
Enhancement of Metabolite Production in High-Altitude Microalgal Strains by Optimized C/N/P Ratio


 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Experimental Design
2.3. Quantification of Biomass and Metabolites
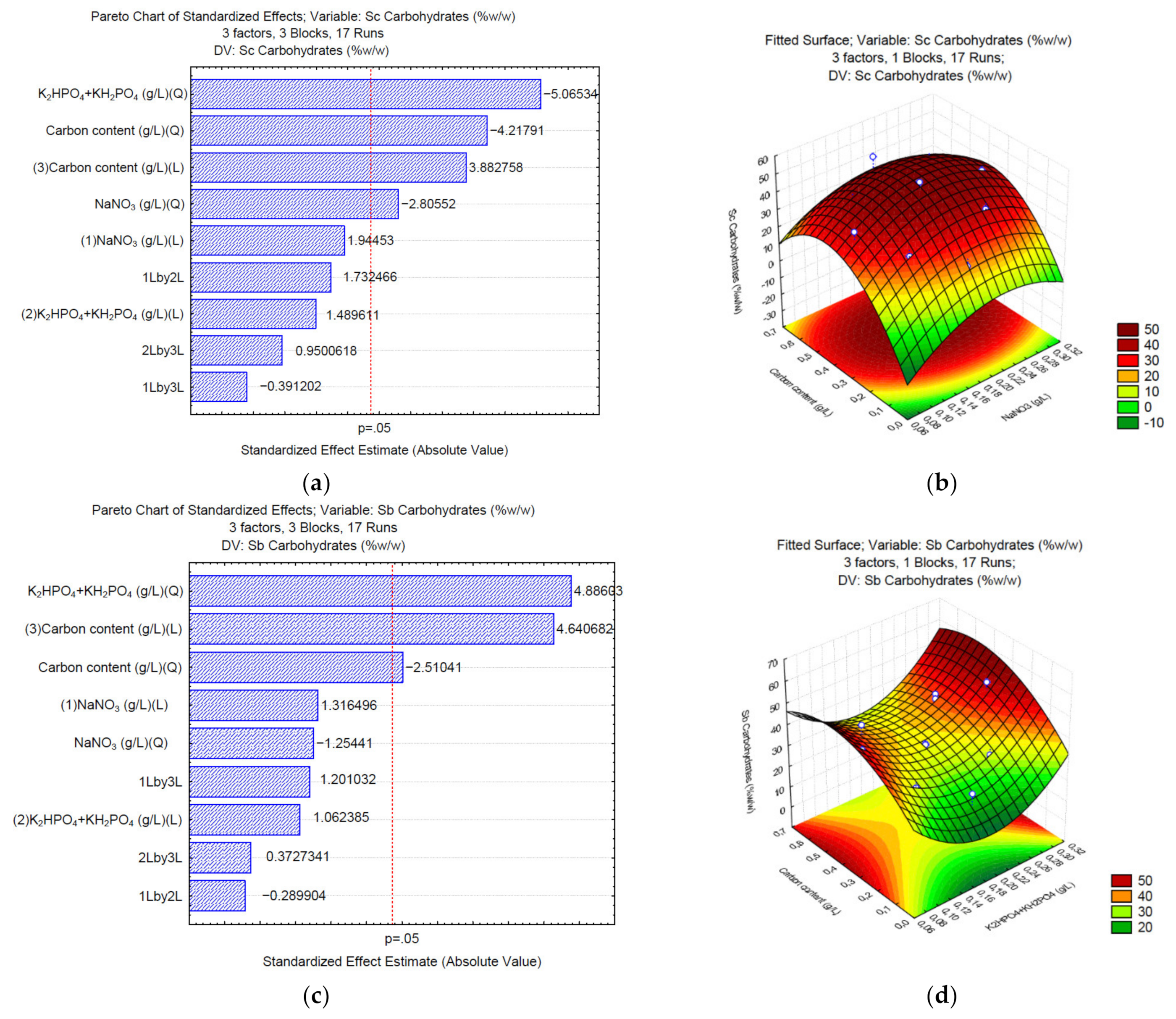
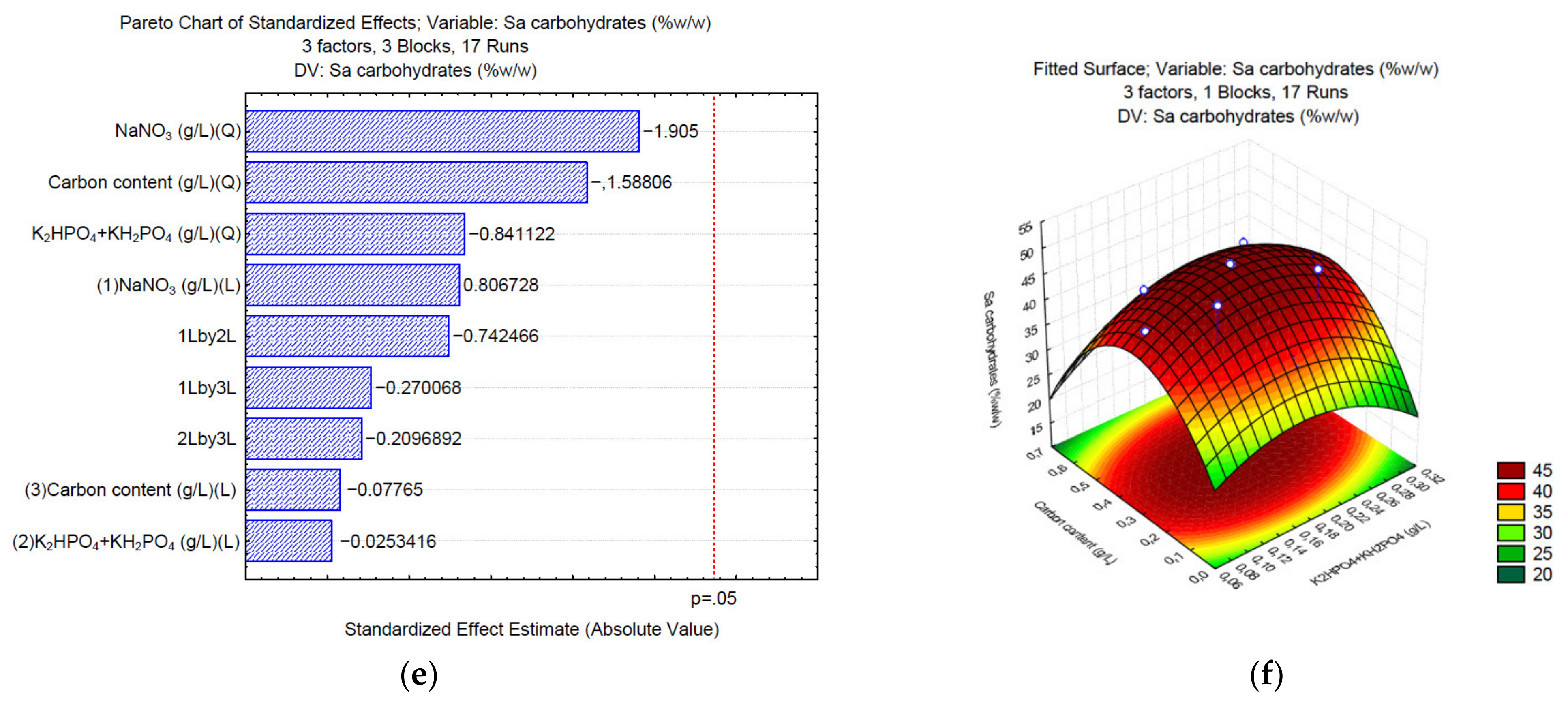
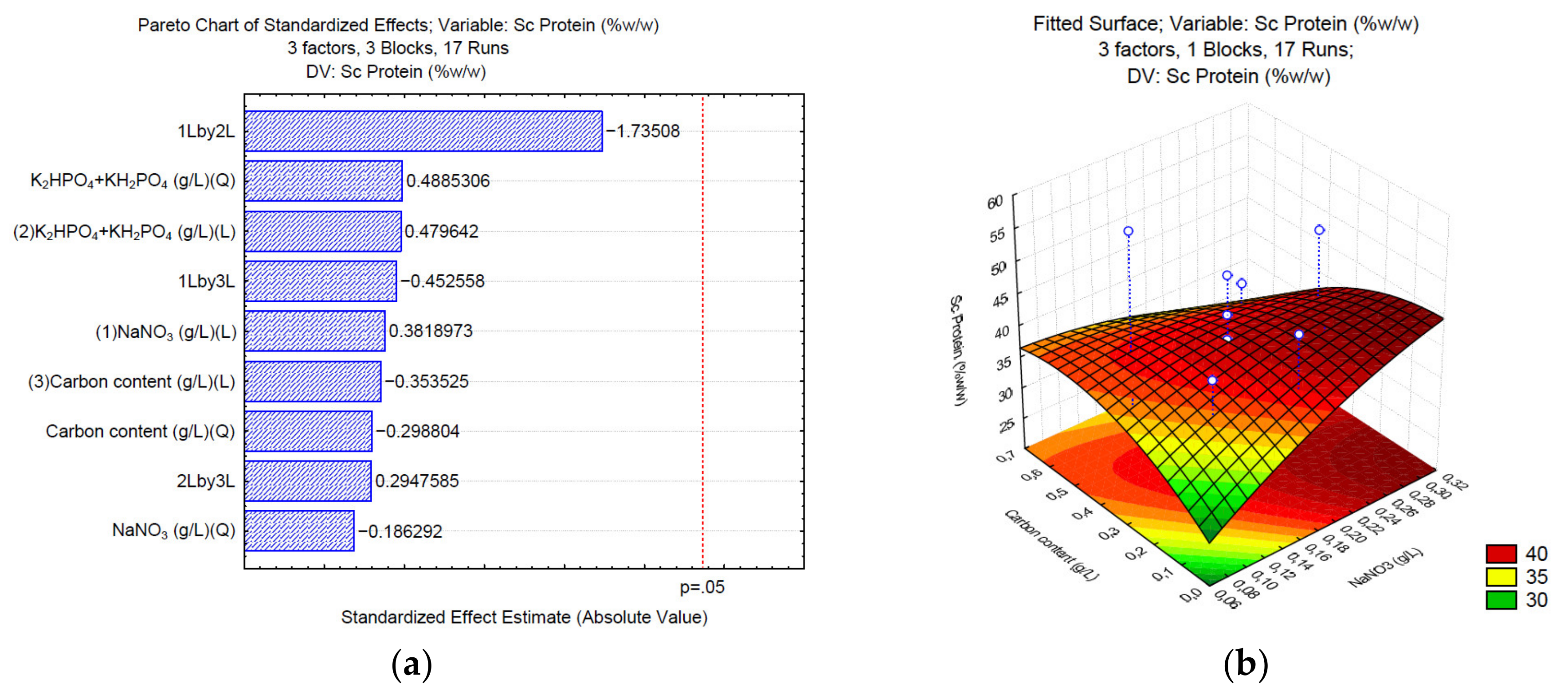
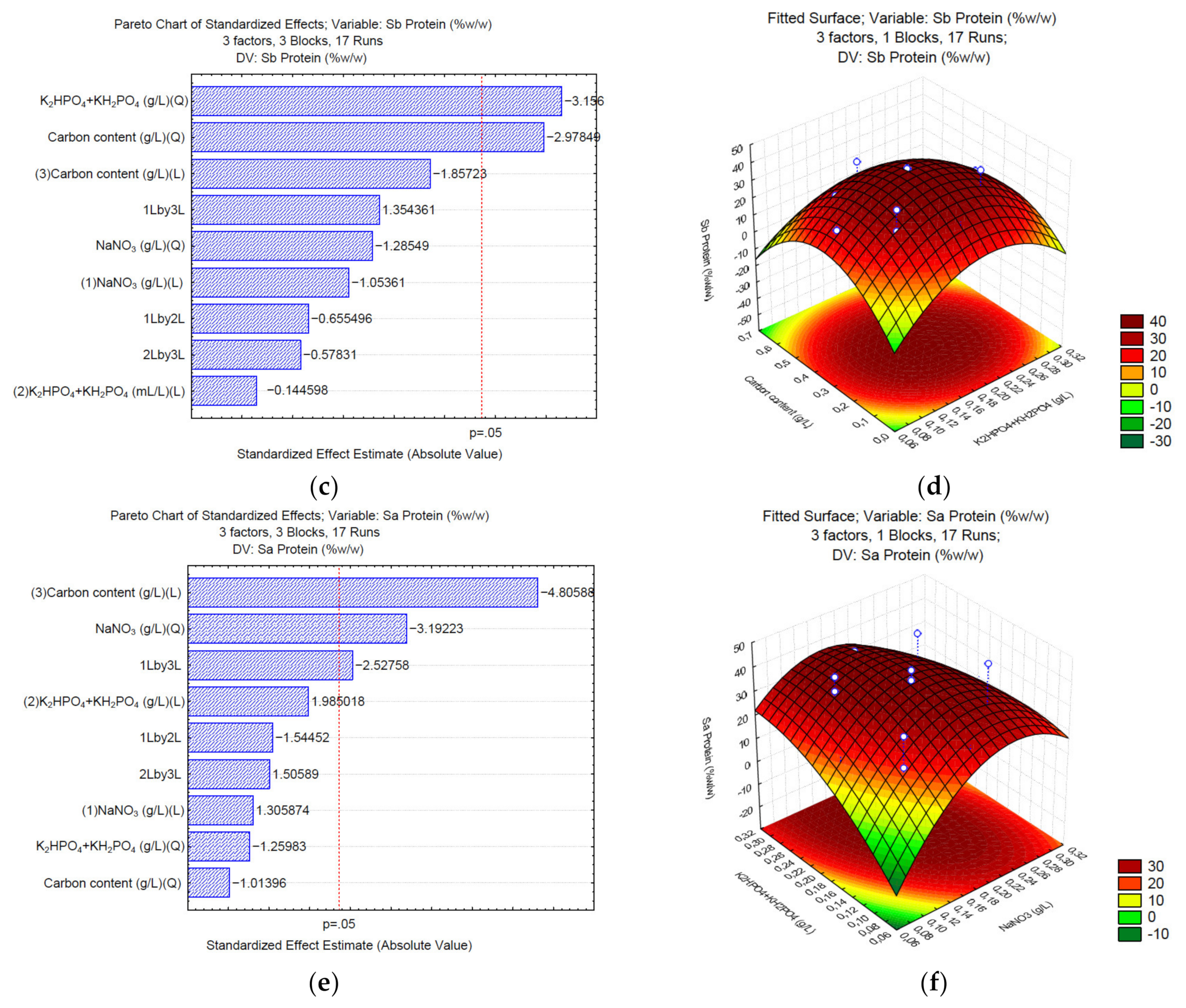
3. Results
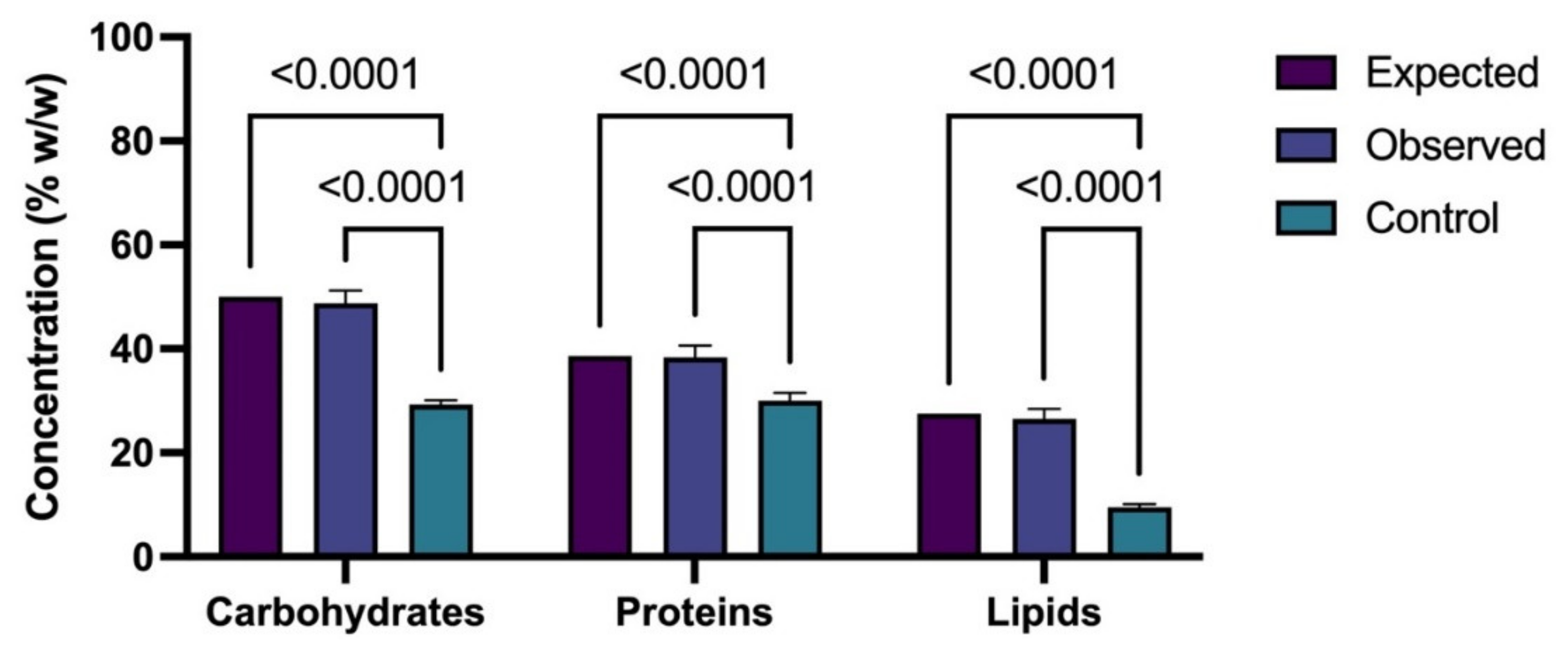
3.1. Production of Carbohydrates under Different C/N/P
3.2. Production of Proteins under Different C/N/P
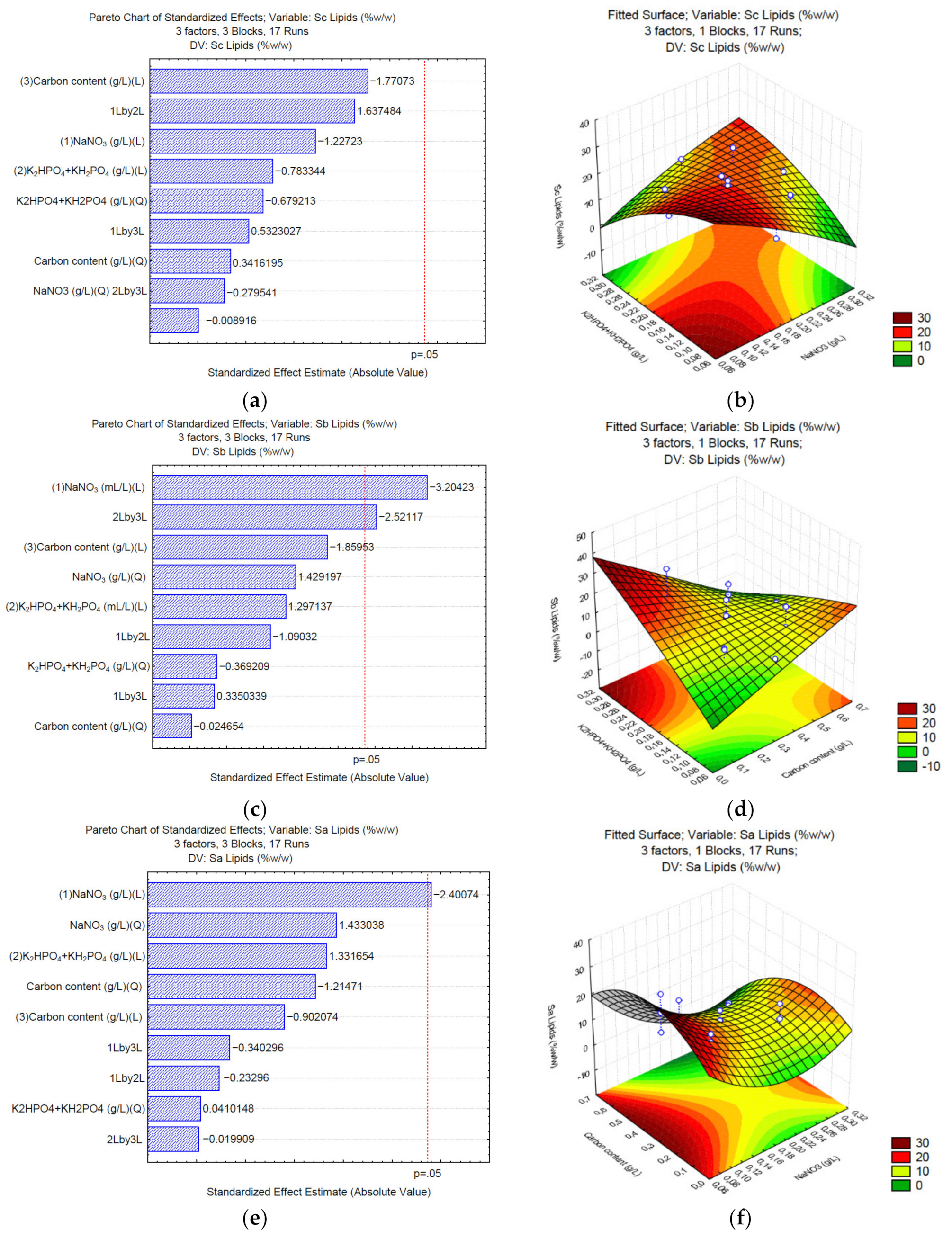
3.3. Production of Lipids under Different C/N/P
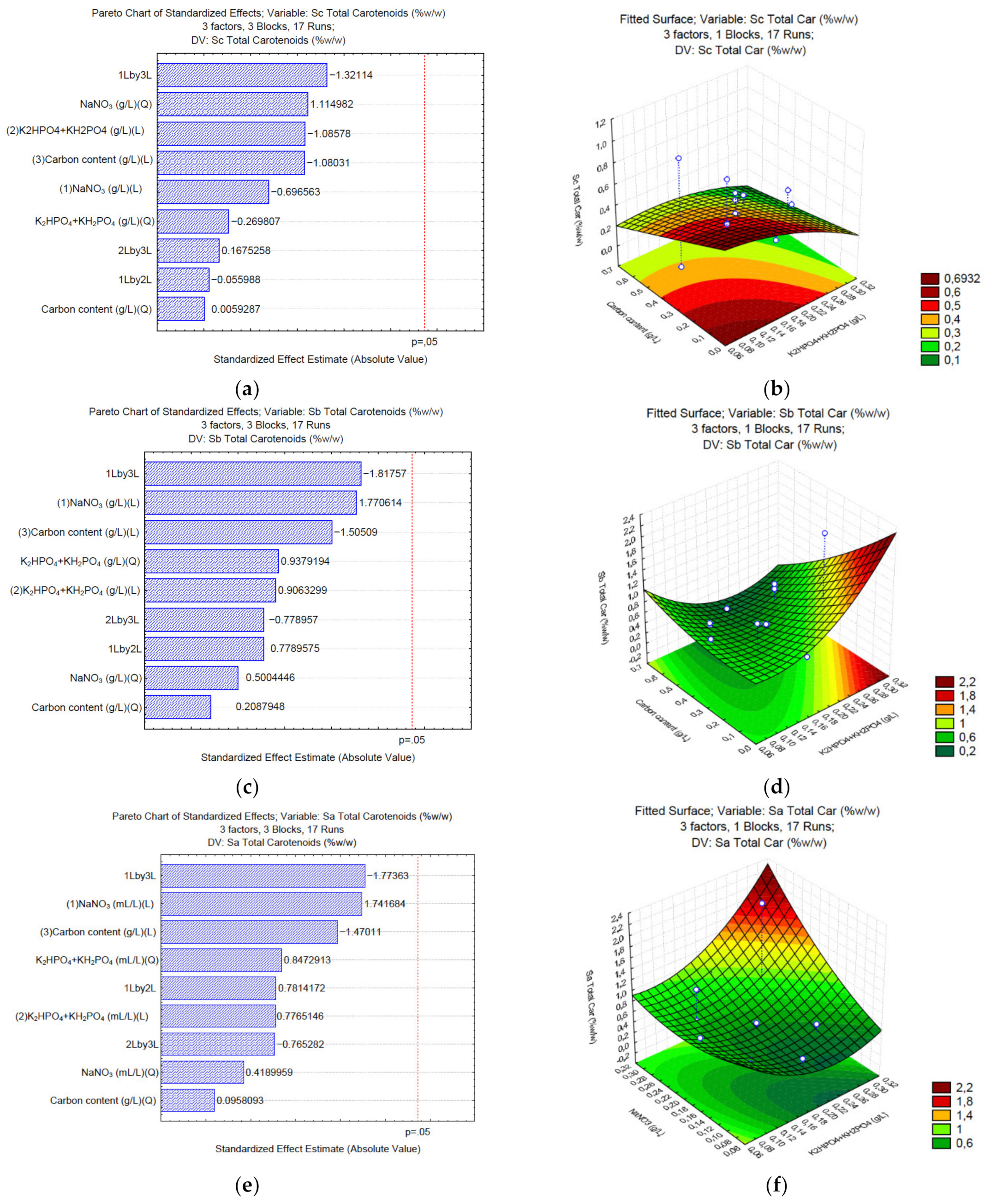
3.4. Production of Total Carotenoids under Different C/N/P
4. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Biomass (g/L) | Productivity (g/L·d−1) | Carbohydrates (% w/w) | Proteins (% w/w) | Lipids (% w/w) | References |
|---|---|---|---|---|---|---|
| Ankistrodesmus falcatus KJ671624 | 1.74 | 0.124 | n/a | 59.6 | [121] | |
| Auxenochlorella protothecoides (formerly Chlorella protothecoides) | 1.06 | 0.71 | n/a | 43.9 | [122] | |
| Auxenochlorella pyrenoidosa (formerly Chlorella pyrenoidosa) | 0.89 | n/a | [98] | |||
| Botryococcus sp. | 0.28 | n/a | 73 | [123] | ||
| B. braunii | 2 | 0.133 | n/a | 70 | [24] | |
| B. braunii LB572 | 4.57 | 0.18 | n/a | 64.9 | [124] | |
| Chlorella sp. | 0.44 | n/a | 55 | [123] | ||
| Chlorella protothecoides UTEX 250 | 1.19 | n/a | 12.9 | [125] | ||
| C. sorokiniana 211-32 | 1.18 | 0.039 | n/a | 38 | [126] | |
| C. sorokiniana UTEX 1602 | 0.68 | n/a | 9 | [127] | ||
| C. sorokiniana UTEX 2805 | 0.66 | n/a | 52 | n/a | [131] | |
| C. vulgaris UTEX 2714 | 0.24 | n/a | 59 | n/a | ||
| C. vulgaris UTEX 1803 | 3.7 | n/a | 60 | n/a | [95] | |
| C. vulgaris FSP-E | 5.51 | 51.3 | [132] | |||
| Chlorococcum oleofaciens | 1.6 | 20 | [97] | |||
| Dunaliella salina | n/a | 0.035 | n/a | 22 | [128] | |
| D. tertiolecta | n/a | 0.044 | n/a | 23 | ||
| D. parva | n/a | 0.045 | n/a | 39 | [129] | |
| Desmodesmus armatus | 1.65 | n/a | 53.6 | n/a | [96] | |
| Scenedesmus sp. ASK22 | 4.67 | 31.06 | 37.1 | [130] | ||
| S. obliquus | n/a | 10–17 | 50–56 | 12–14 | [128] | |
| 2.63 | n/a | 40 | n/a | [132] | ||
| 0.685 | n/a | 39.6 | [93] | |||
| n/a | 66 | [23] | ||||
| S. vacuolatus | 2 | n/a | 4% | [90] | ||
| Chlorella sp. UFPS019 | 0.72 | 0.036 | 48.8 | n/a | This research | |
| 1.5 | 0.048 | n/a | 26.5 | |||
| Desmodesmus sp. UFPS021 | 0.95 | 0.038 | n/a | 38.4 | n/a | |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matos, Â.P. The Impact of Microalgae in Food Science and Technology. JAOCS J. Am. Oil Chem. Soc. 2017, 94, 1333–1350. [Google Scholar] [CrossRef]
- Amini, Z.; Ilham, Z.; Ong, H.C.; Mazaheri, H.; Chen, W.-H. State of the Art and Prospective of Lipase-Catalyzed Transesterification Reaction for Biodiesel Production. Energy Convers. Manag. 2017, 141, 339–353. [Google Scholar] [CrossRef]
- Tekin, K.; Karagöz, S.; Bektaş, S. A Review of Hydrothermal Biomass Processing. Renew. Sustain. Energy Rev. 2014, 40, 673–687. [Google Scholar] [CrossRef]
- Baykara, S.Z. Hydrogen: A Brief Overview on Its Sources, Production and Environmental Impact. Int. J. Hydrgen Energy 2018, 43, 10605–10614. [Google Scholar] [CrossRef]
- Kosourov, S.; Murukesan, G.; Seibert, M.; Allahverdiyeva, Y. Evaluation of Light Energy to H2 Energy Conversion Efficiency in Thin Films of Cyanobacteria and Green Alga under Photoautotrophic Conditions. Algal Res. 2017, 28, 253–263. [Google Scholar] [CrossRef]
- Abbas, J.; Sağsan, M. Impact of Knowledge Management Practices on Green Innovation and Corporate Sustainable Development: A Structural Analysis. J. Clean. Prod. 2019, 229, 611–620. [Google Scholar] [CrossRef]
- Nagarajan, D.; Lee, D.-J.; Kondo, A.; Chang, J.-S. Recent Insights into Biohydrogen Production by Microalgae—From Biophotolysis to Dark Fermentation. Bioresour. Technol. 2017, 227, 373–387. [Google Scholar] [CrossRef]
- Show, P.L.; Tang, M.S.Y.; Nagarajan, D.; Ling, T.C.; Ooi, C.-W.; Chang, J.-S. A Holistic Approach to Managing Microalgae for Biofuel Applications. Int. J. Mol. Sci. 2017, 18, 215. [Google Scholar] [CrossRef]
- Bleakley, S.; Hayes, M. Algal Proteins: Extraction, Application, and Challenges Concerning Production. Foods 2017, 6, 33. [Google Scholar] [CrossRef]
- Koyande, A.K.; Chew, K.W.; Rambabu, K.; Tao, Y.; Chu, D.-T.; Show, P.-L. Microalgae: A Potential Alternative to Health Supplementation for Humans. Food Sci. Hum. Wellness 2019, 8, 16–24. [Google Scholar] [CrossRef]
- Kusmayadi, A.; Leong, Y.K.; Yen, H.-W.; Huang, C.-Y.; Chang, J.-S. Microalgae as Sustainable Food and Feed Sources for Animals and Humans—Biotechnological and Environmental Aspects. Chemosphere 2021, 271, 129800. [Google Scholar] [CrossRef]
- Del Mondo, A.; Smerilli, A.; Sané, E.; Sansone, C.; Brunet, C. Challenging Microalgal Vitamins for Human Health. Microb. Cell Fact. 2020, 19, 201. [Google Scholar] [CrossRef]
- Fu, Y.; Chen, T.; Chen, S.H.Y.; Liu, B.; Sun, P.; Sun, H.; Chen, F. The Potentials and Challenges of Using Microalgae as an Ingredient to Produce Meat Analogues. Trends Food Sci. Technol. 2021, 112, 188–200. [Google Scholar] [CrossRef]
