
The Influence of Silver Nanoparticle Form on the Toxicity in Freshwater Mussels



Abstract
:1. Introduction
2. Methods
2.1. Mussel Exposure Experiments
2.2. Resistance to Air Emersion
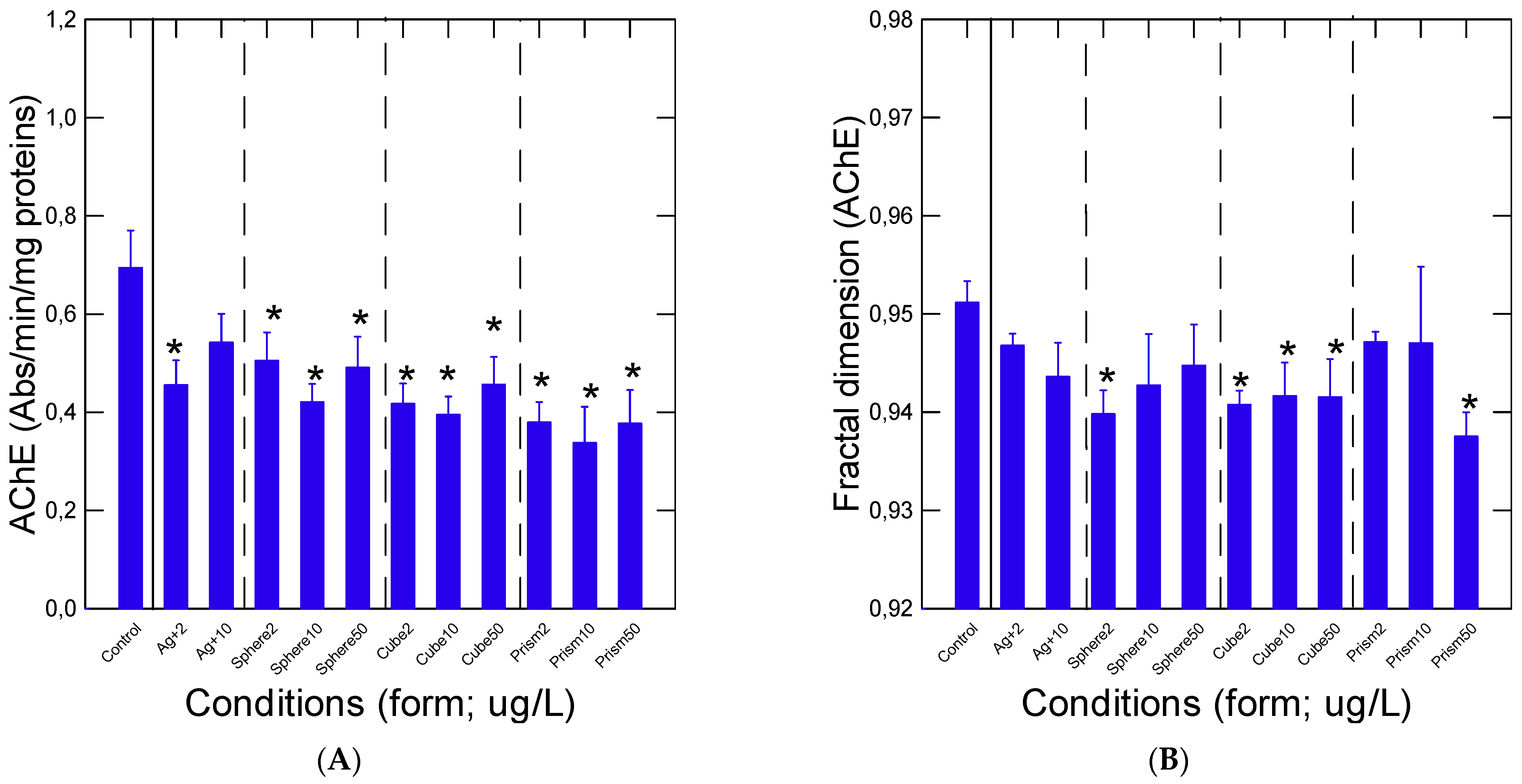
2.3. Acetylcholinesterase Activity
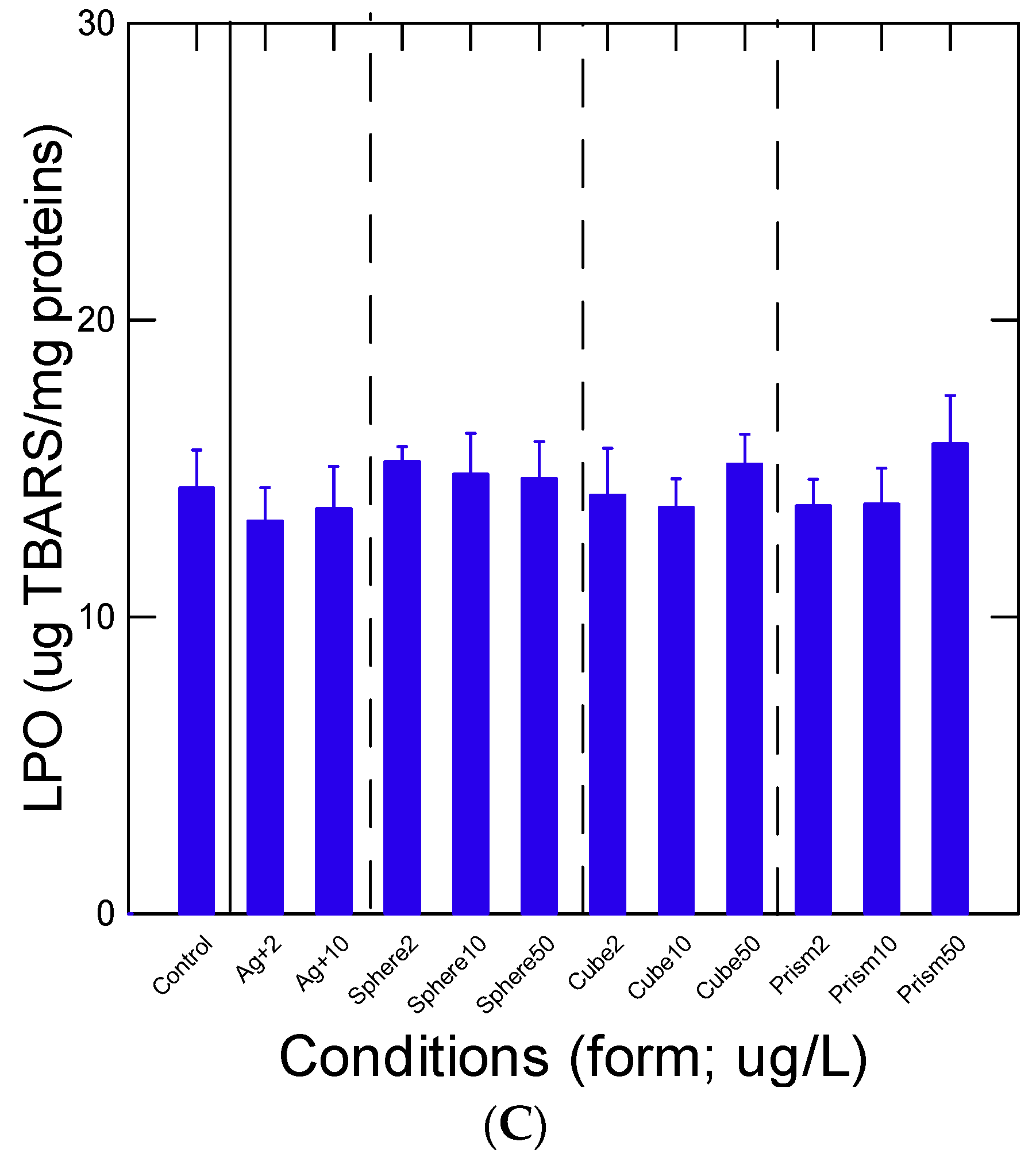
2.4. Oxidative Stress and Inflammation
2.5. Protein Aggregation and Total Lipids
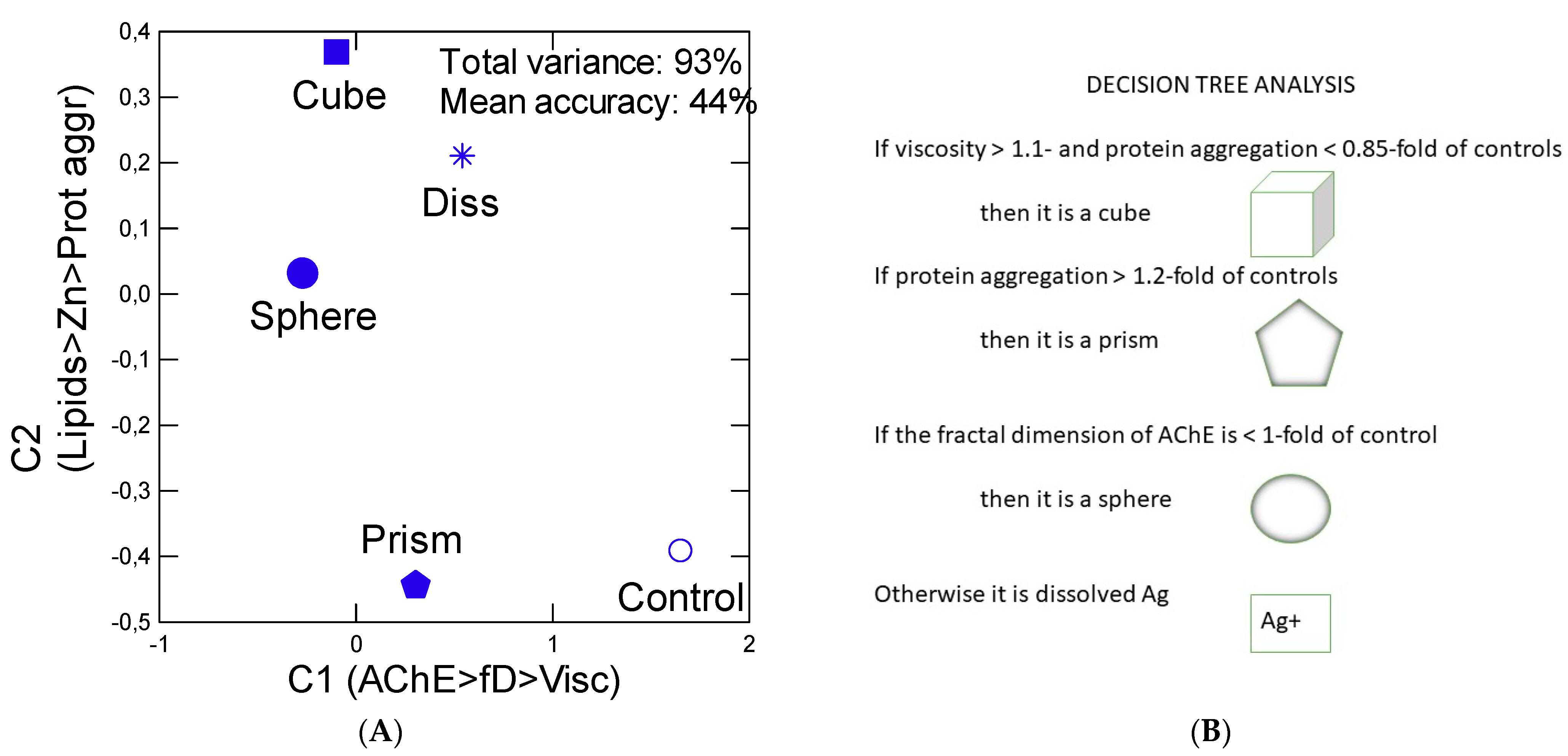
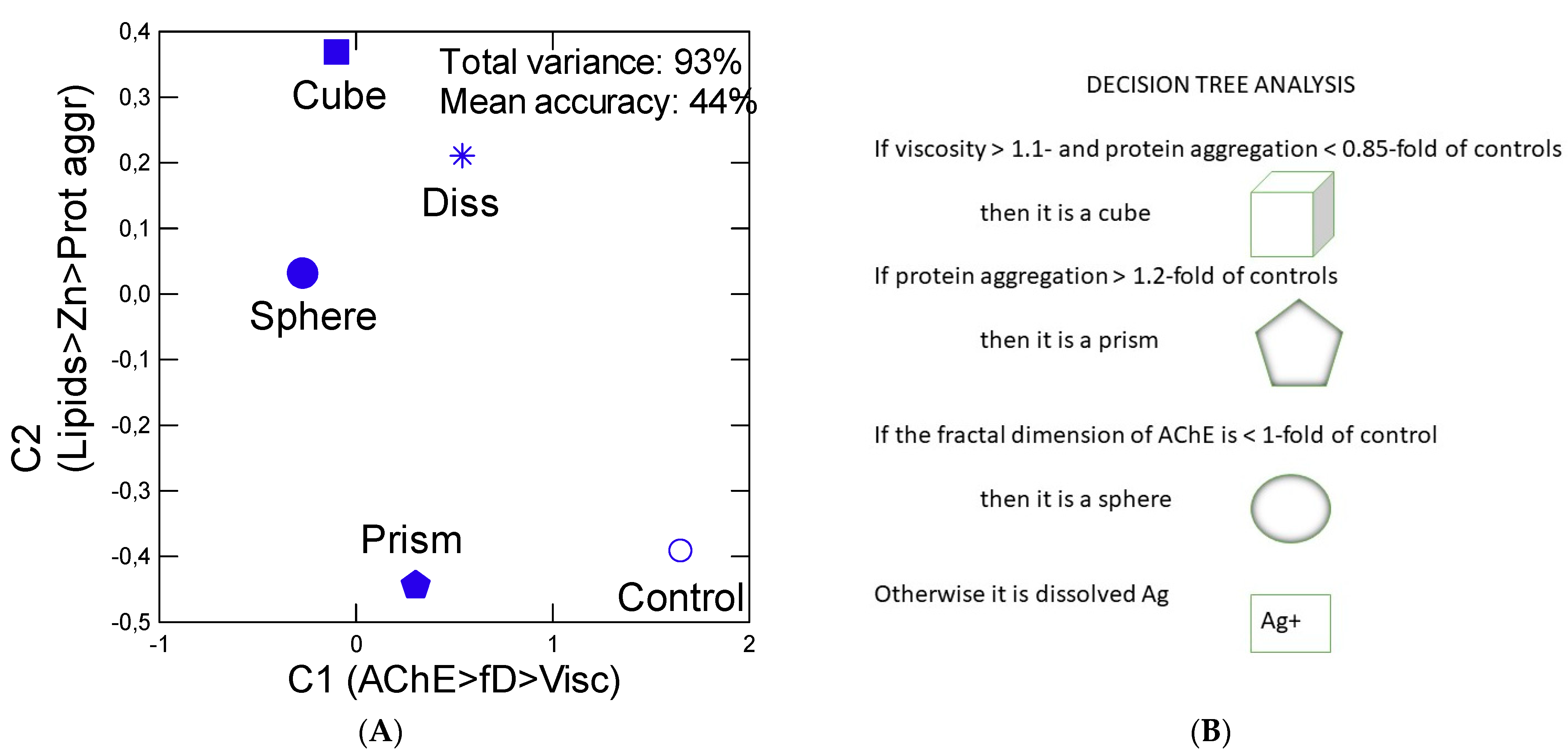
2.6. Data Analysis
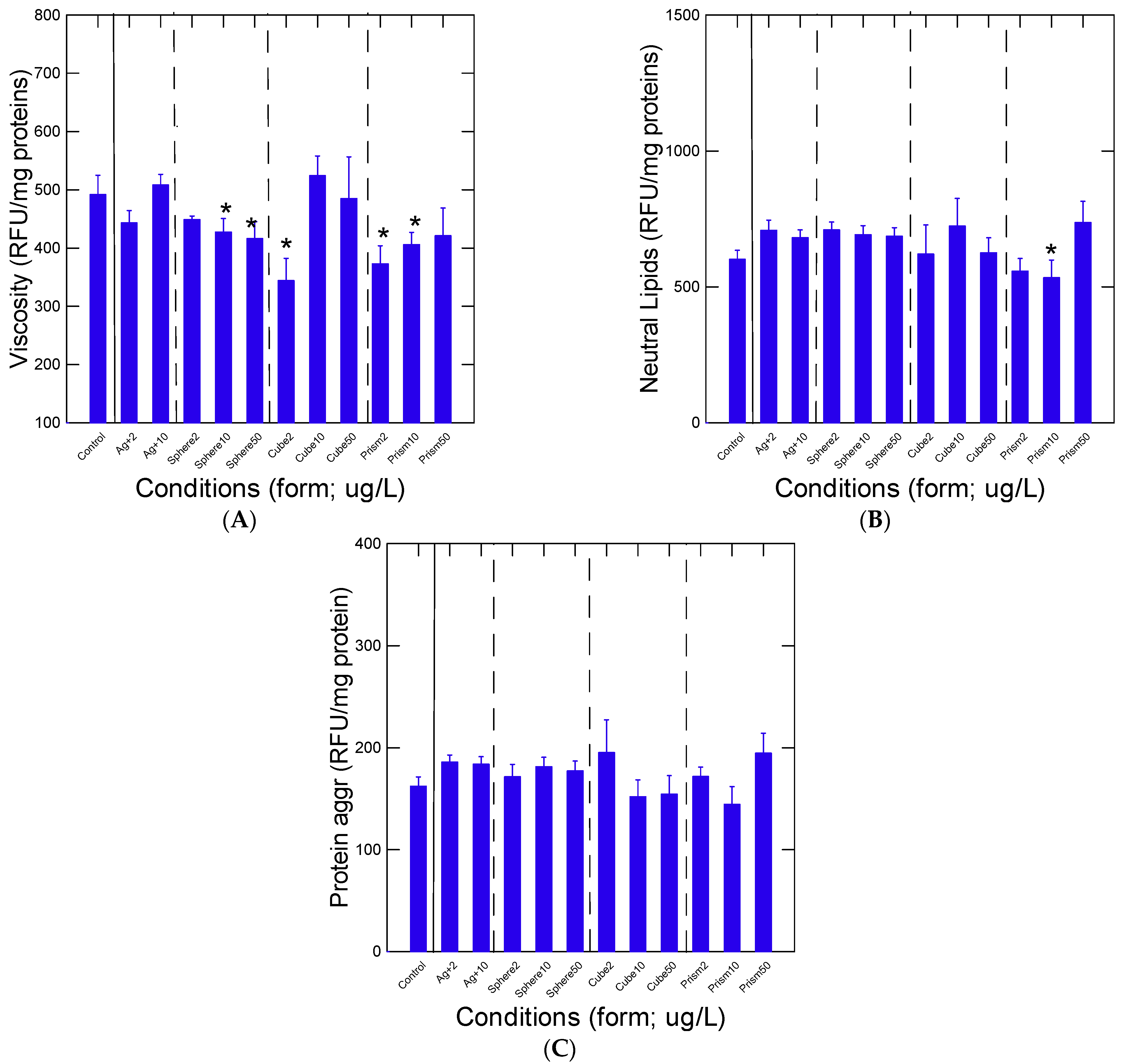
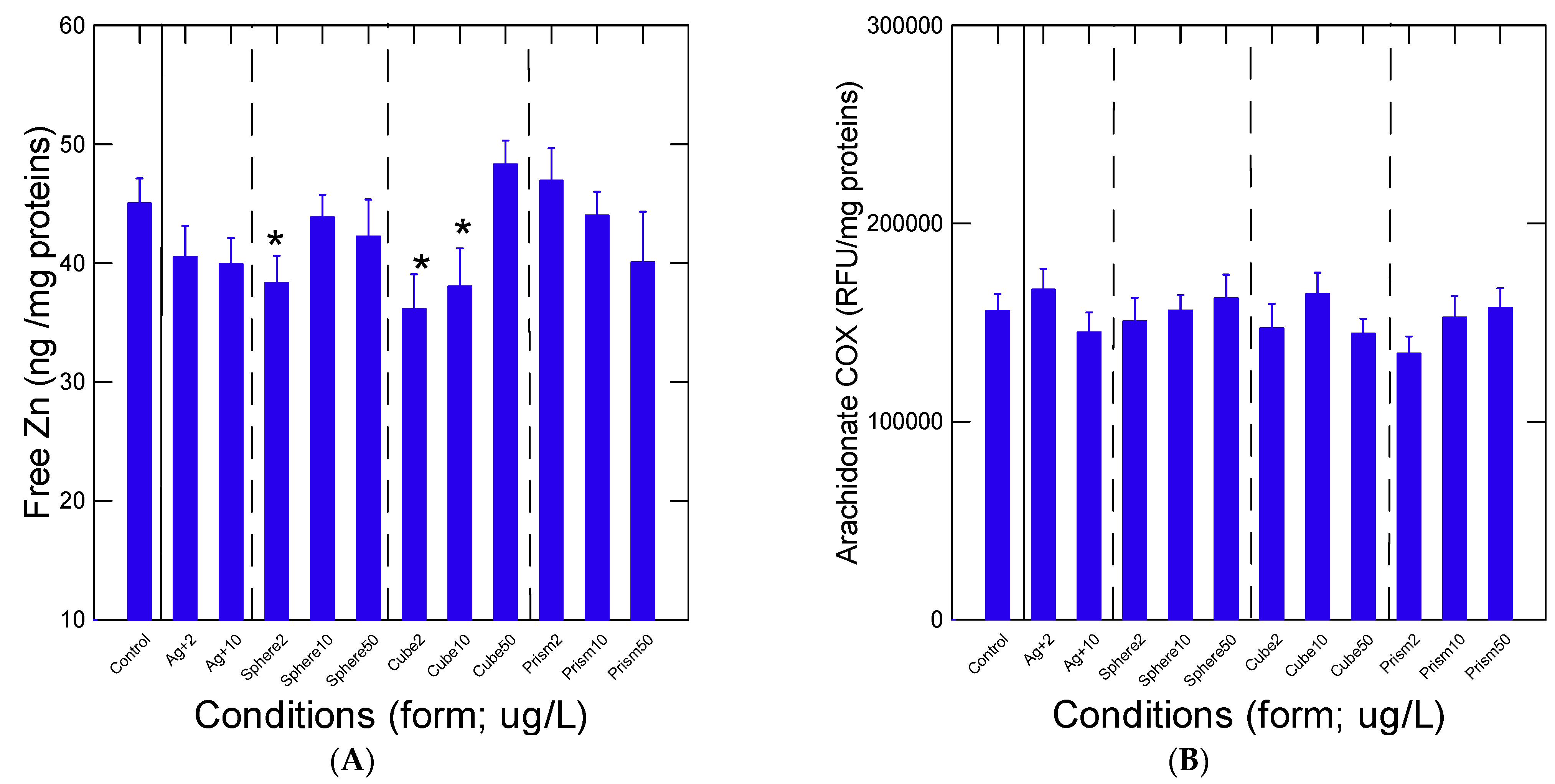
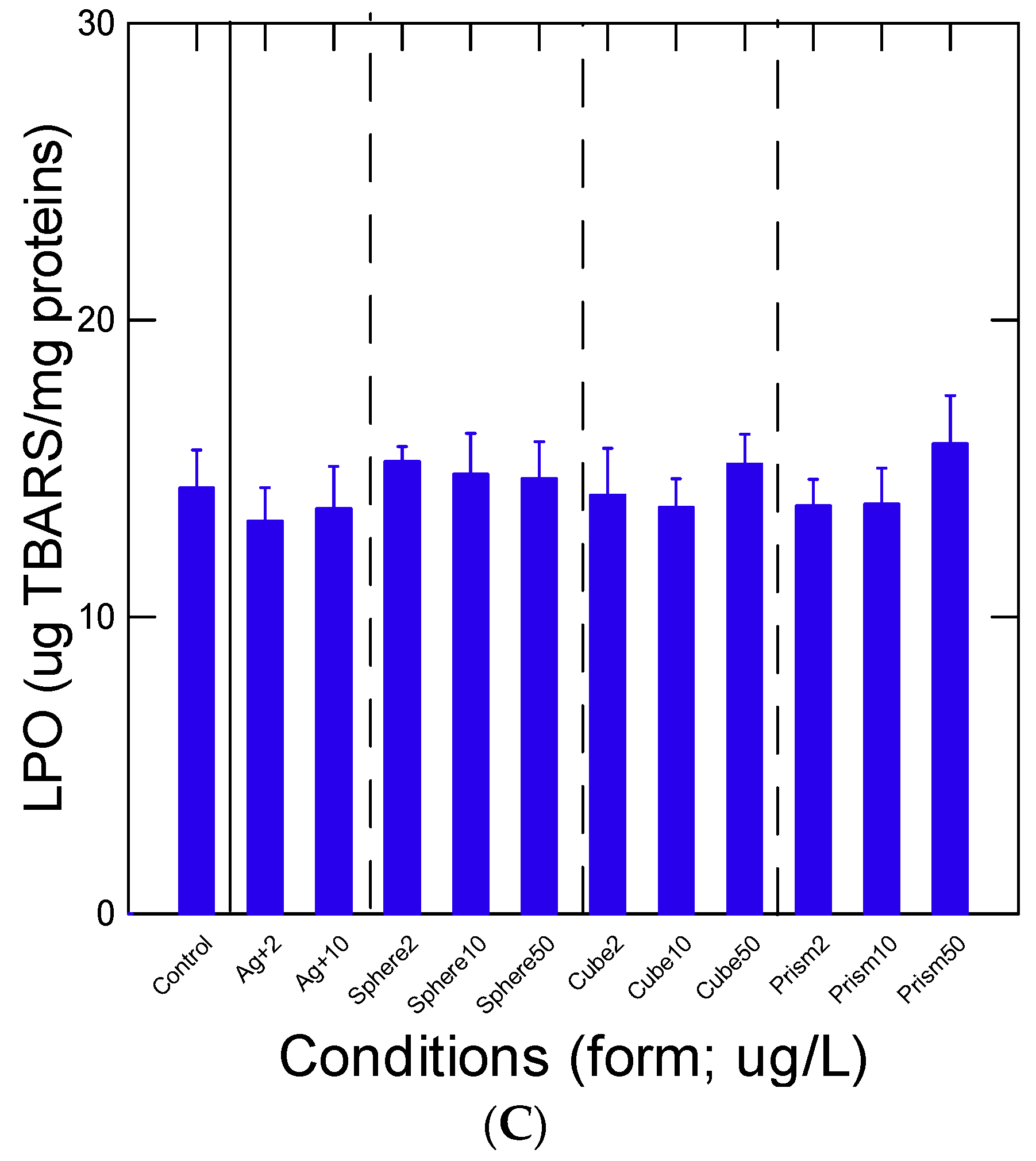
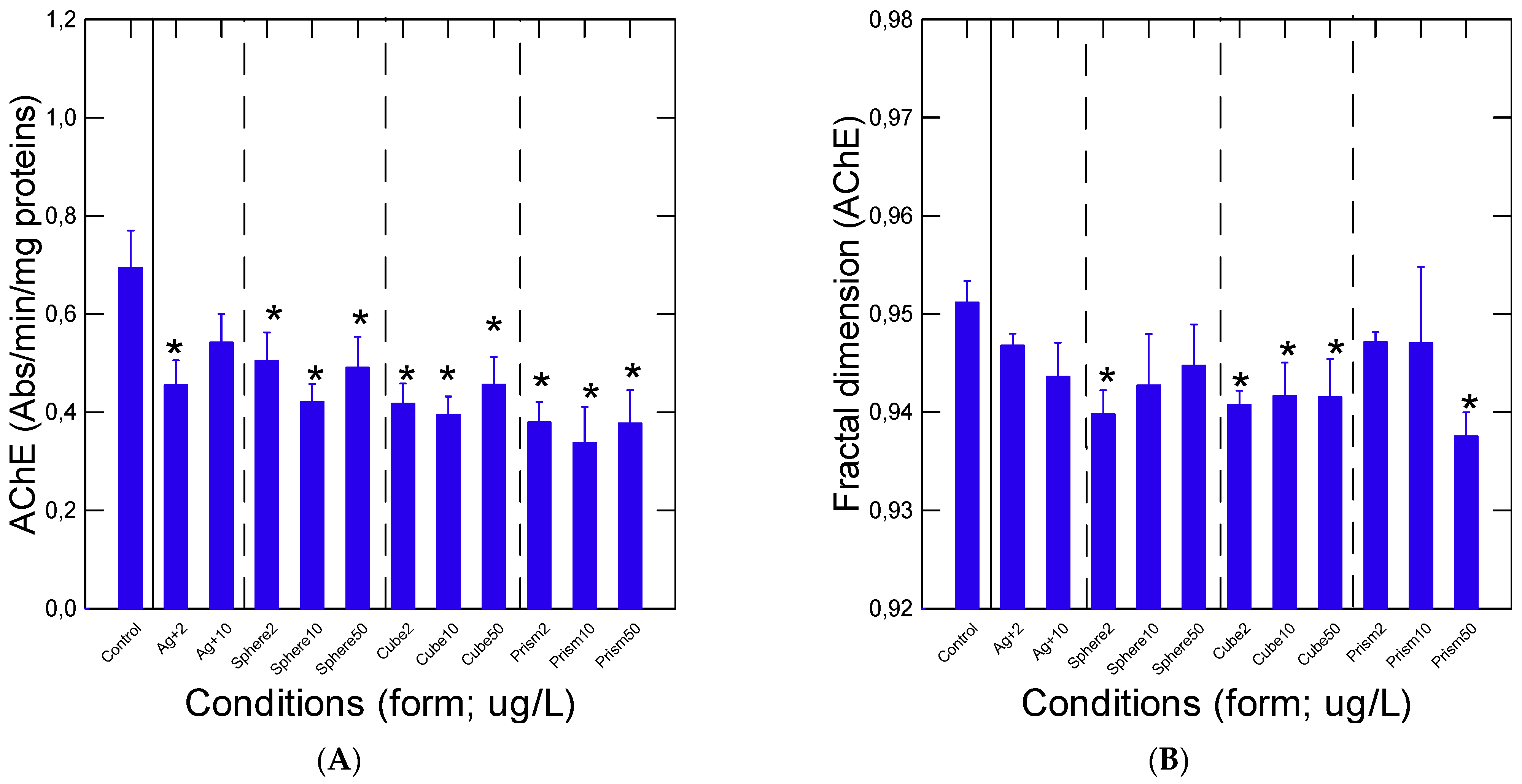
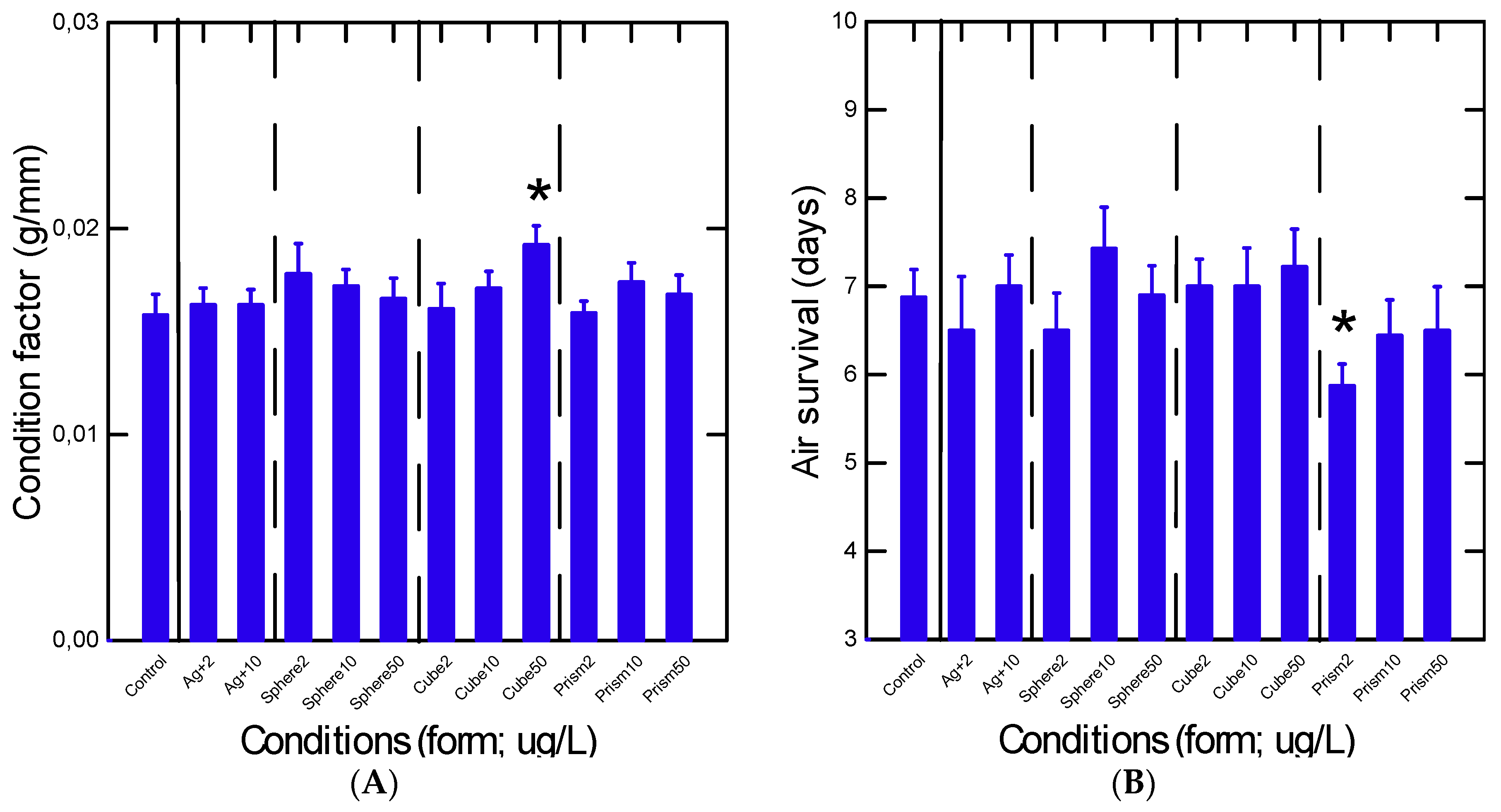
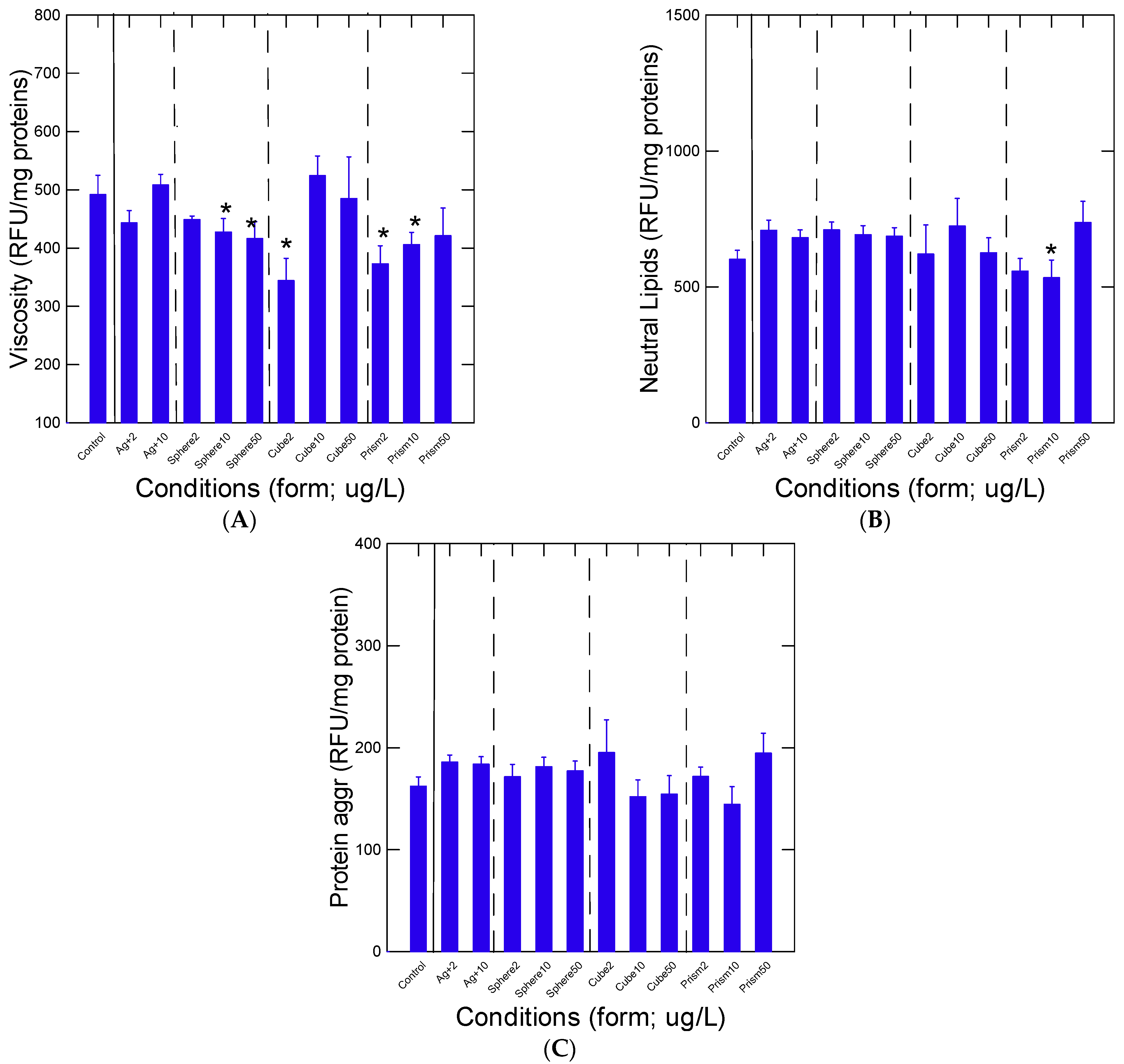
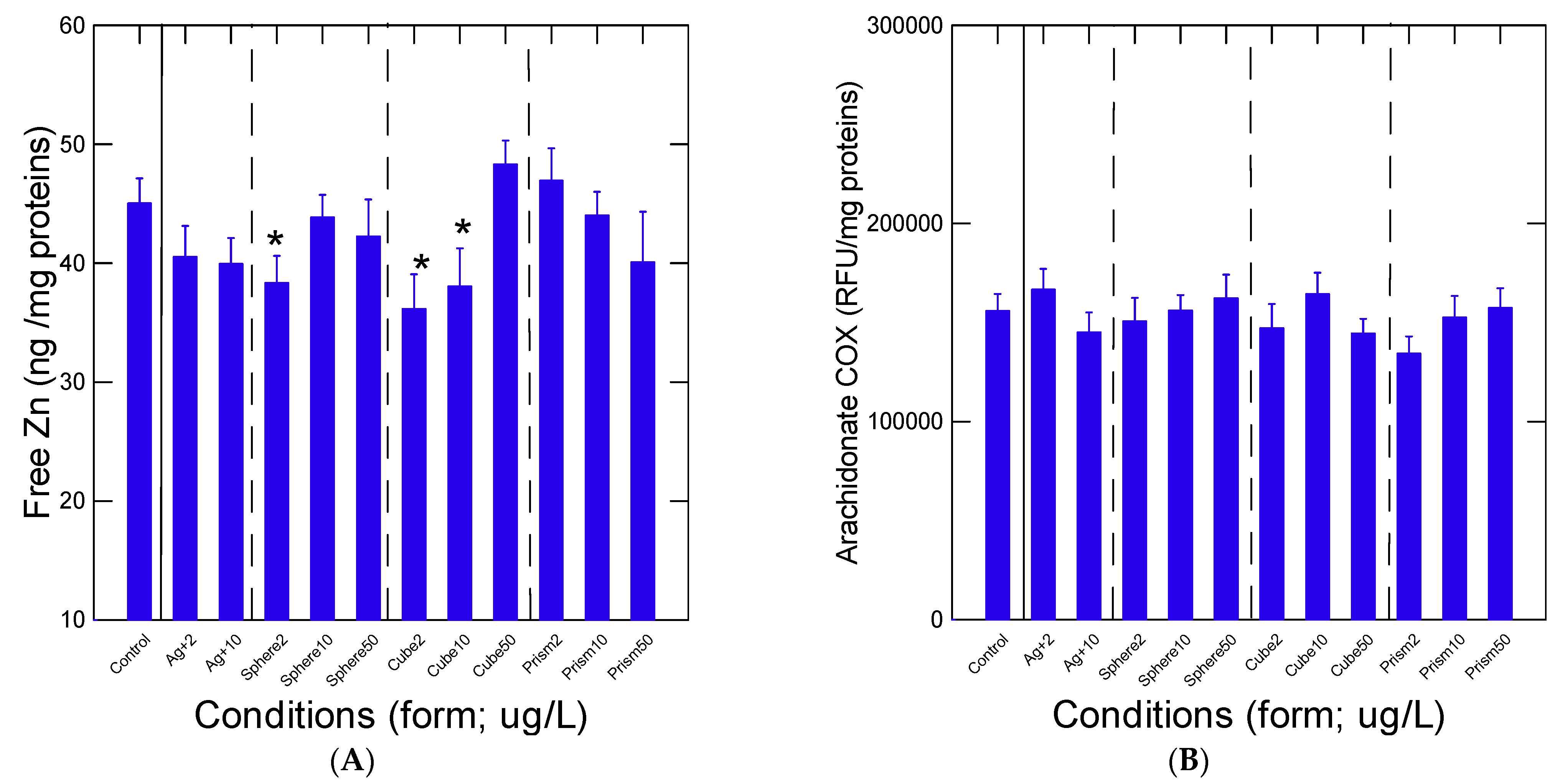
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gottardo, S.; Mech, A.; Drbohlavová, J.; Małyska, A.; Bøwadt, S.; Riego Sintes, J.; Rauscher, H. Towards safe and sustainable innovation in nanotechnology: State-of-play for smart nanomaterials. NanoImpact 2021, 21, 100297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Hu, Z.; Deng, Z. Silver nanoparticles in aquatic environments: Physicochemical behavior and antimicrobial mechanisms. Water Res. 2016, 88, 403–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGillicuddy, E.; Murray, I.; Kavanagh, S.; Morrison, L.; Fogarty, A.; Cormican, M.; Dockery, P.; Prendergast, M.; Rowan, N.; Morris, D. Silver nanoparticles in the environment: Sources, detection and ecotoxicology. Sci. Total Environ. 