
Chemical Investigation of Diketopiperazines and N-Phenethylacetamide Isolated from Aquimarina sp. MC085 and Their Effect on TGF-β-Induced Epithelial–Mesenchymal Transition


 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Experimental Procedure
2.2. Fermentation of Aquimarina sp. MC085 and Preparation of Extracts
2.3. Acid Hydrolysis and C3 Marfey’s Analysis of Compounds 1 and 2
2.4. Cell Culture and Treatment
2.5. Cell Viability Assay
2.6. Western Blotting Analysis
2.7. Gelatin Zymography
2.8. Statistical Analysis
3. Results
3.1. Structural Identification of Compounds 1–3
3.2. The Cell Viability Effect on Comparison of Compounds 1–3 on TGF-β-Induced EMT of A549 Cells
3.3. Comparison of the Protein Expression Profiles of A549 Cells to Determine the Effect of the Treatment with Compounds 1–3 (50 μM), via the Smad2/3 Signaling Pathway, on the TGF-β-Mediated EMT of the Cells
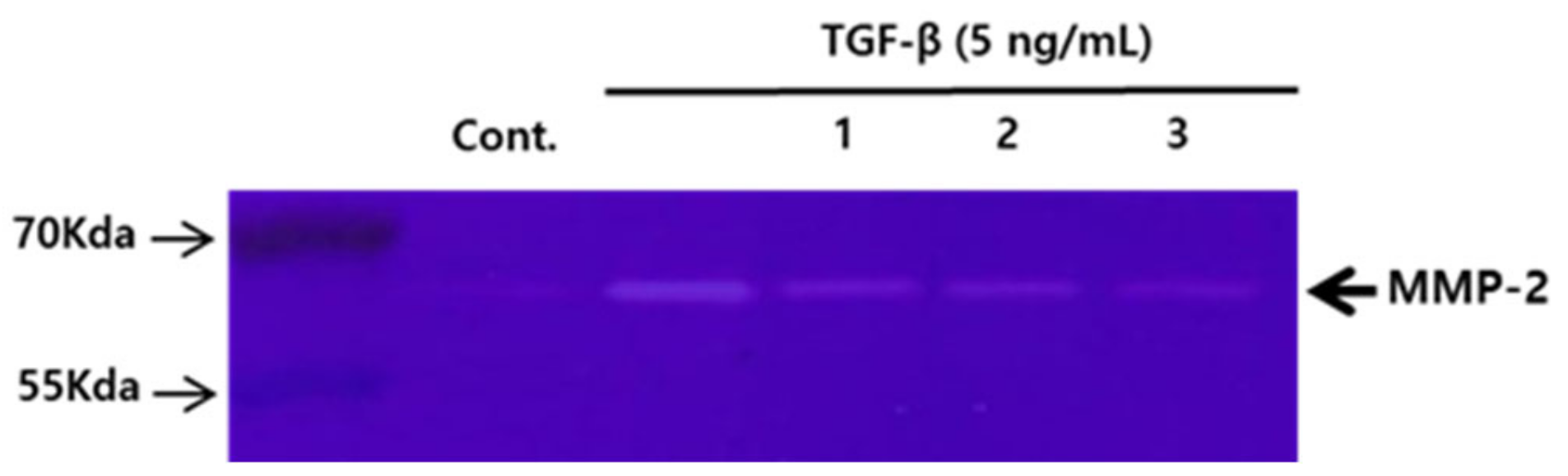
3.4. Comparison of the Effects of the Compounds 1–3 (50 μM) on the Activity of MMP-2 in A549 Human Lung Carcinoma Cells, Tested Using Gelatin Zymography
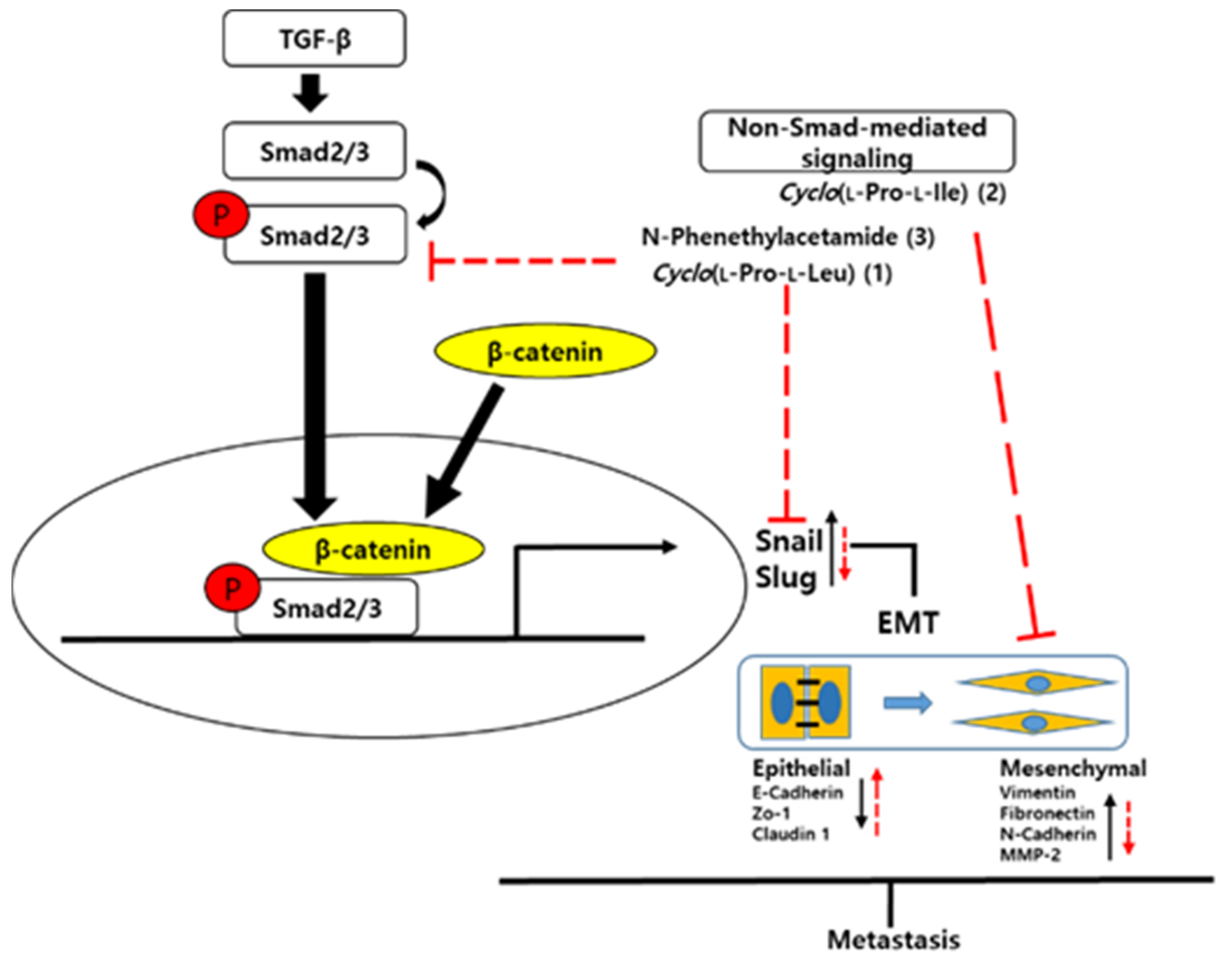
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, W.H.; Moore, B.S. Lessons from the past and charting the future of marine natural products drug discovery and chemical biology. Chem. Biol. 2012, 19, 85–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez, C. Marine Natural Products in Medicinal Chemistry. ACS Med. Chem. Lett. 2018, 9, 959–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Lou, Z.; Lee, S.-H. Arctigenin represses TGF-β-induced epithelial mesenchymal transition in human lung cancer cells. Biochem. Biophys. Res. Commun. 2017, 493, 934–939. [Google Scholar] [CrossRef]
- Sutariya, B.; Saraf, M. Betanin, isolated from fruits of Opuntia elatior Mill attenuates renal fibrosis in diabetic rats through regulating oxidative stress and TGF-β pathway. J. Ethnopharmacol. 2017, 198, 432–443. [Google Scholar] [CrossRef]
- Giacomelli, C.; Daniele, S.; Natali, L.; Iofrida, C.; Flamini, G.; Braca, A.; Trincavelli, M.L.; Martini, C. Carnosol controls the human glioblastoma stemness features through the epithelial-mesenchymal transition modulation and the induction of cancer stem cell apoptosis. Sci. Rep. 2017, 7, 15174. [Google Scholar] [CrossRef]