- Kent, M.; Welladsen, H.M.; Mangott, A.; Li, Y. Nutritional Evaluation of Australian Microalgae as Potential Human Health Supplements. PLoS ONE 2015, 10, e0118985. [Google Scholar] [CrossRef]
- Quintero-Dallos, V.; García-Martínez, J.B.; Contreras-Ropero, J.E.; Barajas-Solano, A.F.; Barajas-Ferrerira, C.; Lavecchia, R.; Zuorro, A. Vinasse as a Sustainable Medium for the Production of Chlorella Vulgaris UTEX 1803. Water 2019, 11, 1526. [Google Scholar] [CrossRef]
- da Silva, M.E.T.; Correa, K.d.P.; Martins, M.A.; da Matta, S.L.P.; Martino, H.S.D.; Coimbra, J.S. dos R. Food Safety, Hypolipidemic and Hypoglycemic Activities, and in Vivo Protein Quality of Microalga Scenedesmus Obliquus in Wistar Rats. J. Funct. Foods 2020, 65, 103711. [Google Scholar] [CrossRef]
- Amorim, M.L.; Soares, J.; Vieira, B.B.; Batista-Silva, W.; Martins, M.A. Extraction of Proteins from the Microalga Scenedesmus Obliquus BR003 Followed by Lipid Extraction of the Wet Deproteinized Biomass Using Hexane and Ethyl Acetate. Bioresour. Technol. 2020, 307, 123190. [Google Scholar] [CrossRef]
- Gateau, H.; Blanckaert, V.; Veidl, B.; Burlet-Schiltz, O.; Pichereaux, C.; Gargaros, A.; Marchand, J.; Schoefs, B. Application of Pulsed Electric Fields for the Biocompatible Extraction of Proteins from the Microalga Haematococcus Pluvialis. Bioelectrochemistry 2021, 137, 107588. [Google Scholar] [CrossRef]
- Grossmann, L.; Wörner, V.; Hinrichs, J.; Weiss, J. Mechanism of the Formation of Insoluble Structures in a Protein Extract of the Microalga Chlorella Protothecoides at PH 3. Food Biosci. 2019, 28, 140–142. [Google Scholar] [CrossRef]
- Cheng, D.; Li, D.; Yuan, Y.; Zhou, L.; Li, X.; Wu, T.; Wang, L.; Zhao, Q.; Wei, W.; Sun, Y. Improving Carbohydrate and Starch Accumulation in Chlorella sp. AE10 by a Novel Two-Stage Process with Cell Dilution. Biotechnol. Biofuels 2017, 10, 75. [Google Scholar] [CrossRef]
- Mathimani, T.; Baldinelli, A.; Rajendran, K.; Prabakar, D.; Matheswaran, M.; Pieter van Leeuwen, R.; Pugazhendhi, A. Review on Cultivation and Thermochemical Conversion of Microalgae to Fuels and Chemicals: Process Evaluation and Knowledge Gaps. J. Clean. Prod. 2019, 208, 1053–1064. [Google Scholar] [CrossRef]
- Barajas-Solano, A.F.; Gonzalez-Delgado, A.D.; Kafarov, V. Effect of Thermal Pre-Treatment On Fermentable Sugar Production of Chlorella Vulgaris. Chem. Eng. Trans. 2014, 37, 655–660. [Google Scholar] [CrossRef]
- Cuellar García, D.J.; Rangel-Basto, Y.A.; Barajas-Solano, A.F.; Muñoz-Peñalosa, Y.A.; Urbina-Suarez, N.A. Towards the Production of Microalgae Biofuels: The Effect of the Culture Medium on Lipid Deposition. Biotechnologia 2019, 100, 273–278. [Google Scholar] [CrossRef]
- Barajas-Solano, A.F.; Guzmán-Monsalve, A.; Kafarov, V. Effect of Carbon-Nitrogen Ratio for the Biomass Production, Hydrocarbons and Lipids on Botryoccus Braunii UIS 003. Chem. Eng. Trans. 2016, 49, 247–252. [Google Scholar] [CrossRef]
- Zuorro, A.; Malavasi, V.; Cao, G.; Lavecchia, R. Use of Cell Wall Degrading Enzymes to Improve the Recovery of Lipids from Chlorella Sorokiniana. Chem. Eng. J. 2019, 377, 120325. [Google Scholar] [CrossRef]
- Zuorro, A.; Lavecchia, R.; Maffei, G.; Marra, F.; Miglietta, S.; Petrangeli, A.; Familiari, G.; Valente, T. Enhanced lipid extraction from unbroken microalgal cells using enzymes. Chem. Eng. Trans. 2015, 43, 211–216. [Google Scholar] [CrossRef]
- Chew, K.W.; Yap, J.Y.; Show, P.L.; Suan, N.H.; Juan, J.C.; Ling, T.C.; Lee, D.-J.; Chang, J.-S. Microalgae Biorefinery: High Value Products Perspectives. Bioresour. Technol. 2017, 229, 53–62. [Google Scholar] [CrossRef]
- Kumar, G.; Shobana, S.; Chen, W.-H.; Bach, Q.-V.; Kim, S.-H.; Atabani, A.E.; Chang, J.-S. A Review of Thermochemical Conversion of Microalgal Biomass for Biofuels: Chemistry and Processes. Green Chem. 2017, 19, 44–67. [Google Scholar] [CrossRef]
- Brigljević, B.; Liu, J.; Lim, H. Green Energy from Brown Seaweed: Sustainable Polygeneration Industrial Process via Fast Pyrolysis of S. Japonica Combined with the Brayton Cycle. Energy Convers. Manag. 2019, 195, 1244–1254. [Google Scholar] [CrossRef]
- Xu, X.; Gu, X.; Wang, Z.; Shatner, W.; Wang, Z. Progress, Challenges and Solutions of Research on Photosynthetic Carbon Sequestration Efficiency of Microalgae. Renew. Sustain. Energy Rev. 2019, 110, 65–82. [Google Scholar] [CrossRef]
- Yu, K.L.; Show, P.L.; Ong, H.C.; Ling, T.C.; Chen, W.-H.; Salleh, M.A.M. Biochar Production from Microalgae Cultivation through Pyrolysis as a Sustainable Carbon Sequestration and Biorefinery Approach. Clean Technol. Environ. Policy 2018, 20, 2047–2055. [Google Scholar] [CrossRef]
- Zuorro, A.; García-Martínez, J.B.; Barajas-Solano, A.F. The Application of Catalytic Processes on the Production of Algae-Based Biofuels: A Review. Catalysts 2021, 11, 22. [Google Scholar] [CrossRef]
- Mohamed, A.G.; Abo-El-Khair, B.E.; Shalaby, S.M. Quality of novel healthy processed cheese analogue enhanced with marine microalgae Chlorella vulgaris biomass. World Appl. Sci. J. 2013, 23, 914–925. [Google Scholar]
- Furbeyre, H.; van Milgen, J.; Mener, T.; Gloaguen, M.; Labussière, E. Effects of Dietary Supplementation with Freshwater Microalgae on Growth Performance, Nutrient Digestibility and Gut Health in Weaned Piglets. Animal 2017, 11, 183–192. [Google Scholar] [CrossRef]
