
The Efficacy of Functional Composts Manufactured Using Spent Coffee Ground, Rice Bran, Biochar, and Functional Microorganisms

Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Selection of Plant Growth-Promoting Bacteria and Plant Pathogen Inhibiting Bacteria, and Their Growth Medium Preparation from Spent Coffee Grounds
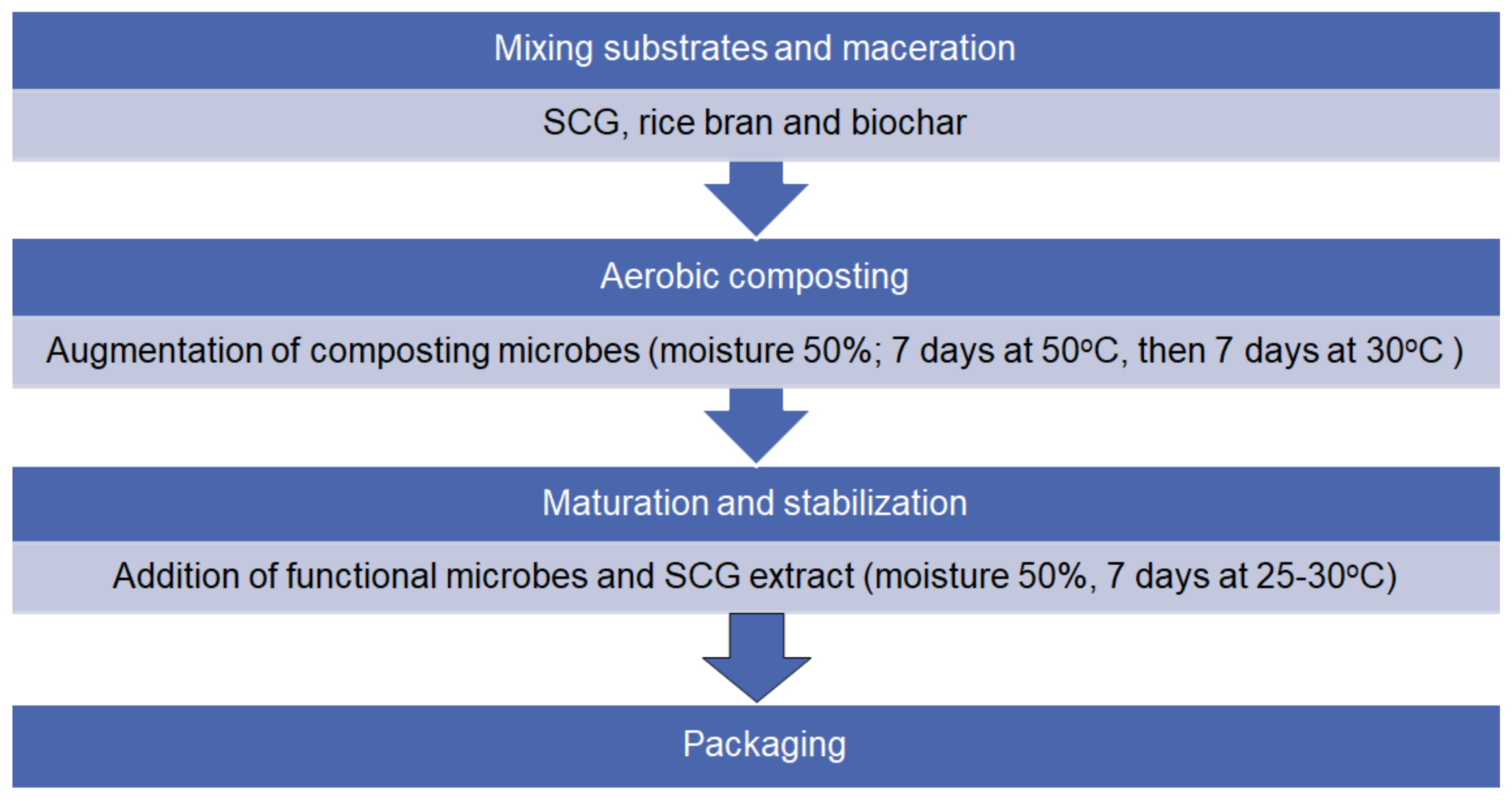
2.2. Procedures for Composting Using Spent Coffee Grounds, Rice Bran, and Biochar, and Phytotoxicity Test for the Manufactured Composts
2.3. Microbial Community Analysis of the Manufactured Complete Composts
2.4. Crop Growth Conditions for Test of the Manufactured Composts
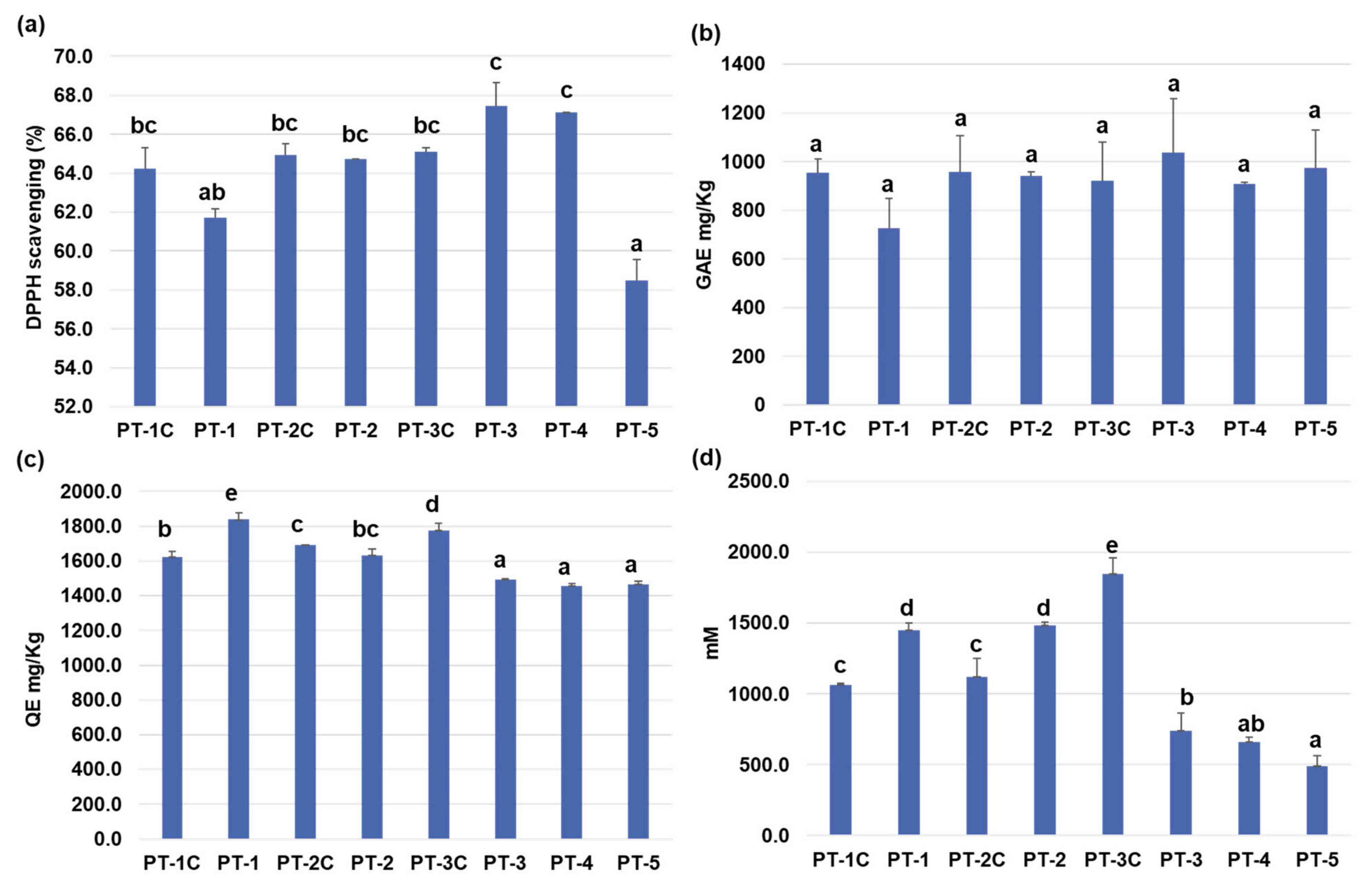
2.5. Analysis of Antioxidants from Leaves of Pepper Plants Grown on the Composts
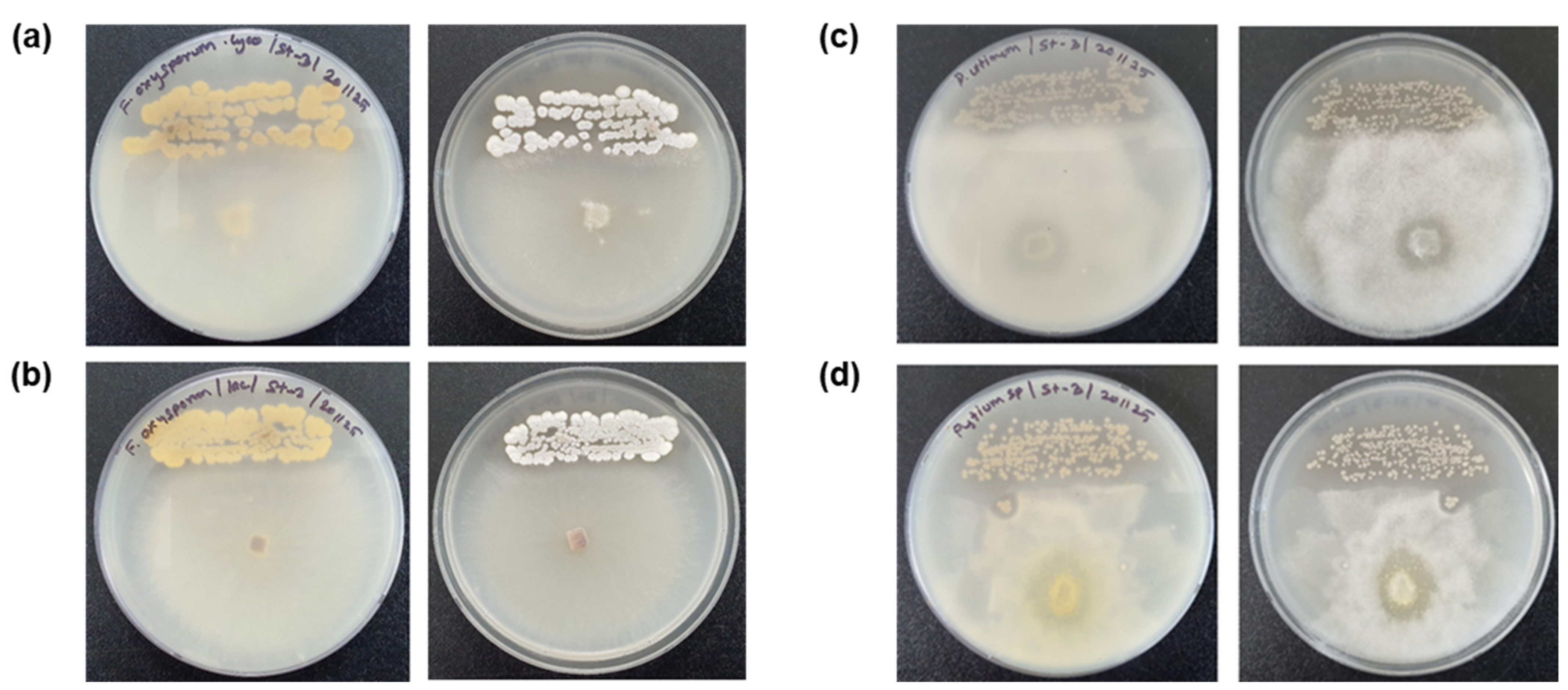
2.6. Inhibition Effects of Streptomyces sasae St-3 on Root Rot Plant Pathogens in Petri Dishes and Pots
2.7. Statistical Analysis
3. Results
3.1. Isolation and Selection of Plant Growth-Promoting Bacteria and Plant Pathogen-Inhibiting Bacteria, and Their Growth Medium Preparation from Spent Coffee Ground
