
Improved Biotransformation of Platycoside E into Deapiose-Xylosylated Platycodin D by Cytolase PCL5 under High Hydrostatic Pressure

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Enzyme Assay
2.3. Optimization of Reaction Conditions
2.4. Bioconversion
2.5. HPLC Analysis
3. Results and Discussion
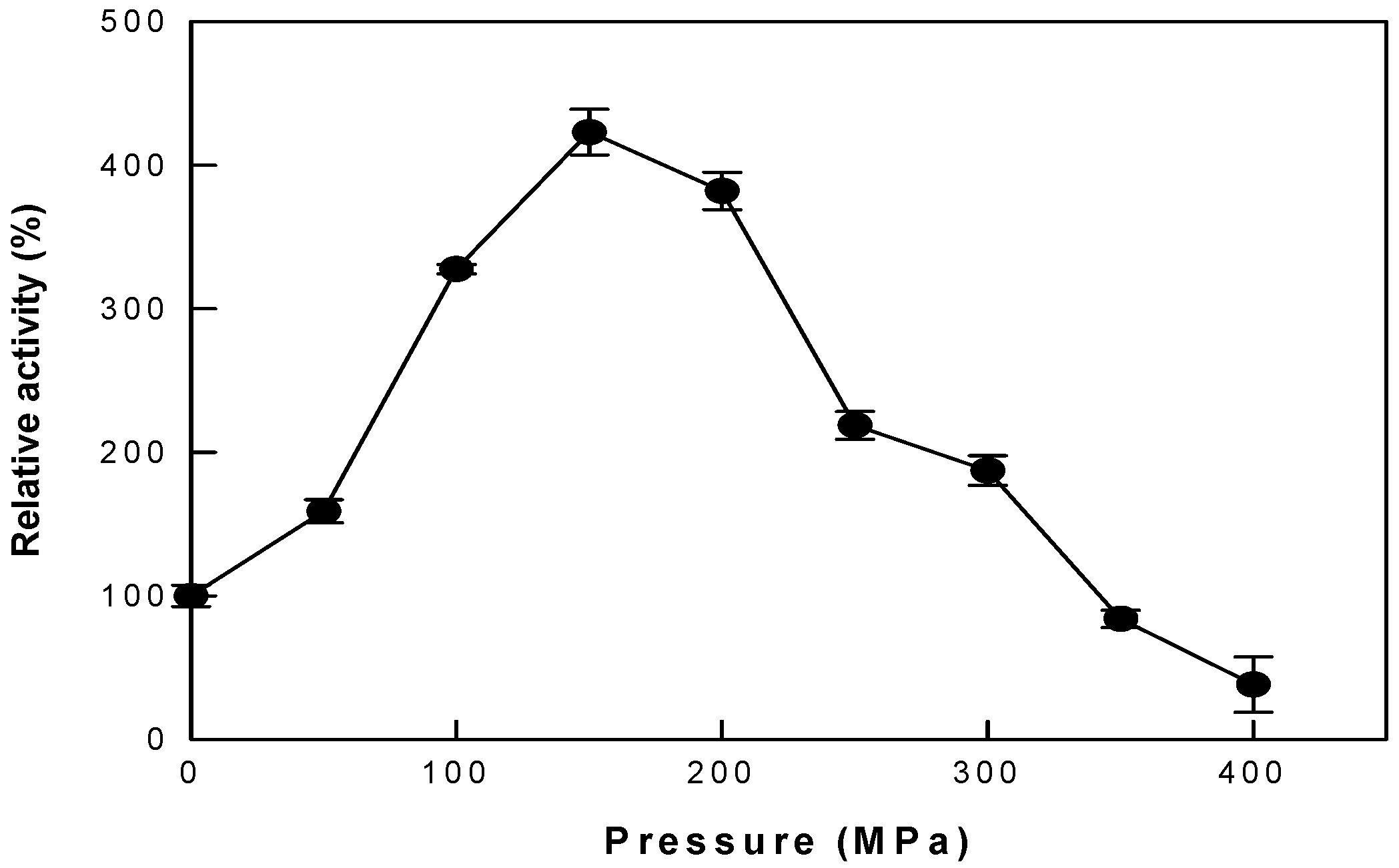
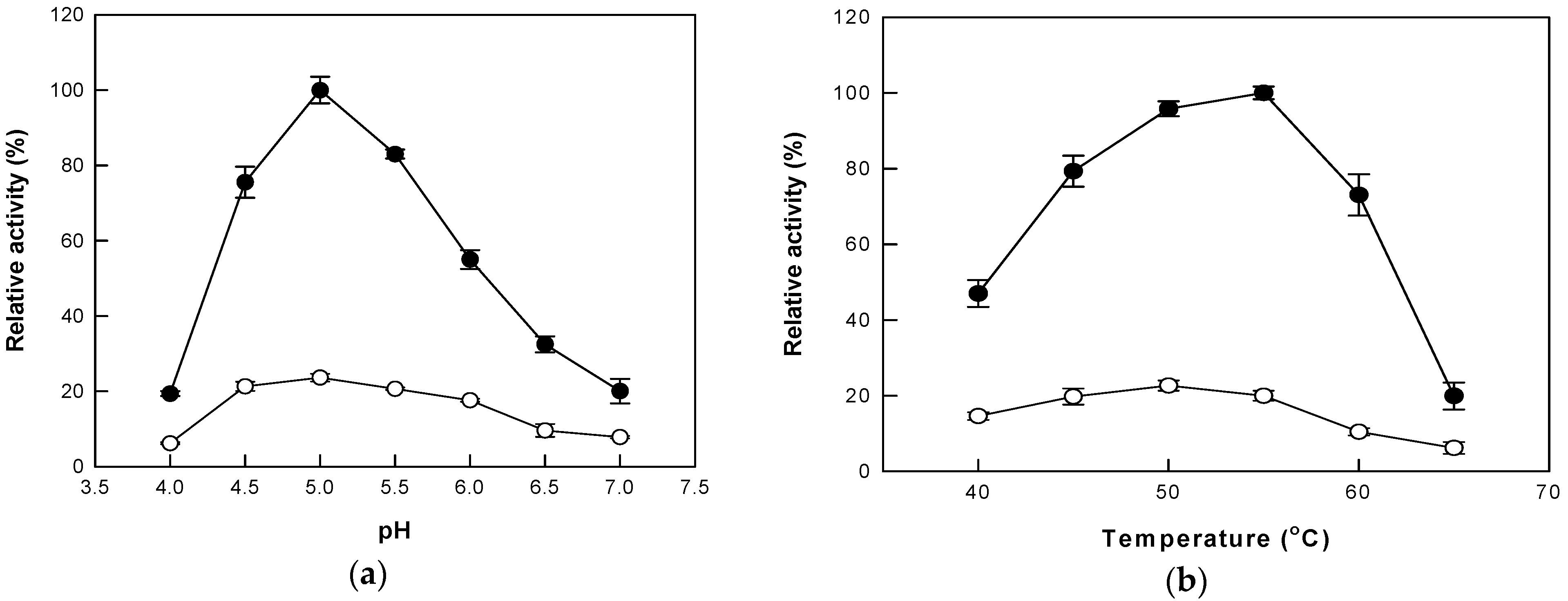
3.1. Effects of Pressure, pH, and Temperature under AP and HHP on Cytolase PCL5 Activity
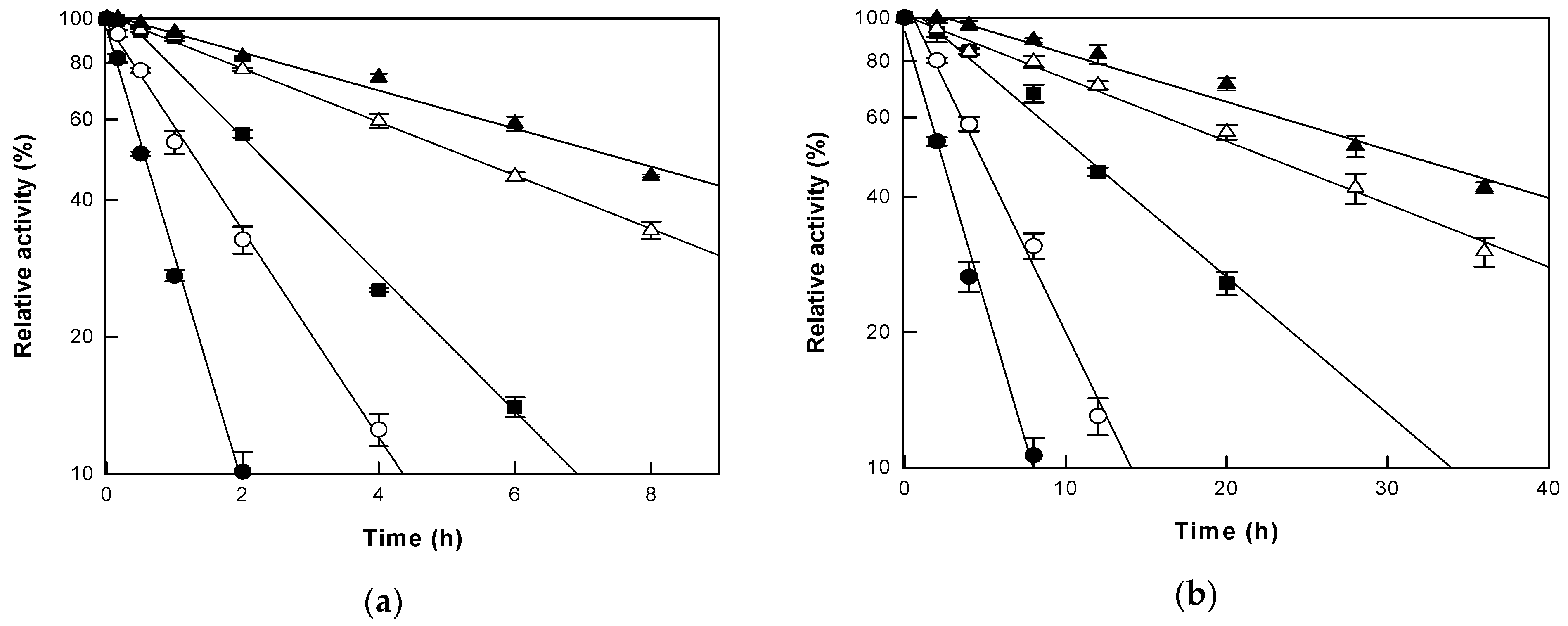
3.2. Thermal Stability of Cytolase PCL5 under AP and HHP
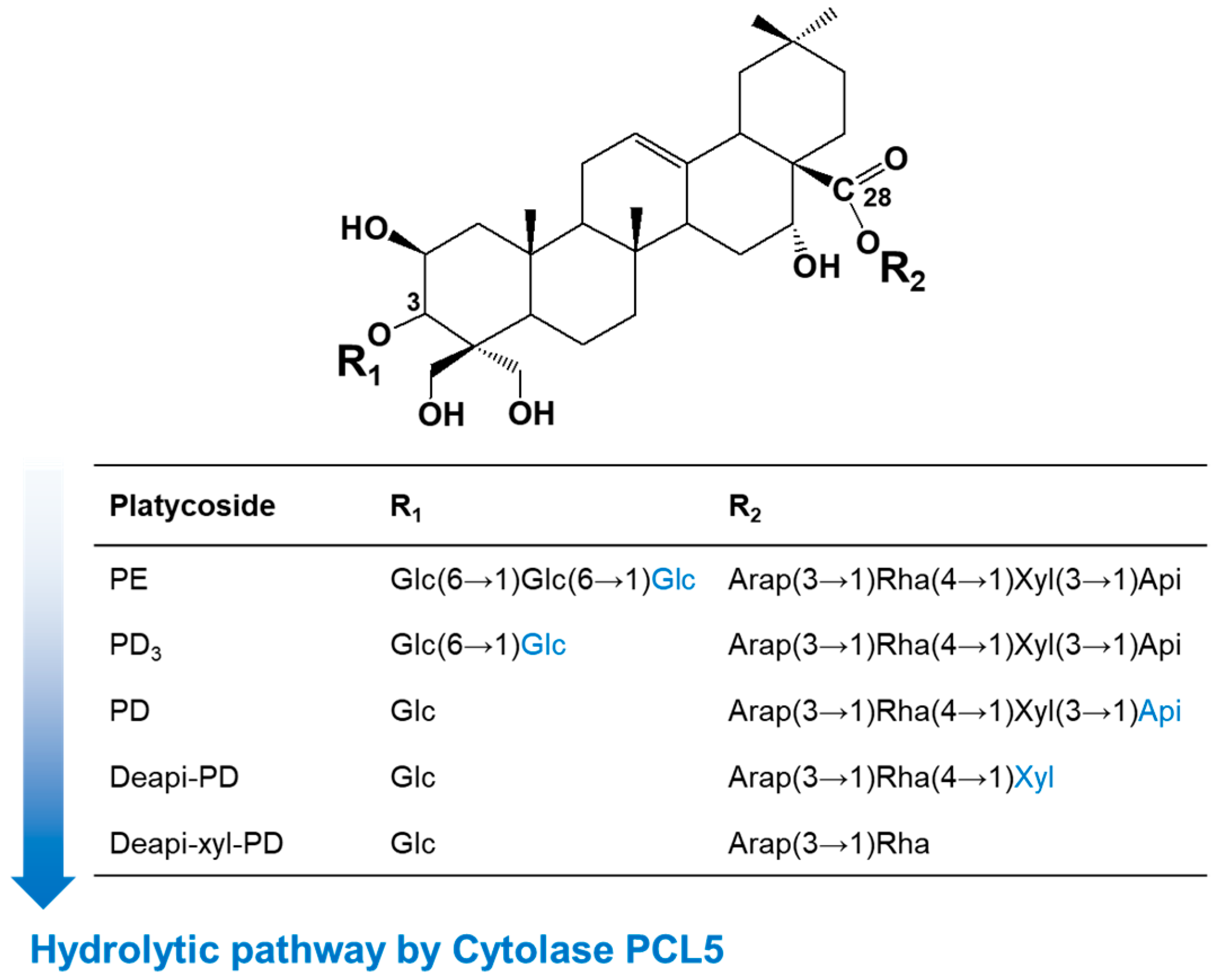
3.3. Changes in Substrate Specificity with Pressure
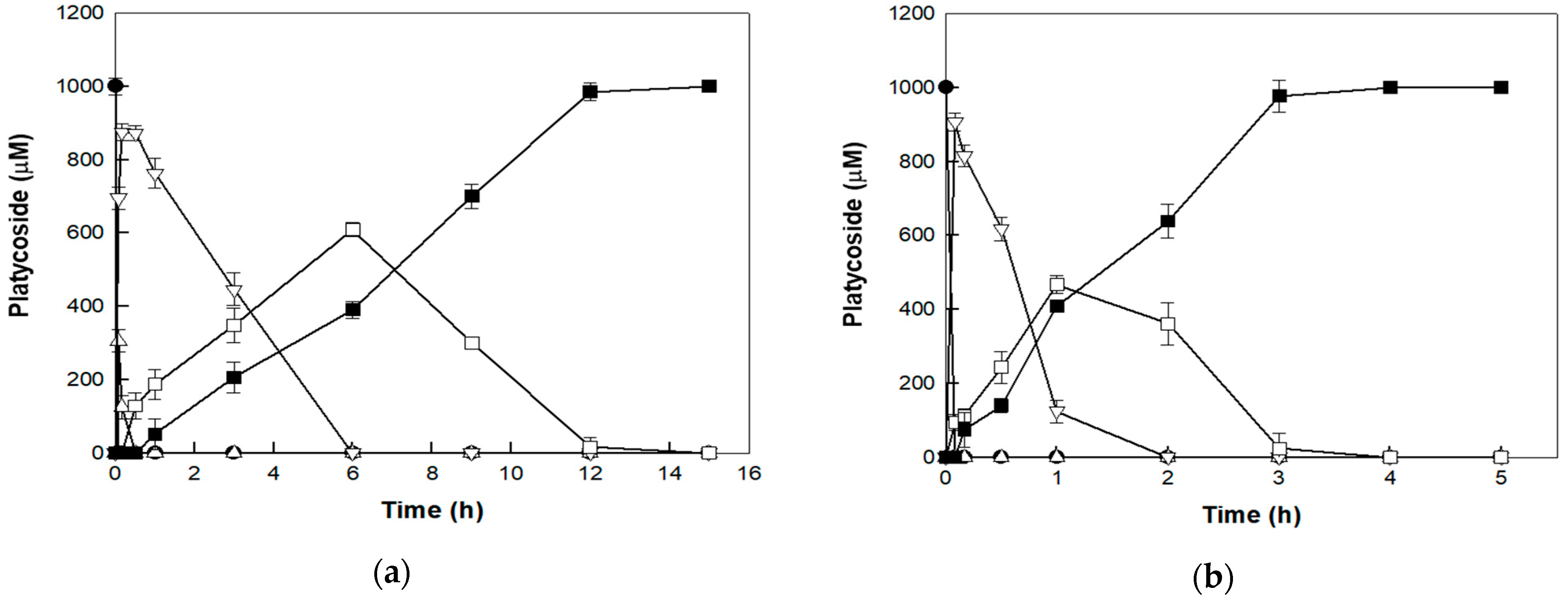
3.4. Bioconversion of Platycoside E to Deapiose-Xylosylated Platycodin D under AP and HHP
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lee, E.B. Pharmacological studies on Platycodon grandiflorum A. DC. IV. A comparison of experimental pharmacological effects of crude platycodin with clinical indications of platycodi radix. Yakugaku Zasshi 1973, 93, 1188–1194. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Song, N.; Liu, Y.; Guo, J. Platycodin D inhibits beta-amyloid-induced inflammation and oxidative stress in BV-2 cells via suppressing