- Oh, S.T.; Zheng, L.; Kwon, H.J.; Choo, Y.K.; Lee, K.W.; Kang, C.W.; An, B.K. Effects of Dietary Fermented Chlorella Vulgaris (CBT®) on Growth Performance, Relative Organ Weights, Cecal Microflora, Tibia Bone Characteristics, and Meat Qualities in Pekin Ducks. Asian-Australas. J. Anim. Sci. 2015, 28, 95–101. [Google Scholar] [CrossRef]
- Kang, H.K.; Salim, H.M.; Akter, N.; Kim, D.W.; Kim, J.H.; Bang, H.T.; Kim, M.J.; Na, J.C.; Hwangbo, J.; Choi, H.C.; et al. Effect of Various Forms of Dietary Chlorella Supplementation on Growth Performance, Immune Characteristics, and Intestinal Microflora Population of Broiler Chickens. J. Appl. Poult. Res. 2013, 22, 100–108. [Google Scholar] [CrossRef]
- El-Baz, F.K.; Abdo, S.M.; Hussein, A.M.S. Microalgae Dunaliella salina for use as food supplement to improve pasta quality. Int. J. Pharm. Sci. Rev. Res. 2017, 46, 45–51. [Google Scholar]
- Xu, Y.; Ibrahim, I.M.; Wosu, C.I.; Ben-Amotz, A.; Harvey, P.J. Potential of New Isolates of Dunaliella Salina for Natural β-Carotene Production. Biology 2018, 7, 14. [Google Scholar] [CrossRef]
- Sui, Y.; Mazzucchi, L.; Acharya, P.; Xu, Y.; Morgan, G.; Harvey, P.J. A Comparison of β-Carotene, Phytoene and Amino Acids Production in Dunaliella salina DF 15 (CCAP 19/41) and Dunaliella salina CCAP 19/30 Using Different Light Wavelengths. Foods 2021, 10, 2824. [Google Scholar] [CrossRef]
- Chan, K.; Chen, S.; Chen, P. Astaxanthin Attenuated Thrombotic Risk Factors in Type 2 Diabetic Patients. J. Funct. Foods 2019, 53, 22–27. [Google Scholar] [CrossRef]
- Sheikhzadeh, N.; Tayefi-Nasrabadi, H.; Khani Oushani, A.; Najafi Enferadi, M.H. Effects of Haematococcus Pluvialis Supplementation on Antioxidant System and Metabolism in Rainbow Trout (Oncorhynchus Mykiss). Fish Physiol. Biochem. 2012, 38, 413–419. [Google Scholar] [CrossRef]
- Do, T.-T.; Ong, B.-N.; Le, T.-L.; Nguyen, T.-C.; Tran-Thi, B.-H.; Thu Hien, B.T.; Melkonian, M.; Tran, H.-D. Growth of Haematococcus pluvialis on a Small-Scale Angled Porous Substrate Photobioreactor for Green Stage Biomass. Appl. Sci. 2021, 11, 1788. [Google Scholar] [CrossRef]
- Rizzo, A.; Ross, M.E.; Norici, A.; Jesus, B. A Two-Step Process for Improved Biomass Production and Non-Destructive Astaxanthin and Carotenoids Accumulation in Haematococcus pluvialis. Appl. Sci. 2022, 12, 1261. [Google Scholar] [CrossRef]
- Neves, M.; Ferreira, A.; Antunes, M.; Laranjeira Silva, J.; Mendes, S.; Gil, M.M.; Tecelão, C. Nannochloropsis oceanica as a Sustainable Source of n-3 Polyunsaturated Fatty Acids for Enrichment of Hen Eggs. Appl. Sci. 2021, 11, 8747. [Google Scholar] [CrossRef]
- du Preez, R.; Majzoub, M.E.; Thomas, T.; Panchal, S.K.; Brown, L. Nannochloropsis oceanica as a Microalgal Food Intervention in Diet-Induced Metabolic Syndrome in Rats. Nutrients 2021, 13, 3991. [Google Scholar] [CrossRef]
- Saito, T.; Ichihara, T.; Inoue, H.; Uematsu, T.; Hamada, S.; Watanabe, T.; Takimura, Y.; Webb, J. Comparison of Areal Productivity of Nannochloropsis Oceanica Between Lab-Scale and Industrial-Scale Raceway Pond. Mar. Biotechnol. 2020, 22, 836–841. [Google Scholar] [CrossRef]
- Zhang, R.; Parniakov, O.; Grimi, N.; Lebovka, N.; Marchal, L.; Vorobiev, E. Emerging Techniques for Cell Disruption and Extraction of Valuable Bio-Molecules of Microalgae Nannochloropsis sp. Bioprocess Biosyst. Eng. 2019, 42, 173–186. [Google Scholar] [CrossRef]
- Mitra, M.; Mishra, S. A Biorefinery from Nannochloropsis spp. Utilizing Wastewater Resources BT—Application of Microalgae in Wastewater Treatment: Volume 2: Biorefinery Approaches of Wastewater Treatment; Gupta, S.K., Bux, F., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 123–145. [Google Scholar] [CrossRef]
- Branco-Vieira, M.; San Martin, S.; Agurto, C.; Santos, M.A.d.; Freitas, M.A.V.; Mata, T.M.; Martins, A.A.; Caetano, N.S. Potential of Phaeodactylum tricornutum for Biodiesel Production under Natural Conditions in Chile. Energies 2018, 11, 54. [Google Scholar] [CrossRef]
- Afonso, C.; Bragança, A.R.; Rebelo, B.A.; Serra, T.S.; Abranches, R. Optimal Nitrate Supplementation in Phaeodactylum tricornutum Culture Medium Increases Biomass and Fucoxanthin Production. Foods 2022, 11, 568. [Google Scholar] [CrossRef]
- Neumann, U.; Derwenskus, F.; Flaiz Flister, V.; Schmid-Staiger, U.; Hirth, T.; Bischoff, S.C. Fucoxanthin, A Carotenoid Derived from Phaeodactylum tricornutum Exerts Antiproliferative and Antioxidant Activities In Vitro. Antioxidants 2019, 8, 183. [Google Scholar] [CrossRef]
- Eilers, U.; Bikoulis, A.; Breitenbach, J.; Büchel, C.; Sandmann, G. Limitations in the Biosynthesis of Fucoxanthin as Targets for Genetic Engineering in Phaeodactylum Tricornutum. J. Appl. Phycol. 2016, 28, 123–129. [Google Scholar] [CrossRef]
- Kwon, D.Y.; Vuong, T.T.; Choi, J.; Lee, T.S.; Um, J.-I.; Koo, S.Y.; Hwang, K.T.; Kim, S.M. Fucoxanthin Biosynthesis Has a Positive Correlation with the Specific Growth Rate in the Culture of Microalga Phaeodactylum Tricornutum. J. Appl. Phycol. 2021, 33, 1473–1485. [Google Scholar] [CrossRef]
- Li, Y.; Ma, Q.; Pan, Y.; Chen, Q.; Sun, Z.; Hu, P. Development of an Effective Flocculation Method by Utilizing the Auto-Flocculation Capability of Phaeodactylum Tricornutum. Algal Res. 2021, 58, 102413. [Google Scholar] [CrossRef]