- Li, W.; Jiang, Z.; Xiao, X.; Wang, Z.; Wu, Z.; Ma, Q.; Cao, L. Curcumin inhibits superoxide dismutase-induced epithelial-to-mesenchymal transition via the PI3K/Akt/NF-κB pathway in pancreatic cancer cells. Int. J. Oncol. 2018, 52, 1593–1602. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, Z.; Yu, G.; Nie, X.; Jia, W.; Liu, R.-E.; Xu, R. Paeoniflorin inhibits migration and invasion of human glioblastoma cells via suppression transforming growth factor β-induced epithelial–mesenchymal transition. Neurochem. Res. 2018, 43, 760–774. [Google Scholar] [CrossRef] [Green Version]
- Pattarayan, D.; Sivanantham, A.; Krishnaswami, V.; Loganathan, L.; Palanichamy, R.; Natesan, S.; Muthusamy, K.; Rajasekaran, S. Tannic acid attenuates TGF-β1-induced epithelial-to-mesenchymal transition by effectively intervening TGF-β signaling in lung epithelial cells. J. Cell. Physiol. 2018, 233, 2513–2525. [Google Scholar] [CrossRef]
- Dybdal-Hargreaves, N.F.; Risinger, A.L.; Mooberry, S.L. Eribulin mesylate: Mechanism of action of a unique microtubule-targeting agent. Clin. Cancer Res. 2015, 21, 2445–2452. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Braun, D.R.; Ananiev, G.E.; Hoffmann, F.M.; Tsai, I.-W.; Rajski, S.R.; Bugni, T.S. Biemamides A–E, inhibitors of the TGF-β pathway that block the epithelial to mesenchymal transition. Org. Lett. 2018, 20, 5529–5532. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Zhang, C.; Liu, F.; Ma, J.; Jia, F.; Han, Z.; Xie, W.; Li, X. Actinomycin V inhibits migration and invasion via suppressing snail/slug-mediated epithelial-mesenchymal transition progression in human breast cancer MDA-MB-231 cells in vitro. Mar. Drugs 2019, 17, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih-Wei, W.; Chih-Ling, C.; Kao, Y.-C.; Martin, R.; Knölker, H.-J.; Shiao, M.-S.; Chen, C.-L. Pentabromopseudilin: A myosin V inhibitor suppresses TGF-β activity by recruiting the type II TGF-β receptor to lysosomal degradation. J. Enzym. Inhib. Med. Chem. 2018, 33, 920–935. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Gamage, C.D.; Kim, G.J.; Hillman, P.F.; Lee, C.; Lee, E.Y.; Choi, H.; Kim, H.; Nam, S.-J.; Fenical, W. Androsamide, a cyclic tetrapeptide from a marine Nocardiopsis sp., suppresses motility of colorectal cancer cells. J. Nat. Prod. 2020, 83, 3166–3172. [Google Scholar] [CrossRef]