2017, 575, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Polesel, F.; Farkas, J.; Kjos, M.; Carvalho, P.A.; Flores-Alsina, X.; Gernaey, K.V.; Hansen, S.F.; Plósz, B.G.; Booth, A.M. Occurrence, characterisation and fate of (nano)particulate Ti and Ag in two Norwegian wastewater treatment plants. Water Res. 2018, 141, 19–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagnon, C.; Turcotte, P.; Gagné, F.; Smyth, S.A. Occurrence and size distribution of silver nanoparticles in wastewater effluents from various treatment processes in Canada. Environ. Sci. Pollut. Res. Int. 2021, 28, 65952–65959. [Google Scholar] [CrossRef]
- Formo, E.V.; Potterf, C.B.; Yang, M.; Unocic, R.R.; Leonard, D.N.; Pawel, M. How a Nanostructure’s Shape Affects its Lifetime in the Environment: Comparing a Silver Nanocube to a Nanoparticle When Dispersed in Aqueous Media. Environ. Sci. Technol. 2016, 50, 7082–7089. [Google Scholar] [CrossRef]
- Tsuji, M.; Gomi, S.; Maeda, Y.; Matsunaga, M.; Hikino, S.; Uto, K.; Tsuji, T.; Kawazumi, H. Rapid transformation from spherical nanoparticles, nanorods, cubes, or bipyramids to triangular prisms of silver with PVP, citrate, and H2O2. Langmuir 2012, 28, 8845–8861. [Google Scholar] [CrossRef]
- Wang, X.; Wu, H.-F.; Kuang, Q.; Huang, R.-B.; Xie, Z.-X.; Zheng, L.S. Shape-dependent antibacterial activities of Ag2O polyhedral particles. Langmuir 2010, 26, 2774–2778. [Google Scholar] [CrossRef]
- Ashkarran, A.A.; Ghavami, M.; Aghaverdi, H.; Stroeve, P.; Mahmoudi, M. Bacterial effects and protein corona evaluations: Crucial ignored factors in the prediction of bio-efficacy of various forms of silver nanoparticles. Chem. Res. Toxicol. 2012, 25, 1231–1242. [Google Scholar] [CrossRef]