- Pereira, H.; Sá, M.; Maia, I.; Rodrigues, A.; Teles, I.; Wijffels, R.H.; Navalho, J.; Barbosa, M. Fucoxanthin Production from Tisochrysis Lutea and Phaeodactylum Tricornutum at Industrial Scale. Algal Res. 2021, 56, 102322. [Google Scholar] [CrossRef]
- Kadalag, N.L.; Pawar, P.R.; Prakash, G. Co-Cultivation of Phaeodactylum Tricornutum and Aurantiochytrium Limacinum for Polyunsaturated Omega-3 Fatty Acids Production. Bioresour. Technol. 2022, 346, 126544. [Google Scholar] [CrossRef]
- Han, S.-I.; Jeon, M.S.; Park, Y.H.; Kim, S.; Choi, Y.-E. Semi-Continuous Immobilized Cultivation of Porphyridium Cruentum for Sulfated Polysaccharides Production. Bioresour. Technol. 2021, 341, 125816. [Google Scholar] [CrossRef]
- Medina-Cabrera, E.V.; Gansbiller, M.; Rühmann, B.; Schmid, J.; Sieber, V. Rheological Characterization of Porphyridium Sordidum and Porphyridium Purpureum Exopolysaccharides. Carbohydr. Polym. 2021, 253, 117237. [Google Scholar] [CrossRef]
- Seemashree, M.H.; Chauhan, V.S.; Sarada, R. Phytohormone Supplementation Mediated Enhanced Biomass Production, Lipid Accumulation, and Modulation of Fatty Acid Profile in Porphyridium Purpureum and Dunaliella Salina Cultures. Biocatal. Agric. Biotechnol. 2022, 39, 102253. [Google Scholar] [CrossRef]
- da Silva, M.E.T.; Leal, M.A.; Resende, M.d.O.; Martins, M.A.; Coimbra, J.S.d.R. Scenedesmus Obliquus Protein Concentrate: A Sustainable Alternative Emulsifier for the Food Industry. Algal Res. 2021, 59, 102468. [Google Scholar] [CrossRef]
- Tao, R.; Lakaniemi, A.-M.; Rintala, J.A. Cultivation of Scenedesmus Acuminatus in Different Liquid Digestates from Anaerobic Digestion of Pulp and Paper Industry Biosludge. Bioresour. Technol. 2017, 245, 706–713. [Google Scholar] [CrossRef]
- Singh, D.V.; Upadhyay, A.K.; Singh, R.; Singh, D.P. Implication of Municipal Wastewater on Growth Kinetics, Biochemical Profile, and Defense System of Chlorella Vulgaris and Scenedesmus Vacuolatus. Environ. Technol. Innov. 2022, 26, 102334. [Google Scholar] [CrossRef]
- Tavanandi, H.A.; Raghavarao, K.S.M.S. Ultrasound-Assisted Enzymatic Extraction of Natural Food Colorant C-Phycocyanin from Dry Biomass of Arthrospira Platensis. LWT 2020, 118, 108802. [Google Scholar] [CrossRef]
- İlter, I.; Akyıl, S.; Demirel, Z.; Koç, M.; Conk-Dalay, M.; Kaymak-Ertekin, F. Optimization of Phycocyanin Extraction from Spirulina Platensis Using Different Techniques. J. Food Compos. Anal. 2018, 70, 78–88. [Google Scholar] [CrossRef]
- Ayekpam, C.; Hamsavi, G.K.; Raghavarao, K.S.M.S. Efficient Extraction of Food Grade Natural Blue Colorant from Dry Biomass of Spirulina Platensis Using Eco-Friendly Methods. Food Bioprod. Process. 2021, 129, 84–93. [Google Scholar] [CrossRef]
- Ferreira-Santos, P.; Nunes, R.; De Biasio, F.; Spigno, G.; Gorgoglione, D.; Teixeira, J.A.; Rocha, C.M.R. Influence of Thermal and Electrical Effects of Ohmic Heating on C-Phycocyanin Properties and Biocompounds Recovery from Spirulina Platensis. LWT 2020, 128, 109491. [Google Scholar] [CrossRef]
- Tavanandi, H.A.; Mittal, R.; Chandrasekhar, J.; Raghavarao, K.S.M.S. Simple and Efficient Method for Extraction of C-Phycocyanin from Dry Biomass of Arthospira Platensis. Algal Res. 2018, 31, 239–251. [Google Scholar] [CrossRef]
- Devi, A.C.; Tavanandi, H.A.; Govindaraju, K.; Raghavarao, K.S.M.S. An Effective Method for Extraction of High Purity Phycocyanins (C-PC and A-PC) from Dry Biomass of Arthrospira Maxima. J. Appl. Phycol. 2020, 32, 1141–1151. [Google Scholar] [CrossRef]
- Carullo, D.; Pataro, G.; Donsì, F.; Ferrari, G. Pulsed Electric Fields-Assisted Extraction of Valuable Compounds from Arthrospira Platensis: Effect of Pulse Polarity and Mild Heating. Front. Bioeng. Biotechnol. 2020, 8, 1–15. [Google Scholar] [CrossRef]
- Khandual, S.; Sanchez, E.O.L.; Andrews, H.E.; de la Rosa, J.D.P. Phycocyanin Content and Nutritional Profile of Arthrospira Platensis from Mexico: Efficient Extraction Process and Stability Evaluation of Phycocyanin. BMC Chem. 2021, 15, 1–13. [Google Scholar] [CrossRef]
- del Pilar Sánchez-Saavedra, M.; Maeda-Martínez, A.N.; Acosta-Galindo, S. Effect of Different Light Spectra on the Growth and Biochemical Composition of Tisochrysis Lutea. J. Appl. Phycol. 2016, 28, 839–847. [Google Scholar] [CrossRef]
- Matsui, H.; Intoy, M.M.B.; Waqalevu, V.; Ishikawa, M.; Kotani, T. Suitability of Tisochrysis Lutea at Different Growth Phases as an Enrichment Diet for Brachionus Plicatilis sp. Complex Rotifers. J. Appl. Phycol. 2020, 32, 3933–3947. [Google Scholar] [CrossRef]
- Marchetti, J.; da Costa, F.; Bougaran, G.; Quéré, C.; Soudant, P.; Robert, R. The Combined Effects of Blue Light and Dilution Rate on Lipid Class and Fatty Acid Composition of Tisochrysis Lutea. J. Appl. Phycol. 2018, 30, 1483–1494. [Google Scholar] [CrossRef]
- Gao, F.; Cabanelas, I.T.D.; Wijffels, R.H.; Barbosa, M.J. Fucoxanthin and Docosahexaenoic Acid Production by Cold-Adapted Tisochrysis Lutea. New Biotechnol. 2022, 66, 16–24. [Google Scholar] [CrossRef]
- Mohamadnia, S.; Tavakoli, O.; Faramarzi, M.A. Production of Fucoxanthin from the Microalga Tisochrysis Lutea in the Bubble Column Photobioreactor Applying Mass Transfer Coefficient. J. Biotechnol. 2022, 348, 47–54. [Google Scholar] [CrossRef]
