Abstract
Tamarix aphylla is a commonly grown plant in Saudi Arabia and its therapeutic potential is widely recognized. Here, in this study, the aim was to explore the role of T. aphylla leaf extract (TAE) on lipopolysaccharide (LPS)-induced acute lung injury (ALI) in mice and LPS-stimulated RAW 264.7 cells. The antioxidant and anti-inflammatory parameters of TAE were performed through in vitro assays by measuring H2O2 free radical scavenging and protein denaturation inhibition. After LPS administration, broncho alveolar lavage fluid (BALF) and lung tissue were obtained to evaluate the pro-inflammatory cytokine levels, histopathological alterations, immunohistochemistry and TUNEL staining. The results designate that TAE shows anti-inflammatory activity through a decrease in pro-inflammatory cytokines, TNF-α and IL-6 in LPS-stimulated RAW 264.7 cells and BALF. The antioxidant enzyme level (SOD) decreased and MDA content was increased in the LPS-induced mice. However, TAE pretreatment markedly reversed these parameters compared to the group administered LPS only (p < 0.05). The histopathological analysis demonstrated that TAE maintained the lung tissue architecture by reducing LPS-induced inflammation and congestion. The expression pattern of the Cox-2 protein and apoptosis was high in the group administered LPS only, whereas TAE pretreatment showed a reduction in the expression pattern and apoptosis. Moreover, the in vitro assay of TAE demonstrated a concentration-dependent free radical scavenging and anti-inflammatory activity. In conclusion, our results advocate the health-promoting effects of T. aphylla on LPS-induced acute lung injury, which might be due to its antioxidative and anti-inflammatory potential.
1. Introduction
Acute lung injury (ALI) is a clinical ailment that causes disturbance in the lung endothelium, as well as loss of lung function [1,2]. The incidence of acute lung injury is still high, and it is linked with high morbidity and mortality globally, particularly in developing countries [3]. Moreover, ALI may progress to its most severe form, acute respiratory distress syndrome (ARDS) [4].
Gram-negative bacterial infection is one of the utmost significant causes of ALI, and lipopolysaccharide (LPS), the main component of Gram-negative bacterial outer membranes, can cause the inflammatory response and lung injury [5,6]. Additionally, LPS is widely recognized to induce lung injury via stimulation of TNF-α, Interlukin-6 and IL-1β, etc. [7,8].
Inhibition, as well as prevention, of pro-inflammatory cytokines and their regulatory pathway has proven great results in modulating LPS-caused lung inflammation [8,9]. Despite developments in the treatment of ALI, currently used treatments cause adverse effects on health. Thus, effective treatments with fewer side effects need to be developed to overcome such problems and increase the survival rate.
Natural products have established much consideration as a hopeful source of bioactive ingredients that are potentially beneficial in the drug discovery [10]. In this regard, natural products or their bioactive components play a significant role in the management of pathogenesis, including lung injury, through modulation of various biological activities [11,12,13,14,15].
Tamarix aphylla L. is an average-sized tree commonly distributed in Saudi Arabia and has been traditionally used in folk medicine to cure various ailments [16]. Islamic religious scripture, the Holy Quran, recognizes T. aphylla with names such as Athel and Tarfaa. The Arabic names of this tree are Abal, Tarfaa, Ghaz and Athel, while the English name is Athel tamarisk [17]. T. aphylla is a rich source of different constituents such as Ellagitannins, gallotannins and gallo-ellagitannins [18]. In addition, an important compound (5-dihydroxy-4′,7-dimethoxyflavone) was isolated from T. aphylla and proved its role in the inhibition of liver pathogenesis. This compound enhances the antioxidant level and reduces lipid peroxidation, also modulating apoptotic protein expression [19]. Tamrix aphylla plays a vital role in diseases management and its role in health management has been confirmed as having anti-bacterial, anti-inflammatory, anti-pyretic, analgesic and wound-healing effects [20,21,22]. Some other plants from in Saudi Arabia show high protein content and are used to prevent oxidative damage of proteins and lipids through its rich antioxidant source [23].
This study aimed to investigate the ameliorative anti-inflammatory and antioxidant effect of the methanolic extract of Tamarix aphylla (TAE) on LPS-induced lung injury in mice, and its mechanisms associated with oxidative stress and inflammation were assessed.
2. Materials and Methods
2.1. Reagents and Chemicals
The present study used Folin–Ciocalteu’s phenol reagent, 2,2-Diphenyl-1-picrylhydrazyl (DPPH), gallic acid and lipopolysaccharide from E. coli (LPS), obtained from Sigma Aldrich Co. (St. Louis, MO, USA). Dulbecco’s modified Eagle’s medium (DMEM) and Fetal bovine serum (FBS) were obtained from Invitrogen; Thermo Fisher Scientific, Carlsbad, USA. TNF-α and IL-6 were used to measure the level of inflammatory markers by using ELISA kits, bought from Abcam, Cambridge, UK. Primary monoclonal antibodies against cyclooxygenase-2 (cox-2) were purchased from Abcam, Cambridge, UK. Biotinylated goat anti-mouse and streptavidin peroxidase were purchased from Abcam, Cambridge, UK. Superoxide dismutase (SOD) kits were purchased from Abcam, Cambridge, UK. The TUNEL assay used to detect DNA fragmentation was purchased from Abcam, Cambridge, UK. (TUNEL HRP-DAB Assay kit). Other materials were obtained from commercial sources and guaranteed the highest available grade.
2.2. Collection and Extraction of T. aphylla Leaves
The leaves of T. aphylla (Family: Tamaricaceae) were acquired from the Kingdom of Saudi Arabia. After collection, the leaves were air-dried in the dark, then milled into a dried powder. The powder was kept at 4 °C in grey-colored bottles for further examination. The extraction procedure was carefully followed to retain maximum biological components responsible for biological activities of T. aphylla. The extraction method was carried out in triplicate. The extract was prepared by using methanol (70%). 100 g dry powder was extracted with 1000 mL of 70% methanol by maceration at room temperature for three days. The extract was purified by two-step filtration using N°1 Whatman paper and filter paper (0.45 μm porosity). The collected filtrates were dried separately at 40° C using a laboratory rotary evaporator. The obtained residue or crude active extract was dissolved in water, frozen and lyophilized. The lyophilized powder was kept for storage at 4 °C and was used for phytochemical investigation, determination of antioxidant, anti-inflammatory screening and used in in vitro and in vivo studies.
The extract % yield was estimated by the following equation: Yield (%) = [extract weight/Initial sample weight] × 100.
2.3. Phytochemical Screening
The phytochemical components of the methanol extract were screened using the methods described in our previously published articles in our laboratory [24]. The components analyzed included phenolic, flavonoids, alkaloids, carbohydrates, anthraquinone, tannins and saponins.
2.4. Total Phenol Content
The total phenol content of T. aphylla leaf extract was investigated using Folin–Ciocalteu (FC) reagent, as described by Anwar and colleagues, and the absorbance of the developed color was determined at 725 nm [24]. 1 mL of crude methanol extract of T. aphylla (1 mg/mL) was mixed with 5 mL of 10% FC reagent, and the mixture was allowed to stand for 5 min at 22 °C. Finally, the mixture was mixed with 4 mL of sodium carbonate (7.5%). The absorbance was noted using a UV–Vis spectrometer after placing the mixture inside a dark space at room temperature for 90 min, and the absorbance was recorded at a constant wavelength of 760 nm. The total phenol content was investigated using a standard curve prepared with gallic acid (50–250 g/mL). Results were expressed as mg GAE (Gallic Acid Equivalent) per 100 g dry weight of lyophilized crude extract. The phenol content was carried out in triplicate.
where K means gallic acid content (mg/mL), L denotes plant extract volume (mL) and M signifies the weight of the pure plant extract (g).