- Auclair, J.; Peyrot, C.; Wilkinson, K.J.; Gagné, F. The geometry of the toxicity of silver nanoparticles to freshwater mussels. Comp. Biochem. Physiol. 2021, 239C, 108841. [Google Scholar] [CrossRef]
- Baltazar, G.C.; Guha, S.; Lu, W.; Lim, J.; Boesze-Battaglia, K.; Laties, A.M.; Tyagi, P.; Kompella, U.B.; Mitchell, C.H. Acidic nanoparticles are trafficked to lysosomes and restore an acidic lysosomal pH and degradative function to compromised ARPE-19 cells. PLoS ONE 2012, 7, e49635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaumbekova, S.; Torkmahalleh, M.A.; Shah, D. Impact of ultrafine particles and secondary inorganic ions on early onset and progression of amyloid aggregation: Insights from molecular simulation. Environ. Poll. 2021, 284, 117147. [Google Scholar] [CrossRef] [PubMed]
- Myrzakhanova, M.; Gambardella, C.; Falugi, C.; Gatti, A.M.; Tagliafierro, G.; Ramoino, P.; Bianchini, P.; Diaspro, A. Effects of nanosilver exposure on cholinesterase activities, CD41, and CDF/LIF-like expression in zebrafish (Danio rerio) larvae. Biomed. Res. Int. 2013, 2013, 205183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Pan, J.-F.; Hunt, D.E.; Chen, M.; Wang, B. Organic matter modifies biochemical but not most behavioral responses of the clam Ruditapes philippinarum to nanosilver exposure. Mar. Environ. Res. 2018, 133, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Zucker, R.M.; Ortenzio, J.; Degn, L.L.; Lerner, J.M.; Boyes, W.K. Biophysical comparison of four silver nanoparticles coatings using microscopy, hyperspectral imaging and flow cytometry. PLoS ONE 2019, 14, e0219078. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Blaise, C.; Gagné, F.; Burgeot, T. Three simple biomarkers useful in conducting water quality assessments with bivalve mollusks. Environ. Sci. Pollut. Res. 2016, 24, 27662–27669. [Google Scholar] [CrossRef]