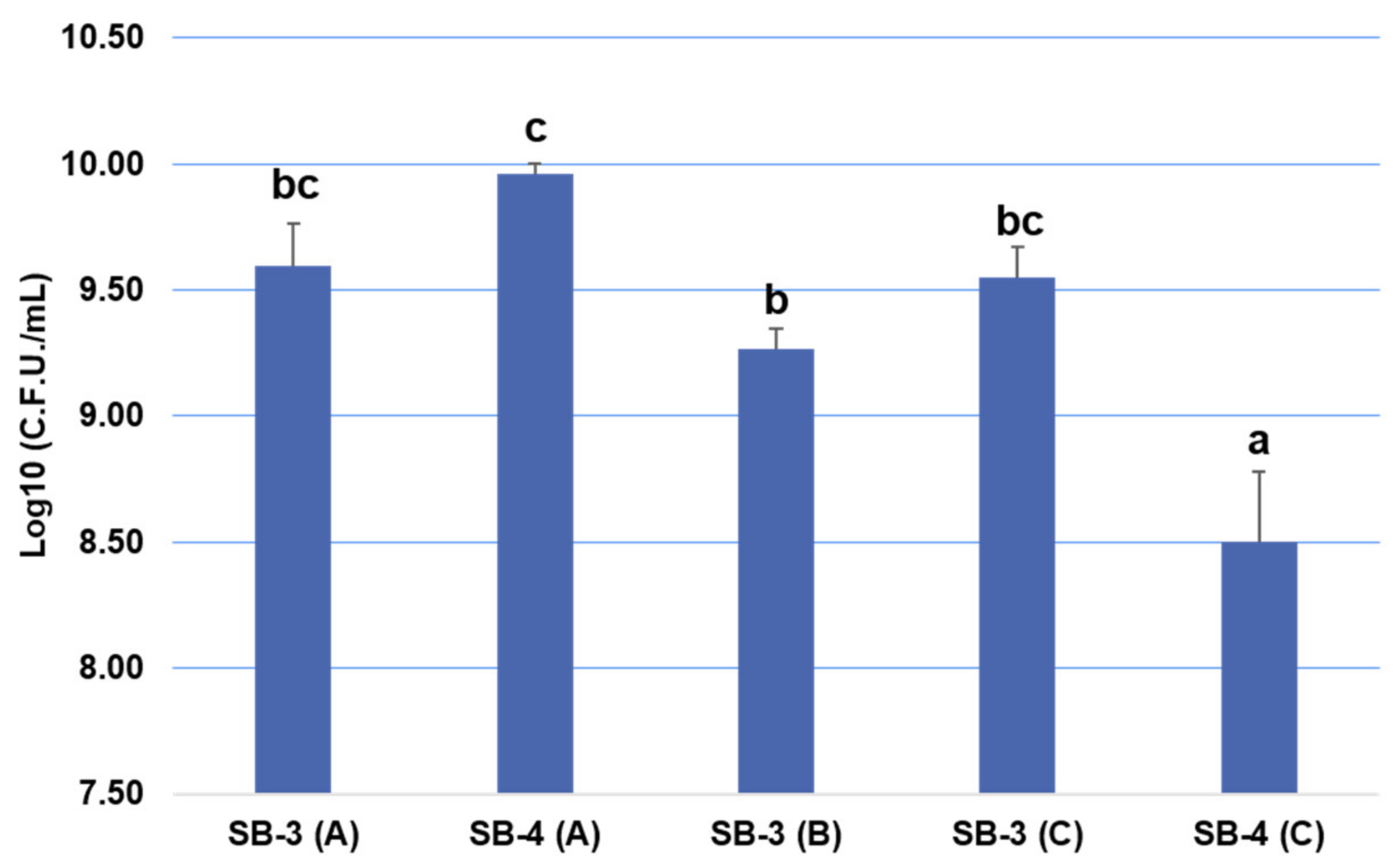
3.2. Impacts of the Various SCG Extracts on the Growth of Potential Plant Growth-Promoting Bacteria
3.3. Composting Process Monitoring, Physico-Chemical Analysis, and Maturity Quality Test of the Manufactured Composts
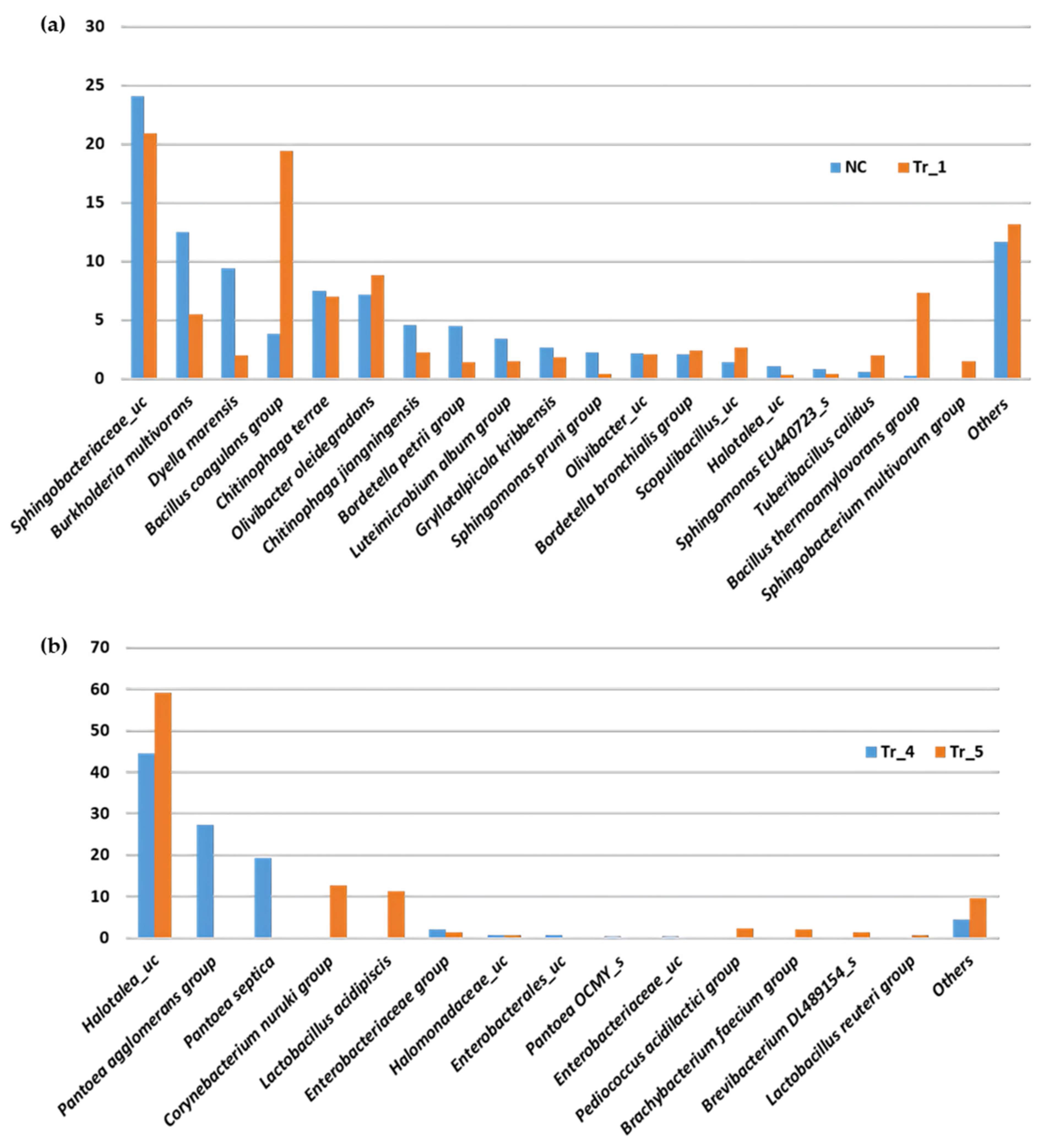
3.4. Microbial Community Analysis of the Manufactured Composts
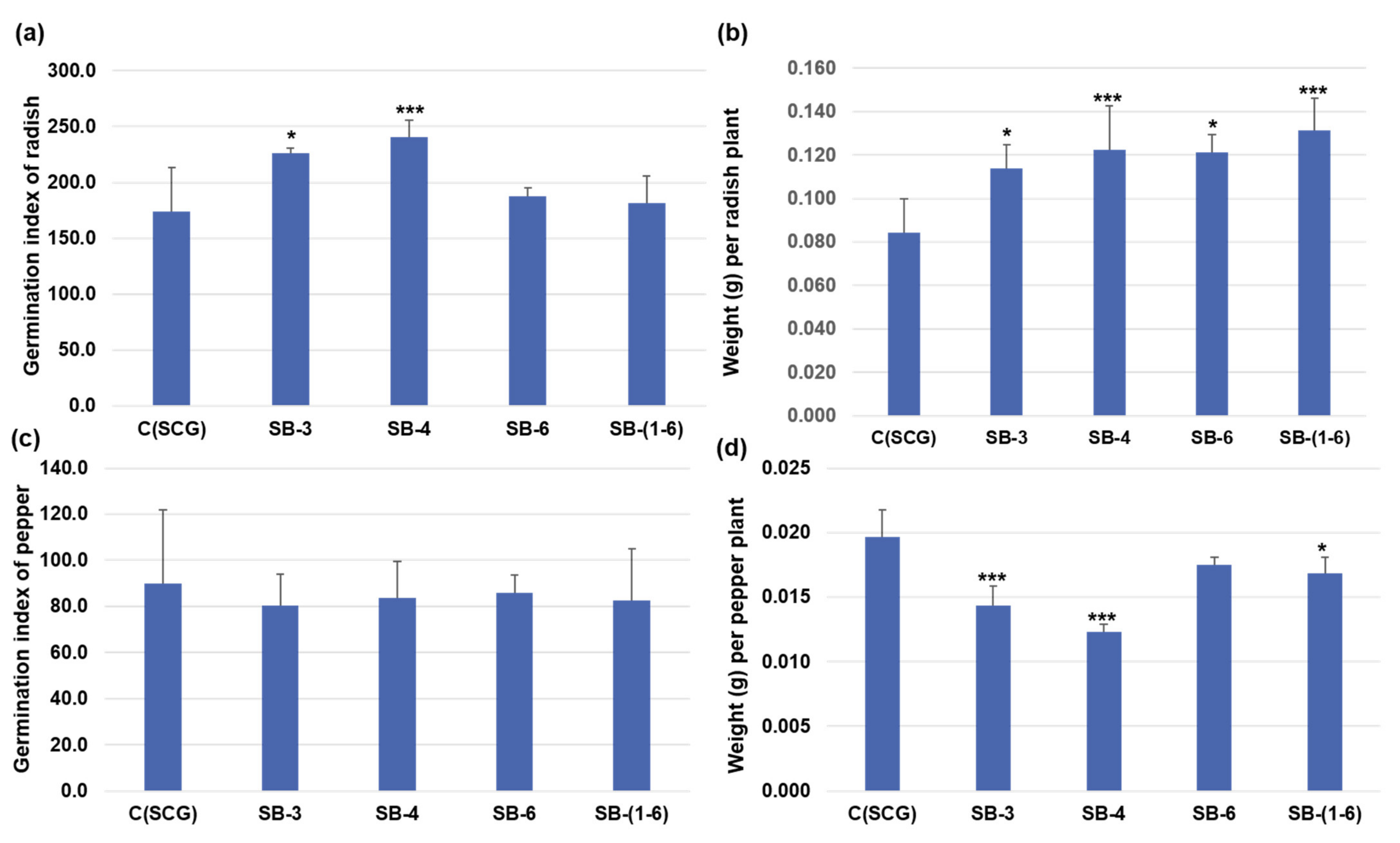
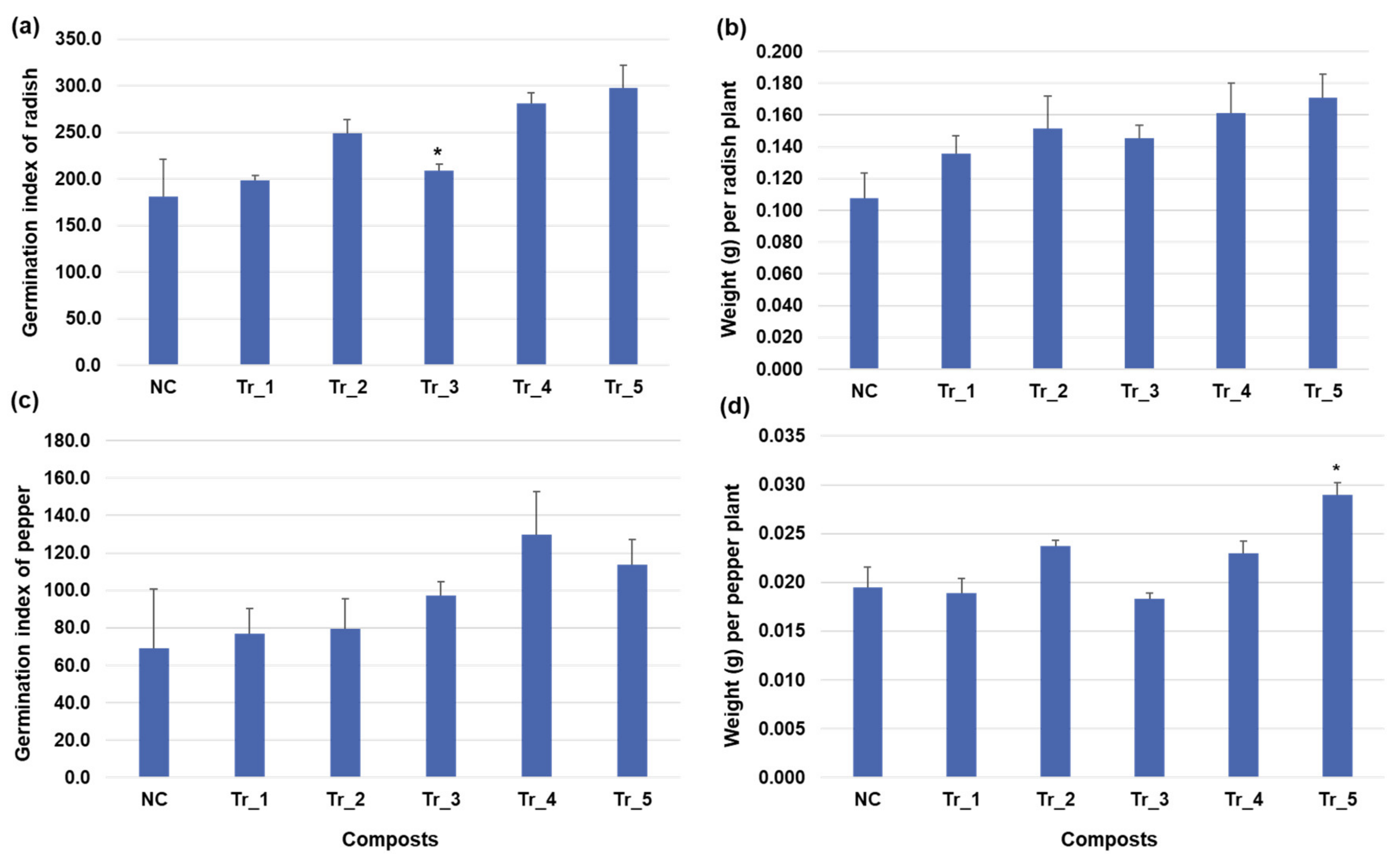
3.5. Analysis of Pepper Growth and Antioxidant Production by Treatment with Functional Composts in Pot Experiments
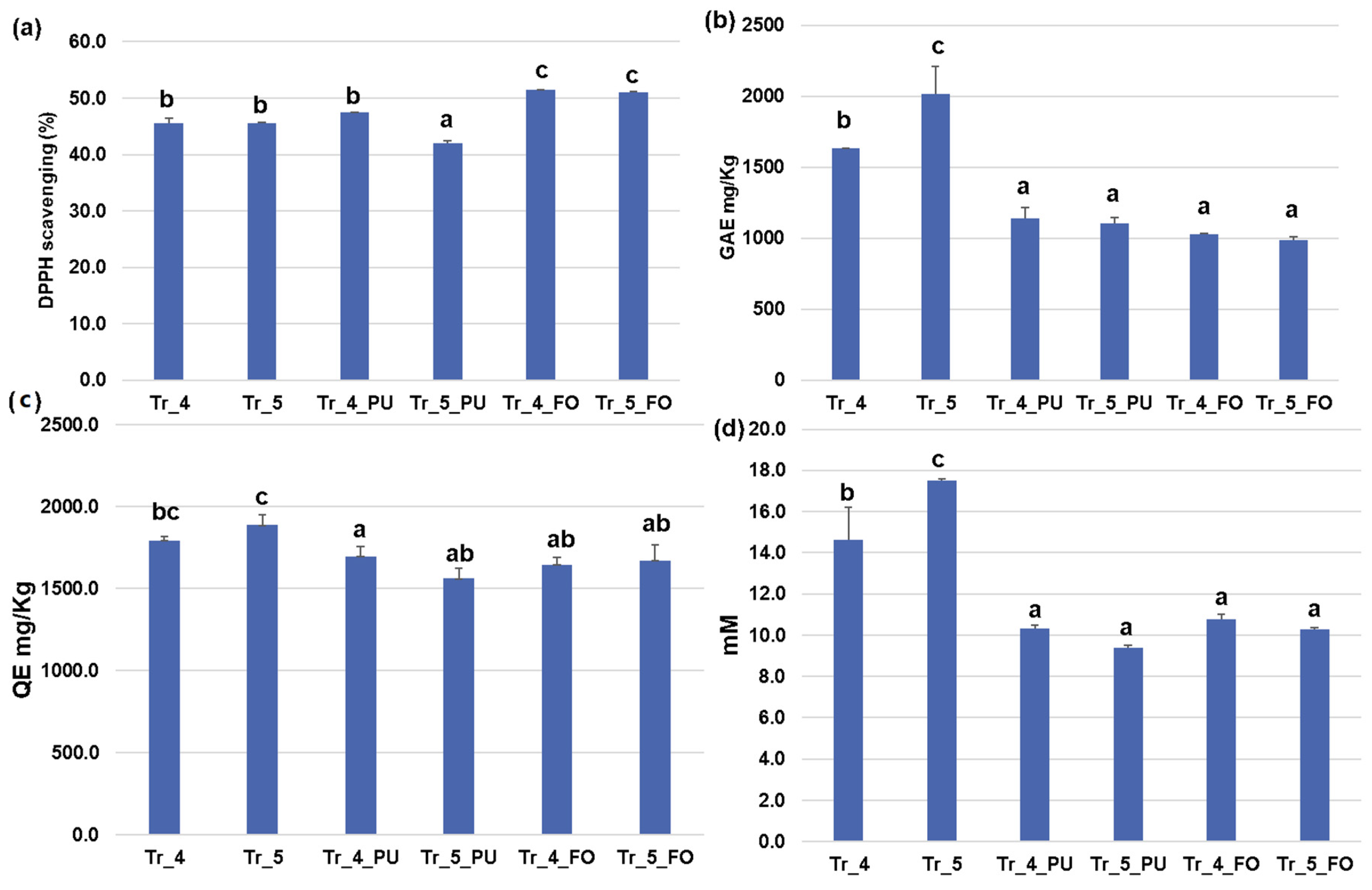
3.6. Inhibition Effect of Streptomyces Sasae St-3 on Plant Pathogens during Radish Germination in the Presence of Functional Compost Extract
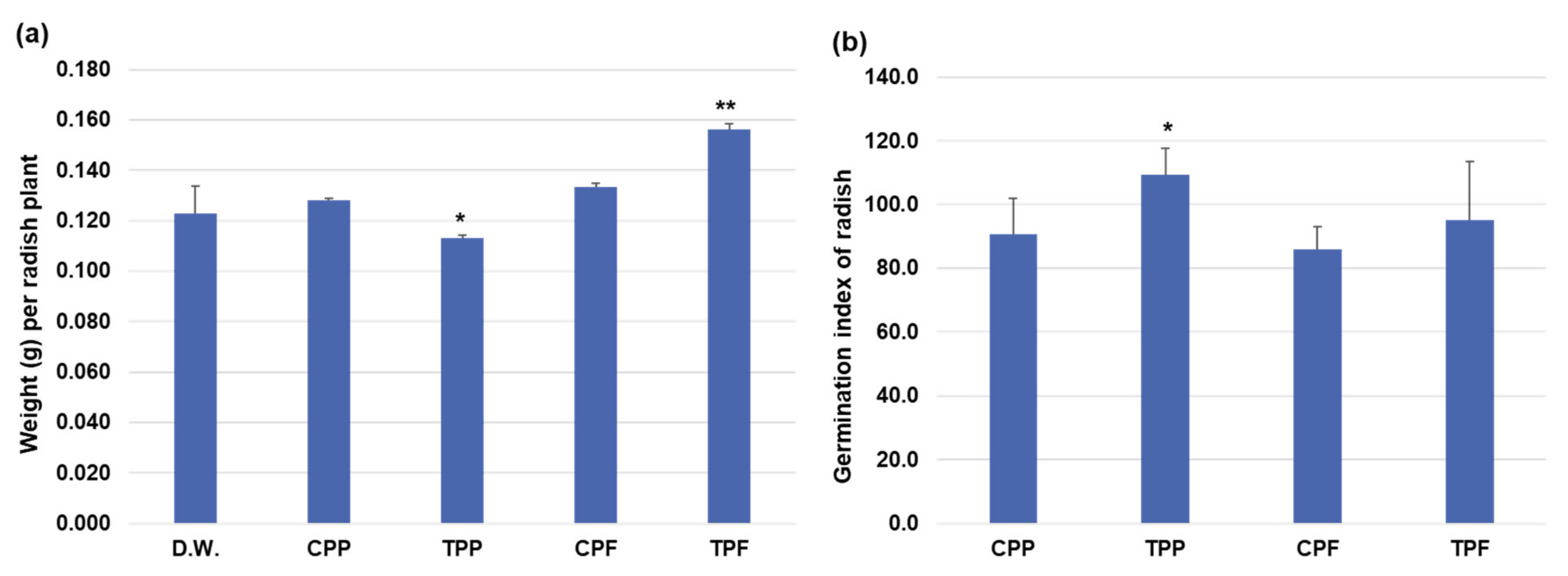
3.7. Effects of the Potential Plant Growth-Promoting Bacteria and Plant Pathogen-Inhibiting Bacterium on Pepper Plants Infected with Root Rot Plant Pathogens
