Lemon Balm and Dandelion Leaf Extracts Synergistically Protect against Carbon Tetrachloride-Induced Acute Liver Injury in Mice

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of MLD and Reagents
2.2. Animal Husbandry and Treatment
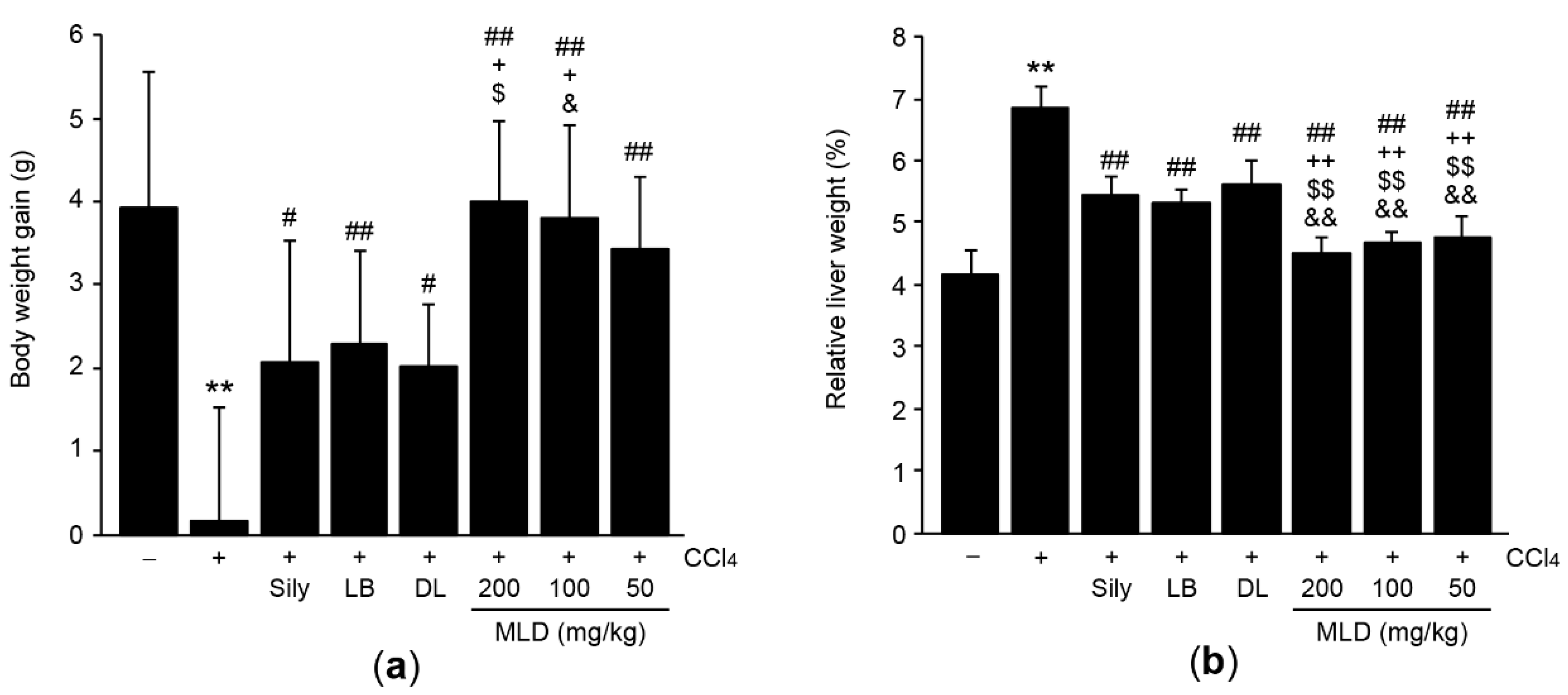
2.3. Body Weight Gain and Relative Liver Weight
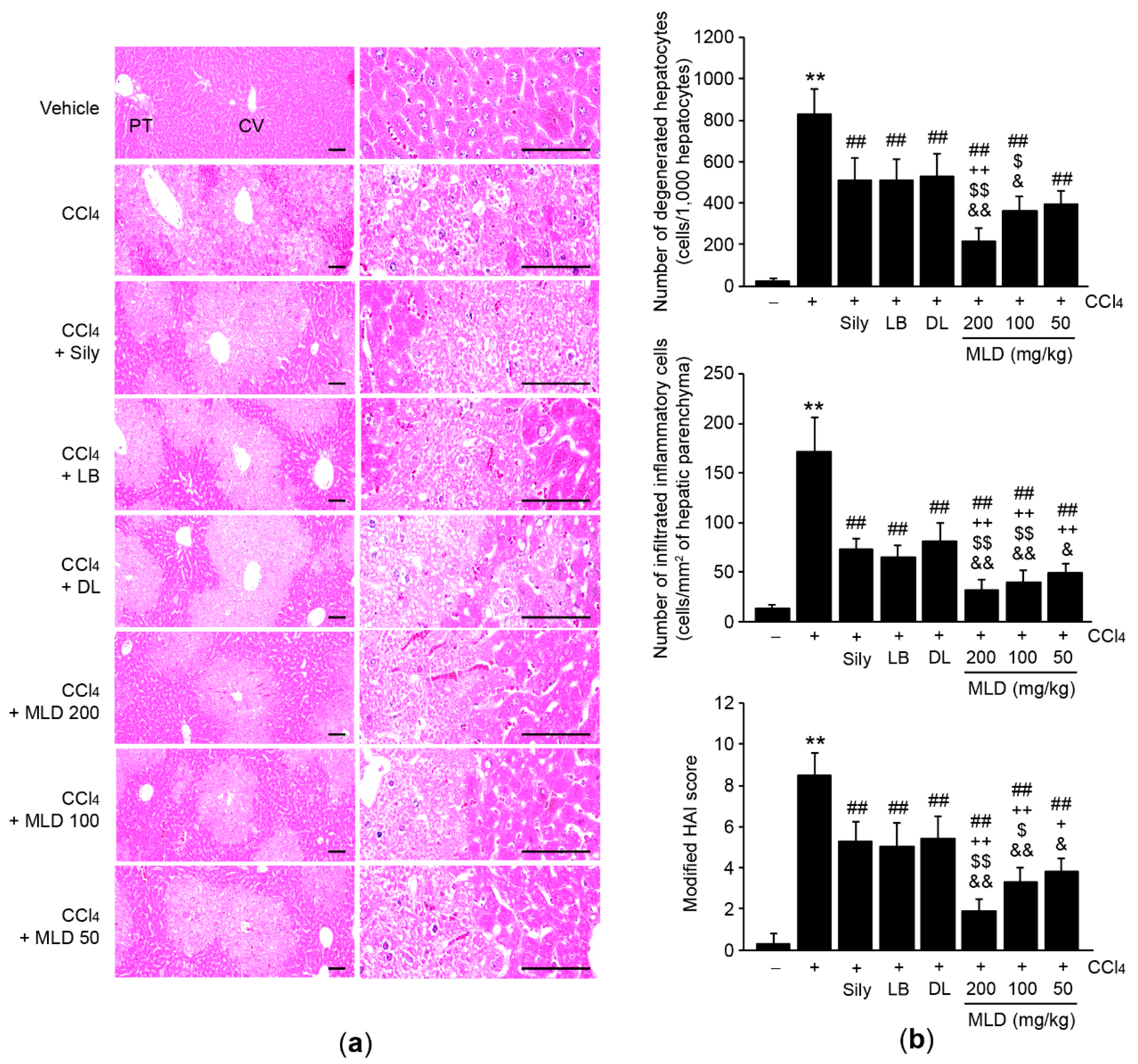
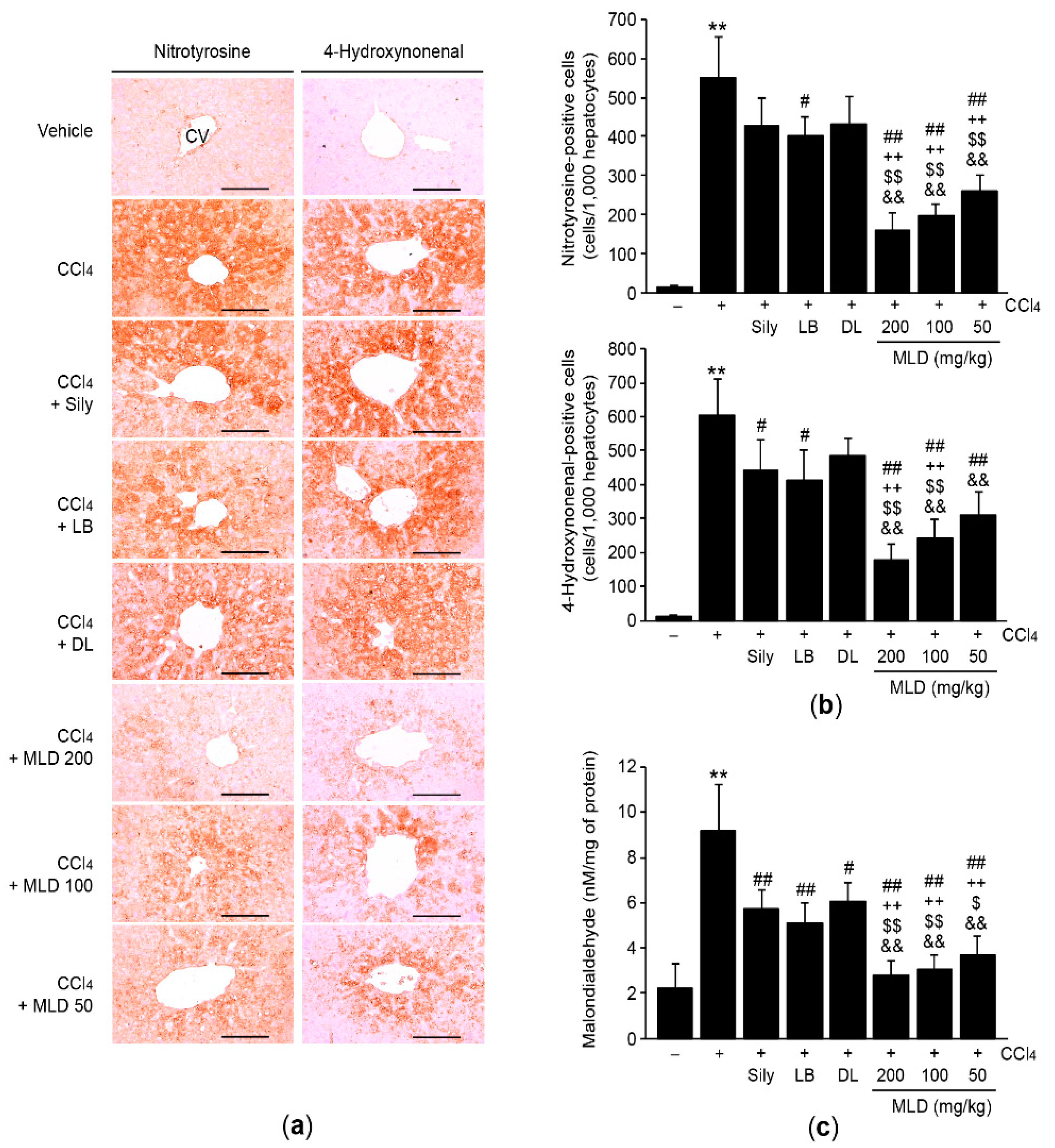
2.4. Histology and Immunohistochemistry
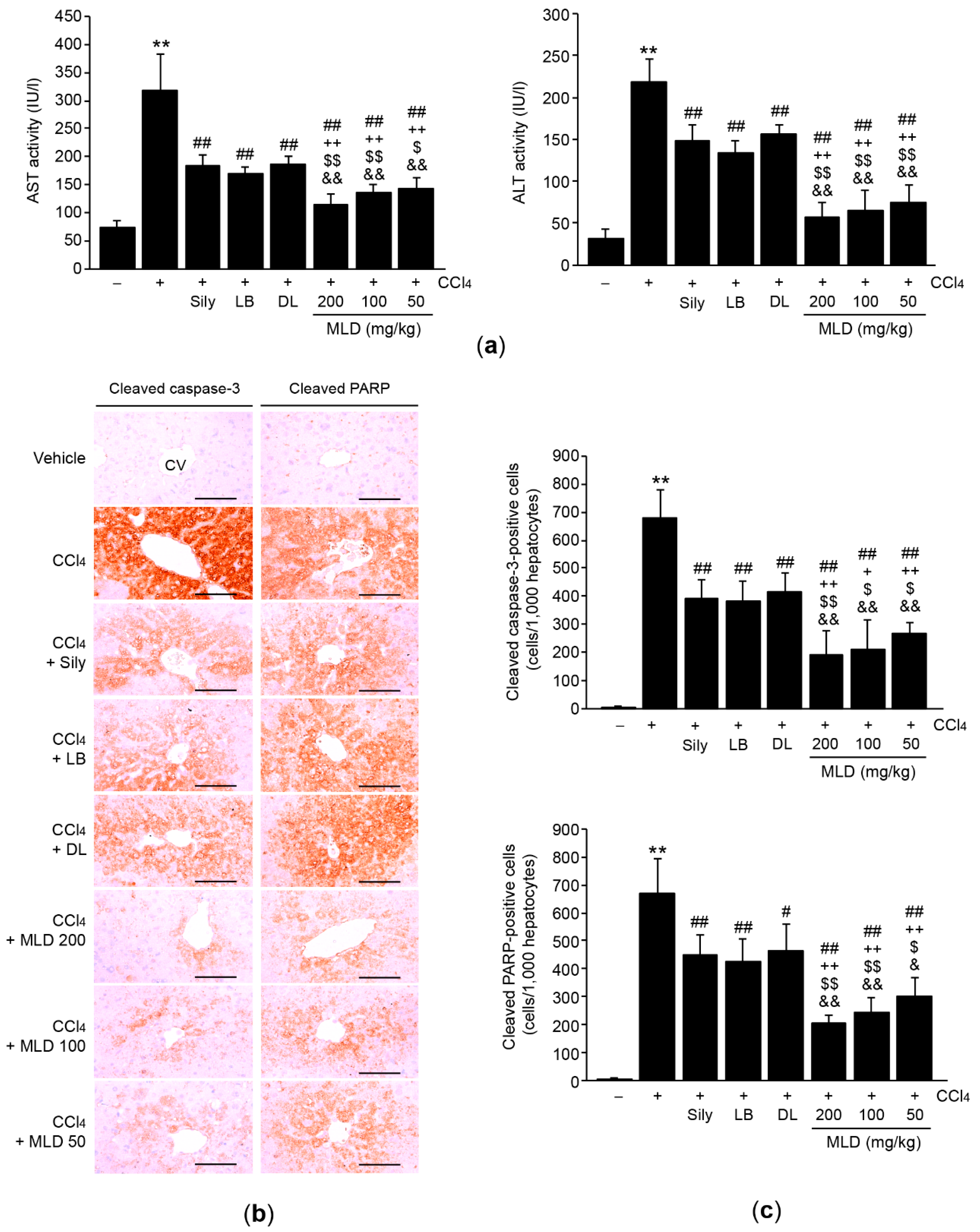
2.5. Measurement of Serum Aspartate Aminotransferase (AST) and Alanine Aminotransferase (ALT) Activities
2.6. Measurement of Lipid Peroxidation
2.7. Real-Time Polymerase Chain Reaction (PCR)
2.8. Determination of Glutathione Level, and Superoxide Dismutase (SOD) and Catalase Activities
2.9. Enzyme-Linked Immunosorbent Assay (ELISA)
2.10. Statistical Analyses
3. Results
3.1. MLD Synergistically Protects the Liver in CCl4-Treated Mice
3.2. MLD Synergistically Attenuates Hepatocyte Apoptosis
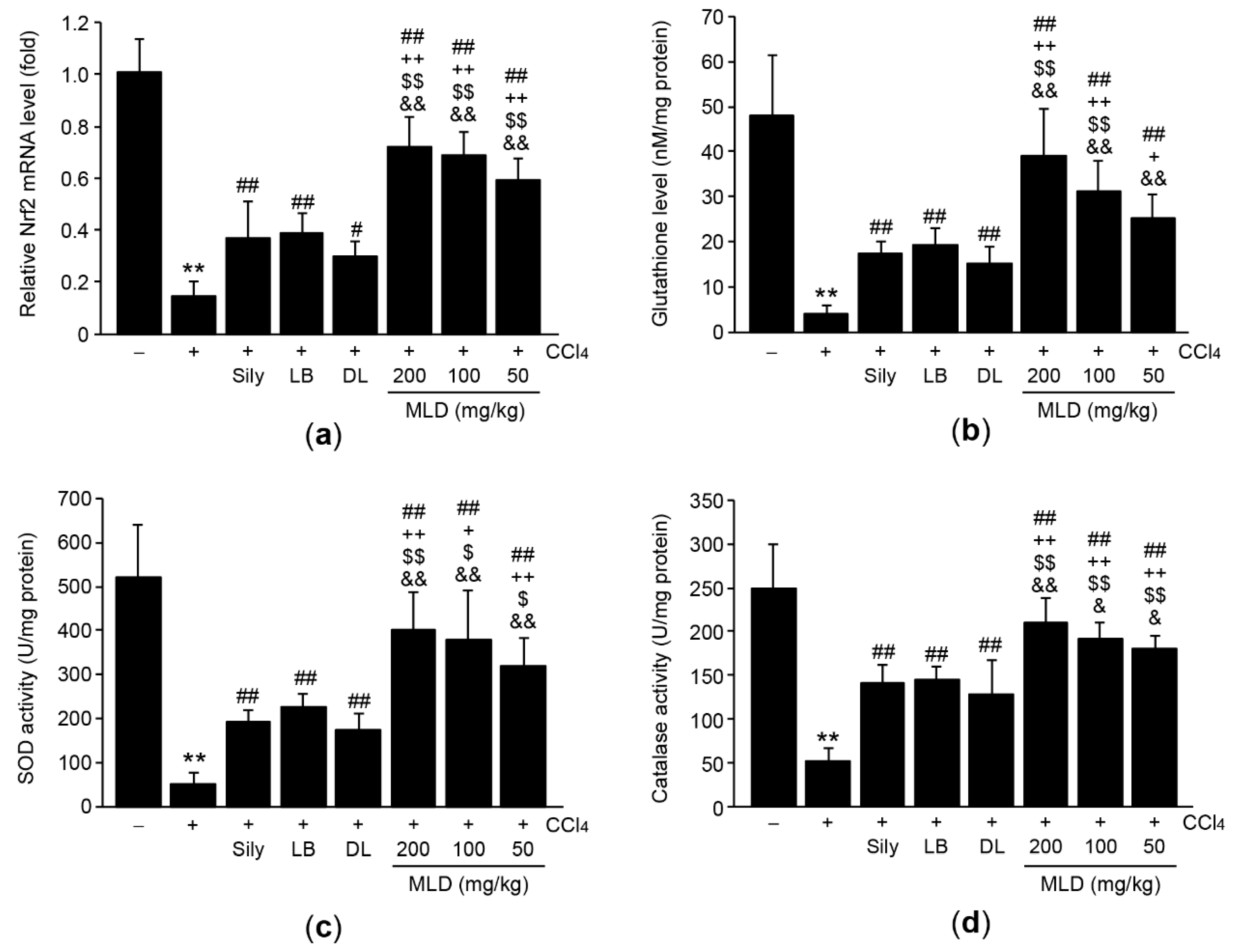
3.3. MLD Synergistically Alleviates CCl4-Mediated Oxidative Stress by Restoring Antioxidant Activity
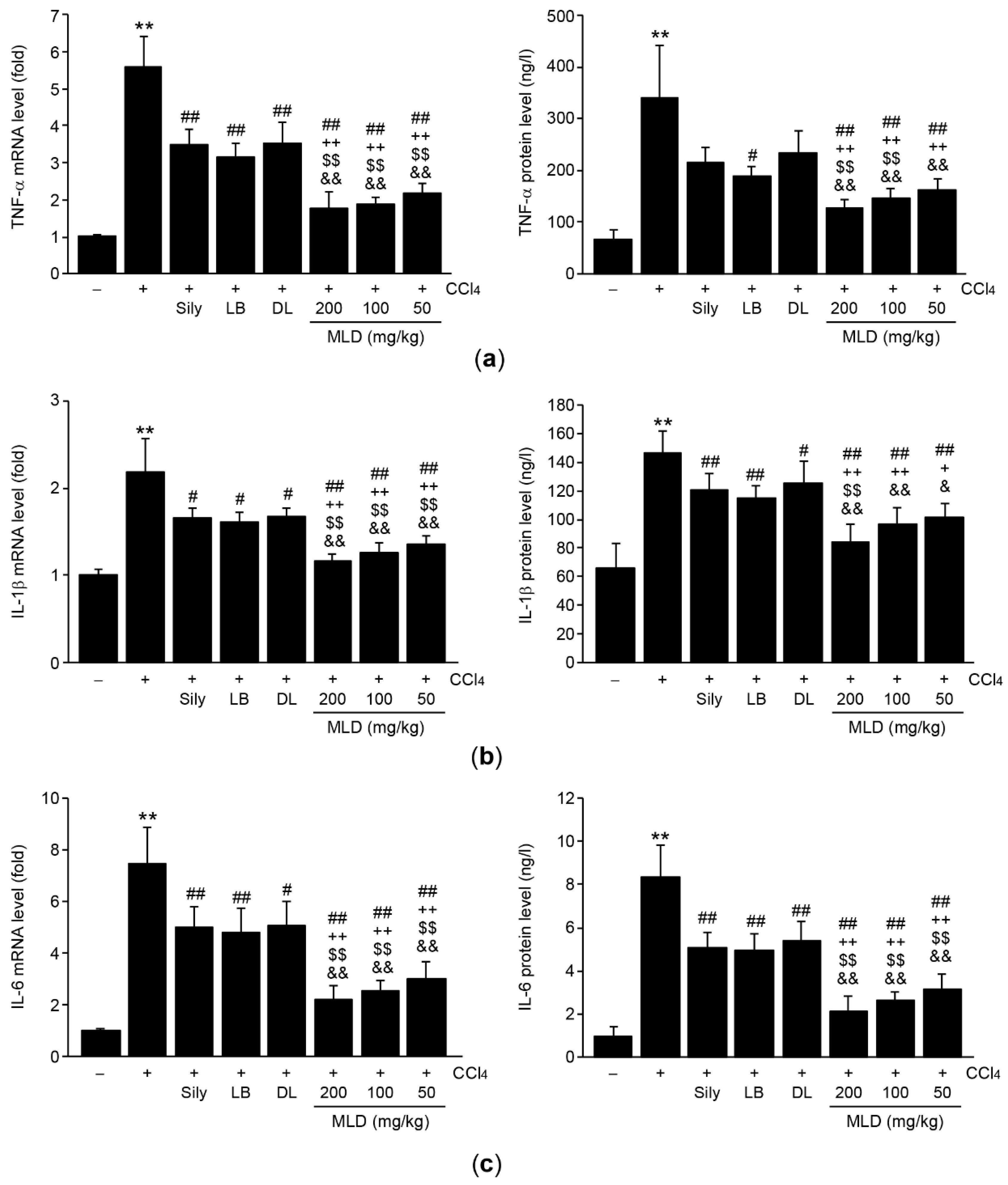
3.4. MLD Synergistically Decreases the Levels of Proinflammatory Cytokines in CCl4-Treated Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, J.; Giordano, S.; Zhang, J. Autophagy, mitochondria and oxidative stress: Cross-talk and redox signaling. Biochem. J. 2012, 441, 523–540. [Google Scholar] [CrossRef] [PubMed]
- Cichoż-Lach, H.; Michalak, A. Oxidative stress as a crucial factor in liver diseases. World J. Gastroenterol. 2014, 20, 8082–8091. [Google Scholar] [CrossRef] [PubMed]