- Chen, W.M.; Sheu, F.S.; Sheu, S.Y. Novel L-amino acid oxidase with algicidal activity against toxic cyanobacterium Microcystis aeruginosa synthesized by a bacterium Aquimarina sp. Enzym. Microb. Technol. 2011, 49, 372–379. [Google Scholar] [CrossRef]
- Helfrich, E.J.; Ueoka, R.; Dolev, A.; Rust, M.; Meoded, R.A.; Bhushan, A.; Califano, G.; Costa, R.; Gugger, M.; Steinbeck, C. Automated structure prediction of trans-acyltransferase polyketide synthase products. Nat. Chem. Biol. 2019, 15, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Vijayasarathy, S.; Prasad, P.; Fremlin, L.J.; Ratnayake, R.; Salim, A.A.; Khalil, Z.; Capon, R.J. C3 and 2D C3 Marfey’s methods for amino acid analysis in natural products. J. Nat. Prod. 2016, 79, 421–427. [Google Scholar] [CrossRef] [Green Version]
- Llinarés, J.; Elguero, J.; Faure, R.; Vincent, E.J. Carbon-13 NMR studies of nitrogen compounds. I—substituent effects of amino, acetamido, diacetamido, ammonium and trimethylammonium groups. Org. Magn. Reson. 1980, 14, 20–24. [Google Scholar] [CrossRef]
- Luo, W.; Liu, Q.; Jiang, N.; Li, M.; Shi, L. Isorhamnetin inhibited migration and invasion via suppression of Akt/ERK-mediated epithelial-to-mesenchymal transition (EMT) in A549 human non-small-cell lung cancer cells. Biosci. Rep. 2019, 39, BSR20190159. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.-J.; Li, Q.; Wen, C.-M.; Duan, Z.-X.; Zhang, J.Y.; Xu, C.; Wang, J.-M. Bleomycin (BLM) induces epithelial-to-mesenchymal transition in cultured A549 cells via the TGF-β/Smad signaling pathway. J. Cancer 2016, 7, 1557. [Google Scholar] [CrossRef] [Green Version]
- Agraval, H.; Yadav, U.C. MMP-2 and MMP-9 mediate cigarette smoke extract-induced epithelial-mesenchymal transition in airway epithelial cells via EGFR/Akt/GSK3β/β-catenin pathway: Amelioration by fisetin. Chem.-Biol. Interact. 2019, 314, 108846. [Google Scholar] [CrossRef]