Total phenolic content = K × L/M
2.5. Evaluation of the Amount of Total Flavonoids
The total flavonoid content of the methanol extract of T. aphylla was evaluated using a spectrophotometer using an aluminum chloride test, as described in our previously published article [24], with slight modifications. The standard quercetin absorbance and extract were estimated at 420 nm. A standard curve using quercetin was produced to measure the total flavonoid concentration. The results were represented in mg of quercetin equivalent per g of crude extract (mg QUE/g).
where K indicates quercetin concentration (mg/mL), L denotes extract volume (mL) and M implies pure plant extract (g) weight.
Total flavonoid amount = K × L/M
2.6. In Vitro Study
2.6.1. Antioxidant Activity: H2O2 Free Radical Scavenging Activity
The H2O2 free radical scavenging potential of the obtained extract was tested using a previously published work to confirm its antioxidant activity. A 40 mM H2O2 solution in extract (50, 75, 100, 150, 200, 250, 300, 400, 500 and 600 µg/mL) set under the dark far from sunlight. After 10 min of incubation, the H2O2 absorbance was estimated against a blank solution at 230 nm. The blank contained phosphate buffer without H2O2. The results were compared to a reference for the % scavenging capability of ascorbic acid (100 µg/mL).
where L describes the control absorbance. Conversely, M signifies sample absorbance in the presence of extract.
Percentage of scavenged H2O2 = [(L − M)/L] × 100
2.6.2. DPPH Method of Free Radical Scavenging Activity
The methanol extract of T. aphylla was analyzed by a 1,1 difenyl-2-picryl-hydrazyl (DPPH) test to measure the free radical scavenging [24]. DPPH solution (0.3 mM) was prepared in methanol, and 1 mL of this solution was introduced to 2.5 mL of different concentrations of extract (50, 75, 100, 150, 200, 250, 300, 400, 500 and 600 µg/mL) prepared by serial dilution of stock solution (10 mg/mL) in methanol. Then, the reaction mixtures were thoroughly mixed, and various vials were stored at room temperature for 30 min in the absence of sunlight. The absorbance changes of each reaction mixture were recorded at 517 nm using methanol and ascorbic acid as a standard. The extract was substituted with methanol to make the control.
where L describes the control absorbance. Conversely, M signifies sample absorbance in the presence of extract.
DPPH scavenging activity (%) = [(L − M)/L] × 100
2.6.3. An Evaluation of In Vitro Anti-Inflammatory Activity by Protein Denaturation Inhibition
The anti-inflammatory activity of the methanol extract of T. aphylla was determined using a modified version of bovine serum albumin (BSA) assay, as described in our previously published articles [24]. BSA solution (1%) was prepared in tris-buffered saline (0.05 M Tris and 0.15 M sodium chloride, pH 7.6, at 25 °C). 100 µL of T. aphylla methanol extract (50, 75, 100, 200, 300, 400, 500 and 600 µg/mL) or ibuprofen (100 or 200 µg/mL) was introduced to separate vials containing BSA (500 µL, 1%). The vials were placed in the dark for 20 min. The mixtures were then heated at 71 °C for 10 min and cooled for 20 min under laboratory conditions. The level of protein precipitation or turbidity of the solutions was measured at 660 nm. The experiment was done three times.
where L describes control absorbance. Conversely, M signifies sample absorbance in the presence of extract/ibuprofen.
Percentage inhibition of protein precipitation = [(L − M)/L] × 100
2.6.4. Anti-Proteinase Action
This assay was performed according to a modified version of previously published articles [24,25]. The reaction mixtures contained 1 mL extract (50, 75, 100, 200, 300, 400, 500 and 600 μg/mL) or diclofenac sodium (100, and 200 μg/mL), 0.06 mg trypsin, as well as tris HCl buffer. Casein was added in each tube after the incubation. Furthermore, 70% perchloric acid (2 mL) was mixed to arrest the reaction. The resultant suspension was centrifuged, and the supernatant absorbance was recorded at 210 nm. The % inhibition of proteinase inhibitory activity was estimated as.
where L represents control absorbance. Conversely, M signifies sample absorbance.
Inhibition % = [(L − M)/L] × 100
2.6.5. Cell Culture
RAW 264.7 cells were cultured and maintained in DMEM comprising 10% fetal bovine serum and cells were incubated at 37 °C in 5% CO2. Cells were pretreated with extract for 1 h before LPS (1 μg/mL) stimulation for 6 h. Vehicle (1% DMSO in PBS)-treated cells were used as controls.
2.6.6. Cell Viability Assay
To measure the cytotoxicity of the extract, MTT experiments were performed. In brief, RAW 264.7 cells were planted into 96-well plates at a density of 1 × 105 /mL. The cells were then treated with extract (25 and 50 μg/mL) for 24 h. Subsequently, MTT solution (15 μL) was added to each well and incubated at 37 °C for 4 h. The supernatant was removed, and optical density was measured [26].
2.6.7. Measurement of Inflammatory Cytokines
Culture RAW 264.7 cells were seeded in a 24-well plate, with almost 2 × 105 cells/well for 24 h; 1 × 105 cells were pretreated with extract for 1 h, and then initiated an inflammatory response by LPS (1 μg/mL) for 18 h. The concentrations of TNF-α and IL-6 in the supernatant were measured by using ELISA kits. The absorbance was measured at 450 nm by using a microplate reader.
2.7. In Vivo Study
2.7.1. Animal Model
All experimental mice weighing 18–24 g was purchased from King Saud University, Saudi Arabia, and used in this study to achieve the objectives accordingly. The animal house was used to keep all animals for complete one-week acclimatization to minimize stress. Conditions were maintained at approximately 23 ± 2 °C temperature, 45–65% relative humidity and a 12 h/12 h dark/light cycle in the animal house, and mice received normal water and food ad libitum. The CAMS ethical committee, Qassim University, approved the use of the animal model for this study (No.10194-Cams1-2020-1-3-I). Guidelines of NIH, USA, were followed for the care and experimental use of animals.
2.7.2. Grouping of Animals and Treatment Plan
Animals were randomly divided into four groups of eight mice in each group: 1. control group: mice were given the vehicle once; 2. LPS group: 5 mg/kg of LPS [27,28] was given intraperitoneally to induce acute lung injury; 3. LPS + extract (100 mg/kg) group [21]. 4. LPS + Dexamethasone group, considered as positive control group as dexamethasone (10 mg/kg) [29] prior to LPS administration. All the mice were alive 24 h after the LPS treatment. After 24 h of LPS injection, all the mice were executed by cervical dislocation. Then, the blood sample, bronchoalveolar lavage fluid (BALF) and lung tissue were collected to achieve the aim of the study. The left lung was collected to measure the wet/dry ratio (W/D), whereas the right lobe of the lung was collected and fixed in 10% formalin for a histopathological, immunohistochemistry and TUNEL staining. The remaining lung tissues were properly homogenized and stored at −20 °C for analysis of other biochemical parameters.
2.7.3. Measurement of Wet-to-Dry Weight (W/D) Ratio of Lung
Mice were sacrificed at 24 h after administration of LPS (5 mg/kg). The lungs were collected, and the wet-to-dry weight (W/D) ratio of lung was determined. Briefly, the fresh lung tissues from all experimental groups were removed to determine the wet weight. Later, the samples were dried in an oven at 80 °C for 48 h to obtain the dry weight. The values of the wet weight divided by the dry weight were measured as the ratio of wet to dry [30].
2.7.4. Measurement of Inflammatory Cytokines in BALF
To access the effects of the extract on the production of inflammatory cytokines in LPS-induced ALI rats, levels of TNF-α and IL-6 in bronchial alveolar lavage fluid (BALF) were determined as per the manufacturer’s instruction. Each sample was used in duplicate, mean values were employed for calculation and results were interpreted accordingly.
2.7.5. Measurement of Lipid Peroxidation and Superoxide Dismutase
Malondialdehyde (MDA), the main indicator of lipid peroxidation, was measured in the lung tissues by using a thiobarbituric acid-reactive substance (TBARS). Lung tissues were homogenized as well and then centrifuged. At 535 nm, presence of a TBARS was noticed by examining the changes in absorbance using a microplate reader [31]. Superoxide dismutase (SOD) was determined in lung tissues as per instructions of the manufacturer. The absorbance was taken at a particular wavelength by using a microplate reader, and the result was analyzed consequently.