TLR4/NF-kappaB signaling pathway and activating Nrf2/HO-1 signaling pathway. Neurochem. Res. 2021, 46, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Han, E.H.; Lim, M.K.; Lee, S.H.; Yu, H.J.; Lim, Y.H.; Kang, S. Fermented Platycodon grandiflorum extracts relieve airway inflammation and cough reflex sensitivity in vivo. J. Med. Food 2020, 23, 1060–1069. [Google Scholar] [CrossRef]
- Ryu, C.S.; Kim, C.H.; Lee, S.Y.; Lee, K.S.; Choung, K.J.; Song, G.Y.; Kim, B.H.; Ryu, S.Y.; Lee, H.S.; Kim, S.K. Evaluation of the total oxidant scavenging capacity of saponins isolated from Platycodon grandiflorum. Food Chem. 2012, 132, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Lin, S.; Ren, F.; Wu, L.; Chen, L.; Sun, Y. Antioxidant and antimicrobial capacity of Chinese medicinal herb extracts in raw sheep meat. J. Food Prot. 2007, 70, 1440–1445. [Google Scholar] [CrossRef] [PubMed]
- Yim, N.H.; Hwang, Y.H.; Liang, C.; Ma, J.Y. A platycoside-rich fraction from the root of Platycodon grandiflorum enhances cell death in A549 human lung carcinoma cells via mainly AMPK/mTOR/AKT signal-mediated autophagy induction. J. Ethnopharmacol. 2016, 194, 1060–1068. [Google Scholar] [CrossRef]