- Rhee, K.-H. Isolation and characterization of Streptomyces sp. KH-614 producing anti-VRE (vancomycin-resistant enterococci) antibiotics. J. Gen. Appl. Microbiol. 2002, 48, 321–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, K.-H. Purification and identification of an antifungal agent from Streptomyces sp. KH-614 antagonistic to rice blast fungus, Pyricularia oryzae. J. Microbiol. Biotechnol. 2003, 13, 984–988. [Google Scholar]
- Li, X.; Dobretsov, S.; Xu, Y.; Xiao, X.; Hung, O.S.; Qian, P.-Y. Antifouling diketopiperazines produced by a deep-sea bacterium, Streptomyces fungicidicus. Biofouling 2006, 22, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Gowrishankar, S.; Kamaladevi, A.; Ayyanar, K.S.; Balamurugan, K.; Pandian, S.K. Bacillus amyloliquefaciens-secreted cyclic dipeptide–cyclo (L-leucyl-L-prolyl) inhibits biofilm and virulence production in methicillin-resistant Staphylococcus aureus. RSC Adv. 2015, 5, 95788–95804. [Google Scholar] [CrossRef]
- Abed, R.M.; Dobretsov, S.; Al-Fori, M.; Gunasekera, S.P.; Sudesh, K.; Paul, V.J. Quorum-sensing inhibitory compounds from extremophilic microorganisms isolated from a hypersaline cyanobacterial mat. J. Ind. Microbiol. Biotechnol. 2013, 40, 759–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Q.; Fu, L.; Wang, Y.; Lin, J. Identification and characterization of extracellular cyclic dipeptides as quorum-sensing signal molecules from Shewanella baltica, the specific spoilage organism of Pseudosciaena crocea during 4 °C storage. J. Agric. Food Chem. 2013, 61, 11645–11652. [Google Scholar] [CrossRef]
- Lin, C.-K.; Wang, Y.-T.; Hung, E.-M.; Yang, Y.-L.; Lee, J.-C.; Sheu, J.-H.; Liaw, C.-C. Butyrolactones and diketopiperazines from marine microbes: Inhibition effects on dengue virus type 2 replication. Planta Med. 2017, 83, 158–163. [Google Scholar] [CrossRef] [Green Version]
- Jinendiran, S.; Teng, W.; Dahms, H.-U.; Liu, W.; Ponnusamy, V.K.; Chiu, C.C.-C.; Kumar, B.D.; Sivakumar, N. Induction of mitochondria-mediated apoptosis and suppression of tumor growth in zebrafish xenograft model by cyclic dipeptides identified from Exiguobacterium acetylicum. Sci. Rep. 2020, 10, 13721. [Google Scholar] [CrossRef]
- Lind, H.; Sjögren, J.; Gohil, S.; Kenne, L.; Schnürer, J.; Broberg, A. Antifungal compounds from cultures of dairy propionibacteria type strains. FEMS Microbiol. Lett. 2007, 271, 310–315. [Google Scholar] [CrossRef] [Green Version]