2.7.6. Histopathological Evaluation
The lung tissues from all experimental animals were cut, collected and fixed in 10% formalin. Then, tissues were processed using an automated tissue processor, and paraffin-embedded blocks were made. 5-µm thin sections were cut using a rotatory microtome, and hematoxylin and eosin (HE) staining was performed to stain the tissues properly. The alterations in lung tissues, including edema, congestion, intra-alveolar cell infiltration and hemorrhage, were examined using a light microscope, and the results were interpreted consequently.
2.7.7. Immunohistochemical Analysis of Protein Expressions
The expression pattern of the Cox-2 protein in the all-experimental group of lung tissue was examined using immunohistochemical staining. In short, the tissue sections were deparaffinized with series of xylene treatment and the tissues were rehydrated. The antigen site was unmasked through antigen retrieval in 50 mM citrate buffer (pH 6.0) for 22 min at boiling temperature (100 °C) in a pressure cooker, followed by 10–15 min cooling at room temperature. The endogenous peroxidase activity was quenched using in 3% H2O2 solution for 15 min. Afterward, a protein block was used to block the unwanted site for 20 min. The primary antibody cox-2 (Abcam, Cambridge, UK) (1:250 dilution) was used on all sections and incubated for 1 h at room temperature. The 2° biotinylated antibody and streptavidin peroxidase were added using the DAB staining kit from Abcam. The scoring for protein expressions was calculated by two independent pathologists in a blinded way. The positivity score was evaluated semi-quantitatively, in which the staining intensity was multiplied by the percentage of positive cells. Images were captured using a Leica light microscope attached with camera.
2.7.8. TUNEL Assay
TUNEL assay was run on all animal lung tissue sections to measure the apoptosis with a kit from Abcam. The stained nuclei were evaluated using an Olympus light microscope (Olympus, Tokyo, Japan) for the identification of DNA fragmentation. The nuclei with TUNEL positivity were calculated, and the images were captured using a Leica light microscope attached with a camera.
2.8. Statistical Analysis
All obtained data were expressed in mean ± SD. One-way analysis of variance (ANOVA) was used to make significance of the differences among different groups. The significant difference (p < 0.05) was measured between the different experimental groups.
3. Results
3.1. Flavonoid, Phenolic and Preliminary Screening
The medicinal properties of T. aphylla are linked with its phytochemical contents producing definite physiological actions in the human body. The most important of these components are alkaloids, tannins, flavonoids and phenolic compounds. Table 1 shows the odor, color, texture, as well as % yield of T. aphylla leaf extract. Table 2 shows the results of leaf extract phytochemical screening of several phyto-constituents. Total phenol content levels in leaf extracts were 371.30 ± 1.27 mg gallic acid (GA) equivalent/g extract dry weight. Carbohydrate reduction, especially fructose, ascorbic acid and protein, show an impact on total polyphenol content. Total flavonoid content in methanolic extracts of leaves was 215.31 ± 1.212 mg quercetin (Qu) equivalents/g extract dry weight, respectively.

Table 1.
Preliminary assessment of T. aphylla methanol extract.
Table 2.
Phytochemical screening of T. aphylla leaf extract.
3.2. H2O2 Reducing Ability
There is a clear link between antioxidant activity and reducing ability. Additionally, antioxidant activity can be evaluated by investigating hydrogen peroxide reducing potential and DPPH radical scavenging capabilities. The current study shows that H2O2 reducing activity of T. aphylla leaf extract increases with the increase in its concentration (Figure 1). The capacity of the phenolic components of the methanol extract to donate electrons may explain the reduction in H2O2. The dissolution of H2O2 results in the formation of hydroxyl radicals. Therefore, this activity of T. aphylla is considered to be very beneficial.
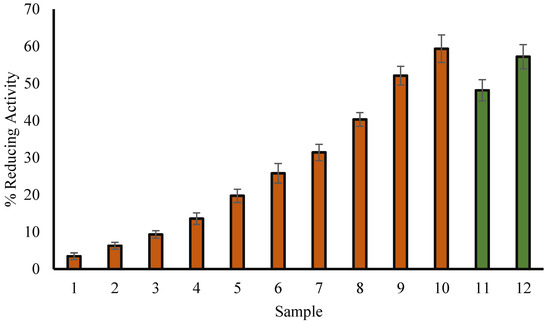
Figure 1.
Percentage H2O2 reducing activity of methanol extract of T. aphylla leaves (red column) and ascorbic acid (green column). Samples 1 to 10 correspond to various concentrations of extract (50, 75, 100, 150, 200, 250, 300, 400, 500 and 600 µg/mL). Samples 11 and 12 (green column) correspond to 100 and 200 µg/mL of ascorbate. The results are represented as means ± SEM (n =3, p < 0.05).
3.3. Scavenging Assay for DPPH Radicals
It has already been reported that the concentration of natural product extracts and the percentage of DPPH radical scavenging activity are correlated to each other [24]. The present study shows concentration-dependent DPPH scavenging activity comparable to ascorbic acid, indicating the antioxidant nature of T. aphylla (Figure 2). In a bar graph, Figure 2 compares the percent of free radical scavenging activity of T. aphylla leaf extract (50, 75, 100, 150, 200, 250, 300, 400, 500 and 600 µg/mL; red) and ascorbic acid (100 and 200 µg/mL; green).
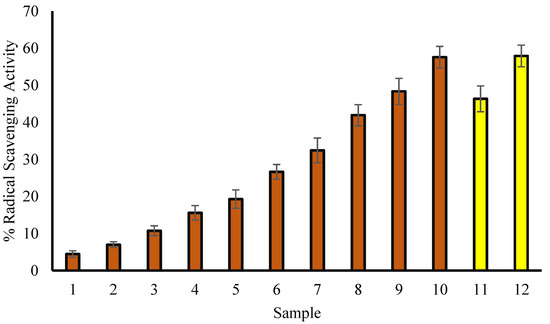
Figure 2.
The percent of free radical scavenging activity. Samples 1 to 10 correspond to various concentrations of extract (50, 75, 100, 200, 300, 400, 500 and 600 µg/mL). Samples 11 and 12 (yellow column) correspond to 100 and 200 µg/mL of ascorbic acid. The results are described as means ± SEM (n = 3, p < 0.05).
3.4. Protein Denaturation Inhibition: An Evaluation of Anti-Inflammatory Activity
Several factors, such as chemicals or stress, can lead to denaturation of proteins, and it plays a key role in inflammation. Hence, the possible anti-inflammatory effect of T. aphylla methanol extract was studied in terms of its ability to protect proteins from denaturation. According to the current study, methanol extract of T. aphylla leaves is effective against denaturation of bovine serum albumin (BSA) (Figure 3), especially at 600 µg/mL (Figure 3). At 200 µg/mL concentration, ibuprofen (anti-inflammatory drug) exhibited the inhibition of BSA by 66.89%.
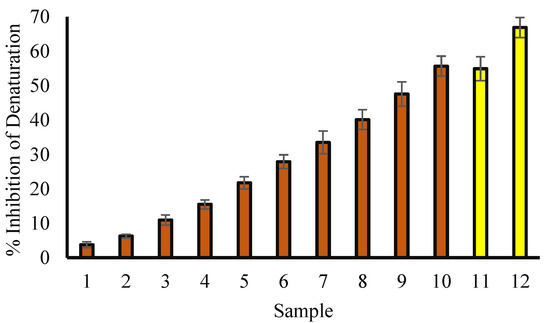
Figure 3.
Percentage protection of heat-induced protein denaturation. Red columns (from 1–10) at X-axis represent concentrations of 50, 75, 100, 150, 200, 250, 300, 400, 500 and 600 µg/mL of T. aphylla methanol extract. Yellow columns (11 and 12) at X-axis represent concentrations of ibuprofen of 100 and 200 µg/mL.