- Li, W.; Tian, Y.H.; Liu, Y.; Wang, Z.; Tang, S.; Zhang, J.; Wang, Y.P. Platycodin D exerts anti-tumor efficacy in H22 tumor-bearing mice via improving immune function and inducing apoptosis. J. Toxicol. Sci. 2016, 41, 417–428. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.L.; Harding, S.V.; Marinangeli, C.P.; Kim, Y.S.; Jones, P.J. Hypocholesterolemic and anti-obesity effects of saponins from Platycodon grandiflorum in hamsters fed atherogenic diets. J. Food Sci. 2008, 73, H195–H200. [Google Scholar] [CrossRef]
- Kim, H.L.; Park, J.; Jung, Y.; Ahn, K.S.; Um, J.Y. Platycodin D, a novel activator of AMP-activated protein kinase, attenuates obesity in db/db mice via regulation of adipogenesis and thermogenesis. Phytomedicine 2019, 52, 254–263. [Google Scholar] [CrossRef]
- Noh, E.M.; Kim, J.M.; Lee, H.Y.; Song, H.K.; Joung, S.O.; Yang, H.J.; Kim, M.J.; Kim, K.S.; Lee, Y.R. Immuno-enhancement effects of Platycodon grandiflorum extracts in splenocytes and a cyclophosphamide-induced immunosuppressed rat model. BMC Complement. Altern. Med. 2019, 19, 322. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Sun, H.X.; Li, D. Platycodin D is a potent adjuvant of specific cellular and humoral immune responses against recombinant hepatitis B antigen. Vaccine 2009, 27, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.C.; Oh, D.K. Classification of glycosidases that hydrolyze the specific positions and types of sugar moieties in ginsenosides. Crit. Rev. Biotechnol. 2016, 36, 1036–1049. [Google Scholar] [CrossRef] [PubMed]