- Powers, J.C.; Baker, B.L.; Brown, J.; Chelm, B.K. Inhibition of chymotrypsin A. alpha. with N-acyl-and N-peptidyl-2-phenylethylamines. Subsite binding free energies. J. Am. Chem. Soc. 1974, 96, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Bang, I.H.; Noh, Y.J.; Kim, D.K.; Bae, E.J.; Hwang, I.H. Metabolites Produced by the Oral Commensal Bacterium Corynebacterium durum Extend the Lifespan of Caenorhabditis elegans via SIR-2.1 Overexpression. Int. J. Mol. Sci. 2020, 21, 2212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunimoto, S.; Chin-Zhi, X.; Naganawa, H.; Hamada, M.; Masuda, T.; Tajeuchi, T.; Umezawa, H. Reversal of resistance by N-acetyltyramine or N-acetyl-2-phenylethylamine in doxorubicin-resistant leukemia P388 cells. J. Antibiot. 1987, 40, 1651–1652. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.J.; Kim, G.J.; Shin, M.-S.; Moon, J.; Kim, S.; Nam, J.-W.; Kang, K.S.; Choi, H. Chemical Investigation of Diketopiperazines and N-Phenethylacetamide Isolated from Aquimarina sp. MC085 and Their Effect on TGF-β-Induced Epithelial–Mesenchymal Transition. Appl. Sci. 2021, 11, 8866. https://doi.org/10.3390/app11198866
Lee MJ, Kim GJ, Shin M-S, Moon J, Kim S, Nam J-W, Kang KS, Choi H. Chemical Investigation of Diketopiperazines and N-Phenethylacetamide Isolated from Aquimarina sp. MC085 and Their Effect on TGF-β-Induced Epithelial–Mesenchymal Transition. Applied Sciences. 2021; 11(19):8866. https://doi.org/10.3390/app11198866
Chicago/Turabian StyleLee, Myong Jin, Geum Jin Kim, Myoung-Sook Shin, Jimin Moon, Sungjin Kim, Joo-Won Nam, Ki Sung Kang, and Hyukjae Choi. 2021. "Chemical Investigation of Diketopiperazines and N-Phenethylacetamide Isolated from Aquimarina sp. MC085 and Their Effect on TGF-β-Induced Epithelial–Mesenchymal Transition" Applied Sciences 11, no. 19: 8866. https://doi.org/10.3390/app11198866
APA StyleLee, M. J., Kim, G. J., Shin, M.-S., Moon, J., Kim, S., Nam, J.-W., Kang, K. S., & Choi, H. (2021). Chemical Investigation of Diketopiperazines and N-Phenethylacetamide Isolated from Aquimarina sp. MC085 and Their Effect on TGF-β-Induced Epithelial–Mesenchymal Transition. Applied Sciences, 11(19), 8866. https://doi.org/10.3390/app11198866