- Hernández-López, I.; Benavente Valdés, J.R.; Castellari, M.; Aguiló-Aguayo, I.; Morillas-España, A.; Sánchez-Zurano, A.; Acién-Fernández, F.G.; Lafarga, T. Utilisation of the Marine Microalgae Nannochloropsis sp. and Tetraselmis sp. as Innovative Ingredients in the Formulation of Wheat Tortillas. Algal Res. 2021, 58, 102361. [Google Scholar] [CrossRef]
- Magpusao, J.; Giteru, S.; Oey, I.; Kebede, B. Effect of High Pressure Homogenization on Microstructural and Rheological Properties of A. Platensis, Isochrysis, Nannochloropsis and Tetraselmis Species. Algal Res. 2021, 56, 102327. [Google Scholar] [CrossRef]
- Khatoon, H.; Penz, K.R.; Banerjee, S.; Rahman, M.R.; Minhaz, T.M.; Islam, Z.; Mukta, F.A.; Nayma, Z.; Sultana, R.; Amira, K.I. Immobilized Tetraselmis sp. for Reducing Nitrogenous and Phosphorous Compounds from Aquaculture Wastewater. Bioresour. Technol. 2021, 338, 125529. [Google Scholar] [CrossRef]
- Farahin, A.W.; Natrah, I.; Nagao, N.; Katayama, T.; Imaizumi, Y.; Mamat, N.Z.; Yusoff, F.M.; Shariff, M. High Intensity of Light: A Potential Stimulus for Maximizing Biomass by Inducing Photosynthetic Activity in Marine Microalga, Tetraselmis Tetrathele. Algal Res. 2021, 60, 102523. [Google Scholar] [CrossRef]
- Souto, M.; Saavedra, M.; Pousão-Ferreira, P.; Herrero, C. Riboflavin Enrichment throughout the Food Chain from the Marine Microalga Tetraselmis Suecica to the Rotifer Brachionus Plicatilis and to White Sea Bream (Diplodus Sargus) and Gilthead Sea Bream (Sparus Aurata) Larvae. Aquaculture 2008, 283, 128–133. [Google Scholar] [CrossRef]
- Araújo, R.; Vázquez Calderón, F.; Sánchez López, J.; Azevedo, I.C.; Bruhn, A.; Fluch, S.; Garcia Tasende, M.; Ghaderiardakani, F.; Ilmjärv, T.; Laurans, M.; et al. Current Status of the Algae Production Industry in Europe: An Emerging Sector of the Blue Bioeconomy. Front. Mar. Sci. 2021, 7, 626389. [Google Scholar] [CrossRef]
- Nascimento, I.A.; Marques, S.S.I.; Cabanelas, I.T.D.; de Carvalho, G.C.; Nascimento, M.A.; de Souza, C.O.; Druzian, J.I.; Hussain, J.; Liao, W. Microalgae Versus Land Crops as Feedstock for Biodiesel: Productivity, Quality, and Standard Compliance. BioEnergy Res. 2014, 7, 1002–1013. [Google Scholar] [CrossRef]
- Neofotis, P.; Huang, A.; Sury, K.; Chang, W.; Joseph, F.; Gabr, A.; Twary, S.; Qiu, W.; Holguin, O.; Polle, J.E.W. Characterization and Classification of Highly Productive Microalgae Strains Discovered for Biofuel and Bioproduct Generation. Algal Res. 2016, 15, 164–178. [Google Scholar] [CrossRef]
- Hobbs, W.O.; Telford, R.J.; Birks, H.J.B.; Saros, J.E.; Hazewinkel, R.R.O.; Perren, B.B.; Saulnier-Talbot, É.; Wolfe, A.P. Quantifying Recent Ecological Changes in Remote Lakes of North America and Greenland Using Sediment Diatom Assemblages. PLoS ONE 2010, 5, e10026. [Google Scholar] [CrossRef] [PubMed]
- Dunck, B.; Felisberto, S.A.; de Souza Nogueira, I. Effects of Freshwater Eutrophication on Species and Functional Beta Diversity of Periphytic Algae. Hydrobiologia 2019, 837, 195–204. [Google Scholar] [CrossRef]
- Schuster, K.F.; Tremarin, P.I.; de Souza-Franco, G.M. Alpha and beta diversity of phytoplankton in two subtropical eutrophic streams in southern Brazil. Acta Bot. Bras. 2015, 29, 597–607. [Google Scholar] [CrossRef][Green Version]
- Gunkel, G.; Casallas, J. Limnology of an Equatorial High Mountain Lake—Lago San Pablo, Ecuador: The Significance of Deep Diurnal Mixing for Lake Productivity. Limnologica 2002, 32, 33–43. [Google Scholar] [CrossRef]
- Zuorro, A.; Leal-Jerez, A.G.; Morales-Rivas, L.K.; Mogollón-Londoño, S.O.; Sanchez-Galvis, E.M.; García-Martínez, J.B.; Barajas-Solano, A.F. Enhancement of Phycobiliprotein Accumulation in Thermotolerant Oscillatoria sp. through Media Optimization. ACS Omega 2021, 6, 10527–10536. [Google Scholar] [CrossRef]
- Sánchez-Zurano, A.; Morillas-España, A.; Gómez-Serrano, C.; Ciardi, M.; Acién, G.; Lafarga, T. Annual Assessment of the Wastewater Treatment Capacity of the Microalga Scenedesmus Almeriensis and Optimisation of Operational Conditions. Sci. Rep. 2021, 11, 21651. [Google Scholar] [CrossRef]
- Lv, J.-M.; Cheng, L.-H.; Xu, X.-H.; Zhang, L.; Chen, H.-L. Enhanced Lipid Production of Chlorella Vulgaris by Adjustment of Cultivation Conditions. Bioresour. Technol. 2010, 101, 6797–6804. [Google Scholar] [CrossRef]
- Mandik, Y.I.; Cheirsilp, B.; Boonsawang, P.; Prasertsan, P. Optimization of Flocculation Efficiency of Lipid-Rich Marine Chlorella sp. Biomass and Evaluation of Its Composition in Different Cultivation Modes. Bioresour. Technol. 2015, 182, 89–97. [Google Scholar] [CrossRef]
- Cuéllar-García, D.J.; Rangel-Basto, Y.A.; Urbina-Suarez, N.A.; Barajas-Solano, A.F.; Muñoz-Peñaloza, Y.A. Lipids production from Scenedesmus obliquus through carbon/nitrogen ratio optimization. J. Phys. Conf. Ser. 2019, 1388, 012043. [Google Scholar] [CrossRef]
- Barajas-Solano, A.F. Optimization of Phycobiliprotein Solubilization from a Thermotolerant Oscillatoria sp. Processes 2022, 10, 836. [Google Scholar] [CrossRef]
- González-Delgado, A.D.; Barajas-Solano, A.F.; Ardila-Álvarez, A.M. Biomass and Protein Production of Chlorella vulgaris Beyerinck (Chlorellales: Chlorellaceae) via the Design of Selective Culture Media. Corpoica Cienc. Tecnol. Agropecu. 2017, 18, 451–461. [Google Scholar] [CrossRef]