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Evanylo, G.; Sherony, C.; Spargo, J.; Starner, D.; Brosius, M.; Haering, K. Soil and water environmental effects of fertilizer-, manure-, and compost-based fertility practices in an organic vegetable cropping system. Agric. Ecosyst. Environ. 2008, 127, 50–58. [Google Scholar] [CrossRef]
- Lakhdar, A.; Scelza, R.; ben Achiba, W.; Scotti, R.; Rao, M.A.; Jedidi, N.; Abdelly, C.; Gianfreda, L. Effect of municipal solid waste compost and sewage sludge on enzymatic activities and wheat yield in a clayey-loamy soil. Soil Sci. 2011, 176, 15–21. [Google Scholar] [CrossRef]
- Zhang, M.; He, L.-Y.; Liu, Y.-S.; Zhao, J.-L.; Zhang, J.-N.; Chen, J.; Zhang, Q.-Q.; Ying, G.-G. Variation of antibiotic resistome during commercial livestock manure composting. Environ. Int. 2020, 136, 105458. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Rossi, M.J.; Glick, B.R. Ethylene and 1-Aminocyclopropane-1-carboxylate (ACC) in Plant–Bacterial Interactions. Front. Plant Sci. 2018, 9, 114. [Google Scholar] [CrossRef]
