
Gluconacetobacter diazotrophicus Inoculation of Two Lettuce Cultivars Affects Leaf and Root Growth under Hydroponic Conditions

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Randomization
2.2. Plant and Bacterial Material
2.3. Inoculant Preparation
2.4. Seed Inoculation, Planting, and Growing Conditions
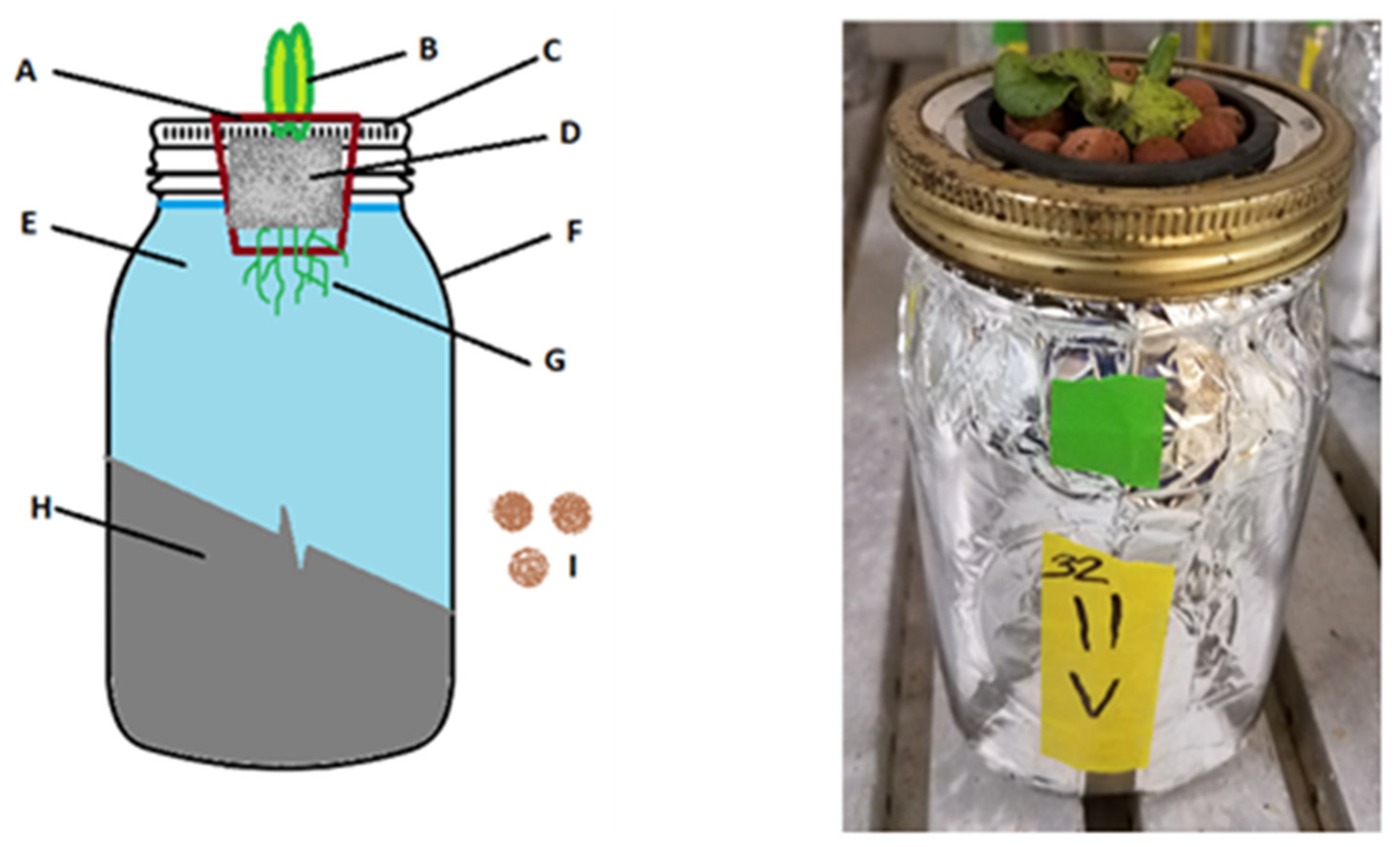
- A.
- 2” circular net cup used to suspend the plant above the nutrient solution through a hole cut in the center of the Mason jar lid.
- B.
- Growing lettuce shoots.
- C.
- Wide-mouth quart jar metal lid and collar. Pre-drilled 7/16” hole in lid center allows net cup to rest inside on the top lip.
- D.
- A036/40 rockwool used as seedling and plant support media. Inert, pH neutral, high wicking growth media with no nutrient content. Bottom 1/4” of rockwool extends into the nutrient solution to allow wicking.
- E.
- Nutrient solution, 800 mL in a 946 mL (1 qt) jar.
- F.
- 946 mL (1 qt) wide-mouth Mason jar.
- G.
- Roots growing into the media solution.
- H.
- Aluminum foil completely wrapping the sides and bottom of jar to prevent exposure to light.
- I.
- Hydroton clay pebbles placed atop the rockwool to limit evaporation and block light.