- Wang, S.; Cao, M.; Wang, B.; Deng, R.; Gao, Y.; Liu, P. Optimization of growth requirements and scale-up cultivation of freshwater algae Desmodesmus armatus using response surface methodology. Aquacult. Res. 2019, 50, 3313–3325. [Google Scholar] [CrossRef]
- Pauline, J.M.N.; Achary, A. Novel media for lipid production of Chlorococcum oleofaciens: A RSM approach. Acta Protozool. 2019, 58, 31–41. [Google Scholar] [CrossRef]
- Mubarak, M.; Shaija, A.; Suchithra, T.V. Cost effective approach for production of Chlorella pyrenoidosa: A RSM based study. Waste Biomass Valorization 2019, 10, 3307–3319. [Google Scholar] [CrossRef]
- Fawley, M.W.; Fawley, K.P. A Simple and Rapid Technique for The Isolation Of DNA From Microalgae. J. Phycol. 2004, 40, 223–225. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Proto- Cols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., Whire, T.J., Eds.; Academic Press: London, UK, 1990; pp. 315–322. [Google Scholar]
- Fei, C.; Zou, S.; Wang, T.; Wang, C.; Kemuma, N.D.; He, M.; Amin, S.A.; Wang, C. A Quick Method for Obtaining High-Quality DNA Barcodes without DNA Extraction in Microalgae. J. Appl. Phycol. 2020, 32, 1165–1175. [Google Scholar] [CrossRef]
- Andersen, R.A.; Berges, J.A.; Harrison, P.J.; Watanabe, M.M. Appendix A—Recipes for Freshwater and Seawater Media. In Algal Culturing Techniques; Andersen, R.A., Ed.; Elsevier Academic Press: Burlington, MA, USA, 2005; pp. 429–538. [Google Scholar]
- Sanchez-Galvis, E.M.; Cardenas-Gutierrez, I.Y.; Contreras-Ropero, J.E.; García-Martínez, J.B.; Barajas-Solano, A.F.; Zuorro, A. An Innovative Low-Cost Equipment for Electro-Concentration of Microalgal Biomass. Appl. Sci. 2020, 10, 4841. [Google Scholar] [CrossRef]
- Mishra, S.K.; Suh, W.I.; Farooq, W.; Moon, M.; Shrivastav, A.; Park, M.S.; Yang, J.W. Rapid Quantification of Microalgal Lipids in Aqueous Medium by a Simple Colorimetric Method. Bioresour. Technol. 2014, 155, 330–333. [Google Scholar] [CrossRef]
- García-Martínez, J.B.; Ayala-Torres, E.; Reyes-Gómez, O.; Zuorro, A.; Barajas-Solano, A.F.; Barajas-Ferreira, C. Evaluation of a Two-Phase Extraction System of Carbohydrates and Proteins from Chlorella Vulgaris Utex 1803. Chem. Eng. Trans. 2016, 49, 355–360. [Google Scholar] [CrossRef]
- Mota, M.F.S.; Souza, M.F.; Bon, E.P.S.; Rodrigues, M.A.; Freitas, S.P. Colorimetric Protein Determination in Microalgae (Chlorophyta): Association of Milling and SDS Treatment for Total Protein Extraction. J. Phycol. 2018, 54, 577–580. [Google Scholar] [CrossRef]
- Hynstova, V.; Sterbova, D.; Klejdus, B.; Hedbavny, J.; Huska, D.; Adam, V. Separation, Identification and Quantification of Carotenoids and Chlorophylls in Dietary Supplements Containing Chlorella Vulgaris and Spirulina Platensis Using High-Performance Thin Layer Chromatography. J. Pharm. Biomed. Anal. 2018, 148, 108–118. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Zhao, X.-Q.; Yen, H.-W.; Ho, S.-H.; Cheng, C.-L.; Lee, D.-J.; Bai, F.-W.; Chang, J.-S. Microalgae-Based Carbohydrates for Biofuel Production. Biochem. Eng. J. 2013, 78, 1–10. [Google Scholar] [CrossRef]
- Ruangsomboon, S. Effects of Different Media and Nitrogen Sources and Levels on Growth and Lipid of Green Microalga Botryococcus Braunii KMITL and Its Biodiesel Properties Based on Fatty Acid Composition. Bioresour. Technol. 2015, 191, 377–384. [Google Scholar] [CrossRef]
- Whangchai, K.; Mathimani, T.; Sekar, M.; Shanmugam, S.; Brindhadevi, K.; Van Hung, T.; Chinnathambi, A.; Alharbi, S.A.; Pugazhendhi, A. Synergistic Supplementation of Organic Carbon Substrates for Upgrading Neutral Lipids and Fatty Acids Contents in Microalga. J. Environ. Chem. Eng. 2021, 9, 105482. [Google Scholar] [CrossRef]
- Rehman, Z.U.; Anal, A.K. Enhanced Lipid and Starch Productivity of Microalga (Chlorococcum sp. TISTR 8583) with Nitrogen Limitation Following Effective Pretreatments for Biofuel Production. Biotechnol. Rep. 2019, 21, e00298. [Google Scholar] [CrossRef]
- Xu, J.; Li, T.; Li, C.-L.; Zhu, S.-N.; Wang, Z.-M.; Zeng, E.Y. Lipid Accumulation and Eicosapentaenoic Acid Distribution in Response to Nitrogen Limitation in Microalga Eustigmatos Vischeri JHsu-01 (Eustigmatophyceae). Algal Res. 2020, 48, 101910. [Google Scholar] [CrossRef]
- Zarrinmehr, M.J.; Farhadian, O.; Heyrati, F.P.; Keramat, J.; Koutra, E.; Kornaros, M.; Daneshvar, E. Effect of Nitrogen Concentration on the Growth Rate and Biochemical Composition of the Microalga, Isochrysis Galbana. Egypt. J. Aquat. Res. 2020, 46, 153–158. [Google Scholar] [CrossRef]
- Kokabi, K.; Gorelova, O.; Ismagulova, T.; Itkin, M.; Malitsky, S.; Boussiba, S.; Solovchenko, A.; Khozin-Goldberg, I. Metabolomic Foundation for Differential Responses of Lipid Metabolism to Nitrogen and Phosphorus Deprivation in an Arachidonic Acid-Producing Green Microalga. Plant Sci. 2019, 283, 95–115. [Google Scholar] [CrossRef]
- Chakravarty, S.; Mallick, N. Engineering a Cultivation Strategy for Higher Lipid Accretion and Biodiesel Production by the Marine Microalga Picochlorum Soloecismus. Sustain. Chem. Pharm. 2022, 26, 100635. [Google Scholar] [CrossRef]