- Silva, C.F.; Azevedo, R.S.; Braga, C.; Da Silva, R.; Dias, E.S.; Schwan, R.F. Microbial diversity in a bagasse-based compost prepared for the production of Agaricus brasiliensis. Braz. J. Microbiol. 2009, 40, 590–600. [Google Scholar] [CrossRef][Green Version]
- Tucker, C.M. Coffee Culture, 2nd ed.; Anthropology of Stuff Series; Routledge: New York, NY, USA, 2018. [Google Scholar]
- Getachew, A.T.; Chun, B.S. Influence of pretreatment and modifiers on subcritical water liquefaction of spent coffee grounds: A green waste valorization approach. J. Clean. Prod. 2017, 142, 3719–3727. [Google Scholar] [CrossRef]
- Cruz, R.; Mendes, E.; Torrinha, Á.; Morais, S.; Pereira, J.A.; Baptista, P.; Casal, S. Revalorization of spent coffee residues by a direct agronomic approach. Food Res. Int. 2015, 73, 190–196. [Google Scholar] [CrossRef]
- Cruz, R.; Gomes, T.; Ferreira, A.; Mendes, E.; Baptista, P.; Cunha, S.; Pereira, J.A.; Ramalhosa, E.; Casal, S. Antioxidant activity and bioactive compounds of lettuce improved by espresso coffee residues. Food Chem. 2014, 145, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Price, G.W. Evaluation of three composting systems for the management of spent coffee grounds. Bioresour. Technol. 2011, 102, 7966–7974. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.P.; Vicente, E.; Gomes, A.P.D.; Nunes, M.I.; Alves, C.; Tarelho, L.A.C. Effect of industrial and domestic ash from biomass combustion, and spent coffee grounds, on soil fertility and plant growth: Experiments at field conditions. Environ. Sci. Pollut. Res. 2017, 24, 15270–15277. [Google Scholar] [CrossRef]
- Hardgrove, S.J.; Livesley, S.J. Applying spent coffee grounds directly to urban agriculture soils greatly reduces plant growth. Urban For. Urban Green. 2016, 18, 1–8. [Google Scholar] [CrossRef]
- Gomes, T.; Pereira, J.; Ramalhosa, E.; Casal, S.; Baptista, P. Effect of Fresh and Composted Spent Coffee Grounds on Lettuce Growth, Photosynthetic Pigments and Mineral Composition. VII Congreso Ibérico de Agroingeniería y Ciencias Horticolas; SECH e SEAgIng. Available online: http://hdl.handle.net/10198/8719 (accessed on 21 June 2021).