2.5. Harvesting and Analysis
2.6. Statistical Analysis
3. Results
3.1. Black Seeded Simpson (BSS) Trials
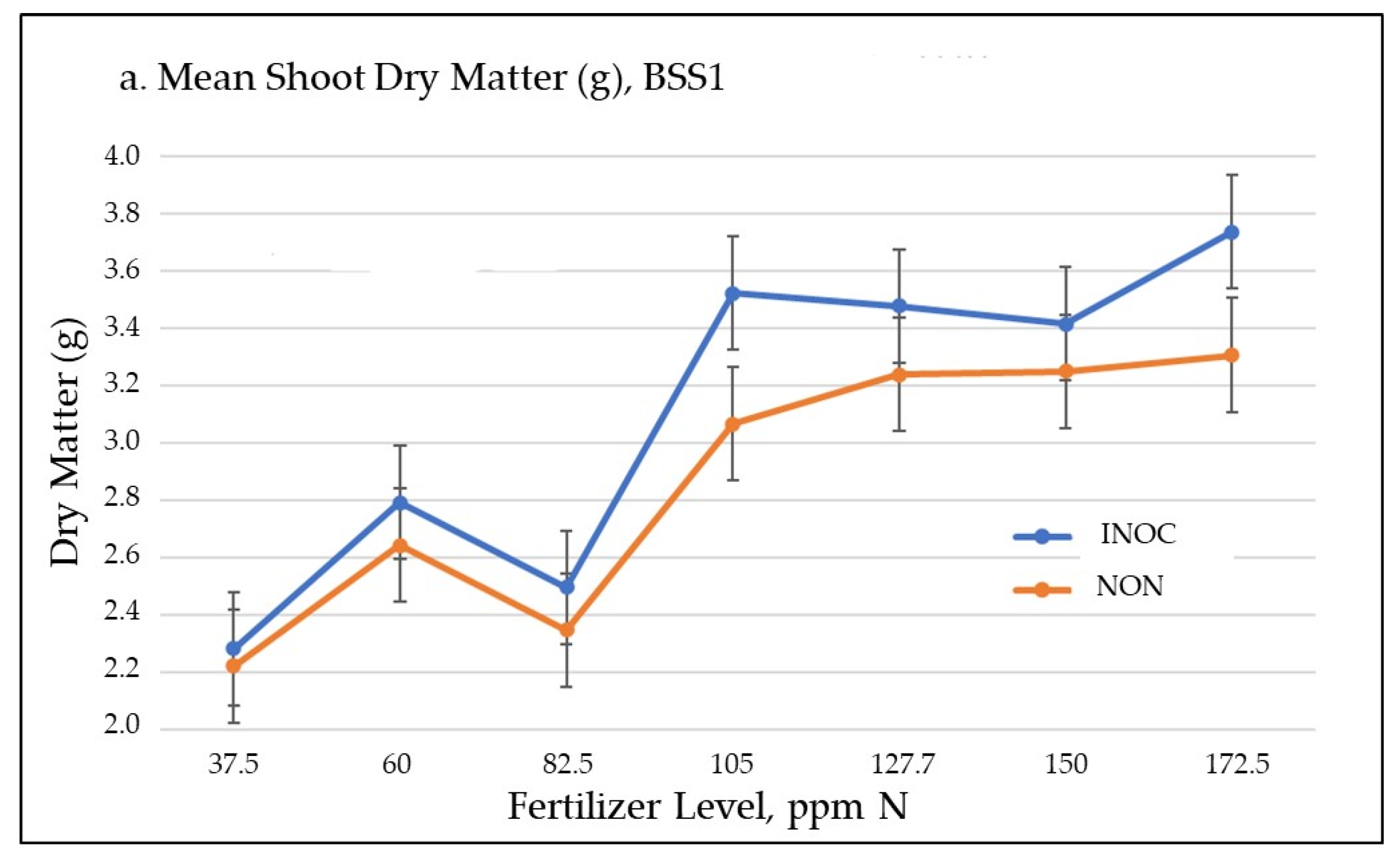
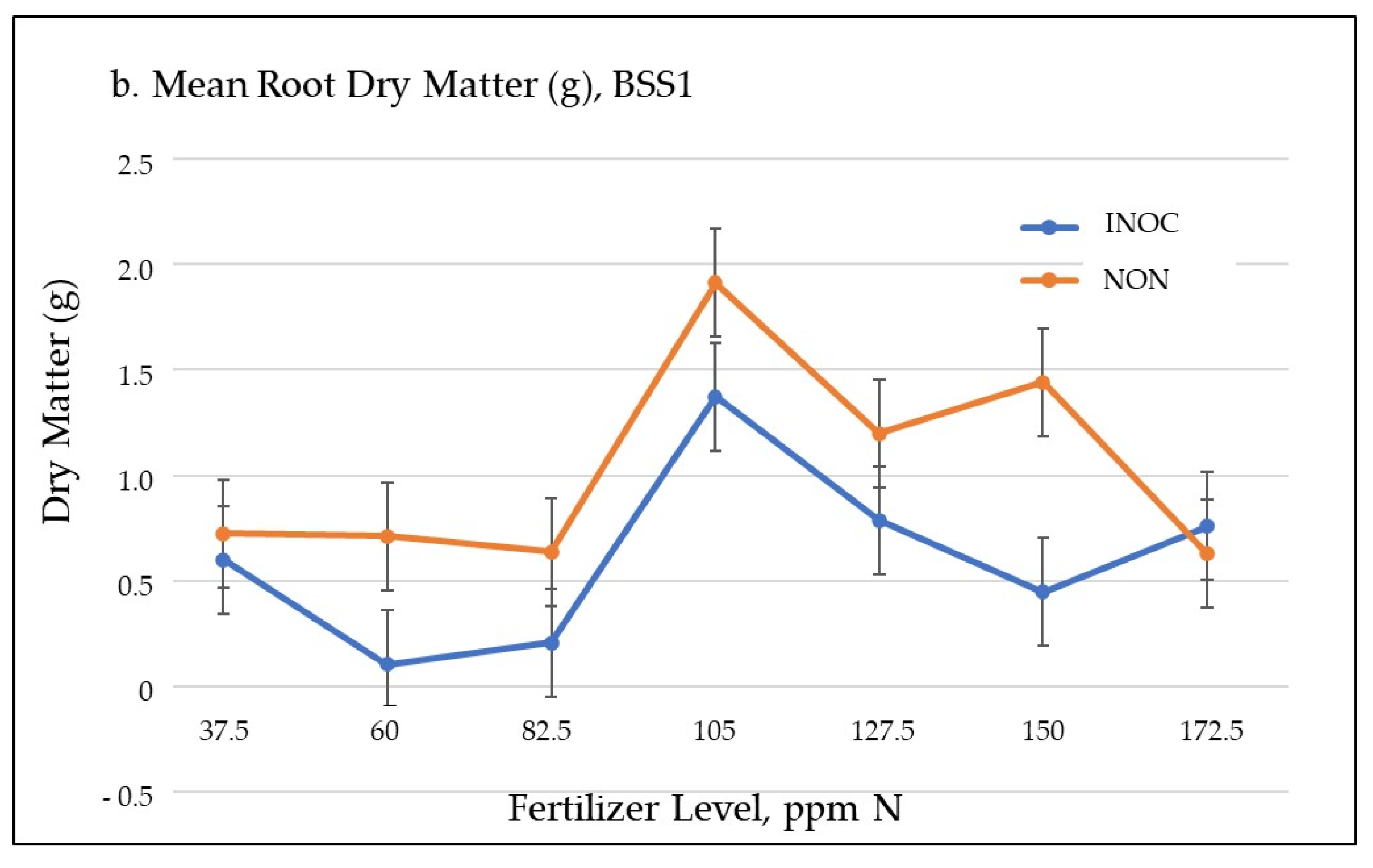
3.1.1. Biomass Accumulation
3.1.2. Carbon Nitrogen Ratios and Nitrogen Accumulation
3.2. Bibb/Limestone (BIB) Trials
3.2.1. Biomass Accumulation
3.2.2. Carbon Nitrogen Ratios and Nitrogen Accumulation
4. Discussion
4.1. Black Seeded Simpson (BSS) Trials
4.2. Bibb/Limestone (BIB) Trials
4.3. Cross Experiment Observations and Implications
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- USDA National Agricultural Statistics Service. 2020; Vegetables 2019 Summary. Available online: https://www.nass.usda.gov/Publications/Todays_Reports/reports/vegean20.pdf (accessed on 22 September 2021).
- Agricultural Marketing Resource Center. 2018. Lettuce. Available online: https://www.agmrc.org/commodities-products/vegetables/lettuce (accessed on 19 April 2021).
- Smith, R.; Cahn, M.; Daugovish, O.; Koike, S.; Natwick, E.; Smith, H.; Subbarao, K.; Takele, E.; Turini, T. Leaf Lettuce production in California. UC Davis Vegetable Research and Information Center. Publication 7216. 2011. Available online: https://anrcatalog.ucanr.edu/pdf/7216.pdf (accessed on 13 April 2021).
- Cometti, N.N.; Matias, G.C.S.; Zonta, E.; Mary, W.; Fernandes, M.S. Efeito da concentração da solução nutritiva no crescimento da alface em cultivo hidropônico-sistema NFT. Hortic. Bras. 2008, 26, 262–267. [Google Scholar] [CrossRef][Green Version]
- Barbosa, G.L.; Gadelha, F.D.A.; Kublik, N.; Proctor, A.; Reichelm, L.; Weissinger, E.; Wohlleb, G.M.; Halden, R.U. Comparison of Land, Water, and Energy Requirements of Lettuce Grown Using Hydroponic vs. Conventional Agricultural Methods. Int. J. Environ. Res. Public Health 2015, 12, 6879–6891. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, P.; Schlaeppi, K.; Sessitsch, A. miCROPe 2019—emerging research priorities towards microbe-assisted crop production. FEMS Microbiol. Ecol. 2020, 96, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Cavalcante, V.A.; Dobereiner, J. A new acid-tolerant nitrogen-fixing bacterium associated with sugarcane. Plant Soil 1988, 108, 23–31. [Google Scholar] [CrossRef]
- Muthukumarasamy, R.; Revathi, G.; Seshadri, S.; Lakshminarasimhan, C. Gluconacetobacter diazotrophicus (syn. Acetobacter diazotrophicus), a promising diazotrophic endophyte in tropics. Curr. Sci. 2002, 83, 137–145. [Google Scholar]
- Stephan, P.M.; Oliveira, M.; Teixeira, K.R.S.; Martinez-Drets, G.; Döbereiner, J. Physiology and dinitrogen fixation ofAcetobacter diazotrophicus. FEMS Microbiol. Lett. 1991, 77, 67–72. [Google Scholar] [CrossRef][Green Version]
