
Identification of Seasonal Honey Based on Quantitative Detection of Typical Pollen DNA


Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Major Nectar Plants
2.2. Honey Samples
2.3. DNA Extraction
2.4. Primer Design and Standard DNA Construction
2.5. Polymerase Chain Reaction
2.6. Standard Curves for DNA Copy Number Estimation
3. Results
3.1. Standard Linear Regression Curves and Limit of Detection
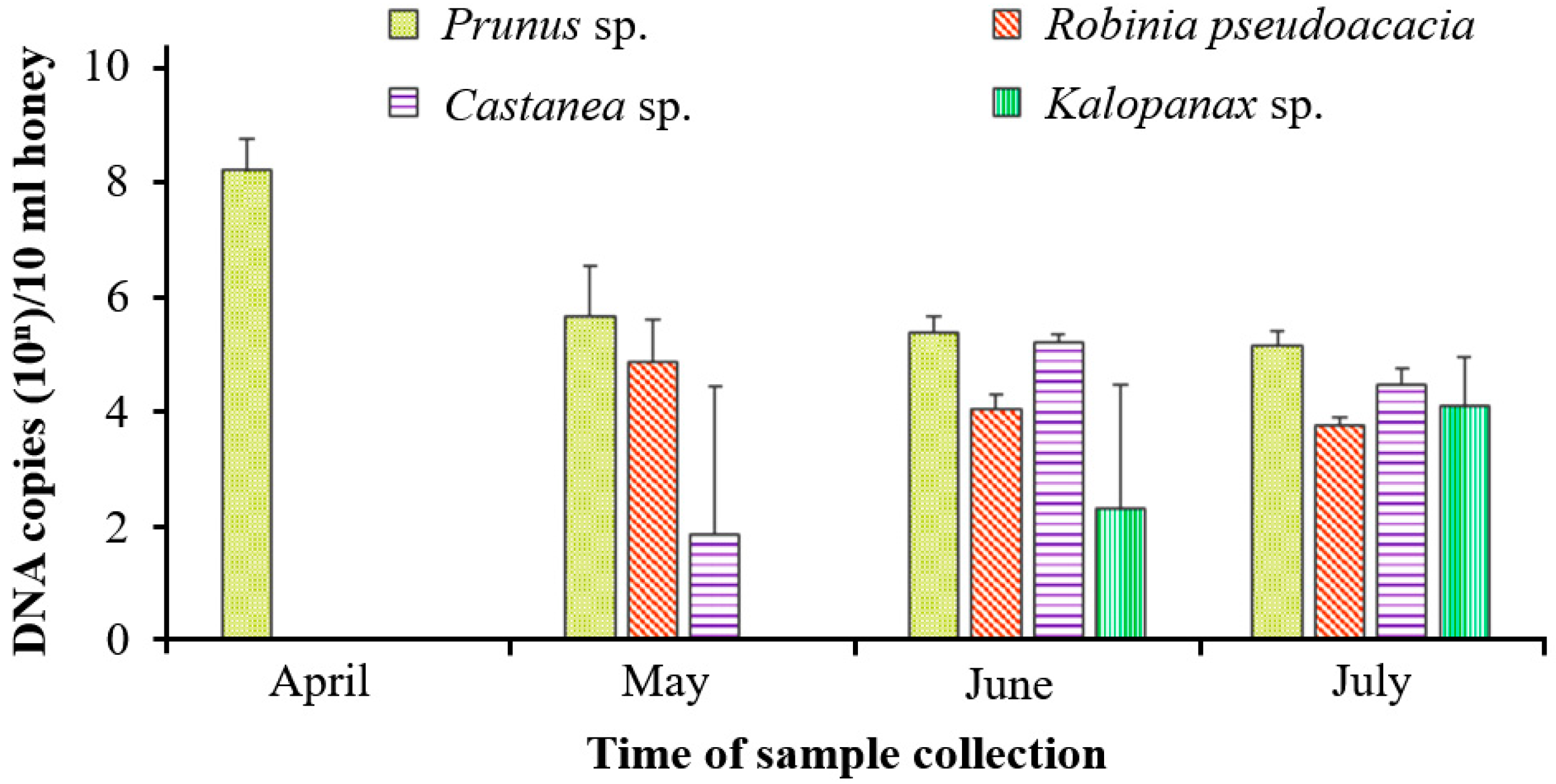
3.2. Quantification of the Target Pollen DNA in Natural Honey Samples
3.3. Detection of the Targeted Pollen DNA in Commercial Honey Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- García, N.L. The Current Situation on the International Honey Market. Bee World 2018, 95, 89–94. [Google Scholar] [CrossRef]
- Andrade, P.B.; Amaral, M.T.; Isabel, P.; Carvalho, J.C.M.F.; Seabra, R.M.; da Cunha, A.P. Physicochemical attributes and pollen spectrum of Portuguese heather honeys. Food Chem. 1999, 66, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Manzanares, A.B.; García, Z.H.; Galdón, B.R.; Rodríguez-Rodríguez, E.M.; Romero, C.D. Physicochemical characteristics and pollen spectrum of monofloral honeys from Tenerife, Spain. Food Chem. 2017, 228, 441–446. [Google Scholar] [CrossRef]
- Corvucci, F.; Nobili, L.; Mellucci, D.; Grillenzoni, F.V. The discrimination of honey origin using melissopalynology and Raman spectroscopy techniques coupled with multivariate analysis. Food Chem. 2015, 169, 297–304. [Google Scholar] [CrossRef]
- El-Senduny, F.F.; Hegazi, N.M.; Elghani, G.E.A.; Farag, M.A. Manuka honey, a unique mono-floral honey. A comprehensive review of its bioactives, metabolism, action mechanisms, and therapeutic merits. Food Biosci. 2021, 42, 101038. [Google Scholar] [CrossRef]
- Ahmed, S.; Othman, N.H. Review of the medicinal effects of tualang honey and a comparison with manuka honey. Malays. J. Med. Sci. 2013, 20, 6–13. [Google Scholar]
- Yaghoobi, R.; Kazerouni, A.; Kazerouni, O. Evidence for clinical use of honey in wound healing as an anti-bacterial, anti-inflammatory, anti-oxidant and anti-viral agent: A review. Jundishapur. J. Nat. Pharm. Prod. 2013, 8, 100–104. [Google Scholar] [CrossRef] [Green Version]
- Ronsisvalle, S.; Lissandrello, E.; Fuochi, V.; Petronio, P.G.; Straquadanio, C.; Crascì, L.; Panico, A.; Milito, M.; Cova, A.M.; Tempera, G.; et al. Antioxidant and antimicrobial properties of Casteanea sativa Miller chestnut honey produced on Mount Etna (Sicily). Nat. Prod. Res. 2019, 33, 843–850. [Google Scholar] [CrossRef]
- Bogdanov, S.; Martin, P. Honey authenticity. Mitteilungen aus Lebensmitteluntersuchung und Hygiene 2002, 93, 232–254. [Google Scholar]
