Microcystis aeruginosa and M. wesenbergii Were the Primary Planktonic Microcystin Producers in Several Bulgarian Waterbodies (August 2019)

 ,
,  , ,
, ,
Abstract
Featured Application
Abstract
1. Introduction
2. Materials and Methods
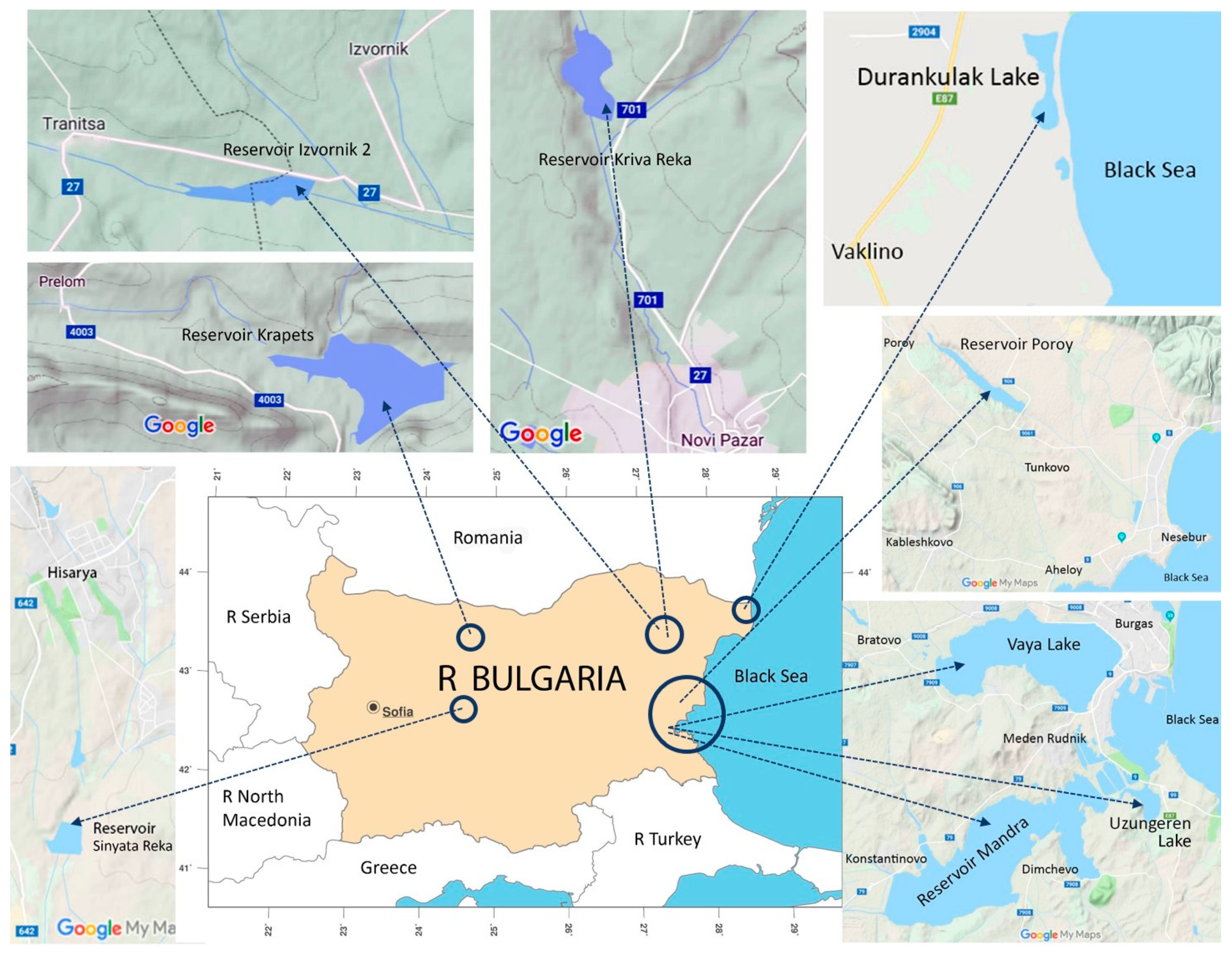
2.1. Sites and Sampling