3.5. Anti-Proteinase Potential
Proteinases are found in lysosomes and contribute to tissue damage in inflammatory reactions. The protecting agents against proteinase action might be ineffective for tissue damage induced by proteinase [32]. In the current study, methanolic leaf extract of T. aphylla is found to inhibit proteinase activity. Furthermore, the extract displayed a concentration-dependent anti-proteinase activity, and the maximum proteinase inhibition activity was noticed at 600 μg/mL (72.16%), as shown by the data (Figure 4). However, diclofenac sodium showed a maximum inhibition of 69.37 at 200 µg/mL.
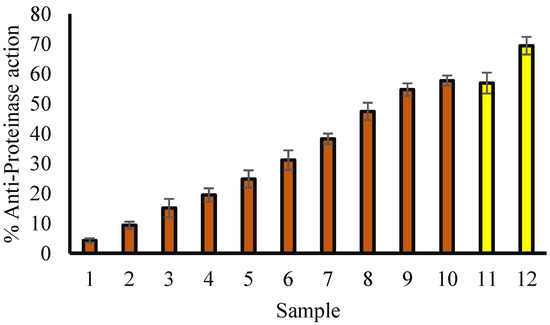
Figure 4.
Anti-proteinase activity of methanol extract of T. aphylla leaves. Red columns (1–10) represent various concentrations (50, 75, 100, 150, 200, 250, 300, 400, 500 and 600 µg/mL) of T. aphylla methanol extract. Yellow columns (11 and 12) show numerous concentrations of ibuprofen (100 and 200 µg/mL).
3.6. Effect of Extract on Cell Viability
MTT assay was carried out to estimate cytotoxicity of the extract at several concentrations (100–1000 μg/mL). The extract showed no cytotoxicity under a concentration of 100–600 μg/mL (Figure 5), and a mild cytotoxic effect was noticed at concentrations of 800 and 1000 μg/mL. No significant difference was observed among concentrations of 100–600 μg/mL.
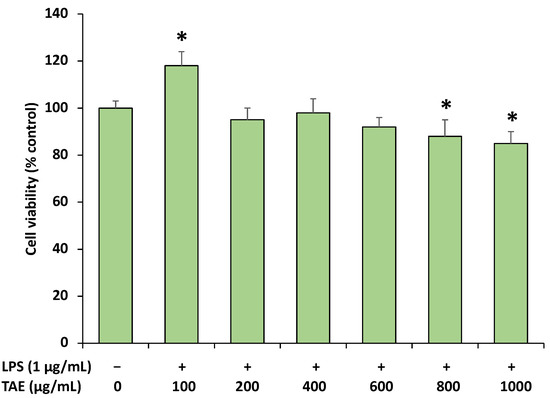
Figure 5.
Effects of extract on cell viability in RAW 264.7 macrophages. The cells were treated with the extract (0, 100, 200, 400, 600, 800 and 1000 μg/mL) in the presence of LPS (1 μg/mL) for 20 h. MTT assay was used to examine the cell viability. Data are presented as mean ± SD (n = 3). Mean values denoted by (*) are significantly different (p < 0.05).
3.6.1. Effects of Leaf Extract on Inflammatory Markers in RAW 264.7 Cells Stimulated by LPS
As a strong pro-inflammatory cytokine, TNF-α exhibits a substantial role in the immune system during inflammatory process [33]. In this study, the inhibitory role of the extract was investigated on the release of proinflammatory cytokines such as TNF-α and IL-6 in RAW 264.7 cells. Treatment of RAW 264.7 macrophages with LPS caused a noteworthy increase in IL-6 and TNF-α in the supernatants. However, the extract significantly decreased the LPS-induced production of TNF-α and IL-6 in a concentration-dependent manner (Figure 6). These results designate that the extract shows anti-inflammatory activity through a decrease in pro-inflammatory cytokines TNF-α and IL-6 in LPS-stimulated RAW 264.7 cells.
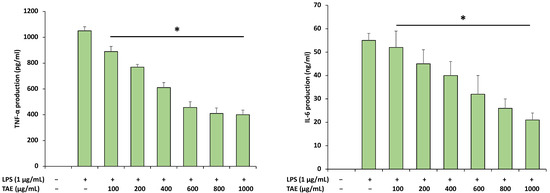
Figure 6.
Anti-inflammatory extract on LPS-induced RAW 264.7 macrophages. Cells were pre-treated with various concentrations of extract for 1 h and treated with LPS for an additional 20 h. IL-6 and TNF-α were measured by an enzyme-linked immunosorbent assay (ELISA) kit. The data are presented as means ± SD (n = 3). (*, p < 0.05 LPS group vs. extract group).
3.6.2. Effects of the Extract on in the Total Number of Cells, Neutrophils and Macrophage Cell Counts
After 24 h of LPS administration, neutrophils, macrophages and total number of cells were counted in different groups in the bronchial alveolar lavage fluid (BALF). LPS meaningfully increased the total number of cells (18.3 × 105 /mL), neutrophils (12.2 × 105 /mL) and macrophages (0.38 × 105 /mL) compared to the control group. However, extract pretreatment of the LPS administration showed a substantial decrease in the total number of cells (12.2 × 105 /mL and 8.4 × 105 /mL), neutrophils (7.3 × 105 /mL and 6.4 × 105 /mL) and macrophages (0.28 × 105 /mL and 0.22 × 105 /mL) (Figure 7).
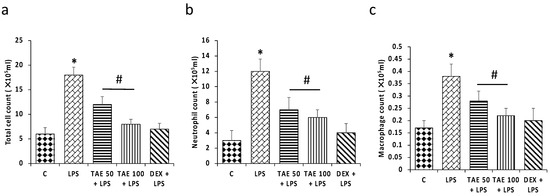
Figure 7.
Effects of extract on neutrophil, macrophages and total cell counts. The figure (a) shows total cells, (b) neutrophils and (c) macrophages in various experimental groups. The noteworthy changes (p < 0.05) are shown by (*) between the control group and LPS treatment groups, and (#) between the LPS treatment group and LPS + extract groups.
3.6.3. Effects of the Extract on TNF-α and IL-6 Level in LPS-Induced ALI
After 24 h of LPS administration, TNF-α and IL-6 in bronchial alveolar lavage fluid (BALF) were assessed to confirm the anti-inflammatory activity of the extract. Compared with the control group, the mice induced with LPS only showed a significant increase in inflammatory markers (TNF-α, and IL-6) (p < 0.05), whereas treatment with the extract (50 and 100 mg/kg) and DEX (5 mg/kg) [28] significantly decreased the levels of inflammatory marker induced by LPS (Figure 8). This finding advocates that Tamarix aphyla has anti-inflammatory effects.
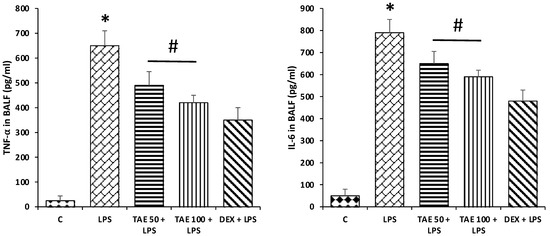
Figure 8.
Effects of extract on inflammatory marker levels. Compared with the control group, the mice induced with LPS only showed a significant increase in inflammatory markers (TNF-α and IL-6) (p < 0.05), whereas treatment with the extract significantly decreases the levels of inflammatory marker induced by LPS. The significant changes (p < 0.05) are shown by (*) between the control group and LPS treatment groupss and (#) between the LPS treatment group and LPS + extract treatment groups.
3.6.4. The Extract Attenuates Lung Oxidative Stress in LPS-Induced Lung Injury in Mice
To evaluate the protective effects of the extract on lung oxidative stress status, the levels of MDA content and SOD level were measured. The SOD level was significantly decreased in the LPS-induced group of mice compared with the control group (p < 0.05) (Figure 9), whereas extract pretreatment markedly reversed SOD enzyme activity in comparison to the group administered LPS only (p < 0.05). Moreover, LPS significantly increased MDA content, while in the extract treatment group, decreased MDA content was noticed (Figure 9).