- Hailu, D.; Belay, A. Melissopalynology and antioxidant properties used to differentiate Schefflera abyssinica and polyfloral honey. PLoS ONE 2020, 15, e0240868. [Google Scholar] [CrossRef]
- Bobi, O.; Mãrghita, L.A.; Dezmirean, D.S.; Bãrnuiu, L.I.; Mãrgãoan, R.; Bogdan Gherman, B.; Bonta, V. The Importance of melissopalynology in addition to physical-chemical analysis on botanical authenticity testing of monofloral honey. Bull. UASVM Anim. Sci. Biotechnol. 2013, 70, 24–30. [Google Scholar]
- Mureșan, C.I.; Cornea-Cipcigan, M.; Suharoschi, R.; Erler, S.; Mărgăoan, R. Honey botanical origin and honey-specific protein pattern: Characterization of some European honeys. LWT 2022, 154, 112883. [Google Scholar] [CrossRef]
- Olivieri, C.; Marota, I.; Rollo, F.; Luciani, S. Tracking plant, fungal, and bacterial DNA in honey specimens. J. Forensic Sci. 2012, 57, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Bagella, S.; Satta, A.; Floris, I.; Caria, M.C.; Rossetti, I.; Podani, J. Effects of plant community composition and flowering phenology on honeybee foraging in Mediterranean sylvo-pastoral systems. Appl. Veg. Sci. 2013, 16, 689–697. [Google Scholar] [CrossRef]
- Bruni, I.; Galimberti, A.; Caridi, L.; Scaccabarozzi, D.; De Mattia, F.; Casiraghi, M.; Labra, M. A DNA barcoding approach to identify plant species in multiflower honey. Food Chem. 2015, 170, 308–315. [Google Scholar] [CrossRef]
- Puścion-Jakubik, A.; Borawska, M.H.; Socha, K. Modern Methods for Assessing the Quality of Bee Honey and Botanical Origin Identification. Foods 2020, 9, 1028. [Google Scholar] [CrossRef]
- Soares, S.; Grazina, L.; Costa, J.; Amaral, J.S.; Maria, B.P.P.; Oliveira, M.B.P.P.; Mafra, I. Botanical authentication of lavender (Lavandula spp.) honey by a novel DNA-barcoding approach coupled to high resolution melting analysis. Food Control 2018, 86, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Manivanan, P.; Rajagopalan, S.M.; Subbarayalu, M. Studies on authentication of true source of honey using pollen DNA barcoding. J. Entomol. Zool. Stud. 2018, 6, 255–261. [Google Scholar]
- Soares, S.; Amaral, J.S.; Oliveira, M.B.P.P.; Mafra, I. Improving DNA isolation from honey for the botanical origin identification. Food Control 2015, 48, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Kohsaka, R.; Park, M.S.; Uchiyama, Y. Beekeeping and honey production in Japan and South Korea: Past and present. J. Ethn. Foods 2017, 4, 72–79. [Google Scholar] [CrossRef]
- Korean Beekeeping Association. Statistical Survey of Korean Beekeeping Industry. Available online: https://korapis.or.kr/jsp/sub3-1_01.jsp (accessed on 29 April 2022).