- Boddey, R.M.; Polidoro, J.C.; Resende, A.S.; Alves, B.J.R.; Urquiaga, S. Use of the15N natural abundance technique for the quantification of the contribution of N2 fixation to sugar cane and other grasses. Funct. Plant Biol. 2001, 28, 889–895. [Google Scholar] [CrossRef]
- Jimenez-Salgado, T.; Fuentes-Ramirez, L.E.; Tapia-Hernandez, A.; Mascarua-Esparza, M.A.; Martinez-Romero, E.; Caballero-Mellado, J. Coffea arabica L., a new host plant for Acetobacter diazotrophicus, and isolation of other nitrogen-fixing acetobacteria. Appl. Environ. Microbiol. 1997, 63, 3676–3683. [Google Scholar] [CrossRef]
- Loganathan, P.; Sunita, R.; Parida, A.K.; Nair, S. Isolation and characterization of two genetically distant groups of Acetobacter diazotrophicus from a new host plant Eleusine coracana L. J. Appl. Microbiol. 1999, 87, 167–172. [Google Scholar] [CrossRef]
- Weber, O.; Baldani, V.L.D.; Teixeira, K.R.S.; Kirchhof, G.; Baldani, J.I.; Dobereiner, J. Isolation and characterization of diazotrophic bacteria from banana and pineapple plants. Plant Soil 1999, 210, 103–113. [Google Scholar] [CrossRef]
- Tapia-Hernández, A.; Bustillos-Cristales, M.; Jiménez-Salgado, T.; Caballero-Mellado, J.; Fuentes-Ramírez, L. Natural Endophytic Occurrence of Acetobacter diazotrophicus in Pineapple Plants. Microb. Ecol. 2000, 39, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Madhaiyan, M.; Saravanan, V.; Jovi, D.S.S.; Lee, H.; Thenmozhi, R.; Hari, K.; Sa, T. Occurrence of Gluconacetobacter diazotrophicus in tropical and subtropical plants of Western Ghats, India. Microbiol. Res. 2004, 159, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Muthukumarasamy, R.; Cleenwerck, I.; Revathi, G.; Vadivelu, M.; Janssens, D.; Hoste, B.; Gum, K.U.; Park, K.D.; Son, C.Y.; Sa, T.; et al. Natural association of Gluconacetobacter diazotrophicus and diazotrophic Acetobacter peroxydans with wetland rice. Syst. Appl. Microbiol. 2005, 28, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Pedraza, R.O. Recent advances in nitrogen-fixing acetic acid bacteria. Int. J. Food Microbiol. 2008, 125, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Riggs, P.J.; Chelius, M.K.; Iniguez, A.L.; Kaeppler, S.M.; Triplett, E.W. Enhanced maize productivity by inoculation with diazotrophic bacteria. Funct. Plant Biol. 2001, 28, 829–836. [Google Scholar] [CrossRef]
- Tian, G.; Pauls, P.; Dong, Z.; Reid, L.M.; Tian, L. Colonization of the nitrogen-fixing bacterium Gluconacetobacter dia-zotrophicus in a large number of Canadian corn plants. Can. J. Plant Sci. 2009, 89, 1009–1016. [Google Scholar] [CrossRef]
- Luna, M.F.; Apreaa, J.; Crespoa, J.M.; Boiardia, J.L. Colonization and yield promotion of tomato by Gluconacetobacter diazotrophicus. Appl. Soil Ecol. 2012, 61, 225–229. [Google Scholar] [CrossRef]