- Rank, J.; Lehtonen, K.K.; Strand, J.; Laursen, M. DNA damage, acetylcholinesterase activity and lysosomal stability in native and transplanted mussels (Mytilus edulis) in areas close to coastal chemical dumping sites in Denmark. Aquat. Toxicol. 2007, 84, 50–61. [Google Scholar] [CrossRef]
- Gagné, F. Ecotoxicology of altered fractal organization in cells. Am. J. Biomed. Sci. Res. 2020, 8, 498–502. [Google Scholar] [CrossRef]
- Schepers, H.E.; van Beek, J.H.G.M.; Bassingthwaighte, J.B. Four methods to estimate the fractal dimension from self-affine signals. IEEE Eng. Med. Biol. Mag. 2002, 11, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Gagné, F.; Auclair, J.; Turcotte, P.; Gagnon, C.; Peyrot, C.; Wilkinson, K. The influence of surface waters on the bioavailability and toxicity of zinc oxide nanoparticles in freshwater mussels. Comp. Biochem. Physiol. 2019, 219C, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Oxidative metal release from metallothionein via zinc-thiol/disulfide interchange. Proc. Natl. Acad. Sci. USA 1994, 91, 237–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wills, E.D. Evaluation oflipid peroxidation in lipids and biological membranes. In Biochemical Toxicology. A Practical Approach; Snell, K., Mullock, B., Eds.; IRL Press: Oxford, UK, 1987; pp. 127–152. [Google Scholar]
- Gagné, F.; Blaise, C. Available intracellular Zn as a potential indicator of heavy metal exposure in rainbow trout hepatocytes. Environ. Toxicol. 1996, 11, 319–325. [Google Scholar] [CrossRef]