- Brenner, C.; Galluzzi, L.; Keep, O.; Kroemer, G. Decoding cell death signals in liver inflammation. J. Hepatol. 2013, 59, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tan, H.Y.; Wang, N.; Zhang, Z.J.; Lao, L.; Wong, C.W.; Feng, Y. The role of oxidative stress and antioxidants in liver disease. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, A.; Sahebkar, A.; Javadi, B. Melissa officinalis L.—A review of its traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 2016, 188, 204–228. [Google Scholar]
- World Health Organization. Folium Melissae. In WHO Monographs on Selected Medicinal Plants, 1st ed.; World Health Organization, Ed.; World Health Organization: Geneva, Switzerland, 2004; Volume 2, pp. 180–187. [Google Scholar]
- Jun, H.J.; Lee, J.H.; Jia, Y.; Hoang, M.H.; Byun, H.; Kim, K.H.; Lee, S.J. Melissa officinalis essential oil reduces plasma triglycerides in human apolipoprotein E2 transgenic mice by inhibiting sterol regulatory element-binding protein-1c-dependent fatty acid synthesis. J. Nutr. 2012, 142, 432–440. [Google Scholar] [CrossRef][Green Version]
- Schütz, K.; Carle, R.; Schieber, A. Taraxacum—A review on its phytochemical and pharmacological profile. J. Ethnopharmacol. 2006, 107, 313–323. [Google Scholar] [CrossRef]
- Davaatseren, M.; Hur, H.J.; Yang, H.J.; Hwang, J.T.; Park, J.H.; Kim, H.J.; Kim, M.J.; Kwon, D.Y.; Sung, M.J. Taraxacum official (dandelion) leaf extract alleviates high-fat diet-induced nonalcoholic fatty liver. Food Chem. Toxicol. 2013, 58, 30–36. [Google Scholar] [CrossRef]
- Davaatseren, M.; Hur, H.J.; Yang, H.J.; Hwang, J.T.; Park, J.H.; Kim, H.J.; Kim, M.S.; Kim, M.J.; Kwon, D.Y.; Sung, M.J. Dandelion leaf extract protects against liver injury induced by methionine- and choline-deficient diet in mice. J. Med. Food. 2013, 16, 26–33. [Google Scholar] [CrossRef]
- Colle, D.; Arantes, L.P.; Gubert, P.; da Luz, S.C.; Athayde, M.L.; Teixeira Rocha, J.B.; Soares, F.A. Antioxidant properties of Taraxacum officinale leaf extract are involved in the protective effect against hepatoxicity induced by acetaminophen in mice. J. Med. Food. 2012, 15, 549–556. [Google Scholar] [CrossRef]