- Paula, M.A.; Reis, V.M. Interactions of Glomus clarum with Acetobacter diazotrophicus in infection of sweet potato (Ipomoea batatas), sugarcane (Saccharum spp.), and sweet sorghum (Sorghum vulgare). Biol. Fertil. Soils 1991, 11, 111–115. [Google Scholar] [CrossRef]
- Yoon, V.; Tian, G.; Vessey, J.K.; MacFie, S.M.; Dangi, O.P.; Kumer, A.K.; Tian, L. Colonization efficiency of different sorghum genotypes by Gluconacetobacter diazotrophicus. Plant Soil 2015, 398, 243–256. [Google Scholar] [CrossRef]
- Rocafell, Y.R.; Álvarez, B.D.; Badía, M.R.; García, M.O.; Daza, N.A.; Sánchez, J.R. Interaction of the bacteria Gluconacetobacter diazotrophicus and root vegetables. Cultiv. Trop. 2016, 37, 28–32. [Google Scholar] [CrossRef]
- Pellegrini, M.; Pagnani, G.; Rossi, M.; D’Egidio, S.; Gallo, M.; Forni, C. Daucus carota L. Seed Inoculation with a Consortium of Bacteria Improves Plant Growth, Soil Fertility Status and Microbial Community. Appl. Sci. 2021, 11, 3274. [Google Scholar] [CrossRef]
- de Oliveira, T.R.A.; Gravina, G.A.; da Cruz, D.P.; Silva, N.D.; de Oliveira, G.H.F.; de Sant’Anna, C.Q.; Magalhães, M.M.; Berbert-Molina, M.A.; Neto, F.A. The performance of bean pod lineage inoculated with Gluconacetobacter diazotrophicus PAL5. Sci. Hortic. 2019, 249, 65–70. [Google Scholar] [CrossRef]
- Lopes, E.A.P.; Da Silva, A.D.A.; Mergulhão, A.C.D.E.S.; Da Silva, E.V.N.; Santiago, A.D.; Figueiredo, M.D.V.B. Co-Inoculation of Growth Promoting Bacteria and Glomus Clarum in Micropropagated Cassava Plants. Rev. Caatinga 2019, 32, 152–166. [Google Scholar] [CrossRef]
- Fisher, K.; Newton, W.E. Nitrogenase proteins from Gluconacetobacter diazotrophicus, a sugarcane-colonizing bacterium. Biochim. et Biophys. Acta (BBA) Proteins Proteom. 2005, 1750, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Andrade, O.; Fuentes-Ramírez, L.E.; Morales-García, Y.E.; Molina-Romero, D.; Bustillos-Cristales, M.R.; Martínez-Contreras, R.D.; Muñoz-Rojas, J. Colonization of sugarcane by Acetobacter diazotrophicus is inhibited by high N-fertilization. FEMS Microbiol. Ecol. 1999, 29, 117–128. [Google Scholar] [CrossRef]
- Rodríguez-Andrade, O.; Fuentes-Ramírez, L.E.; Morales-García, Y.E.; Molina-Romero, D.; Bustillos-Cristales, M.R.; Martínez-Contreras, R.D.; Muñoz-Rojas, J. The decrease in the population of Gluconacetobacter diazotrophicus in sugarcane after nitrogen fertilization is related to plant physiology in split root experiments. Rev. Argent Microbiol. 2015, 47, 335–343. [Google Scholar] [CrossRef]
- Fuentes-Ramírez, L.E.; Jiménez-Salgado, T.; Abarca-Ocampo, I.R.; Caballero-Mellado, J. Acetobacter diazotrophicus, an indoleacetic acid producing bacterium isolated from sugarcane cultivars of Mexico. Plant Soil 1993, 154, 145–150. [Google Scholar] [CrossRef]