- Ros, M.; Pascual, J.; Garcia, C.; Hernandez, T.; Insam, H. Hydrolase activities, microbial biomass and bacterial community in a soil after long-term amendment with different composts. Soil Biol. Biochem. 2006, 38, 3443–3452. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Carneiro, L.M.; Silva, J.P.A.; Roberto, I.C.; Teixeira, J.A. A study on chemical constituents and sugars extraction from spent coffee grounds. Carbohydr. Polym. 2011, 83, 368–374. [Google Scholar] [CrossRef]
- Kondamudi, N.; Mohapatra, S.K.; Misra, M. Spent coffee grounds as a versatile source of green energy. J. Agric. Food Chem. 2008, 56, 11757–11760. [Google Scholar] [CrossRef] [PubMed]
- Cruz, R.; Morais, S.; Mendes, E.; Pereira, J.A.; Baptista, P.; Casal, S. Improvement of vegetables elemental quality by espresso coffee residues. Food Chem. 2014, 148, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Zuorro, A.; Lavecchia, R. Spent coffee grounds as a valuable source of phenolic compounds and bioenergy. J. Clean. Prod. 2012, 34, 49–56. [Google Scholar] [CrossRef]
- Budihardjo, M.A.; Sutrisno, E.; Fatimah, M. Leaves composting process and the influence of rumen content and bran addition. MATEC Web Conf. 2018, 159, 2033. [Google Scholar] [CrossRef][Green Version]
- Chang, J.I.; Chen, Y. Effects of bulking agents on food waste composting. Bioresour. Technol. 2010, 101, 5917–5924. [Google Scholar] [CrossRef] [PubMed]
- Moongngarm, A.; Daomukda, N.; Khumpika, S. Chemical compositions, phytochemicals, and antioxidant capacity of rice bran, rice bran layer, and rice germ. APCBEE Procedia 2012, 2, 73–79. [Google Scholar] [CrossRef]
- Khan, M.A.I.; Ueno, K.; Horimoto, S.; Komai, F.; Tanaka, K.; Ono, Y. Evaluation of the use of rice bran compost for eco-friendly weed control in organic farming systems. Am. J. Environ. Sci. 2007, 3, 235–240. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, X.; Xing, G.; Yang, L. Large-scale biochar production from crop residue: A new idea and the biogas-energy pyrolysis system. BioResources 2012, 8, 8–11. [Google Scholar] [CrossRef]
- Sanchez-Monedero, M.; Cayuela, M.L.; Roig, A.; Jindo, K.; Mondini, C.; Bolan, N. Role of biochar as an additive in organic waste composting. Bioresour. Technol. 2018, 247, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Malinowski, M.; Wolny-Koładka, K.; Vaverková, M. Effect of biochar addition on the OFMSW composting process under real conditions. Waste Manag. 2019, 84, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Mierzwa-Hersztek, M.; Wolny-Koładka, K.; Gondek, K.; Gałązka, A.; Gawryjołek, K. Effect of coapplication of biochar and nutrients on microbiocenotic composition, dehydrogenase activity index and chemical properties of sandy soil. Waste Biomass Valorization 2020, 11, 3911–3923. [Google Scholar] [CrossRef]
- Deportes, I.; Guyod, B.; Zmirou, D.; Bouvier, M.C. Microbial disinfection capacity of municipal solid waste (MSW) composting. J. Appl. Microbiol. 1998, 85, 238–246. [Google Scholar] [CrossRef]
- Fatunla, K.; Inam, E.; Essien, J.; Dan, E.; Odon, A.; Kang, S.; Semple, K.T. Influence of composting and thermal processing on the survival of microbial pathogens and nutritional status of Nigeria sewage sludge. Int. J. Recycl. Org. Waste Agric. 2017, 6, 301–310. [Google Scholar] [CrossRef]
- Li, K.; Cao, R.; Mo, S.; Yao, R.; Ren, Z.; Wu, J. Swine manure composting with compound microbial inoculants: Removal of antibiotic resistance genes and their associations with microbial community. Front. Microbiol. 2020, 11, 2686. [Google Scholar] [CrossRef]
- Danish, S.; Zafar-Ul-Hye, M.; Mohsin, F.; Hussain, M. ACC-deaminase producing plant growth promoting rhizobacteria and biochar mitigate adverse effects of drought stress on maize growth. PLoS ONE 2020, 15, e0230615. [Google Scholar] [CrossRef]
- Danish, S.; Zafar-Ul-Hye, M. Combined role of ACC deaminase producing bacteria and biochar on cereals productivity under drought. Phyton 2020, 89, 217–227. [Google Scholar] [CrossRef]
- Emmanuel, S.A.; Yoo, J.; Kim, E.-J.; Chang, J.-S.; Park, Y.-I.; Koh, S.-C. Development of functional composts using spent coffee grounds, poultry manure and biochar through microbial bioaugmentation. J. Environ. Sci. Health Part B 2017, 52, 802–811. [Google Scholar] [CrossRef]
- Kim, I.-S.; Ekpeghere, K.I.; Ha, S.-Y.; Kim, B.-S.; Song, B.; Kim, J.-T.; Kim, H.-G.; Koh, S.-C. Full-scale biological treatment of tannery wastewater using the novel microbial consortium BM-S-1. J. Environ. Sci. Health Part A 2014, 49, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.-C.; Jang, H.-D.; Lin, W.-D.; Duan, P.-F. Antioxidant and antimicrobial activities of commercial rice wine extracts of Taiwanese Allium fistulosum. Food Chem. 2016, 190, 724–729. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-S.; Jung, S.-T.; Kang, S.-G.; Heo, B.G.; Arancibia-Avila, P.; Toledo-Montiel, F.; Drzewiecki, J.; Namiesnik, J.; Gorinstein, S. Antioxidants and proteins in ethylene-treated kiwifruits. Food Chem. 2008, 107, 640–648. [Google Scholar] [CrossRef]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Byrne, D.H. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Jothy, S.L.; Zakaria, Z.; Chen, Y.; Lau, Y.L.; Latha, L.Y.; Sasidharan, S. Acute oral toxicity of methanolic seed extract of cassia fistula in mice. Molecules 2011, 16, 5268–5282. [Google Scholar] [CrossRef]
- Gu, W.; Zhang, F.; Xu, P.; Tang, S.; Xie, K.; Huang, X.; Huang, Q. Effects of sulphur and Thiobacillus thioparus on cow manure aerobic composting. Bioresour. Technol. 2011, 102, 6529–6535. [Google Scholar] [CrossRef]
- Stefan, V.; Van Herpen, E.; Tudoran, A.A.; Lähteenmäki, L. Avoiding food waste by Romanian consumers: The importance of planning and shopping routines. Food Qual. Prefer. 2013, 28, 375–381. [Google Scholar] [CrossRef]
- Chitra, S.; Bargavi, P.; Balasubramaniam, M.; Chandran, R.R.; Balakumar, S. Impact of copper on in-vitro biomineralization, drug release efficacy and antimicrobial properties of bioactive glasses. Mater. Sci. Eng. C 2020, 109, 110598. [Google Scholar] [CrossRef]
- Bidondo, L.F.; Pergola, M.; Silvani, V.; Colombo, R.; Bompadre, J.; Godeas, A. Continuous and long-term monoxenic culture of the arbuscular mycorrhizal fungus Gigaspora decipiens in root organ culture. Fungal Biol. 2012, 116, 729–735. [Google Scholar] [CrossRef]
- Kang, S.-M.; Radhakrishnan, R.; Lee, I.-J. Bacillus amyloliquefaciens subsp. Plantarum GR53, a potent biocontrol agent resists Rhizoctonia disease on Chinese cabbage through hormonal and antioxidants regulation. World J. Microbiol. Biotechnol. 2015, 31, 1517–1527. [Google Scholar] [CrossRef]
- Kuan, K.B.; Othman, R.; Rahim, K.A.; Shamsuddin, Z.H. Plant growth-promoting rhizobacteria inoculation to enhance vegetative growth, nitrogen fixation and nitrogen remobilisation of maize under greenhouse conditions. PLoS ONE 2016, 11, e0152478. [Google Scholar] [CrossRef]