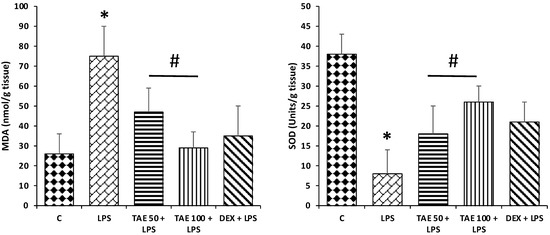
Figure 9.
Effects of extract on oxidative stress. Compared with the control group, the mice induced with LPS only showed a significant increase in MDA content, whereas SOD levels were decreased significantly (p < 0.05). However, treatment with the extract significantly decreased the levels of MDA content induced by LPS and increased the SOD level. The significant changes (p < 0.05) are represented by (*) between the control group and LPS groups, and (#) between the LPS group and LPS + extract groups.
3.7. The Role of the Extract on Wet/Dry Lung Ratios
Lung W/D ratios were evaluated 24 h after LPS administration. LPS-challenged mice showed a significant increase in the lung wet/dry (W/D) weight ratio compared to the control group. Moreover, the extract (1 g/kg) meaningfully reduced the lung W/D ratio increased due to LPS administration (Figure 10). Furthermore, Lung W/D ratios also decreased by DEX (5 mg/kg) treatment after LPS stimulation.
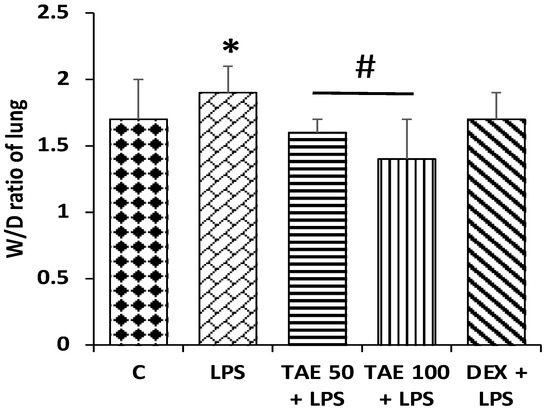
Figure 10.
Different animal groups showing lung W/D ratios. The values denote mean ± SEM, with eight animals in each group. The significant difference (p < 0.05) is shown by an asterisk (*) between the control group and LPS treatment groups, and by (#) between the LPS and LPS + extract groups.
3.7.1. Role of the Extract on Lung Tissue Architectures
LPS, an endotoxin, is a serious factor to inducing a severe, as well as systemic, inflammatory response and acute tissue injury on such organs as the lung, kidney and liver during sepsis [34]. There were no evident histological changes in lung tissues in the control group. After administration of LPS, different types of alteration were noticed in tissues, including congestion, hemorrhage and infiltration of inflammatory cells. However, the LPS-induced lung tissue changes were reduced by pretreatment with the extract (100 mg/kg) (Figure 11). Furthermore, lung alterations were also decreased by DEX (5 mg/kg) treatment after LPS stimulation.
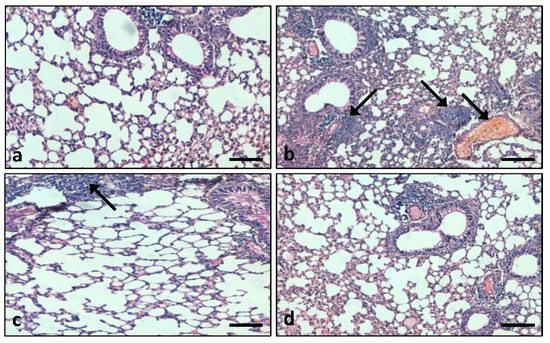
Figure 11.
Lung tissue histopathological analysis in different experimental groups: (a) control animals exhibit normal lung tissue architecture; (b) LPS-induced (5 mg/kg body weight) animals showed severe damage of the alveolar architecture, inflammatory cell infiltration and congestion; (c) the tissue alterations were significantly reduced in the extract treatment group; (d) in the LPS + DEX-treated group, damage of the lung tissue architecture was prevented. (Arrow indicates congestion and inflammation) (Scale bar = 100 μm).
3.7.2. Effects of the Extract on the Expression Pattern of the Cox-2 Protein in Lung Tissue
The immunohistochemistry (IHC) staining result demonstrates that Cox-2 protein expression was increased in the lung tissues of LPS-induced group compared to control group tissues (Cox-2 protein not expressed). In the LPS + extract-treated group, the expression pattern of the Cox-2 protein was low compared to the LPS-treated group only (Figure 12). Moreover, the DEX + LPS group showed mild expression of the Cox-2 protein.
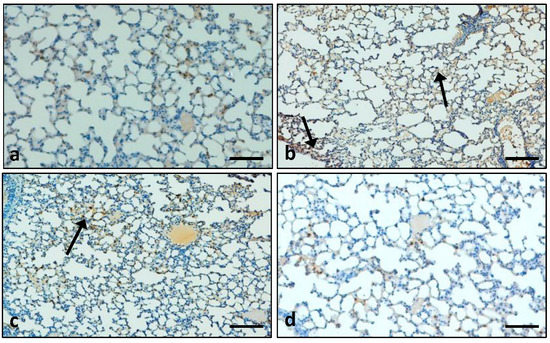
Figure 12.
The expression pattern of Cox-2 was measured. (a) In the control group, Cox-2 protein expression was not detected; (b) In the LPS-treated group, expression pattern of Cox-2 protein was high; (c) In the extract plus LPS treatment group, the Cox-2 protein expression was decreased; (d) In the LPS + DEX treatment group only, there was mild expression of Cox-2 protein. (Arrow indicates cytoplasmic expression) (Scale bar = 100 µm).
3.7.3. Role of the Extract on LPS-Administered Lung Tissue Apoptosis
TUNEL-positive cells were not observed in the control group. When compared with the control group, TUNEL positivity was enhanced in the LPS-induced group significantly. In comparison with the LPS treatment group, pretreatment with the extract also showed TUNEL-positive cells, but the number of TUNEL-positive cells was less compared to the LPS group (Figure 13). However, in the LPS + DEX-treated group, TUNEL-positive cells were decreased significantly (Figure 13).
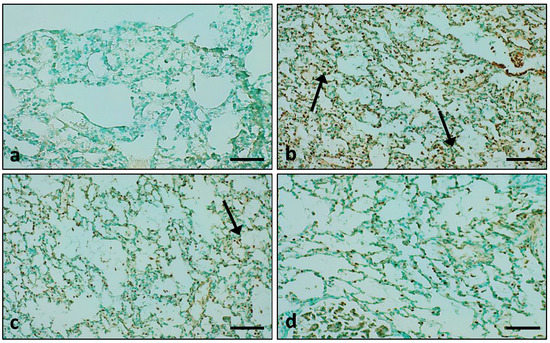
Figure 13.
The effect of the extract was measure on apoptosis. (a) In control animal group, the nuclei stained brown not seen; (b) A significant number was seen in the LPS-treated group only; (c) The group treated with extract plus LPS also showed nuclei stained brown; (d) The group treated with LPS + DEX showed fewer brown-stained nuclei. (Arrow indicates apoptosis) (Scale bar = 100 μm).
4. Discussion
Reactive oxygen species (ROS) are generated in aerobic metabolism processes and are capable of controlling many pathways through direct reaction with proteins, controlling gene expression and transcription factors [35]. An imbalance between the antioxidant production and the production of free radical generation causes various types of pathogenesis. In this perspective, previous studies reported that medicinal plants or its active compounds prevent the pathogenesis through regulation of oxidative stress, inflammation and enhancement of antioxidant levels [11,12,13]. In this study, antioxidant and anti-inflammatory activity of Tamarix aphylla leaf extract were examined, and it was reported that the extract showed free radical scavenging effects through DPPH assay-based findings. These leaves are a rich source of various ingredients, including polyphenols, which are responsible for scavenging the free radical formation. Previous findings were in accordance with the current finding, as T. aphylla leaf and stem extracts were evaluated for antioxidant potential. It was reported that enhanced activity was noticed against DPPH radicals, superoxide anion radicals and nitric oxide radicals [36]. Other findings also reported free radical scavenging and antioxidant potential of Tamarix aphylla bark and A. javanica (aerial parts), respectively [37]. Current findings show that heat-induced albumin denaturation inhibition was concentration-dependent, and the % inhibition of denaturation is enhanced by an increase in extract concentration.
