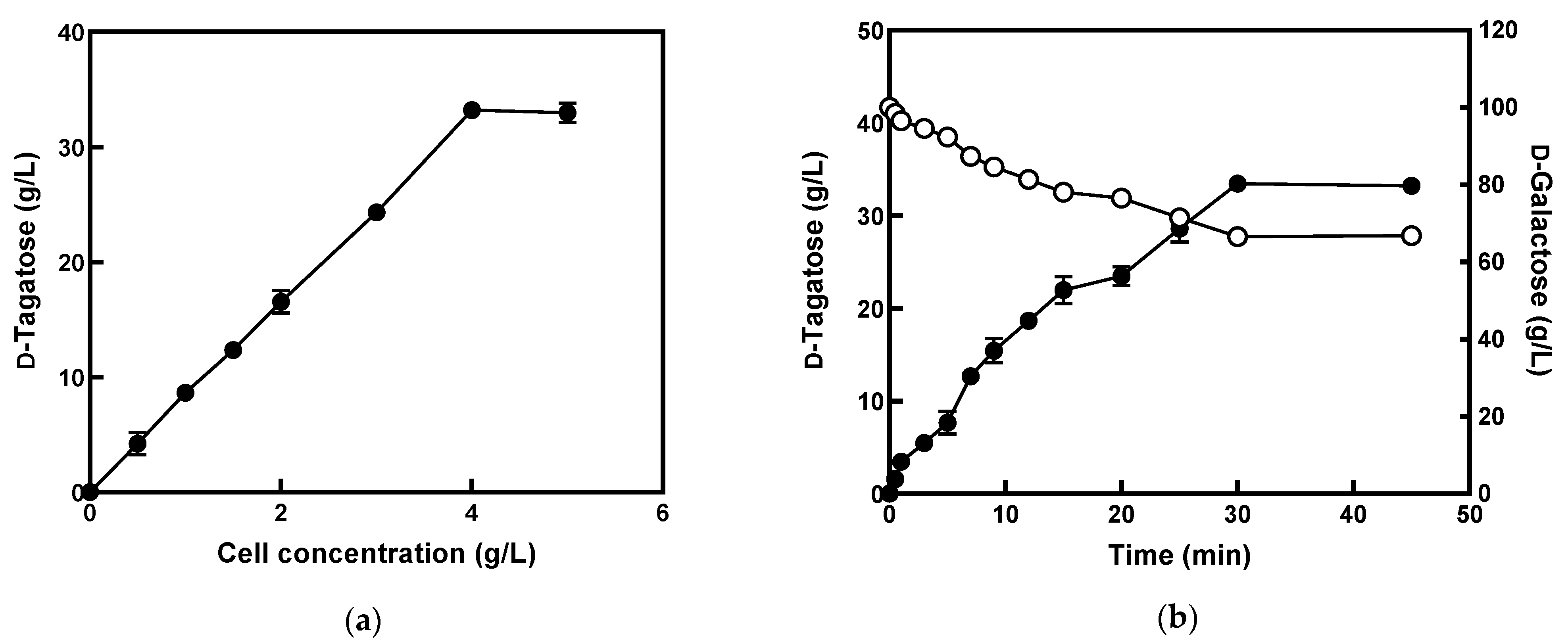
3.5. Production of d-Tagatose from d-Galactose Using E. coli Cells Expressing l-Arabinose Isomerase from K. pneumoniae
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Mendoza, M.R.; Olano, A.; Villamiel, M. Chemical indicators of heat treatment in fortified and special milks. J. Agric. Food Chem. 2005, 53, 2995–2999. [Google Scholar] [CrossRef]
- Oh, D.K. Tagatose: Properties, applications, and biotechnological processes. Appl. Microbiol. Biotechnol. 2007, 76, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Levin, G.V.; Zehner, L.R.; Saunders, J.P.; Beadle, J.R. Sugar substitutes: Their energy values, bulk characteristics, and potential health benefits. Am. J. Clin. Nutr. 1995, 62, 1161s–1168s. [Google Scholar] [CrossRef]
- Levin, G.V. Tagatose, the new GRAS sweetener and health product. J. Med. Food 2002, 5, 23–36. [Google Scholar] [CrossRef]
- Espinosa, I.; Fogelfeld, L. Tagatose: From a sweetener to a new diabetic medication? Expert Opin. Investig. Drugs 2010, 19, 285–294. [Google Scholar] [CrossRef]
- Kim, P. Current studies on biological tagatose production using l-arabinose isomerase: A review and future perspective. Appl. Microbiol. Biotechnol. 2004, 65, 243–249. [Google Scholar] [CrossRef]
- Lu, Y.; Levin, G.V.; Donner, T.W. Tagatose, a new antidiabetic and obesity control drug. Diabetes Obes. Metab. 2008, 10, 109–134. [Google Scholar] [CrossRef] [PubMed]
- Beadle, J.R.; Saunders, J.P.; Wajda, T.J. Process for Manufacturing Tagatose. U.S. Patent US5078796A, 7 January 1992. [Google Scholar]
- Xu, Z.; Li, S.; Feng, X.; Liang, J.; Xu, H. l-Arabinose isomerase and its use for biotechnological production of rare sugars. Appl. Microbiol. Biotechnol. 2014, 98, 8869–8878. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Mu, W.; Zhang, T.; Jiang, B. An l-arabinose isomerase from Acidothermus cellulolytics ATCC 43068: Cloning, expression, purification, and characterization. Appl. Microbiol. Biotechnol. 2010, 86, 1089–1097. [Google Scholar] [CrossRef]
- Lee, S.J.; Lee, D.W.; Choe, E.A.; Hong, Y.H.; Kim, S.B.; Kim, B.C.; Pyun, Y.R. Characterization of a thermoacidophilic l-arabinose isomerase from Alicyclobacillus acidocaldarius: Role of Lys-269 in pH optimum. Appl. Environ. Microbiol. 2005, 71, 7888–7896. [Google Scholar] [CrossRef] [Green Version]
- Wanarska, M.; Kur, J. A method for the production of d-tagatose using a recombinant Pichia pastoris strain secreting β-D-galactosidase from Arthrobacter chlorophenolicus and a recombinant l-arabinose isomerase from Arthrobacter sp. 22c. Microb. Cell Fact. 2012, 11, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhu, Y.; Liu, A.; Sun, Y. Identification and characterization of a novel l-arabinose isomerase from Anoxybacillus flavithermus useful in d-tagatose production. Extrem. Life Under Extrem. Cond. 2011, 15, 441–450. [Google Scholar] [CrossRef]