- Cabaleiro-Lago, C.; Quinlan-Pluck, F.; Lynch, I.; Dawson, K.A.; Linse, S. Dual effect of amino modified polystyrene nanoparticles on amyloid β protein fibrillation. ACS Chem. Neurosci. 2010, 1, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Greenspan, P.; Mayer, E.P.; Fowler, S.D. Nile red: A selective fluorescent stain for intracellular lipid droplets. J. Cell Biol. 1985, 100, 973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellali, M.; Khallouli, A.; Harrath, A.H.; Falodah, F.; Alwasel, S.; Beyrem, H.; Gyedu-Ababio, T.; Rohal-Lupher, M.; Boufahja, F. Effects of Au/TiO2 metallic nanoparticles on Unio ravoisieri: Assessment through an oxidative stress and toxicity biomarkers. Environ. Sci. Poll. Res. Int. 2021, 28, 18176–18185. [Google Scholar] [CrossRef]
- Auclair, J.; Turcotte, P.; Gagnon, C.; Peyrot, C.; Wilkinson, K.J.; Gagné, F. The influence of surface coatings of silver nanoparticles on the bioavailability and toxicity to Elliptio complanata Mussels. J. Nanomater. 2019, 2019, 7843025. [Google Scholar] [CrossRef]
- Young, A.; Protheroe, A.; Lukowiak, K. Silver nanoparticles alter learning and memory formation in an aquatic organism, Lymnaea stagnalis. Environ. Pollut. 2017, 225, 403–411. [Google Scholar] [CrossRef]
- Pan, J.-F.; Buffet, P.E.; Poirier, L.; Amiard-Triquet, C.; Gilliland, D.; Joubert, Y.; Pilet, P.; Guibbolini, M.; de Faverney, C.R.; Roméo, M.; et al. Size dependent bioaccumulation and ecotoxicity of gold nanoparticles in an endobenthic invertebrate: The Tellinid clam Scrobicularia plana. Environ. Pollut. 2012, 168, 37–43. [Google Scholar] [CrossRef]