Acute lung injury (ALI) shows high mortality and morbidity among patients and effective drugs are not available in the clinical practice [38,39]. LPS is measured to be one of the vital aspects leading to ALI, which causes entrance of fluid to the lung tissue, as well as microvascular permeability increases, triggering the signs of acute inflammation in the tissue [40,41]. Moreover, it causes inflammatory cell infiltration and alveolar epithelial permeability, which in turn causes in an accumulation of inflammatory cytokines in lung and BALF [42], similar to the pathology of ALI/ARDS in humans [43,44].
After being challenged with LPS, macrophages are activated, and numerous cytokines and pro-inflammatory factors released; too much release can result in a lot of tissue damage and pathological changes [45,46]. The treatment module in the treatment of ALI is still limited. Effective treatments with fewer side effects are still of central importance to health sciences.
Tamarix aphylla, grown in Saudi Arabia, has been shown to have anti-inflammatory and antipyretic properties [47]. In this study, it was reported that treatment of RAW 264.7 macrophages with LPS caused a significant increase in IL-6 and TNF-α. Meanwhile, the extract significantly decreased the LPS-induced production of TNF-α and IL-6 in a concentration-dependent manner. A previous study based on Tamarix aphylla reported that its extracts inhibited the production of TNF-α by LPS-activated THP-1 cells and extracts inhibited TNF-α production with IC50 = 3.7 ± 0.3 and 44.9 ± 8.9 µg/mL, respectively [48]. Cytotoxicity of the extract was evaluated, and it was reported that a low concentration did not show any cytotoxicity and high concentration showed mild cytotoxicity. An earlier study also reported that a mild cytotoxic effect was noticed at high concentrations of T. aphylla leaf extract on the Vero cell line with a 50% cytotoxicity concentration >100 g/mL. Moreover, findings of the cytotoxicity test of extracts on Vero cells designated that this plant is possibly nontoxic [49].
In the current study, it was reported that mice induced with LPSs only showed a significant increase in inflammatory markers (TNF-α and IL-6), whereas treatment with the extract significantly decreased the levels of inflammatory markers induced by LPS. Moreover, the SOD level was significantly decreased in the LPS-induced group of mice compared with the control group, whereas extract pretreatment markedly reversed SOD enzyme activity in comparison to the group administered LPS only. In this regard, an experiment based on leaf extract reported that catalase, SOD and GPX decreased significantly in a CCl4-treated group of mice liver tissues. Moreover, pretreatment with an extract (500 mg/kg) dose resulted in the significant increase in levels of antioxidant enzymes [19]. Moreover, a study based on Portulaca oleracea extract reported that the extract significantly controlled the expression of TNF-α, as well as IL-6, compared with the LPS group [50].
After administration of LPS, inflammatory cell infiltration, congestion and hemorrhage was noticed in lung tissues. However, extract treatment noticeably alleviated the lung tissue changes induced by LPS. Another study was in accordance with the current study, as the study, based on 3,5-dihydroxy-4′,7-dimethoxyflavone isolated from Tamarix aphylla L., reported that this flavone inhibits liver injury [19].
Several proinflammatory mediators such as IL-6 and TNF-α play a significant role in inflammation. Moreover, cyclooxygenase 2 (Cox-2), a vital enzyme, plays a role in inflammation and increased Cox-2 expression as a feature reported to be common to cancers [51] (Shelby et al., 2005). In the current study, it was reported that Cox-2 protein expression was increased in lung tissues of the LPS-induced group compared to the control group. In the LPS + extract-treated group, the expression pattern of the Cox-2 protein was low compared to the LPS-treated group. A previous study was in accordance with the current findings, as Cox-2 was expressed immensely in the LPS model group [52]. Another study based on western blotting reported that Cox-2 expression was significantly higher in the LPS group than in the control group [53]. In this regard, previous findings designated that Cox-2 protein distribution was high after 24 h LPS administration, whereas pretreatment with quercetin significantly prevented the increase in Cox-2 expression [54].
In the current study, it was reported that, when compared with the control group, TUNEL positivity increased in the LPS-treated group significantly. Compared with the LPS group, pretreatment with the extract also showed an increase in the number of TUNEL-positive cells. A previous study was in accordance with the current study, as the number of TUNEL-positive cells was remarkably increased in mice lungs after LPS induction, whereas the number of TUNEL-positive endothelial cells decreased in the lungs of ruscogenin treatment groups [55]. Moreover, increased proportions of TUNEL-positive cells were seen in the LPS group compared with the vehicle group, whereas Capsaicin pretreatment significantly decreased the percentage of TUNEL-positive cells [56].
5. Conclusions
Tamarix aphylla, which holds flavonoids and phenolic compounds, showed potent anti-inflammatory and antioxidant activities through decreasing the IL-6 and TNF-alpha production in RAW 264.7 macrophages treated with LPS, and enhancing the antioxidant levels in the lung BALF. T. aphylla showed an important role in the maintenance of lung tissue architecture and inhibited the protein expression of cox-2. Thus, T. aphylla is a possible herbal medicine that could be developed as a therapeutic anti-inflammatory and antioxidant remedy for the prevention and treatment of lung-associated pathogenesis.
Author Contributions
Conceptualization, S.A.A., A.H.R., M.A.S. and A.A.K.; methodology, S.A.A., M.A.S., A.A.K., A.A.A. and A.H.R.; investigation, S.A.A., A.A.K. and A.H.R.; writing—original draft preparation, S.A.A., A.A.K. and A.H.R.; writing—review and editing, S.A.A., A.A.K., A.A.A. and A.H.R.; supervision, S.A.A., A.A.K. and A.H.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded and supported by the Qassim University, Deanship of Scientific Research (Grant No. 10241-cams1-2020-1-3-I).