- Truong, A.-T.; Yoo, M.S.; Yun, B.R.; Kang, J.E.; Noh, J.; Hwang, T.J.; Seo, S.K.; Yoon, S.S.; Cho, Y.S. Prevalence and pathogen detection of Varroa and Tropilaelaps mites in Apis mellifera (Hymenoptera, Apidae) apiaries in South Korea. J. Apic. Res. 2022. [Google Scholar] [CrossRef]
- Kim, Y.K.; Kim, M.S.; Nam, J.I.; Song, J.H.; Kim, S.H. Analysis on floral nectar characteristics among the selected black locust (Robinia spp.) individuals. J. Apic. Res. 2021. [Google Scholar] [CrossRef]
- Jung, C.; Cho, S.K. Relationship between honey bee population and honey production in Korea: A historical trend analysis. Korean J. Apic. 2015, 30, 7–12. [Google Scholar]
- Ryu, J.; Jang, J. Major Nectar Plant for Beekeeping in Korea; Fuzzycom Media: Daegu, Korea, 2007. [Google Scholar]
- Kamo, T.; Kusumoto, Y.; Tokuoka, Y.; Okubo, S.; Hayakawa, H.; Yoshiyama, M.; Kimura, K.; Konuma, A. A DNA barcoding method for identifying and quantifying the composition of pollen species collected by European honeybees, Apis mellifera (Hymenoptera: Apidae). Appl. Entomol. Zool. 2018, 53, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Holton, T.A.; Graham, M.W. A simple and efficient method for direct cloning of PCR products using ddT-tailed vectors. Nucleic Acids Res. 1991, 19, 1156. [Google Scholar] [CrossRef] [Green Version]
- Pires, J.; Estevinho, M.L.; Feas, X.; Cantalapiedra, J.; Iglesias, A. Pollen spectrum and physico-chemical attributes of heather (Erica sp.) honeys of north Portugal. J. Sci. Food Agric. 2009, 89, 1862–1870. [Google Scholar] [CrossRef] [Green Version]
- Soares, S.; Amaral, J.S.; Oliveira, M.B.P.P.; Mafra, I. A comprehensive review on the main honey authentication issues: Production and origin. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1072–1100. [Google Scholar] [CrossRef] [Green Version]
- Carroll, M.J.; Brown, N.; Goodall, C.; Downs, A.M.; Sheenan, T.H.; Anderson, K.E. Honey bees preferentially consume freshly-stored pollen. PLoS ONE 2017, 12, e0175933. [Google Scholar] [CrossRef] [Green Version]
- Bouseta, A.; Scheirman, V.; Collin, S. Flavor and free amino acid composition of lavender and eucalyptus honeys. J. Food Sci. 1996, 61, 683–687. [Google Scholar] [CrossRef]
- Gomes, T.; Feás, X.; Iglesias, A.; Estevinho, L.M. Study of organic honey from the northeast of Portugal. Molecules 2011, 16, 5374–5386. [Google Scholar] [CrossRef] [PubMed]
- Gune, M.E.; Şahin, S.; Demir, C.; Borum, E.; Tosunoglu, A. Determination of phenolic compounds profile in chestnut and floral honeys and their antioxidant and antimicrobial activities. J. Food Biochem. 2016, 41, e12345. [Google Scholar] [CrossRef]
- Louveaux, J.; Maurizio, A.; Vorwohl, G. Methods of melissopalynology. Bee World 1978, 59, 139–157. [Google Scholar] [CrossRef]
- Milla, L.; Schmidt-Lebuhn, A.; Bovill, J.; Encinas-Viso, F. Monitoring of honey bee floral resources with pollen DNA metabarcoding as a complementary tool to vegetation surveys. Ecol. Solut. Evid. 2022, 3, e12120. [Google Scholar] [CrossRef]
- Utzeri, V.J.; Ribani, A.; Schiavo, G.; Bertolini, F.; Bovo, S.; Fontanesi, L. Application of next generation semiconductor based sequencing to detect the botanical composition of monofloral, polyfloral and honeydew honey. Food Control 2018, 86, 342–349. [Google Scholar] [CrossRef]
- Chin, N.L.; Sowndhararajan, K. A Review on Analytical Methods for Honey Classification, Identification and Authentication. In Honey Analysis—New Advances and Challenges; de Alencar Arnaut de Toledo, V., Chambó, E.D., Eds.; IntechOpen: London, UK, 2020. [Google Scholar]