- Bastián, F.; Cohen, A.; Piccoli, P.; Luna, V.; Bottini, R.; Baraldi, R. Production of indole-3-acetic acid and gibberellins A1 and A3 by Acetobacter diazotrophicus and Herbaspirillum seropedicae in chemically-defined culture media. Plant Growth Regul. 1998, 24, 7–11. [Google Scholar] [CrossRef]
- Piñón, D.; Casas, M.; Blanch, M.; Fontaniella, B.; Blanco, Y.; Vicente, C.; Solas, M.-T.; Legaz, M.-E. Gluconacetobacter diazotrophicus, a sugar cane endosymbiont, produces a bacteriocin against Xanthomonas albilineans, a sugar cane pathogen. Res. Microbiol. 2002, 153, 345–351. [Google Scholar] [CrossRef]
- Arencibia, A.D.; Vinagre, F.; Estevez, Y.; Bernal, A.; Perez, J.; Cavalcanti, J.; Santana, I.; Hemerly, A.S. Gluconacetobacter diazotrophicus elicits a sugarcane defense response against a pathogenic bacteria Xanthomonas albilineans. Plant Signal. Behav. 2006, 1, 265–273. [Google Scholar] [CrossRef]
- Vargas, L.; Brígida, A.B.S.; Filho, J.P.M.; De Carvalho, T.G.; Rojas, C.; Vaneechoutte, D.; Van Bel, M.; Farrinelli, L.; Ferreira, P.C.G.; Vandepoele, K.; et al. Drought Tolerance Conferred to Sugarcane by Association with Gluconacetobacter diazotrophicus: A Transcriptomic View of Hormone Pathways. PLoS ONE 2014, 9, e114744. [Google Scholar] [CrossRef] [PubMed]
- Filgueiras, L.; Silva, R.; Almeida, I.; Vidal, M.; Baldani, J.I.; Meneses, C.H.S.G. Gluconacetobacter diazotrophicus mitigates drought stress in Oryza sativa L. Plant Soil 2019, 451, 57–73. [Google Scholar] [CrossRef]
- Saravanan, V.S.; Osborne, J.; Madhaiyan, M.; Mathew, L.; Chung, K.; Ahn, K.; Sa, T. Zinc Metal Solubilization by Gluconacetobacter diazotrophicus and Induction of Pleomorphic Cells. J. Microbiol. Biotechnol. 2007, 17, 1477–1482. [Google Scholar] [PubMed]
- Delaporte-Quintana, P.; Lovaisa, N.C.; Rapisarda, V.A.; Pedraza, R.O. The plant growth promoting bacteria Gluconacetobacter diazotrophicus and Azospirillum brasilense contribute to the iron nutrition of strawberry plants through siderophores production. Plant Growth Regul. 2020, 91, 185–199. [Google Scholar] [CrossRef]
- Kratky, B.A. A suspended pot, non-circulating hydroponic method. Proceedings of the South Pacific Soilless Culture Conference. Acta Hort. 2004, 648, 83–89. [Google Scholar] [CrossRef]
- Cocking, E.C.; Stone, P.J.; Davey, M.R. Intracellular colonization of roots of Arabidopsis and crop plants by Gluconacetobacter diazotrophicus. In Vitro Cell. Dev. Biol. Plant 2006, 42, 74–82. [Google Scholar] [CrossRef]
- Mattson, N.S.; Peters, C. A Recipe for Hydroponic Success. Inside Grower 16–19 January 2014. Available online: http://www.greenhouse.cornell.edu/crops/factsheets/hydroponic-recipes.pdf (accessed on 24 September 2021).