Institutional Review Board Statement
The animals were maintained at animal facility of the College of Applied Medical Sciences (CAMS) accordance with the guidelines of the Qassim University on Animal Care. The animal experiments were carried out as per the guidelines of CAMS, Qassim University and approved by the Institutional Animal Ethics Committee (10241-cams1-2020-1-3-I), Qassim University.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data used to support the findings of this study are included within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sarwar, S.; Lin, Z.S.; Ku, C.F.; Guan, Y.F.; Xiao, H.T.; Shi, X.K.; Wang, H.Q.; Bian, Z.X.; Tsang, S.W.; Zhang, H.J. Protective effect of dihydro-resveratrol against lung injury in rats with cerulein-induced acute pancreatitis. Planta Med. 2016, 82, P959. [Google Scholar] [CrossRef]
- Tanaka, R.; Ishima, Y.; Enoki, Y.; Kimachi, K.; Shirai, T.; Watanabe, H.; Chuang, V.T.; Maruyama, T.; Otagiri, M. Therapeutic impact of human serum albumin–thioredoxin fusion protein on influenza virus-induced lung injury mice. Front. Immunol. 2014, 5, 561. [Google Scholar] [CrossRef]
- Sahetya, S.K.; Goligher, E.C.; Brower, R.G. Fifty years of research in ARDS. Setting positive end-expiratory pressure in acute respiratory distress syndrome. Am. J. Respir. Crit. Care Med. 2017, 195, 1429–1438. [Google Scholar] [CrossRef] [PubMed]
- Matthay, M.A.; Ware, L.B.; Zimmerman, G.A. The acute respiratory distress syndrome. J. Clin. Investig. 2012, 122, 2731–2740. [Google Scholar] [CrossRef]
- Hsieh, Y.H.; Deng, J.S.; Pan, H.P.; Liao, J.C.; Huang, S.S.; Huang, G.J. Sclareol ameliorate lipopolysaccharide-induced acute lung injury through inhibition of MAPK and induction of HO-1 signaling. Int. Immunopharmacol. 2017, 44, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Kolomaznik, M.; Nova, Z.; Calkovska, A. Pulmonary surfactant and bacterial lipopolysaccharide: The interaction and its functional consequences. Physiol. Res. 2017, 66, S147–S157. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Sun, M.; Wan, Z.; Liang, J.; Betti, M.; Hrynets, Y.; Xue, X.; Wu, L.; Wang, K. Bee pollen extracts modulate serum metabolism in lipopolysaccharide-induced acute lung injury mice with anti-inflammatory effects. J. Agric. Food Chem. 2019, 67, 7855–7868. [Google Scholar] [CrossRef]
- Ding, H.; Ci, X.; Cheng, H.; Yu, Q.; Li, D. Chicoric acid alleviates lipopolysaccharide-induced acute lung injury in mice through anti-inflammatory and anti-oxidant activities. Int. Immunopharmacol. 2019, 66, 169–176. [Google Scholar] [CrossRef]
- Huang, X.; Liu, Y.; Lu, Y.; Ma, C. Anti-inflammatory effects of eugenol on lipopolysaccharide-induced inflammatory reaction in acute lung injury via regulating inflammation and redox status. Int. Immunopharmacol. 2015, 26, 265–271. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from January 1981 to September 2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Alzohairy, M.A.; Khan, A.A.; Alsahli, M.A.; Almatroodi, S.A.; Rahmani, A.H. Protective effects of thymoquinone, an active compound of nigella sativa, on rats with Benzo (a) pyrene-Induced Lung injury through regulation of oxidative stress and inflammation. Molecules 2021, 26, 3218. [Google Scholar] [CrossRef]
- Alzohairy, M.A.; Khan, A.A.; Ansari, M.A.; Babiker, A.Y.; Alsahli, M.A.; Almatroodi, S.A.; Rahmani, A.H. Protective Effect of Quercetin, a Flavonol against Benzo (a) pyrene-Induced Lung Injury via Inflammation, Oxidative Stress, Angiogenesis and Cyclooxygenase-2 Signalling Molecule. Appl. Sci. 2021, 11, 8675. [Google Scholar] [CrossRef]
- Park, S.M.; Jung, C.J.; Lee, D.G.; Choi, B.R.; Ku, T.H.; La, I.J.; Cho, I.J.; Ku, S.K. Adenophora Stricta Root Extract Protects Lung Injury from Exposure to Particulate Matter 2.5 in Mice. Antioxidants 2022, 11, 1376. [Google Scholar] [CrossRef] [PubMed]
- Mao, M.; Li, J.; Bi, A.; Jia, H.; Li, Q.; Liu, Y.; Jiang, X.; Huang, D.; Xia, S. Thymoquinone ameliorates the PM2.5-induced lung injury in rats. Exp. Lung Res. 2020, 46, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Pei, C.; Wang, F.; Huang, D.; Shi, S.; Wang, X.; Wang, Y.; Li, S.; Wu, Y.; Wang, Z. Astragaloside IV protects from PM2.5-induced lung injury by regulating autophagy via inhibition of PI3K/Akt/mTOR signaling in vivo and in vitro. J. Inflamm. Res. 2021, 14, 4707. [Google Scholar] [CrossRef]
- Panhwar, A.Q.; Abro, H. Ethnobotanical studies of Mahal Kohistan (Khirthar national park). Pak. J. Bot. 2007, 39, 2301–2315. [Google Scholar]
- Ahmad, M.; Zafar, M.; Sultana, S. Salvadora persica, Tamarix aphylla and Zizyphus mauritiana-Three woody plant species mentioned in Holy Quran and Ahadith and their ethnobotanical uses in north western part (DI Khan) of Pakistan. Pak. J. Nutr. 2009, 8, 542–547. [Google Scholar]
- Orabi, M.A.; Yoshimura, M.; Amakura, Y.; Hatano, T. Ellagitannins, gallotannins, and gallo-ellagitannins from the galls of Tamarix aphylla. Fitoterapia 2015, 104, 55–63. [Google Scholar] [CrossRef]
- El-Aarag, B.; Khairy, A.; Khalifa, S.A.M.; El-Seedi, H.R. Protective Effects of Flavone from Tamarix aphylla against CCl4-Induced Liver Injury in Mice Mediated by Suppression of Oxidative Stress, Apoptosis and Angiogenesis. Int. J. Mol. Sci. 2019, 20, 5215. [Google Scholar] [CrossRef]
- Khalid, M.; Hassani, D.; Bilal, M.; Butt, Z.A.; Hamayun, M.; Ahmad, A.; Huang, D.; Hussain, A. Identification of oral cavity biofilm forming bacteria and determination of their growth inhibition by Acacia arabica, Tamarix aphylla L. and Melia azedarach L. medicinal plants. Arch. Oral Biol. 2017, 81, 175–185. [Google Scholar] [CrossRef]
- Qadir, M.I.; Abbas, K.; Hamayun, R.; Ali, M. Analgesic, anti-inflammatory and anti-pyretic activities of aqueous ethanolic extract of Tamarix aphylla L. (Saltcedar) in mice. Pak. J. Pharm. Sci. 2014, 27, 1985–1988. [Google Scholar] [PubMed]
- Soliman Yu, H.; Ibrahim Al, S. Anti-inflammatory and Wound Healing Activities of Herbal Gel Containing an Antioxidant Tamarix aphylla Leaf Extract. Int. J. Pharmacol. 2011, 7, 829–835. [Google Scholar] [CrossRef]
- Hamed, A.I.; Said, R.B.; Kontek, B.; Al-Ayed, A.S.; Kowalczyk, M.; Moldoch, J.; Stochmal, A.; Olas, B. LC–ESI-MS/MS profile of phenolic and glucosinolate compounds in samh flour (Mesembryanthemum forsskalei Hochst. ex Boiss) and the inhibition of oxidative stress by these compounds in human plasma. Food Res. Int. 2016, 85, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Anwar, S.; Raut, R.; Alsahli, M.A.; Almatroudi, A.; Alfheeaid, H.; Alzahrani, F.M.; Khan, A.A.; Allemailem, K.S.; Almatroodi, S.A.; Rahmani, A.H. Role of Ajwa date fruit pulp and seed in the management of diseases through in vitro and in silico analysis. Biology 2022, 11, 78. [Google Scholar] [CrossRef] [PubMed]
- Sakat, S.; Juvekar, A.R.; Gambhire, M.N. In vitro antioxidant and anti-inflammatory activity of methanol extract of Oxalis corniculata Linn. Int. J. Pharm. Pharm. Sci. 2010, 2, 146–155. [Google Scholar]
- Gao, H.; Sun, W.; Zhao, W.; Hao, W.; Leung, C.H.; Lu, J.; Chen, X. Total tanshinones-induced apoptosis and autophagy via reactive oxygen species in lung cancer 95D cells. Am. J. Chin. Med. 2015, 43, 1265–1279. [Google Scholar] [CrossRef]