- Zhou, X.; Wu, J.C. Heterologous expression and characterization of Bacillus coagulans l-arabinose isomerase. World J. Microbiol. Biotechnol. 2012, 28, 2205–2212. [Google Scholar] [CrossRef]
- Lee, D.W.; Choe, E.A.; Kim, S.B.; Eom, S.H.; Hong, Y.H.; Lee, S.J.; Lee, H.S.; Lee, D.Y.; Pyun, Y.R. Distinct metal dependence for catalytic and structural functions in the l-arabinose isomerases from the mesophilic Bacillus halodurans and the thermophilic Geobacillus stearothermophilus. Arch. Biochem. Biophys. 2005, 434, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, P.; Tiwari, M.K.; Jeya, M.; Gunasekaran, P.; Kim, I.W.; Lee, J.K. Cloning and characterization of a novel l-arabinose isomerase from Bacillus licheniformis. Appl. Microbiol. Biotechnol. 2008, 81, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Rhimi, M.; Bejar, S. Cloning, purification and biochemical characterization of metallic-ions independent and thermoactive L-arabinose isomerase from the Bacillus stearothermophilus US100 strain. Biochim. Biophys. Acta 2006, 1760, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Prabhu, P.; Jeya, M.; Tiwari, M.K.; Moon, H.J.; Singh, R.K.; Lee, J.K. Characterization of an l-arabinose isomerase from Bacillus subtilis. Appl. Microbiol. Biotechnol. 2010, 85, 1839–1847. [Google Scholar] [CrossRef]
- Seo, M.J. Characterization of an l-arabinose isomerase from Bacillus thermoglucosidasius for d-tagatose production. Biosci. Biotechnol. Biochem. 2013, 77, 385–388. [Google Scholar] [CrossRef]
- Salonen, N.; Nyyssölä, A.; Salonen, K.; Turunen, O. Bifidobacterium longum l-arabinose isomerase--overexpression in Lactococcus lactis, purification, and characterization. Appl. Biochem. Biotechnol. 2012, 168, 392–405. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Kim, P.; Oh, D.K. Properties of l-arabinose isomerase from Escherichia coli as biocatalyst for tagatose production. World J. Microbiol. Biotechnol. 2003, 19, 47–51. [Google Scholar] [CrossRef]
- Cao, T.P.; Choi, J.M.; Lee, S.J.; Lee, Y.J.; Lee, S.K.; Jun, Y.; Lee, D.W.; Lee, S.H. Crystallization and preliminary X-ray crystallographic analysis of l-arabinose isomerase from thermophilic Geobacillus kaustophilus. Acta Crystallogr. F-Struct. Biol. Commun. 2014, 70, 108–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Kim, J.H.; Oh, H.J.; Oh, D.K. Characterization of a mutated Geobacillus stearothermophilus l-arabinose isomerase that increases the production rate of d-tagatose. J. Appl. Microbiol. 2006, 101, 213–221. [Google Scholar] [CrossRef]