- Vysotskaya, L.B.; Veselov, S.Y.; Kudoyarova, G.R. Effect of Competition and Treatment with Inhibitor of Ethylene Perception on Growth and Hormone Content of Lettuce Plants. J. Plant Growth Regul. 2017, 36, 450–459. [Google Scholar] [CrossRef]
- Miceli, A.; Moncada, A.; Sabatino, L.; Vetrano, F. Effect of Gibberellic Acid on Growth, Yield, and Quality of Leaf Lettuce and Rocket Grown in a Floating System. Agronomy 2019, 9, 382. [Google Scholar] [CrossRef]
- Di Gioia, F.; Gonnella, M.; Buono, V.; Santamaria, P. Nitrogen use efficiency, yield and quality response of lettuce crop to nitrogen input. In Proceedings of the Conference on Nitrogen, Environment and Vegetables (NEV2013), Torino, Italy, 15—17 April 2013. [Google Scholar]
- Amirouche, M.; Smadhi, D.; Zella, L. Modeling of Nitrogen Use Efficiency in Lettuce Culture (Lactuca sativa): Isotopic Nitrogen (15 N) and AquaCrop. In Nitrogen in Agriculture—Physiological, Agricultural and Ecological Aspec; IntechOpen: London, UK, 2020; pp. 1–15. [Google Scholar] [CrossRef]
- Gang, S.; Sharma, S.; Saraf, M.; Buck, M.; Schumacher, J. Bacterial Indole-3-Acetic Acid Influences Soil Nitrogen Acquisition in Barley and Chickpea. Plants 2021, 10, 780. [Google Scholar] [CrossRef]
- Eskin, N.; Vessey, K.; Tian, L. Research Progress and Perspectives of Nitrogen Fixing Bacterium, Gluconacetobacter diazotrophicus, in Monocot Plants. Int. J. Agron. 2014, 2014, 1–13. [Google Scholar] [CrossRef]
- de Souza, A.L.S.R.; De Souza, S.A.; De Oliveira, M.V.V.; Ferraz, T.M.; Figueiredo, F.A.M.M.A.; Da Silva, N.D.; Rangel, P.L.; Panisset, C.R.S.; Olivares, F.L.; Campostrini, E.; et al. Endophytic colonization of Arabidopsis thaliana by Gluconacetobacter diazotrophicus and its effect on plant growth promotion, plant physiology, and activation of plant defense. Plant Soil 2015, 399, 257–270. [Google Scholar] [CrossRef]
- Di Gioia, F.; Gonnella, M.; Buono, V.; Ayala, O.; Santamaria, P. Agronomic, physiological and quality response of romaine and red oak-leaf lettuce to nitrogen input. Ital. J. Agron. 2017, 12, 47–58. [Google Scholar] [CrossRef]
- Sebring, R.L.; Schlossberg, M.; Regan, J.; Bryant, R.B. Establishment of Kentucky Bluegrass Inoculated with the Novel Nitrogen Fixing Bacterial Endophyte, Gluconacetobacter Diazotrophicus. Ph.D. Thesis, The Pennsylvania State Univiversity, University Park, State College, PA, USA, 2017. [Google Scholar]
- Day, J.A.; Diener, C.; Otwell, A.E.; Tams, K.E.; Bebout, B.; Detweiler, A.M.; Lee, M.D.; Scott, M.T.; Ta, W.; Ha, M.; et al. Lettuce (Lactuca sativa) productivity influenced by microbial inocula under nitrogen-limited conditions in aquaponics. PLoS ONE 2021, 16, e0247534. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fertilizer Treatment Provided Nitrogen Concentration (ppm) | |||||||
| Trial and Measure | 37.5 | 60 | 82.5 | 105 | 127.5 | 150 | 172.5 |
| BSS1 | |||||||
| Shoot Dry Matter (g) * | LSD: 0.235 s.e.: 0.1057 df = 28 p ≤ 0.0345 | ||||||
| INOC | 2.2843 | 2.7927 | 2.499 | 3.524 | 3.4793 | 3.416 | 3.7387 |
| NON | 2.224 | 2.645 | 2.3493 | 3.0683 | 3.241 | 3.2527 | 3.3087 |
| Difference | 0.0603 | 0.1477 | 0.1497 | 0.4557 | 0.2383 | 0.1633 | 0.43 |
| Root Dry Matter (g) ** | LSD: −0.4251 s.e.: 0.1366 df = 28 p ≤ 0.0042 | ||||||
| INOC | 0.602 | 0.1067 | 0.207 | 1.3737 | 0.7867 | 0.4473 | 0.7597 |
| NON | 0.7243 | 0.7117 | 0.639 | 1.913 | 1.1987 | 1.4433 | 0.629 |
| Difference | −0.122 | −0.605 | −0.432 | −0.539 | −0.412 | −0.996 | 0.1307 |
| Total Dry Matter (g) | n/s | ||||||
| INOC | 2.8863 | 2.8993 | 2.706 | 4.8977 | 4.266 | 3.8633 | 4.4983 |
| NON | 2.9483 | 3.3567 | 2.9883 | 4.9813 | 4.4397 | 4.696 | 3.9377 |
| Difference | −0.062 | −0.457 | −0.282 | −0.084 | −0.174 | −0.833 | 0.5606 |
| BSS2 | |||||||
| Shoot Dry Matter (g) | n/s | ||||||
| INOC | - | 4.7867 | 5.7825 | 5.705 | 6.3525 | 6.6475 | 6.2067 |
| NON | - | 4.47 | 6 | 5.6875 | 7.0975 | 5.715 | 6.7767 |