- Zhao, Z.; Li, G.; Liu, Q.S.; Liu, W.; Qu, G.; Hu, L.; Long, Y.; Cai, Z.; Zhao, X.; Jiang, G. Identification and interaction mechanism of protein corona on silver nanoparticles with different sizes and the cellular responses. J. Hazard. Mater. 2021, 414, 125582. [Google Scholar] [CrossRef]
- Saidani, W.; Sellami, B.; Khazri, A.; Mezni, A.; Dellali, M.; Joubert, O.; Sheehan, D.; Beyrem, H. Metal accumulation, biochemical and behavioral responses on the Mediterranean clams Ruditapes decussatus exposed to two photocatalyst nanocomposites (TiO2 NPs and AuTiO2 NPs). Aquat. Toxicol. 2019, 208, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Efthimiou, I.; Kalamaras, G.; Papavasileiou, K.; Anastasi-Papathanasi, N.; Georgiou, Y.; Dailianis, S.; Deligiannakis, Y.; Vlastos, D. ZnO, Ag and ZnO-Ag nanoparticles exhibit differential modes of toxic and oxidative action in hemocytes of mussel Mytilus galloprovincialis. Sci. Total Environ. 2021, 767, 144699. [Google Scholar] [CrossRef] [PubMed]
- Marinho, C.S.; Matias, M.V.F.; Toledo, E.K.M.; Smaniotto, S.; Ximenes-da-Silva, A.; Tonholo, J.; Santos, E.L.; Machado, S.S.; Zanta, C.L.P.S. Toxicity of silver nanoparticles on different tissues in adult Danio rerio. Fish Physiol. Biochem. 2021, 47, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Gagné, F.; Gagnon, C.; Blaise, C. Aquatic Nanotoxicology: A review. Curr. Top. Toxicol. 2007, 4, 51–64. [Google Scholar]