- Zuorro, A. Enhanced Lycopene Extraction from Tomato Peels by Optimized Mixed-Polarity Solvent Mixtures. Molecules 2020, 25, 2038. [Google Scholar] [CrossRef] [PubMed]
- Zuorro, A.; Lavecchia, R. Polyphenols and Energy Recovery from Spent Coffee Grounds. Chem. Eng. Trans. 2011, 25, 285–290. [Google Scholar] [CrossRef]
- Zuorro, A.; Iannone, A.; Natali, S.; Lavecchia, R. Green Synthesis of Silver Nanoparticles Using Bilberry and Red Currant Waste Extracts. Processes 2019, 7, 193. [Google Scholar] [CrossRef]
- Montanaro, D.; Lavecchia, R.; Petrucci, E.; Zuorro, A. UV-Assisted Electrochemical Degradation of Coumarin on Boron-Doped Diamond Electrodes. Chem. Eng. J. 2017, 323, 512–519. [Google Scholar] [CrossRef]
- Zuorro, A.; Maffei, G.; Lavecchia, R. Kinetic Modeling of Azo Dye Adsorption on Non-Living Cells of Nannochloropsis Oceanica. J. Environ. Chem. Eng. 2017, 5, 4121–4127. [Google Scholar] [CrossRef]
- Singh, P.; Guldhe, A.; Kumari, S.; Rawat, I.; Bux, F. Investigation of Combined Effect of Nitrogen, Phosphorus and Iron on Lipid Productivity of Microalgae Ankistrodesmus Falcatus KJ671624 Using Response Surface Methodology. Biochem. Eng. J. 2015, 94, 22–29. [Google Scholar] [CrossRef]
- Polat, E.; Yüksel, E.; Altınbaş, M. Mutual Effect of Sodium and Magnesium on the Cultivation of Microalgae Auxenochlorella Protothecoides. Biomass Bioenergy 2020, 132, 105441. [Google Scholar] [CrossRef]
- Vishwakarma, R.; Dhar, D.W.; Pabbi, S. Formulation of a Minimal Nutritional Medium for Enhanced Lipid Productivity in Chlorella sp. and Botryococcus sp. Using Response Surface Methodology. Water Sci. Technol. 2018, 77, 1660–1672. [Google Scholar] [CrossRef]
- Tran, H.-L.; Kwon, J.-S.; Kim, Z.-H.; Oh, Y.; Lee, C.-G. Statistical Optimization of Culture Media for Growth and Lipid Production of Botryococcus Braunii LB572. Biotechnol. Bioprocess Eng. 2010, 15, 277–284. [Google Scholar] [CrossRef]
- Cheng, K.C.; Ren, M.; Ogden, K.L. Statistical Optimization of Culture Media for Growth and Lipid Production of Chlorella protothecoides UTEX 250. Bioresour. Technol. 2013, 128, 44–48. [Google Scholar] [CrossRef]
- León-Vaz, A.; León, R.; Díaz-Santos, E.; Vigara, J.; Raposo, S. Using Agro-Industrial Wastes for Mixotrophic Growth and Lipids Production by the Green Microalga Chlorella Sorokiniana. New Biotechnol. 2019, 51, 31–38. [Google Scholar] [CrossRef]
- Li, T.; Zheng, Y.; Yu, L.; Chen, S. Mixotrophic Cultivation of a Chlorella Sorokiniana Strain for Enhanced Biomass and Lipid Production. Biomass Bioenergy 2014, 66, 204–213. [Google Scholar] [CrossRef]
- Ra, C.H.; Kang, C.-H.; Kim, N.K.; Lee, C.-G.; Kim, S.-K. Cultivation of Four Microalgae for Biomass and Oil Production Using a Two-Stage Culture Strategy with Salt Stress. Renew. Energy 2015, 80, 117–122. [Google Scholar] [CrossRef]
- Fawzy, M.A.; Alharthi, S. Use of Response Surface Methodology in Optimization of Biomass, Lipid Productivity and Fatty Acid Profiles of Marine Microalga Dunaliella Parva for Biodiesel Production. Environ. Technol. Innov. 2021, 22, 101485. [Google Scholar] [CrossRef]
- Pandey, A.; Gupta, A.; Sunny, A.; Kumar, S.; Srivastava, S. Multi-Objective Optimization of Media Components for Improved Algae Biomass, Fatty Acid and Starch Biosynthesis from Scenedesmus sp. ASK22 Using Desirability Function Approach. Renew. Energy 2020, 150, 476–486. [Google Scholar] [CrossRef]
- Choix, F.J.; De-Bashan, L.E.; Bashan, Y. Enhanced Accumulation of Starch and Total Carbohydrates in Alginate-Immobilized Chlorella Spp. Induced by Azospirillum Brasilense: I. Autotrophic Conditions. Enzyme Microb. Technol. 2012, 51, 294–299. [Google Scholar] [CrossRef]
- Ho, S.-H.; Chen, C.-Y.; Yeh, K.-L.; Chen, W.-M.; Lin, C.-Y.; Chang, J.-S. Characterization of Photosynthetic Carbon Dioxide Fixation Ability of Indigenous Scenedesmus Obliquus Isolates. Biochem. Eng. J. 2010, 53, 57–62. [Google Scholar] [CrossRef]
- de Farias Silva, C.E.; Sforza, E.; Bertucco, A. Stability of Carbohydrate Production in Continuous Microalgal Cultivation under Nitrogen Limitation: Effect of Irradiation Regime and Intensity on Tetradesmus Obliquus. J. Appl. Phycol. 2018, 30, 261–270. [Google Scholar] [CrossRef]
- McClain, A.M.; Sharkey, T.D. Triose Phosphate Utilization and beyond: From Photosynthesis to End Product Synthesis. J. Exp. Bot. 2019, 70, 1755–1766. [Google Scholar] [CrossRef]
- Ghosh, A.; Samadhiya, K.; Kashyap, M.; Anand, V.; Sangwan, P.; Bala, K. The Use of Response Surface Methodology for Improving Fatty Acid Methyl Ester Profile of Scenedesmus Vacuolatus. Environ. Sci. Pollut. Res. 2020, 27, 27457–27469. [Google Scholar] [CrossRef] [PubMed]
- González-Fernández, C.; Ballesteros, M. Linking Microalgae and Cyanobacteria Culture Conditions and Key-Enzymes for Carbohydrate Accumulation. Biotechnol. Adv. 2012, 30, 1655–1661. [Google Scholar] [CrossRef] [PubMed]
- Dragone, G.; Fernandes, B.D.; Abreu, A.P.; Vicente, A.A.; Teixeira, J.A. Nutrient Limitation as a Strategy for Increasing Starch Accumulation in Microalgae. Appl. Energy 2011, 88, 3331–3335. [Google Scholar] [CrossRef]