- Masocha, W. Systemic lipopolysaccharide (LPS)-induced microglial activation results in different temporal reduction of CD200 and CD200 receptor gene expression in the brain. J. Neuroimmunol. 2009, 214, 78–82. [Google Scholar] [CrossRef]
- Huang, C.Y.; Deng, J.S.; Huang, W.C.; Jiang, W.P.; Huang, G.J. Attenuation of Lipopolysaccharide-Induced Acute Lung Injury by Hispolon in Mice, Through Regulating the TLR4/PI3K/Akt/mTOR and Keap1/Nrf2/HO-1 Pathways, and Suppressing Oxidative Stress-Mediated ER Stress-Induced Apoptosis and Autophagy. Nutrients 2020, 12, 1742. [Google Scholar] [CrossRef]
- Ali, H.; Khan, A.; Ali, J.; Ullah, H.; Khan, A.; Ali, H.; Irshad, N.; Khan, S. Attenuation of LPS-induced acute lung injury by continentalic acid in rodents through inhibition of inflammatory mediators correlates with increased Nrf2 protein expression. BMC Pharmacol. Toxicol. 2020, 21, 81. [Google Scholar] [CrossRef]
- Peng, L.Y.; Shi, H.T.; Yuan, M.; Li, J.H.; Song, K.; Huang, J.N.; Yi, P.F.; Shen, H.Q.; Fu, B.D. Madecassoside protects against LPS-induced acute lung injury via inhibiting TLR4/NF-κB activation and blood-air barrier permeability. Front. Pharmacol. 2020, 11, 807. [Google Scholar] [CrossRef]
- Ali, J.; Khan, A.U.; Shah, F.A.; Ali, H.; Islam, S.U.; Kim, Y.S.; Khan, S. Mucoprotective effects of Saikosaponin-A in 5-fluorouracil-induced intestinal mucositis in mice model. Life Sci. 2019, 239, 116888. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.U. Evaluation of in vitro and in vivo therapeutic efficacy of Ribes alpestre Decne in Rheumatoid arthritis. Braz. J. Pharm. Sci. 2019, 55, 1–8. [Google Scholar] [CrossRef]
- Rogler, G.; Andus, T. Cytokines in inflammatory bowel disease. World J. Surg. 1998, 22, 382–389. [Google Scholar] [CrossRef]
- Salomao, R.; Brunialti, M.K.; Rapozo, M.M.; Baggio-Zappia, G.L.; Galanos, C.; Freudenberg, M. Bacterial sensing, cell signaling, and modulation of the immune response during sepsis. Shock 2012, 38, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Sena, L.A.; Chandel, N.S. Physiological roles of mitochondrial reactive oxygen species. Mol. Cell 2012, 48, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Mahfoudhi, A.; Grosso, C.; Gonçalves, R.F.; Khelifi, E.; Hammami, S.; Achour, S.; Trabelsi-Ayadi, M.; Valentão, P.; Andrade, P.B.; Mighri, Z. Evaluation of antioxidant, anticholinesterase, and antidiabetic potential of dry leaves and stems in Tamarix aphylla growing wild in Tunisia. Chem. Biodivers. 2016, 13, 1747–1755. [Google Scholar] [CrossRef]
- Suleiman, M.H. Ethnobotanical, phytochemical, and biological study of Tamarix aphylla and Aerva javanica medicinal plants growing in the Asir region, Saudi Arabia. Trop. Conserv. Sci. 2019, 12, 1940082919869480. [Google Scholar] [CrossRef]
- Butt, Y.; Kurdowska, A.; Allen, T.C. Acute lung injury: A clinical and molecular review. Arch. Pathol. Lab. Med. 2016, 140, 345–350. [Google Scholar] [CrossRef]
- Frutos-Vivar, F.; Nin, N.; Esteban, A. Epidemiology of acute lung injury and acute respiratory distress syndrome. Curr. Opin. Crit. Care 2004, 10, 1–6. [Google Scholar] [CrossRef]
- Ware, L.B.; Matthay, M.A. The acute respiratory distress syndrome. N. Engl. J. Med. 2000, 342, 1334–1349. [Google Scholar] [CrossRef]
- Takeda, K.; Akira, S. Toll-like receptors in innate immunity. Int. Immunol. 2005, 17, 19686699. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Lu, Q.; Wang, K.; Lu, J.; Gu, X.; Zhu, D.; Liu, F.; Guo, Z. miR-34b-5p inhibition attenuates lung inflammation and apoptosis in an LPS-induced acute lung injury mouse model by targeting progranulin. J. Cell. Physiol. 2018, 233, 6615–6631. [Google Scholar] [CrossRef] [PubMed]
- Wyns, H.; Plessers, E.; De Backer, P.; Meyer, E.; Croubels, S. In vivo porcine lipopolysaccharide inflammation models to study immunomodulation of drugs. Vet. Immunol. Immunopathol. 2015, 166, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Salazar, A.; Gonzalez-Rivera, B.L.; Redus, L.; Parrott, J.M.; O’Connor, J.C. Indoleamine 2, 3-dioxygenase mediates anhedonia and anxiety-like behaviors caused by peripheral lipopolysaccharide immune challenge. Horm. Behav. 2012, 62, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Dobrovolskaia, M.A.; Vogel, S.N. Toll receptors, CD14, and macrophage activation and deactivation by LPS. Microbes Infect. 2002, 4, 903–914. [Google Scholar] [CrossRef]
- Maldonado, R.F.; Sá-Correia, I.; Valvano, M.A. Lipopolysaccharide modification in Gram-negative bacteria during chronic infection. FEMS Microbiol. Rev. 2016, 40, 480–493. [Google Scholar] [CrossRef]
- Ali, M.; Alhazmi, H.A.; Ansari, S.H.; Hussain, A.; Ahmad, S.; Alam, M.S.; Ali, M.S.; El-Sharkawy, K.A.; Hakeem, K.R. Tamarix aphylla (L.) Karst. Phytochemical and bioactive profile compilations of less discussed but effective naturally growing Saudi plant. In Plant and Human Health; Springer: Cham, Switzerland, 2019; Volume 3, pp. 343–352. [Google Scholar]
- Gadallah, A.S.; Yousuf, S.; Jabeen, A.; Swilam, M.M.; Khalifa, S.A.; El-Seedi, H.R.; Choudhary, M.I. Anti-inflammatory principles from Tamarix aphylla L.: A bioassay-guided fractionation study. Molecules 2020, 25, 2994. [Google Scholar] [CrossRef]
- Al Sobeai, S.M. Anticancer, cytotoxic effect of Tamarix aphylla, and antibacterial screening efficiency against multidrug-resistant human pathogens. Asian J. Pharm. Clin. Res. 2018, 11, 241–246. [Google Scholar] [CrossRef]
- Baradaran Rahimi, V.; Rakhshandeh, H.; Raucci, F.; Buono, B.; Shirazinia, R.; Samzadeh Kermani, A.; Maione, F.; Mascolo, N.; Askari, V.R. Anti-inflammatory and anti-oxidant activity of Portulaca oleracea extract on LPS-induced rat lung injury. Molecules 2019, 24, 139. [Google Scholar] [CrossRef]
- Shelby, B.D.; Nelson, A.; Morris, C. γ-Herpesvirus neoplasia: A growing role for COX-2. Microsc. Res. Tech. 2005, 68, 120–129. [Google Scholar] [CrossRef]
- Huang, X.L.; Wei, X.C.; Guo, L.Q.; Zhao, L.; Chen, X.H.; Cui, Y.D.; Yuan, J.; Chen, D.F.; Zhang, J. The therapeutic effects of Jaceosidin on lipopolysaccharide-induced acute lung injury in mice. J. Pharmacol. Sci. 2019, 140, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Am Lee, S.; Lee, S.H.; Kim, J.Y.; Lee, W.S. Effects of glycyrrhizin on lipopolysaccharide-induced acute lung injury in a mouse model. J. Thorac. Dis. 2019, 11, 1287. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, J.; Wang, B.; Wu, D.; Li, H.; Lu, H.; Wu, H.; Chai, Y. Protective effect of quercetin on lipopolysaccharide-induced acute lung injury in mice by inhibiting inflammatory cell influx. Exp. Biol. Med. 2014, 239, 1653–1662. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Y.; Gong, S.; Tang, J.; Zhang, J.; Li, F.; Yu, B.; Zhang, Y.; Kou, J. Ruscogenin alleviates LPS-induced pulmonary endothelial cell apoptosis by suppressing TLR4 signaling. Biomed. Pharmacother. 2020, 125, 109868. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Li, N.; Zhan, X.; Zheng, T.; Huang, X.; Chen, Q.; Song, Z.; Yang, F.; Nie, H.; Zhang, Y.; et al. Capsaicin Protects against Lipopolysaccharide-Induced Acute Lung Injury Through the HMGB1/NF-κB and PI3K/AKT/mTOR Pathways. J. Inflamm. Res. 2021, 14, 5291. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).