- Luis, L.G.; Barreto, A.; Trindade, T.; Soares, A.M.V.M.; Oliveira, M. Effects of emerging contaminants on neurotransmission and biotransformation in marine organisms—An in vitro approach. Mar. Pollut. Bull. 2016, 106, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Farkas, J.; Bergum, S.; Nilsen, E.W.; Olsen, A.J.; Salaberria, I.; Ciesielski, T.M.; Bączek, T.; Konieczna, L.; Salvenmoser, W.; Jenssen, B.M. The impact of TiO2 nanoparticles on uptake and toxicity of benzo(a)pyrene in the blue mussel (Mytilus edulis). Sci. Total Environ. 2015, 511, 469–476. [Google Scholar] [CrossRef] [Green Version]
- Spagnoletti, F.N.; Kronberg, F.; Spedalieri, C.; Munarriz, E.; Giacometti, E. Protein corona on biogenic silver nanoparticles provides higher stability and protects cells from toxicity in comparison to chemical nanoparticles. J. Environ. Manag. 2021, 297, 113434. [Google Scholar] [CrossRef]
- Chen, R.; Choudhary, P.; Schurr, R.N.; Bhattacharya, P.; Brown, J.M.; Ke, P.C. Interaction of lipid vesicle with silver nanoparticle-seru m albumin protein corona. Appl. Phys. Lett. 2010, 100, 13703–137034. [Google Scholar] [CrossRef] [Green Version]
- Auclair, J.; Gagné, F. The influence of polystyrene nanoparticles on the fractal kinetics of lactate dehydrogenase. Biochem. Biophys. Rep. 2020, 23, 100793. [Google Scholar] [CrossRef]
- Usman, A.; Lobb, K.; Pletschke, B.I.; Whiteley, C.G.; Wilhelmi, B.S. Interaction of silver nanoparticles with catechol O- methyltransferase: Spectroscopic and simulation analyses. Biochem. Biophys. Rep. 2021, 26, 101013. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Coating | Size (nm) | Surface Area (nm2) | Zeta Potential (mvolt) |
|---|---|---|---|---|
| Prismatic nAg | PVP | 78 ± 10 (equilateral prism) | 6000 | −40 |