- Theodorou, M.E.; Plaxton, W. Metabolic adaptations of plant respiration to nutritional phosphate deprivation. Plant Physiol. 1993, 101, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.-M.; Radhakrishnan, R.; Khan, A.L.; Kim, M.-J.; Park, J.-M.; Kim, B.-R.; Shin, D.-H.; Lee, I.-J. Gibberellin secreting rhizobacterium, Pseudomonas putida H-2-3 modulates the hormonal and stress physiology of soybean to improve the plant growth under saline and drought conditions. Plant Physiol. Biochem. 2014, 84, 115–124. [Google Scholar] [CrossRef]
- Zeng, Q.; Xie, J.; Li, Y.; Gao, T.; Xu, C.; Wang, Q. Comparative genomic and functional analyses of four sequenced Bacillus cereus genomes reveal conservation of genes relevant to plant-growth-promoting traits. Sci. Rep. 2018, 8, 17009. [Google Scholar] [CrossRef] [PubMed]
- Pérez, G.C.; González, C.G.Z. Practices and conceptions of feedback in initial teacher training. Educ. Pesqui. 2019, 45, e192953. [Google Scholar] [CrossRef]
- Rojas-Solis, D.; Vences-Guzmán, M.Á.; Sohlenkamp, C.; Santoyo, G. Antifungal and plant growth–promoting bacillus under saline stress modify their membrane composition. J. Soil Sci. Plant Nutr. 2020, 20, 1549–1559. [Google Scholar] [CrossRef]
- Lyu, A.; Liu, H.; Che, H.; Yang, L.; Zhang, J.; Wu, M.; Chen, W.; Li, G. Reveromycins A and B from Streptomyces sp. 3–10: Antifungal activity against plant pathogenic fungi in vitro and in a strawberry food model system. Front. Microbiol. 2017, 8, 550. [Google Scholar] [CrossRef]
- Getha, K.; Vikineswary, S. Antagonistic effects of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f. sp. cubense race 4: Indirect evidence for the role of antibiosis in the antagonistic process. J. Ind. Microbiol. Biotechnol. 2002, 28, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Al-Askar, A.A.; Baka, Z.A.; Rashad, Y.M.; Ghoneem, K.M.; Abdulkhair, W.M.; Hafez, E.E.; Shabana, Y.M. Evaluation of Streptomyces griseorubens E44G for the biocontrol of Fusarium oxysporum f. sp. lycopersici: Ultrastructural and cytochemical investigations. Ann. Microbiol. 2015, 65, 1815–1824. [Google Scholar] [CrossRef]
- Sudiana, I.M.; Putri, A.; Napitupulu, T.P.; Purnaningsih, I.; Kanti, A. Growth inhibition of Fusarium solani and F. oxysporum by Streptomyces sasae TG01, and its ability to solubilize insoluble phosphate. Biodiversitas J. Biol. Divers. 2020, 21. [Google Scholar] [CrossRef]
- Hassanisaadi, M.; Bonjar, G.S.; Hosseinipour, A.; Abdolshahi, R.; Barka, E.A.; Saadoun, I. Biological control of Pythium aphanidermatum, the causal agent of tomato root rot by two Streptomyces root symbionts. Agronomy 2021, 11, 846. [Google Scholar] [CrossRef]
- Medina, J.J.R.; Ramírez, K.; Rangel-Peraza, J.G.; Bustos-Terrones, Y.A. Increase of content and bioactivity of total phenolic compounds from spent coffee grounds through solid state fermentation by Bacillus clausii. J. Food Sci. Technol. 2018, 55, 915–923. [Google Scholar] [CrossRef]
- González-Moreno, M.; Gracianteparaluceta, B.G.; Sádaba, S.M.; Urdin, J.Z.; Domínguez, E.R.; Ezcurdia, M.P.; Meneses, A.S. Feasibility of vermicomposting of spent coffee grounds and silverskin from coffee industries: A laboratory study. Agronomy 2020, 10, 1125. [Google Scholar] [CrossRef]
- Priftis, A.; Mitsiou, D.; Halabalaki, M.; Ntasi, G.; Stagos, D.; Skaltsounis, L.A.; Kouretas, D. Roasting has a distinct effect on the antimutagenic activity of coffee varieties. Mutat. Res. Toxicol. Environ. Mutagen. 2018, 829–830, 33–42. [Google Scholar] [CrossRef]
- Etesami, H.; Hosseini, H.M.; Alikhani, H.A. Bacterial biosynthesis of 1-aminocyclopropane-1-caboxylate (ACC) deaminase, a useful trait to elongation and endophytic colonization of the roots of rice under constant flooded conditions. Physiol. Mol. Biol. Plants 2014, 20, 425–434. [Google Scholar] [CrossRef]
- Islam, M.; Kasim, S.; Alam, K.; Amin, A.; Hun, T.G.; Haque, M. Changes in chemical properties of banana pseudostem, mushroom media waste, and chicken manure through the co-composting process. Sustainability 2021, 13, 8458. [Google Scholar] [CrossRef]
- Khan, M.A.I.; Ueno, K.; Horimoto, S.; Komai, F.; Tanaka, K.; Ono, Y. Evaluation of the upland weed control potentiality of green tea waste—Rice bran compost and its effect on spinach growth. Am. J. Agric. Biol. Sci. 2007, 2, 142–148. [Google Scholar] [CrossRef][Green Version]
- Mujtaba, G.; Hayat, R.; Hussain, Q.; Ahmed, M. Physio-chemical characterization of biochar, compost and co-composted biochar derived from green waste. Sustainability 2021, 13, 4628. [Google Scholar] [CrossRef]
- Saini, A.; Aggarwal, N.K.; Sharma, A.; Yadav, A. Actinomycetes: A source of lignocellulolytic enzymes. Enzym. Res. 2015, 2015, 279381. [Google Scholar] [CrossRef] [PubMed]
- Tortosa, G.; Castellano-Hinojosa, A.; Correa-Galeote, D.; Bedmar, E.J. Evolution of bacterial diversity during two-phase olive mill waste (“alperujo”) composting by 16S rRNA gene pyrosequencing. Bioresour. Technol. 2017, 224, 101–111. [Google Scholar] [CrossRef]
- Martins, L.F.; Antunes, L.P.; Pascon, R.C.; De Oliveira, J.C.F.; Digiampietri, L.A.; Barbosa, D.; Peixoto, B.M.; Vallim, M.; Viana-Niero, C.; Ostroski, E.H.; et al. Metagenomic analysis of a tropical composting operation at the São Paulo zoo park reveals diversity of biomass degradation functions and organisms. PLoS ONE 2013, 8, e61928. [Google Scholar] [CrossRef]
- Bosma, E.F.; Van De Weijer, A.H.P.; Van Der Vlist, L.; De Vos, W.M.; Van Der Oost, J.; Van Kranenburg, R. Establishment of markerless gene deletion tools in thermophilic Bacillus smithii and construction of multiple mutant strains. Microb. Cell Factories 2015, 14, 99. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, W.; Liu, R.; Li, Q.; Li, B.; Wang, S.; Song, C.; Qiao, C.; Mulchandani, A. Phylogenetic diversity and metabolic potential of activated sludge microbial communities in full-scale wastewater treatment plants. Environ. Sci. Technol. 2011, 45, 7408–7415. [Google Scholar] [CrossRef]
- Pellegrini, M.; Spera, D.; Ercole, C.; Del Gallo, M. Allium cepa L. Inoculation with a consortium of plant growth-promoting bacteria: Effects on plants, soil, and the autochthonous microbial community. Microorganisms 2021, 9, 639. [Google Scholar] [CrossRef]
- Tungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabai, A. Flavonoids and other phenolic compounds from medicinal plants for pharmaceutical and medical aspects: An overview. Medicines 2018, 5, 93. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Gaurav, A.K.; Srivastava, S.; Verma, J.P. Plant growth-promoting bacteria: Biological tools for the mitigation of salinity stress in plants. Front. Microbiol. 2020, 11, 1216. [Google Scholar] [CrossRef]
- Beckers, T.; Burkhardt, C.; Wieland, H.; Gimmnich, P.; Ciossek, T.; Maier, T.; Sanders, K. Distinct pharmacological properties of second generation HDAC inhibitors with the benzamide or hydroxamate head group. Int. J. Cancer 2007, 121, 1138–1148. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Guo, J.; Qu, J.; Lian, J.; Jefferson, W.; Yang, J.; Li, H. Catalyzing denitrification of Paracoccus versutus by immobilized 1,5-dichloroanthraquinone. Biodegradation 2011, 23, 399–405. [Google Scholar] [CrossRef]
- Clark, L. DigitalCommons@University of Nebraska-Lincoln: A Review of Pathogens of Agricultural and Human Health Interest Found in Canada Geese. Available online: https://digitalcommons.unl.edu/icwdm_usdanwrc/205/ (accessed on 21 June 2021).