- Park, C.S.; Yoo, M.H.; Noh, K.H.; Oh, D.K. Biotransformation of ginsenosides by hydrolyzing the sugar moieties of ginsenosides using microbial glycosidases. Appl. Microbiol. Biotechnol. 2010, 87, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.Y.; Kim, J.M.; Han, S.B.; Lee, S.K.; Kim, N.D.; Park, M.K.; Kim, C.K.; Park, J.H. Steaming of ginseng at high temperature enhances biological activity. J. Nat. Prod. 2000, 63, 1702–1704. [Google Scholar] [CrossRef]
- Bae, E.A.; Han, M.J.; Kim, E.J.; Kim, D.H. Transformation of ginseng saponins to ginsenoside Rh2 by acids and human intestinal bacteria and biological activities of their transformants. Arch. Pharm. Res. 2004, 27, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.C.; Kim, T.H.; Choi, J.H.; Oh, D.K. Complete biotransformation of protopanaxadiol-type ginsenosides to 20-O-beta-glucopyranosyl-20(S)-protopanaxadiol using a novel and thermostable beta-glucosidase. J. Agric. Food Chem. 2018, 66, 2822–2829. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.J.; You, H.J.; Park, M.S.; Johnston, T.V.; Ku, S.; Ji, G.E. Biocatalysis of platycoside E and platycodin D3 using fungal extracellular β-glucosidase responsible for rapid platycodin D production. Int. J. Mol. Sci. 2018, 19, 2671. [Google Scholar] [CrossRef] [Green Version]
- Kil, T.G.; Kang, S.H.; Kim, T.H.; Shin, K.C.; Oh, D.K. Enzymatic biotransformation of balloon flower root saponins into bioactive platycodin D by deglucosylation with Caldicellulosiruptor bescii β-glucosidase. Int. J. Mol. Sci. 2019, 20, 3854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, K.C.; Seo, M.J.; Kim, D.W.; Yeom, S.J.; Kim, Y.S. Characterization of β-glycosidase from Caldicellulosiruptor owensensis and its application in the production of platycodin D from balloon flower leaf. Catalysts 2019, 9, 1025. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.H.; Kim, T.H.; Shin, K.C.; Ko, Y.J.; Oh, D.K. Biotransformation of food-derived saponins, platycosides, into deglucosylated saponins including deglucosylated platycodin D and their anti-inflammatory activities. J. Agric. Food Chem. 2019, 67, 1470–1477. [Google Scholar] [CrossRef]
- Ha, I.J.; Ha, Y.W.; Kang, M.; Lee, J.; Park, D.; Kim, Y.S. Enzymatic transformation of platycosides and one-step separation of platycodin D by high-speed countercurrent chromatography. J. Sep. Sci. 2010, 33, 1916–1922. [Google Scholar] [CrossRef]