- Kim, H.J.; Oh, D.K. Purification and characterization of an l-arabinose isomerase from an isolated strain of Geobacillus thermodenitrificans producing d-tagatose. J. Biotechnol. 2005, 120, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Qing, Y.; Li, S.; Feng, X.; Xu, H.; Ouyang, P. A novel l-arabinose isomerase from Lactobacillus fermentum CGMCC2921 for d-tagatose production: Gene cloning, purification and characterization. J. Mol. Catal. B-Enzym. 2011, 70, 1–7. [Google Scholar] [CrossRef]
- Chouayekh, H.; Bejar, W.; Rhimi, M.; Jelleli, K.; Mseddi, M.; Bejar, S. Characterization of an l-arabinose isomerase from the Lactobacillus plantarum NC8 strain showing pronounced stability at acidic pH. FEMS Microbiol. Lett. 2007, 277, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Rhimi, M.; Ilhammami, R.; Bajic, G.; Boudebbouze, S.; Maguin, E.; Haser, R.; Aghajari, N. The acid tolerant l-arabinose isomerase from the food grade Lactobacillus sakei 23K is an attractive d-tagatose producer. Bioresour. Technol. 2010, 101, 9171–9177. [Google Scholar] [CrossRef]
- Men, Y.; Zhu, Y.; Zhang, L.; Kang, Z.; Izumori, K.; Sun, Y.; Ma, Y. Enzymatic conversion of d-galactose to d-tagatose: Cloning, overexpression and characterization of l-arabinose isomerase from Pediococcus pentosaceus PC-5. Microbiol. Res. 2014, 169, 171–178. [Google Scholar] [CrossRef]
- Rhimi, M.; Bajic, G.; Ilhammami, R.; Boudebbouze, S.; Maguin, E.; Haser, R.; Aghajari, N. The acid-tolerant l-arabinose isomerase from the mesophilic Shewanella sp. ANA-3 is highly active at low temperatures. Microb. Cell Fact. 2011, 10, 96. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, F.; Hansen, O.C.; Stougaard, P. Enzymatic conversion of d-galactose to d-tagatose: Heterologous expression and characterisation of a thermostable l-arabinose isomerase from Thermoanaerobacter mathranii. Appl. Microbiol. Biotechnol. 2004, 64, 816–822. [Google Scholar] [CrossRef]
- Lin, C.J.; Tseng, W.C.; Lin, T.H.; Liu, S.M.; Tzou, W.S.; Fang, T.Y. Characterization of a thermophilic l-rhamnose isomerase from Thermoanaerobacterium saccharolyticum NTOU1. J. Agric. Food Chem. 2010, 58, 10431–10436. [Google Scholar] [CrossRef]
- Lee, D.W.; Jang, H.J.; Choe, E.A.; Kim, B.C.; Lee, S.J.; Kim, S.B.; Hong, Y.H.; Pyun, Y.R. Characterization of a thermostable l-arabinose (d-galactose) isomerase from the hyperthermophilic eubacterium Thermotoga maritima. Appl. Environ. Microbiol. 2004, 70, 1397–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.C.; Lee, Y.H.; Lee, H.S.; Lee, D.W.; Choe, E.A.; Pyun, Y.R. Cloning, expression and characterization of l-arabinose isomerase from Thermotoga neapolitana: Bioconversion of d-galactose to d-tagatose using the enzyme. FEMS Microbiol. Lett. 2002, 212, 121–126. [Google Scholar] [CrossRef]
- Kim, J.W.; Kim, Y.W.; Roh, H.J.; Kim, H.Y.; Cha, J.H.; Park, K.H.; Park, C.S. Production of tagatose by a recombinant thermostable l-arabinose isomerase from Thermus sp. IM6501. Biotechnol. Lett. 2003, 25, 963–967. [Google Scholar] [CrossRef] [PubMed]