- Park, C.M.; Cha, Y.S.; Youn, H.J.; Cho, C.W.; Song, Y.S. Amelioration of oxidative stress by dandelion extract through CYP2E1 suppression against acute liver injury induced by carbon tetrachloride in Sprague-Dawley rats. Phytother. Res. 2010, 24, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- You, Y.; Yoo, S.; Yoon, H.G.; Park, J.; Lee, Y.H.; Kim, S.; Oh, K.T.; Lee, J.; Cho, H.Y.; Jun, W. In vitro and in vivo hepatoprotective effects of the aqueous extract from Taraxacum officinale (dandelion) root against alcohol-induced oxidative stress. Food Chem. Toxicol. 2010, 48, 1632–1637. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.Y.; Cho, H.S.; Lee, J.J.; Lee, H.J.; Jun, S.Y.; Lee, J.H.; Song, H.H.; Choi, S.; Saloura, V.; Park, C.G.; et al. Novel TRAIL sensitizer Taraxacum officinale F.H. Wigg enhances TRAIL-induced apoptosis in Huh7 cells. Mol. Carcinog. 2016, 55, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.R.; Cho, I.J.; Jung, S.J.; Kim, J.K.; Lee, D.G.; Ku, S.K.; Park, K.M. Study on the hepatoprotective effects of lemon balm and dandelion leaf extract combination in carbon tetrachloride-mediated liver injured mice. Herbal Formula Sci. 2019, 27, 199–211. [Google Scholar]
- Jung, J.Y.; Park, S.M.; Ko, H.L.; Lee, J.R.; Park, C.A.; Byun, S.H.; Ku, S.K.; Cho, I.J.; Kim, S.C. Epimedium koreanum ameliorates oxidative stress-mediated liver injury by activating nuclear factor erythroid 2-related factor 2. Am. J. Chin. Med. 2018, 46, 469–488. [Google Scholar] [CrossRef] [PubMed]
- Ishak, K.; Baptista, A.; Bianchi, L.; Callea, F.; De Groote, J.; Gudat, F.; Denk, H.; Desmet, V.; Korb, G.; MacSween, R.N.M.; et al. Histological grading and staging of chronic hepatitis. J. Hepatol. 1995, 22, 696–699. [Google Scholar] [CrossRef]