| Spheric nAg | PVP | 72 ± 7 (diameter) | 31,416 | −45 |
| Cubic nAg | PVP | 75 ± 7 (edge) | 60,000 | −36 |
| CF | Air Surv. | AChE | FD | Free Zn | aCOX | LPO | Prot. Agg. | Lipids | |
|---|---|---|---|---|---|---|---|---|---|
| CF | 1 | ||||||||
| Air surv | 0.08 | 1 | |||||||
| AChE | 0.22 | −0.04 | 1 | ||||||
| FD | 0.04 | 0.06 | −0.09 | 1 | |||||
| Free Zn | 0.15 | −0.04 | −0.02 | 0.13 | 1 | ||||
| aCOX | 0.01 | 0.03 | 0.20 | 0.04 | 0.13 | 1 | |||
| LPO | 0.18 | 0.18 | −0.22 | 0.00 | 0.01 | −0.10 | 1 | ||
| Prot agg | −0.08 | −0.11 | −0.05 | −0.02 | −0.05 | 0.04 | 0.13 | 1 | |
| Lipids | 0.05 | −0.06 | 0.06 | −0.01 | −0.05 | 0.11 | 0.16 | 0.65 | 1 |
| Viscosity | −0.1 | −0.04 | 0.17 | 0.03 | 0.26 | 0.17 | −0.02 | 0.23 | 0.45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Auclair, J.; Peyrot, C.; Wilkinson, K.J.; Gagné, F. The Influence of Silver Nanoparticle Form on the Toxicity in Freshwater Mussels. Appl. Sci. 2022, 12, 1429. https://doi.org/10.3390/app12031429
Auclair J, Peyrot C, Wilkinson KJ, Gagné F. The Influence of Silver Nanoparticle Form on the Toxicity in Freshwater Mussels. Applied Sciences. 2022; 12(3):1429. https://doi.org/10.3390/app12031429
Chicago/Turabian StyleAuclair, Joelle, Caroline Peyrot, Kevin J. Wilkinson, and François Gagné. 2022. "The Influence of Silver Nanoparticle Form on the Toxicity in Freshwater Mussels" Applied Sciences 12, no. 3: 1429. https://doi.org/10.3390/app12031429
APA StyleAuclair, J., Peyrot, C., Wilkinson, K. J., & Gagné, F. (2022). The Influence of Silver Nanoparticle Form on the Toxicity in Freshwater Mussels. Applied Sciences, 12(3), 1429. https://doi.org/10.3390/app12031429