- Mordukhova, E.A.; Lee, H.S.; Pan, J.G. Improved thermostability and acetic acid tolerance of Escherichia coli via directed evolution of homoserine o-succinyltransferase. Appl. Environ. Microbiol. 2008, 74, 7660–7668. [Google Scholar] [CrossRef] [Green Version]
- Gur, E.; Biran, D.; Gazit, E.; Ron, E.Z. In vivo aggregation of a single enzyme limits growth of Escherichia coli at elevated temperatures. Mol. Microbiol. 2002, 46, 1391–1397. [Google Scholar] [CrossRef]
- van de Vossenberg, J.L.; Ubbink-Kok, T.; Elferink, M.G.; Driessen, A.J.; Konings, W.N. Ion permeability of the cytoplasmic membrane limits the maximum growth temperature of bacteria and archaea. Mol. Microbiol. 1995, 18, 925–932. [Google Scholar] [CrossRef] [Green Version]
- Ron, E.Z.; Davis, B.D. Growth rate of Escherichia coli at elevated temperatures: Limitation by methionine. J. Bacteriol. 1971, 107, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Manjasetty, B.A.; Chance, M.R. Crystal structure of Escherichia coli l-arabinose isomerase (ECAI), the putative target of biological tagatose production. J. Mol. Biol. 2006, 360, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Lee, S.J.; Kim, S.B.; Lee, S.J.; Lee, S.H.; Lee, D.W. Structural insights into conserved l-arabinose metabolic enzymes reveal the substrate binding site of a thermophilic l-arabinose isomerase. FEBS Lett. 2014, 588, 1064–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Li, S.; Liang, J.; Feng, X.; Xu, H. Protein purification, crystallization and preliminary X-ray diffraction analysis of l-arabinose isomerase from Lactobacillus fermentum CGMCC2921. Acta Crystallogr. F-Struct. Biol. Commun. 2015, 71, 28–33. [Google Scholar] [CrossRef] [Green Version]
- Torres, P.R.; Manzo, R.M.; Rubiolo, A.C.; Batista-Viera, F.D.; Mammarella, E.J. Purification of an l-arabinose isomerase from Enterococcus faecium DBFIQ E36 employing a biospecific affinity strategy. J. Mol. Catal. B-Enzym. 2014, 102, 99–105. [Google Scholar] [CrossRef]
- Hong, Y.H.; Lee, D.W.; Pyun, Y.R.; Lee, S.H. Creation of metal-independent hyperthermophilic l-arabinose isomerase by homologous recombination. J. Agric. Food Chem. 2011, 59, 12939–12947. [Google Scholar] [CrossRef]
- Liang, M.; Chen, M.; Liu, X.; Zhai, Y.; Liu, X.W.; Zhang, H.; Xiao, M.; Wang, P. Bioconversion of d-galactose to d-tagatose: Continuous packed bed reaction with an immobilized thermostable l-arabinose isomerase and efficient purification by selective microbial degradation. Appl. Microbiol. Biotechnol. 2012, 93, 1469–1474. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.S.; Kim, H.J.; Oh, D.K. Tagatose production by immobilized recombinant Escherichia coli cells containing Geobacillus stearothermophilus l-arabinose isomerase mutant in a packed-bed bioreactor. Biotechnol. Prog. 2005, 21, 1335–1340. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.H.; Lee, D.W.; Lee, S.J.; Choe, E.A.; Kim, S.B.; Lee, Y.H.; Cheigh, C.I.; Pyun, Y.R. Production of d-tagatose at high temperatures using immobilized Escherichia coli cells expressing l-arabinose isomerase from Thermotoga neapolitana. Biotechnol. Lett. 2007, 29, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zabed, H.M.; Yun, J.; Yuan, J.; Zhang, Y.; Wang, Y.; Qi, X. Two-stage biosynthesis of d-tagatose from milk whey powder by an engineered Escherichia coli strain expressing l-arabinose isomerase from Lactobacillus plantarum. Bioresour. Technol. 2020, 305, 123010. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Li, M.; Jiang, B.; Zhang, T.; Chen, J. Whole-cell biosynthesis of d-tagatose from maltodextrin by engineered Escherichia coli with multi-enzyme co-expression system. Enzyme Microb. Technol. 2021, 145, 109747. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metal Ion | Relative Activity (%) |
|---|---|
| Ba2+ | 82 |
| Ca2+ | 101 |
| Co2+ | 105 |
| Cu2+ | 56 |
| Fe2+ | 94 |
| Mg2+ | 115 |
| Mn2+ | 128 |
| EDTA | 6 |
| Negative control | 100 |
| Substrate | Product | Relative Activity (%) | |
|---|---|---|---|
| Pentose | l-Arabinose | l-Ribulose | 315.6 |
| d-Xylose | d-Xylulose | 10.2 | |
| l-Ribulose | l-Arabinose | 30.2 | |
| d-Xylulose | d-Xylose | 27.6 | |
| Hexose | d-Galactose | d-Tagatose | 100.0 |
| d-Glucose | d-Fructose | 2.4 | |
| d-Tagatose | d-Galactose | 4.3 | |
| d-Fructose | d-Glucose | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, K.-C.; Seo, M.-J.; Kim, S.J.; Kim, Y.-S.; Park, C.-S. Characterization of l-Arabinose Isomerase from Klebsiella pneumoniae and Its Application in the Production of d-Tagatose from d-Galactose. Appl. Sci. 2022, 12, 4696. https://doi.org/10.3390/app12094696
Shin K-C, Seo M-J, Kim SJ, Kim Y-S, Park C-S. Characterization of l-Arabinose Isomerase from Klebsiella pneumoniae and Its Application in the Production of d-Tagatose from d-Galactose. Applied Sciences. 2022; 12(9):4696. https://doi.org/10.3390/app12094696
Chicago/Turabian StyleShin, Kyung-Chul, Min-Ju Seo, Sang Jin Kim, Yeong-Su Kim, and Chang-Su Park. 2022. "Characterization of l-Arabinose Isomerase from Klebsiella pneumoniae and Its Application in the Production of d-Tagatose from d-Galactose" Applied Sciences 12, no. 9: 4696. https://doi.org/10.3390/app12094696
APA StyleShin, K.-C., Seo, M.-J., Kim, S. J., Kim, Y.-S., & Park, C.-S. (2022). Characterization of l-Arabinose Isomerase from Klebsiella pneumoniae and Its Application in the Production of d-Tagatose from d-Galactose. Applied Sciences, 12(9), 4696. https://doi.org/10.3390/app12094696