- Hu, J.R.; Chun, Y.S.; Kim, J.K.; Cho, I.J.; Ku, S.K. Ginseng berry aqueous extract prevents scopolamine-induced memory impairment in mice. Exp. Ther. Med. 2019, 18, 4388–4396. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Okamoto, T.; Okabe, S. Carbon tetrachloride treatment induces anorexia independently of hepatitis in rats. Int. J. Mol. Med. 2000, 6, 181–183. [Google Scholar] [CrossRef]
- Kobayashi, M.; Yamamoto, M. Nrf2-Keap1 regulation of cellular defense mechanisms against electrophiles and reactive oxygen species. Adv. Enzyme Regul. 2006, 46, 113–140. [Google Scholar] [CrossRef]
- Schütz, K.; Kammerer, D.R.; Carle, R.; Schieber, A. Characterization of phenolic acids and flavonoids in dandelion (Taraxacum officinale WEB. ex WIGG.) root and herb by high-performance liquid chromatography/electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2005, 19, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.R.; Cho, I.J.; Jung, S.J.; Kim, J.K.; Park, S.M.; Lee, D.G.; Ku, S.K.; Park, K.M. Lemon balm and dandelion leaf extract synergistically alleviate ethanol-induced hepatotoxicity by enhancing antioxidant and anti-inflammatory activity. J. Food Biochem. 2020, 44, e13232. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Lai, Y.; Huang, P.; Xie, L.; Lin, H.; Zhou, Z.; Mo, C.; Deng, G.; Yan, W.; Gao, Z.; et al. Naringin attenuates alcoholic liver injury by reducing lipid accumulation and oxidative stress. Life Sci. 2019, 216, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Shi, A.; Shi, H.; Wang, Y.; Liu, X.; Cheng, Y.; Li, H.; Zhao, H.; Wang, S.; Dong, L. Activation of Nrf2 pathway and inhibition of NLRP3 inflammasome activation contribute to the protective effect of chlorogenic acid on acute liver injury. Int. Immunopharmacol. 