| Difference | - | 0.3167 | −0.218 | 0.0175 | −0.745 | 0.9325 | −0.57 |
| Root Dry Matter (g) | n/s | ||||||
| INOC | - | 1.78 | 2.0675 | 2.135 | 2.0425 | 2.2175 | 2.2333 |
| NON | - | 1.89 | 1.92 | 2.0525 | 2.285 | 1.885 | 2.13 |
| Difference | - | −0.11 | 0.1475 | 0.0825 | −0.243 | 0.3325 | 0.1033 |
| Total Dry Matter (g) | n/s | ||||||
| INOC | - | 6.5667 | 7.85 | 7.84 | 8.395 | 8.865 | 8.44 |
| NON | - | 6.36 | 7.92 | 7.74 | 9.3825 | 7.6 | 8.9067 |
| Difference | - | 0.2067 | −0.07 | 0.1 | −0.988 | 1.265 | −0.467 |
| Fertilizer Treatment Provided Nitrogen Concentration (ppm) | |||||||
| Trial and Measure | 37.5 | 60 | 82.5 | 105 | 127.5 | 150 | 172.5 |
| BSS1 | |||||||
| SDM Nitrogen % ** | LSD: −0.2328 s.e.: 0.06591 df = 28 p ≤ 0.0015 | ||||||
| INOC | 1.290 | 1.465 | 1.930 | 1.898 | 2.268 | 2.596 | 2.390 |
| NON | 1.337 | 1.828 | 2.326 | 2.110 | 2.276 | 2.648 | 2.940 |
| Difference | −0.047 | −0.363 | −0.396 | −0.212 | −0.008 | −0.052 | −0.550 |
| SDM C/N Ratio ** | LSD: 2.2234 s.e.: 0.7305 df = 28 p ≤ 0.0050 | ||||||
| INOC | 30.495 | 26.496 | 20.144 | 20.821 | 17.370 | 15.439 | 16.513 |
| NON | 29.260 | 21.143 | 16.466 | 18.783 | 17.419 | 15.060 | 13.583 |
| Difference | 1.236 | 5.353 | 3.678 | 2.038 | −0.049 | 0.379 | 2.930 |
| RDM Nitrogen % | n/s | ||||||
| INOC | 1.191 | 1.322 | 2.078 | 1.360 | 1.560 | 2.507 | 2.382 |
| NON | 1.339 | 1.475 | 1.589 | 1.475 | 1.656 | 2.193 | 2.404 |
| Difference | −0.148 | −0.153 | 0.489 | −0.116 | −0.096 | 0.314 | −0.022 |
| RDM C/N Ratio | n/s | ||||||
| INOC | 31.839 | 25.611 | 16.846 | 19.224 | 17.898 | 14.019 | 14.047 |
| NON | 27.854 | 22.803 | 18.133 | 18.065 | 17.514 | 14.886 | 13.412 |
| Difference | 3.985 | 2.808 | −1.287 | 1.159 | 0.384 | −0.867 | 0.635 |
| BSS2 | |||||||
| SDM Nitrogen % | n/s | ||||||
| INOC | - | 0.981 | 1.099 | 1.282 | 1.416 | 1.455 | 1.670 |
| NON | - | 0.989 | 1.051 | 1.303 | 1.283 | 1.722 | 1.619 |
| Difference | - | −0.008 | 0.048 | −0.021 | 0.133 | −0.267 | 0.051 |
| SDM C/N Ratio | n/s | ||||||
| INOC | - | 40.344 | 36.258 | 31.033 | 28.425 | 27.444 | 23.917 |
| NON | - | 39.789 | 37.960 | 30.498 | 30.977 | 23.460 | 24.860 |
| Difference | - | 0.555 | −1.701 | 0.535 | −2.552 | 3.983 | −0.943 |
| RDM Nitrogen % | n/s | ||||||
| INOC | - | 1.135 | 1.092 | 1.537 | 1.690 | 1.800 | 1.748 |
| NON | - | 1.099 | 1.320 | 1.380 | 1.326 | 1.985 | 1.737 |
| Difference | - | 0.036 | −0.228 | 0.158 | 0.364 | −0.185 | 0.011 |
| RDM C/N Ratio | n/s | ||||||
| INOC | - | 32.169 | 35.669 | 25.131 | 24.014 | 21.337 | 22.515 |
| NON | - | 36.039 | 29.351 | 28.021 | 28.078 | 21.438 | 22.466 |
| Difference | - | −3.871 | 6.318 | −2.891 | −4.064 | −0.101 | 0.049 |
| Fertilizer Treatment Provided Nitrogen Concentration (ppm) | |||||||
| Trial and Measure | 37.5 | 60 | 82.5 | 105 | 127.5 | 150 | 172.5 |
| BIB1 | |||||||
| Shoot Dry Matter (g) * | LSD: 0.5296 s.e.: 0.2387 df = 32 p ≤ 0.0338 | ||||||
| INOC | - | 4.280 | 6.048 | 5.535 | 5.378 | 5.825 | 5.377 |
| NON | - | 4.217 | 4.805 | 5.025 | 5.250 | 5.188 | 4.780 |
| Difference | - | 0.063 | 1.243 | 0.510 | 0.128 | 0.638 | 0.597 |
| Root Dry Matter (g) | n/s | ||||||
| INOC | - | 1.833 | 1.958 | 2.083 | 1.905 | 1.908 | 1.557 |
| NON | - | 1.637 | 1.805 | 2.000 | 1.863 | 1.680 | 1.563 |
| Difference | - | 0.197 | 0.153 | 0.083 | 0.043 | 0.228 | −0.007 |
| Total Dry Matter (g) * | LSD: 0.6655 s.e.: 0.3015 df = 37 p ≤ 0.0336 | ||||||
| INOC | - | 6.113 | 8.005 | 7.618 | 7.283 | 7.733 | 6.933 |
| NON | - | 5.853 | 6.610 | 7.025 | 7.113 | 6.868 | 6.343 |
| Difference | - | 0.260 | 1.395 | 0.592 | 0.170 | 0.865 | 0.590 |
| BIB2 | |||||||
| Shoot Dry Matter (g) * | LSD: 0.6526 s.e.: 0.2482 df = 34 p ≤ 0.0127 | ||||||
| INOC | - | 4.457 | 6.381 | 4.825 | 3.908 | 3.676 | 3.843 |
| NON | - | 3.937 | 4.588 | 3.672 | 3.526 | 3.594 | 3.949 |
| Difference | - | 0.519 | 1.793 | 1.153 | 0.382 | 0.082 | −0.106 |