- Voyslavov, T.; Mladenova, E.; Balkanska, R. A new approach for determination of the botanical origin of monofloral bee honey, combining mineral content, physicochemical parameters, and self-organizing maps. Molecules 2021, 26, 7219. [Google Scholar] [CrossRef]
- Bianchi, F.; Careri, M.; Musci, M. Volatile norisoprenoids as markers of botanical origin of Sardinian strawberry-tree (Arbutus unedo L.) honey: Characterization of aroma compounds by dynamic headspace extraction and gas chromatography-mass spectrometry. Food Chem. 2005, 89, 527–532. [Google Scholar] [CrossRef]
- Prosser, S.W.J.; Hebert, P.D.N. Rapid identification of the botanical and entomological sources of honey using DNA metabarcoding. Food Chem. 2017, 214, 183–191. [Google Scholar] [CrossRef]
- Wirta, H.; Abrego, N.; Miller, K.; Roslin, T.; Vesterinen, E. DNA traces the origin of honey by identifying plants, bacteria and fungi. Sci. Rep. 2021, 11, 4798. [Google Scholar] [CrossRef]
- McDonald, C.M.; Keeling, S.E.; Brewer, M.J.; Hathaway, S.C. Using chemical and DNA marker analysis to authenticate a high-value food, manuka honey. NPJ Sci. Food 2018, 2, 9. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| No. | Collection Date | Number of Samples | Expected Major Nectar Plant |
|---|---|---|---|
| 1 | 22 April 2019 | 5 | Prunus sp. |
| 2 | 30 May 2019 | 5 | Robinia pseudoacacia |
| 3 | 30 June 2019 | 5 | Castanea sp. |
| 4 | 30 July 2019 | 5 | Kalopanax sp. |
| No. | Primer | Sequence (5′-3′) | Annealing (°C) | Target Plant | Target Gene | Amplicon Size (bp) | Reference |
|---|---|---|---|---|---|---|---|
| 1 | Pruchlo-F | GGTGTACTCTTTCTTCGAGT | 53 | Prunus sp. | ndhF-rpl32 | 205 | This study |
| Pruchlo-R | GAAGTTGATAAAATACAATAC | ||||||
| 2 | Acasia-F | GTGGTGGAACAAAATATCTAGA | 58 | Robinia pseudoacacia | rpoC2 | 299 | |
| Acasia-R | AACGATTTGTTACCGAGCTT | ||||||
| 3 | Castanea-F | CCATGGACCGTATTCTTCG | 61 | Castanea sp. | atpI | 286 | |
| Castanea-R | AGAGGGCAATATGAAATTATG | ||||||
| 4 | Kalopana-F | ACGAAAGAATCGAATATCGA | 57 | Kalopanax sp. | trnL-trnF | 435 | |
| Kalopana-R | GCGAGTTTCAGTATGAATAATT |
| Target Plant | Time of Sample Collection | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| April | May | June | July | |||||||||||||||||
| 1 * | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 | |
| Prunus sp. | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Robinia pseudoacacia | − | − | − | − | − | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Castanea sp. | − | − | − | − | − | + | + | − | − | − | + | + | + | + | + | + | + | + | + | + |
| Kalopanax sp. | − | − | − | − | − | − | − | − | − | − | + | − | + | − | − | + | + | + | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truong, A.-T.; Yoo, M.-S.; Cho, Y.S.; Yoon, B. Identification of Seasonal Honey Based on Quantitative Detection of Typical Pollen DNA. Appl. Sci. 2022, 12, 4846. https://doi.org/10.3390/app12104846
Truong A-T, Yoo M-S, Cho YS, Yoon B. Identification of Seasonal Honey Based on Quantitative Detection of Typical Pollen DNA. Applied Sciences. 2022; 12(10):4846. https://doi.org/10.3390/app12104846
Chicago/Turabian StyleTruong, A-Tai, Mi-Sun Yoo, Yun Sang Cho, and Byoungsu Yoon. 2022. "Identification of Seasonal Honey Based on Quantitative Detection of Typical Pollen DNA" Applied Sciences 12, no. 10: 4846. https://doi.org/10.3390/app12104846
APA StyleTruong, A.-T., Yoo, M.-S., Cho, Y. S., & Yoon, B. (2022). Identification of Seasonal Honey Based on Quantitative Detection of Typical Pollen DNA. Applied Sciences, 12(10), 4846. https://doi.org/10.3390/app12104846