2018, 54, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Kim, S.C.; Kim, K.M.; Jang, C.H.; Cho, S.S.; Kim, S.J.; Ku, S.K.; Cho, I.J.; Ki, S.H. Isorhamnetin attenuates liver fibrosis by inhibiting TGF-β/Smad signaling and relieving oxidative stress. Eur. J. Pharmacol. 2016, 783, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Landmann, M.; Kanuri, G.; Spruss, A.; Stahl, C.; Bergheim, I. Oral intake of chicoric acid reduces acute alcohol-induced hepatic steatosis in mice. Nutrition 2014, 30, 882–889. [Google Scholar] [CrossRef]
- Domitrović, R.; Skoda, M.; Marchesi, V.V.; Cvijanović, O.; Pugel, E.P.; Stefan, M.B. Rosmarinic acid ameliorates acute liver damage and fibrogenesis in carbon tetrachloride-intoxicated mice. Food Chem. Toxicol. 2013, 51, 370–378. [Google Scholar]
- Domitrović, R.; Jakovac, H.; Milin, C.; Radosević-Stasić, B. Dose- and time-dependent effects of luteolin on carbon tetrachloride-induced hepatotoxicity in mice. Exp. Toxicol. Pathol. 2009, 61, 581–589. [Google Scholar] [CrossRef]
- Weber, L.W.; Boll, M.; Stampfl, A. Hepatotoxicity and mechanism of action of haloalkanes: Carbon tetrachloride as a toxicological model. Crit. Rev. Toxicol. 2003, 33, 105–136. [Google Scholar] [CrossRef]
- Choi, H.S.; Kang, J.W.; Lee, S.M. Melatonin attenuates carbon tetrachloride-induced liver fibrosis via inhibition of necroptosis. Transl. Res. 2015, 166, 292–303. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Z.; Nowicki, M.J. Caspase-12 mediates carbon tetrachloride-induced hepatocyte apoptosis in mice. World J. Gastroenterol. 2014, 20, 18189–18198. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.H.; Ting, W.J.; Shen, C.Y.; Hsu, H.H.; Lin, Y.M.; Kuo, C.H.; Tsai, F.J.; Tsai, C.H.; Tsai, Y.; Huang, C.Y. Anti-apoptotic effect of San Huang Shel Shin Tang cyclodextrin complex (SHSSTc) on CCl4-induced hepatotoxicity in rats. Environ. Toxicol. 