- Ju, J.H.; Kang, S.H.; Kim, T.H.; Shin, K.C.; Oh, D.K. Biotransformation of glycosylated saponins in balloon flower root extract into 3-O-beta-D-glucopyranosyl platycosides by deglycosylation of pectinase from Aspergillus aculeatus. J. Microbiol. Biotechnol. 2020, 30, 946–954. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.C.; Kim, D.W.; Woo, H.S.; Oh, D.K.; Kim, Y.S. Conversion of glycosylated platycoside E to deapiose-xylosylated platycodin D by cytolase PCL5. Int. J. Mol. Sci. 2020, 21, 1207. [Google Scholar] [CrossRef] [PubMed]
- Palaniyandi, S.A.; Damodharan, K.; Lee, K.W.; Yang, S.H.; Suh, J.-W. Enrichment of ginsenoside Rd in Panax ginseng extract with combination of enzyme treatment and high hydrostatic pressure. Biotechnol. Bioprocess Eng. 2015, 20, 608–613. [Google Scholar] [CrossRef]
- Kim, D.Y.; Yeom, S.J.; Park, C.S.; Kim, Y.S. Effect of high hydrostatic pressure treatment on isoquercetin production from rutin by commercial alpha-L-rhamnosidase. Biotechnol. Lett. 2016, 38, 1775–1780. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Specific Activity (nmol/min/mg) | |
|---|---|---|
| AP | HHP | |
| PE | 15,601.2 ± 50.2 | 48,738.2 ± 101.2 |
| PD3 | 281.2 ± 18.0 | 1056.4 ± 31.5 |