- Muthuraj, M.; Kumar, V.; Palabhanvi, B.; Das, D. Evaluation of Indigenous Microalgal Isolate Chlorella sp. FC2 IITG as a Cell Factory for Biodiesel Production and Scale up in Outdoor Conditions. J. Ind. Microbiol. Biotechnol. 2014, 41, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Tourang, M.; Baghdadi, M.; Torang, A.; Sarkhosh, S. Optimization of Carbohydrate Productivity of Spirulina Microalgae as a Potential Feedstock for Bioethanol Production. Int. J. Environ. Sci. Technol. 2019, 16, 1303–1318. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Beisson, F.; Riekhof, W. Metabolism of Acyl-Lipids in Chlamydomonas Reinhardtii. Plant J. 2015, 82, 504–522. [Google Scholar] [CrossRef]
- Chakravarty, S.; Mallick, N. Optimization of Lipid Accumulation in an Aboriginal Green Microalga Selenastrum sp. GA66 for Biodiesel Production. Biomass Bioenergy 2019, 126, 1–13. [Google Scholar] [CrossRef]
- Schüler, L.M.; Santos, T.; Pereira, H.; Duarte, P.; Katkam, N.G.; Florindo, C.; Schulze, P.S.C.; Barreira, L.; Varela, J.C.S. Improved Production of Lutein and β-Carotene by Thermal and Light Intensity Upshifts in the Marine Microalga Tetraselmis sp. CTP4. Algal Res. 2020, 45, 101732. [Google Scholar] [CrossRef]
- Aburai, N.; Sumida, D.; Abe, K. Effect of Light Level and Salinity on the Composition and Accumulation of Free and Ester-Type Carotenoids in the Aerial Microalga Scenedesmus sp. (Chlorophyceae). Algal Res. 2015, 8, 30–36. [Google Scholar] [CrossRef]
- de Souza da Silva, S.P.; Perrone, D.; do Valle, A.F. Optimization of Arthrospira Maxima Cultivation for Biomass and Protein Production and Biomass Technological Treatment to Color, Flavor, and Aroma Masking for Addition to Food Products. J. Appl. Phycol. 2022, 34, 65–80. [Google Scholar] [CrossRef]
- Kumaran, J.; Poulose, S.; Joseph, V.; Bright Singh, I.S. Enhanced Biomass Production and Proximate Composition of Marine Microalga Nannochloropsis Oceanica by Optimization of Medium Composition and Culture Conditions Using Response Surface Methodology. Anim. Feed Sci. Technol. 2021, 271, 114761. [Google Scholar] [CrossRef]








| Factor | Units | Low Level (−1) | High Level (+1) |
|---|---|---|---|
| Carbon source | g/L | 0.2 | 0.5 |
| NaNO3 | g/L | 0.125 | 0.25 |
| K2HPO4 | g/L | 0.0375 | 0.075 |
| KH2PO4 | g/L | 0.0875 | 0.175 |
| Standard Run | Block | NaNO3 | K2HPO4 | KH2PO4 | Carbon Content |
|---|---|---|---|---|---|
| g/L | g/L | g/L | (g/L) | ||
| 1 | 1 | 0.125 | 0.0375 | 0.0875 | 0.20 |
| 2 | 0.125 | 0.075 | 0.175 | 0.50 | |
| 3 | 0.25 | 0.0375 | 0.0875 | 0.50 | |
| 4 | 0.25 | 0.075 | 0.175 | 0.20 | |
| 5 (Central point) | 0.1875 | 0.056 | 0.131 | 0.35 | |
| 6 | 2 | 0.125 | 0.0375 | 0.0875 | 0.50 |
| 7 | 0.125 | 0.075 | 0.175 | 0.20 | |
| 8 | 0.25 | 0.0375 | 0.0875 | 0.20 | |
| 9 | 0.25 | 0.075 | 0.175 | 0.50 | |
| 10 (Central point) | 0.1875 | 0.056 | 0.131 | 0.35 | |
| 11 | 3 | 0.083 | 0.056 | 0.131 | 0.35 |
| 12 | 0.292 | 0.056 | 0.131 | 0.35 | |
| 13 | 0.1875 | 0.0249 | 0.0581 | 0.35 | |
| 14 | 0.1875 | 0.0876 | 0.2044 | 0.35 | |
| 15 | 0.1875 | 0.056 | 0.131 | 0.099 | |
| 16 | 0.1875 | 0.056 | 0.131 | 0.61 | |
| 17 (Central point) | 0.1875 | 0.056 | 0.131 | 0.35 |
| Strain | Variable | Units | Value |
|---|---|---|---|
| Chlorella sp. UFPS019 | Na2CO3 | g/L | 3.53 |
| KH2PO4 | 0.06 | ||
| K2HPO4 | 0.14 | ||
| NaNO3 | 0.1875 | ||
| carbohydrates | % w/w | 50 | |
| C2H3NaO2 | g/L | 1.19 | |
| KH2PO4 | 0.056 | ||
| K2HPO4 | 0.131 | ||
| NaNO3 | 0.075 | ||
| Lipids | % w/w | 27.5 | |
| Desmodesmus sp. UFPS021 | C2H3NaO2 | g/L | 2 |
| KH2PO4 | 0.056 | ||
| K2HPO4 | 0.131 | ||
| NaNO3 | 0.25 | ||
| Proteins | % w/w | 38.6 |
| Chlorella sp. UFPS019 (Carbohydrates) | ||
| µ | 0.0029 h−1 | |
| dt | 120 h−1 | |
| NO3 | PO4 | |
| 0.2819 | 0.0698 | |
| 0.1885 | 0.0768 | |
| Chlorella sp. UFPS019 (Lipids) | ||
| µ | 0.00647 h−1 | |
| dt | 120 h−1 | |
| NO3 | PO4 | |
| 0.1566 | 0.0318 | |
| 0.004 | 0.001209 | |
| Desmodesmus sp. UFPS021 (Proteins) | ||
| µ | 0.00567 h−1 | |
| dt | 120 h−1 | |
| NO3 | PO4 | |
| 0.36 | 0.0775 | |
| 0.0284 | 0.0772 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suárez Quintana, W.H.; García-Rico, R.O.; García-Martínez, J.B.; Urbina-Suarez, N.A.; López-Barrera, G.L.; Barajas-Solano, A.F.; Zuorro, A. Enhancement of Metabolite Production in High-Altitude Microalgal Strains by Optimized C/N/P Ratio. Appl. Sci. 2022, 12, 6779. https://doi.org/10.3390/app12136779
Suárez Quintana WH, García-Rico RO, García-Martínez JB, Urbina-Suarez NA, López-Barrera GL, Barajas-Solano AF, Zuorro A. Enhancement of Metabolite Production in High-Altitude Microalgal Strains by Optimized C/N/P Ratio. Applied Sciences. 2022; 12(13):6779. https://doi.org/10.3390/app12136779
Chicago/Turabian StyleSuárez Quintana, William H., Ramón O. García-Rico, Janet B. García-Martínez, Néstor A. Urbina-Suarez, Germán L. López-Barrera, Andrés F. Barajas-Solano, and Antonio Zuorro. 2022. "Enhancement of Metabolite Production in High-Altitude Microalgal Strains by Optimized C/N/P Ratio" Applied Sciences 12, no. 13: 6779. https://doi.org/10.3390/app12136779
APA StyleSuárez Quintana, W. H., García-Rico, R. O., García-Martínez, J. B., Urbina-Suarez, N. A., López-Barrera, G. L., Barajas-Solano, A. F., & Zuorro, A. (2022). Enhancement of Metabolite Production in High-Altitude Microalgal Strains by Optimized C/N/P Ratio. Applied Sciences, 12(13), 6779. https://doi.org/10.3390/app12136779