2016, 31, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Xu, Y.; Xu, L.; Cong, X.; Yin, L.; Li, H.; Peng, J. Mechanism investigation of dioscin against CCl4-induced acute liver damage in mice. Environ. Toxicol. Pharmacol. 2012, 34, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Cavalcante, G.C.; Schaan, A.P.; Cabral, G.F.; Santana-da-Silva, M.N.; Pinto, P.; Vidal, A.F.; Ribeiro-Dos-Santos, Â. A cell’s fate: An overview of the molecular biology and genetics of apoptosis. Int. J. Mol. Sci. 2019, 20, 4133. [Google Scholar] [CrossRef] [PubMed]
- Breitzig, M.; Bhimineni, C.; Lockey, R.; Kolliputi, N. 4-Hydroxy-2-nonenal: A critical target in oxidative stress? Am. J. Physiol. Cell Physiol. 2016, 311, C537–C543. [Google Scholar] [CrossRef]
- Tipoe, G.L.; Leung, T.M.; Liong, E.; So, H.; Leung, K.M.; Lau, T.Y.; Tom, W.M.; Fung, M.L.; Fan, S.T.; Nanji, A.A. Inhibitors of inducible nitric oxide (NO) synthase are more effective than an NO donor in reducing carbon-tetrachloride induced acute liver injury. Histol. Histopathol. 2006, 21, 1157–1165. [Google Scholar]
- Xu, W.; Hellerbrand, C.; Köhler, U.A.; Bugnon, P.; Kan, Y.W.; Werner, S.; Beyer, T.A. The Nrf2 transcription factor protects from toxin-induced liver injury and fibrosis. Lab. Investig. 2008, 88, 1068–1078. [Google Scholar] [CrossRef]
- Lamlé, J.; Marhenke, S.; Borlak, J.; von Wasielewski, R.; Eriksson, C.J.; Geffers, R.; Manns, M.P.; Yamamoto, M.; Vogel, A. Nuclear factor-erythroid 2-related factor 2 prevents alcohol-induced fulminant liver injury. Gastroenterology 2008, 134, 1159–1168. [Google Scholar] [CrossRef]
- Enomoto, A.; Itoh, K.; Nagayoshi, E.; Haruta, J.; Kimura, T.; O’Connor, T.; Harada, T.; Yamamoto, M. High sensitivity of Nrf2 knockout mice to acetaminophen hepatotoxicity associated with decreased expression of ARE-regulated drug metabolizing enzymes and antioxidant genes. Toxicol. Sci. 2001, 59, 169–177. [Google Scholar] [CrossRef]