| PD | 35.1 ± 1.8 | 141.5 ± 2.1 |
| Deapi-PD | 15.3 ± 1.3 | 71.9 ± 2.5 |
| Deapi-xyl-PD | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, K.-C.; Seo, M.-J.; Oh, Y.J.; Kim, D.W.; Na, C.S.; Kim, Y.-S. Improved Biotransformation of Platycoside E into Deapiose-Xylosylated Platycodin D by Cytolase PCL5 under High Hydrostatic Pressure. Appl. Sci. 2021, 11, 10623. https://doi.org/10.3390/app112210623
Shin K-C, Seo M-J, Oh YJ, Kim DW, Na CS, Kim Y-S. Improved Biotransformation of Platycoside E into Deapiose-Xylosylated Platycodin D by Cytolase PCL5 under High Hydrostatic Pressure. Applied Sciences. 2021; 11(22):10623. https://doi.org/10.3390/app112210623
Chicago/Turabian StyleShin, Kyung-Chul, Min-Ju Seo, Yu Jin Oh, Dae Wook Kim, Chae Sun Na, and Yeong-Su Kim. 2021. "Improved Biotransformation of Platycoside E into Deapiose-Xylosylated Platycodin D by Cytolase PCL5 under High Hydrostatic Pressure" Applied Sciences 11, no. 22: 10623. https://doi.org/10.3390/app112210623
APA StyleShin, K.-C., Seo, M.-J., Oh, Y. J., Kim, D. W., Na, C. S., & Kim, Y.-S. (2021). Improved Biotransformation of Platycoside E into Deapiose-Xylosylated Platycodin D by Cytolase PCL5 under High Hydrostatic Pressure. Applied Sciences, 11(22), 10623. https://doi.org/10.3390/app112210623