| Root Dry Matter (g) ** | LSD: 0.2534 s.e.: 0.07679 df = 34 p ≤ 0.0023 | ||||||
| INOC | - | 1.930 | 2.047 | 1.862 | 1.246 | 1.144 | 1.416 |
| NON | - | 1.580 | 1.671 | 1.467 | 1.111 | 1.138 | 1.099 |
| Difference | - | 0.351 | 0.376 | 0.394 | 0.135 | 0.006 | 0.317 |
| Total Dry Matter (g)** | LSD: 0.9060 s.e.: 0.2942 df = 34 p ≤ 0.0041 | ||||||
| INOC | - | 6.387 | 8.428 | 6.687 | 5.154 | 4.820 | 5.258 |
| NON | - | 5.517 | 6.259 | 5.140 | 4.637 | 4.733 | 5.047 |
| Difference | - | 0.870 | 2.169 | 1.547 | 0.517 | 0.088 | 0.211 |
| Fertilizer Treatment Provided Nitrogen Concentration (ppm) | |||||||
| Trial and Measure | 37.5 | 60 | 82.5 | 105 | 127.5 | 150 | 172.5 |
| BIB1 | |||||||
| SDM Nitrogen % ** | n/s | ||||||
| INOC | - | 1.078 | 1.167 | 1.525 | 1.813 | 1.853 | 2.275 |
| NON | - | 1.095 | 1.284 | 1.612 | 1.729 | 2.059 | 2.122 |
| Difference | - | −0.017 | −0.118 | −0.087 | 0.085 | −0.207 | 0.153 |
| SDM C/N Ratio ** | n/s | ||||||
| INOC | - | 36.990 | 34.444 | 26.534 | 22.670 | 21.712 | 17.938 |
| NON | - | 36.442 | 31.337 | 25.226 | 23.529 | 19.697 | 19.051 |
| Difference | - | 0.548 | 3.107 | 1.308 | −0.860 | 2.015 | −1.114 |
| RDM Nitrogen % | n/s | ||||||
| INOC | - | 0.929 | 1.016 | 1.396 | 1.661 | 1.712 | 2.293 |
| NON | - | 0.961 | 1.119 | 1.265 | 1.583 | 1.693 | 2.090 |
| Difference | - | −0.031 | −0.103 | 0.131 | 0.079 | 0.018 | 0.204 |
| RDM C/N Ratio | n/s | ||||||
| INOC | - | 38.137 | 37.768 | 29.106 | 23.449 | 22.393 | 17.555 |
| NON | - | 36.593 | 30.946 | 29.356 | 23.147 | 17.578 | 17.169 |
| Difference | - | 1.544 | 6.822 | −0.250 | 0.303 | 4.815 | 0.385 |
| BIB2 | |||||||
| SDM Nitrogen % * | LSD: −0.2767 s.e.: 0.1311 df = 29 p ≤ 0.0436 | ||||||
| INOC | - | 0.821 | 0.525 | 1.347 | 1.944 | 2.578 | 2.711 |
| NON | - | 1.166 | 1.018 | 1.794 | 2.266 | 2.517 | 2.598 |
| Difference | - | −0.345 | −0.493 | −0.447 | −0.321 | 0.061 | 0.113 |
| SDM C/N Ratio * | LSD: 4.3860 s.e.: 1.8028 df = 29 p ≤ 0.0214 | ||||||
| INOC | - | 46.711 | 54.038 | 34.718 | 21.985 | 18.160 | 15.702 |
| NON | - | 45.598 | 42.960 | 24.143 | 20.420 | 16.329 | 15.546 |
| Difference | - | 1.113 | 11.078 | 10.575 | 1.565 | 1.830 | 0.155 |
| RDM Nitrogen % | n/s | ||||||
| INOC | - | 0.976 | 1.086 | 1.711 | 3.172 | 3.247 | 2.854 |
| NON | - | 1.602 | 1.470 | 1.926 | 2.681 | 2.901 | 2.901 |
| Difference | - | −0.627 | −0.384 | −0.215 | 0.491 | 0.346 | −0.047 |
| RDM C/N Ratio | n/s | ||||||
| INOC | - | 37.729 | 30.613 | 25.942 | 14.957 | 16.632 | 16.155 |
| NON | - | 29.565 | 28.208 | 22.692 | 18.673 | 15.365 | 16.032 |
| Difference | - | 8.164 | 2.405 | 3.250 | −3.717 | 1.267 | 0.123 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sebring, R.L.; Duiker, S.W.; Berghage, R.D.; Regan, J.M.; Lambert, J.D.; Bryant, R.B. Gluconacetobacter diazotrophicus Inoculation of Two Lettuce Cultivars Affects Leaf and Root Growth under Hydroponic Conditions. Appl. Sci. 2022, 12, 1585. https://doi.org/10.3390/app12031585
Sebring RL, Duiker SW, Berghage RD, Regan JM, Lambert JD, Bryant RB. Gluconacetobacter diazotrophicus Inoculation of Two Lettuce Cultivars Affects Leaf and Root Growth under Hydroponic Conditions. Applied Sciences. 2022; 12(3):1585. https://doi.org/10.3390/app12031585
Chicago/Turabian StyleSebring, Ryan L., Sjoerd W. Duiker, Robert D. Berghage, John M. Regan, Joshua D. Lambert, and Ray B. Bryant. 2022. "Gluconacetobacter diazotrophicus Inoculation of Two Lettuce Cultivars Affects Leaf and Root Growth under Hydroponic Conditions" Applied Sciences 12, no. 3: 1585. https://doi.org/10.3390/app12031585
APA StyleSebring, R. L., Duiker, S. W., Berghage, R. D., Regan, J. M., Lambert, J. D., & Bryant, R. B. (2022). Gluconacetobacter diazotrophicus Inoculation of Two Lettuce Cultivars Affects Leaf and Root Growth under Hydroponic Conditions. Applied Sciences, 12(3), 1585. https://doi.org/10.3390/app12031585