- Peng, C.; Zhou, Z.M.; Li, J.; Luo, Y.; Zhou, Y.S.; Ke, X.H.; Huang, K.E. CCl4-induced liver injury was ameliorated by Qi-Ge decoction through the antioxidant pathway. Evid. Based Complement. Alternat. Med. 2019, 2019, 5941263. [Google Scholar] [CrossRef]
- Esmaeili, M.A.; Alilou, M. Naringenin attenuates CCl4-induced hepatic inflammation by the activation of an Nrf2-mediated pathway in rats. Clin. Exp. Pharmacol. Physiol. 2014, 41, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Morio, L.A.; Chiu, H.; Sprowles, K.A.; Zhou, P.; Heck, D.E.; Gordon, M.K.; Laskin, D.L. Distinct role of tumor necrosis factor α and nitric oxide in acute liver injury induced by carbon tetrachloride in mice. Toxicol. Appl. Pharmacol. 2001, 172, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Wertheimer, S.J.; Myers, C.L.; Wallace, R.W.; Parks, T.P. Intercellular adhesion molecule-1 gene expression in human endothelial cells: Differential regulation by tumor necrosis factor-a and phorbol myristate acetate. J. Biol. Chem. 1992, 267, 12030–12035. [Google Scholar] [PubMed]
- Turner, N.A.; Mughal, R.S.; Warburton, P.; O’Regan, D.J.; Ball, S.G.; Porter, K.E. Mechanism of TNFalpha-induced IL-1alpha, IL-1beta and IL-6 expression in human cardiac fibroblasts: Effects of statins and thiazolidinediones. Cardiovasc. Res. 2007, 76, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wang, D.; Li, D.; Chen, X.; Wang, B.; Wang, F.; Liu, X.; Shang, J.; Zheng, Q. Licochalcone E protects against carbon tetrachloride-induced liver toxicity by activating peroxisome proliferator-activated receptor gamma. Mol. Med. Rep. 2017, 16, 5269–5276. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, E.H.; Suzuki, T.; Funayama, R.; Nagashima, T.; Hayashi, M.; Sekine, H.; Tanaka, N.; Moriguchi, T.; Motohashi, H.; Nakayama, K.; et al. Nrf2 suppresses macrophage inflammatory response by blocking cytokine transcription. Nature Commun. 2016, 7, 11624. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer | Backward Primer | GenBank ID | Amplicon Size (bp) |