- Abdel-Monaim, M.F. Improvement of biocontrol of damping-off and root rot/wilt of faba bean by salicylic acid and hydrogen peroxide. Mycobiology 2013, 41, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Palaniyandi, S.A.; Yang, S.H.; Zhang, L.; Suh, J.-W. Effects of actinobacteria on plant disease suppression and growth promotion. Appl. Microbiol. Biotechnol. 2013, 97, 9621–9636. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Unit | Location of Substrates | Mixture of Substrates and Composting Microbes | Inoculation of Functional Microbes | Composting Location | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| SPG | Defatted Rice Bran | Biochar | Microbial Inoculation (MA-1) | Total (kg) | Bacillus sp. (SB-3 and SB-4) | Streptomyces sasae (St-3) | ||||
| NC | % | Lab | 81.5 | 16.0 | 2.5 | 0 | 0 | 0 | Lab | |
| kg | 1.46 | 0.283 | 0.045 | 0 | 1.7 | 0 | 0 | |||
| Tr_1 | % | Lab | 81.5 | 16.0 | 2.5 | 0.2 | 1 | 1 | Lab | |
| kg | 1.46 | 0.283 | 0.045 | 0.004 | 1.7 | 1.7 mL | 1.7 mL | |||
| Tr_2 | % | Factory | 47 | 50 | 3 | 0.2 | 0 | 0 | Lab | |
| kg | 25.85 | 27.25 | 1.65 | 0.11 | 55 | 0 | 0 | |||
| Tr_3 | % | Factory | 47 | 50 | 3 | 0.2 | 1 | 1 | Lab | |
| kg | 25.85 | 27.25 | 1.65 | 0.11 | 55 | 1.5 L | 1.5 L | |||
| Tr_4 | % | Factory | 47 | 50 | 3 | 0.2 | 1 | 1 | Factory | |
| kg | 25.85 | 27.25 | 1.65 | 0.11 | 55 | 1.5 L | 1.5 L | |||
| Tr_5 | % | Factory | 47 | 50 | 3 | 0.2 | 1 | 1 | Factory | |
| kg | 25.85 | 27.25 | 1.65 | 0.11 | 55 | 1.5 L | 1.5 L | |||
| Treatment Code | Kinds of Compost or Fertilizer | SB Cultures (%, w/w) * | St-3 Culture (%, w/w) ** | Compost or Fertilizer Amount (g) |
|---|---|---|---|---|
| PT_1C | NC | 0 | 0 | 24 |
| PT_1 | Tr_1 | 1.0 | 1.0 | 24 |
| PT_2C | Tr_2 | 0 | 0 | 24 |
| PT_2 | Tr_3 | 1.0 | 1.0 | 24 |
| PT_3C | Tr_4 | 0 | 0 | 24 |
| PT_3 | Tr_5 | 1.0 | 1.0 | 24 |
| PT_4 | Commercial compost product | - | - | 9.0 |
| PT_5 | Chemical fertilizer | - | - | 1.54 |
| Treatment Code | Treatment | Compost Extract | Inoculation of Streptomyces sasae * St-3 | Root Rotting Pathogen ** |
|---|---|---|---|---|
| CPP | Control | Tr_4 | − | Pythium ultimum |
| TPP | Treatment | Tr_4 | + | Pythium ultimum |
| CPF | Control | Tr_4 | − | Fusarium oxysporum f. lactucae |
| TPF | Treatment | Tr_4 | + | Fusarium oxysporum f. lactucae |
| Treatment Code | Compost Amended | Inoculation of St-3, SB-3, and SB-4 * | Root Rotting Pathogen ** |
|---|---|---|---|
| Tr_4 | Tr_4 | + | − |
| Tr_5 | Tr_5 | + | − |
| Tr_4_PU | Tr_4 | − | Pythium ultimum |
| Tr_5_PU | Tr_5 | + | Pythium ultimum |
| Tr_4_FO | Tr_4 | − | Fusarium oxysporum f. lactucae |
| Tr_5_FO | Tr_5 | + | Fusarium oxysporum f. lactucae |
| Evaluation Criteria | Unit | Commercial Quality Standard | Tr-1 | Tr-5 |
|---|---|---|---|---|
| Organic matter/N ratio | <45 | 19.68 | 26 | |
| NaCl | % | <2.0 | 0.051 | 0.042 |
| Moisture | % | <55 | 45.08 | 29.08 |
| As | mg kg−1 | <45 | ND | ND |
| Cd | mg kg−1 | <5 | ND | ND |
| Hg | mg kg−1 | <2 | ND | ND |
| Pb | mg kg−1 | <130 | 0.2 | 0.04 |
| Cr | mg kg−1 | <200 | 2.35 | 1.75 |
| Cu | mg kg−1 | <360 | 43.8 | 16.27 |
| Ni | mg kg−1 | <45 | 2.36 | 1.05 |
| Zn | mg kg−1 | <900 | 49.3 | 61.99 |
| Organic matter | % | >30 | 49.6 | 62.68 |
| E. coli O157:H7 | ND | ND | ND | ND |
| Salmonella | ND | ND | ND | ND |
| Maturity | Instrument analysis (CoMMe-100) ** | Complete humification | Complete humification | Complete humification |
| HCl insoluble material | % | < 25 | 0.34 | 0.27 |
| Total N (%) *** | - | 2.52 | 2.41 | |
| Total P (%) *** | - | 1.22 | 3.36 | |
| Total K (%) *** | - | 0.89 | 1.66 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santhanarajan, A.-E.; Han, Y.-H.; Koh, S.-C. The Efficacy of Functional Composts Manufactured Using Spent Coffee Ground, Rice Bran, Biochar, and Functional Microorganisms. Appl. Sci. 2021, 11, 7703. https://doi.org/10.3390/app11167703
Santhanarajan A-E, Han Y-H, Koh S-C. The Efficacy of Functional Composts Manufactured Using Spent Coffee Ground, Rice Bran, Biochar, and Functional Microorganisms. Applied Sciences. 2021; 11(16):7703. https://doi.org/10.3390/app11167703
Chicago/Turabian StyleSanthanarajan, Aalfin-Emmanuel, Yong-Hun Han, and Sung-Cheol Koh. 2021. "The Efficacy of Functional Composts Manufactured Using Spent Coffee Ground, Rice Bran, Biochar, and Functional Microorganisms" Applied Sciences 11, no. 16: 7703. https://doi.org/10.3390/app11167703
APA StyleSanthanarajan, A.-E., Han, Y.-H., & Koh, S.-C. (2021). The Efficacy of Functional Composts Manufactured Using Spent Coffee Ground, Rice Bran, Biochar, and Functional Microorganisms. Applied Sciences, 11(16), 7703. https://doi.org/10.3390/app11167703