|---|---|---|---|---|
| Nrf2 | 5′-CGAGATATACGCAGGA GAGGTAAGA-3′ | 5′-GCTCGACAATGTTCTC CAGCTT-3′ | AH006764.2 | 79 |
| TNF-α | 5′-GATCCGAGACGTGGAA CTGG-3′ | 5′-AGTTCAGTAGACAGA AGAGC-3′ | AB039227.1 | 140 |
| IL-1β | 5′-ATGGCAACTGTTCCTGA ACT-3′ | 5′-CAGGACAGGTATAGA TTCTT-3′ | NM_008361.3 | 563 |
| IL-6 | 5′-TTCCATCCAGTTGCCTT CTT-3′ | 5′-ATTTCCACGATTTCCCA GAG-3′ | DQ788722.1 | 170 |
| β-actin | 5′-GCTGAGAGGGAAATCG TGCGT-3′ | 5′-GAAGCATTTGCGGTGC ACGATG-3′ | NM_007393.5 | 516 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, B.-R.; Cho, I.-J.; Jung, S.-J.; Kim, J.-K.; Lee, D.-G.; Ku, S.-K.; Park, K.-M. Lemon Balm and Dandelion Leaf Extracts Synergistically Protect against Carbon Tetrachloride-Induced Acute Liver Injury in Mice. Appl. Sci. 2021, 11, 390. https://doi.org/10.3390/app11010390
Choi B-R, Cho I-J, Jung S-J, Kim J-K, Lee D-G, Ku S-K, Park K-M. Lemon Balm and Dandelion Leaf Extracts Synergistically Protect against Carbon Tetrachloride-Induced Acute Liver Injury in Mice. Applied Sciences. 2021; 11(1):390. https://doi.org/10.3390/app11010390
Chicago/Turabian StyleChoi, Beom-Rak, Il-Je Cho, Su-Jin Jung, Jae-Kwang Kim, Dae-Geon Lee, Sae-Kwang Ku, and Ki-Moon Park. 2021. "Lemon Balm and Dandelion Leaf Extracts Synergistically Protect against Carbon Tetrachloride-Induced Acute Liver Injury in Mice" Applied Sciences 11, no. 1: 390. https://doi.org/10.3390/app11010390
APA StyleChoi, B.-R., Cho, I.-J., Jung, S.-J., Kim, J.-K., Lee, D.-G., Ku, S.-K., & Park, K.-M. (2021). Lemon Balm and Dandelion Leaf Extracts Synergistically Protect against Carbon Tetrachloride-Induced Acute Liver Injury in Mice. Applied Sciences, 11(1), 390. https://doi.org/10.3390/app11010390