2.2. Phytoplankton Species Composition and Abundance Assessment by Conventional Light Microscopy (LM)
2.3. Phytoplankton According to HPLC Marker Pigment Analysis
2.4. Identification of Toxins
2.5. Molecular-Genetic Studies
3. Results
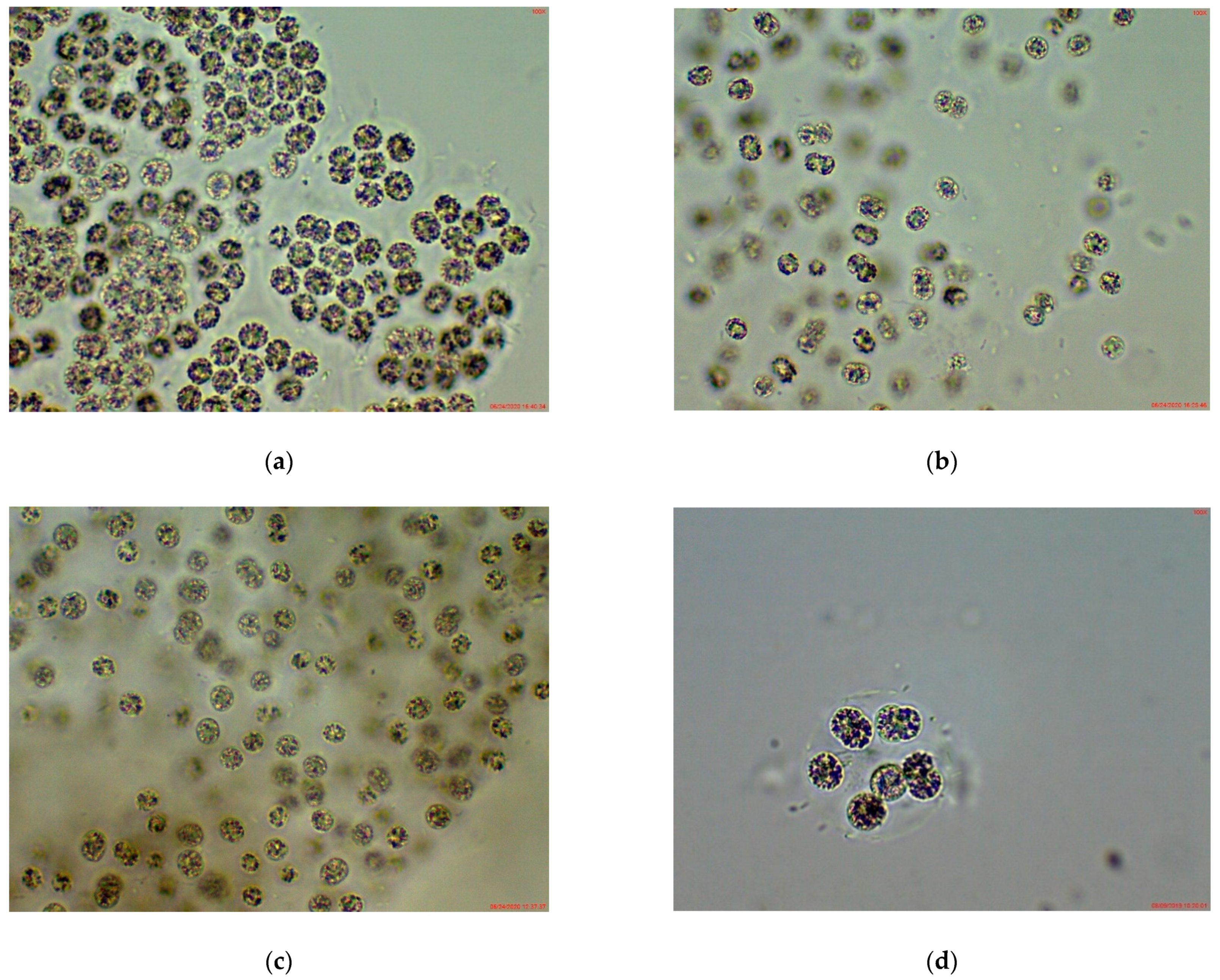
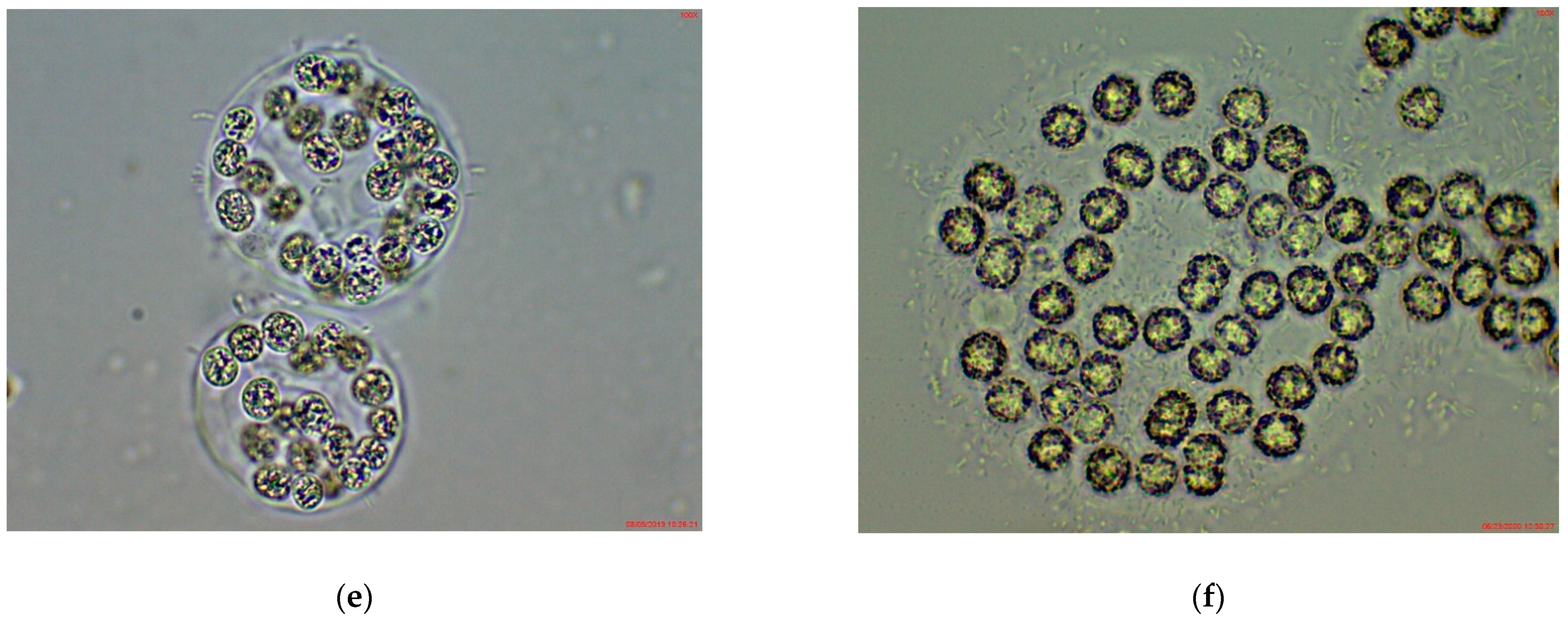
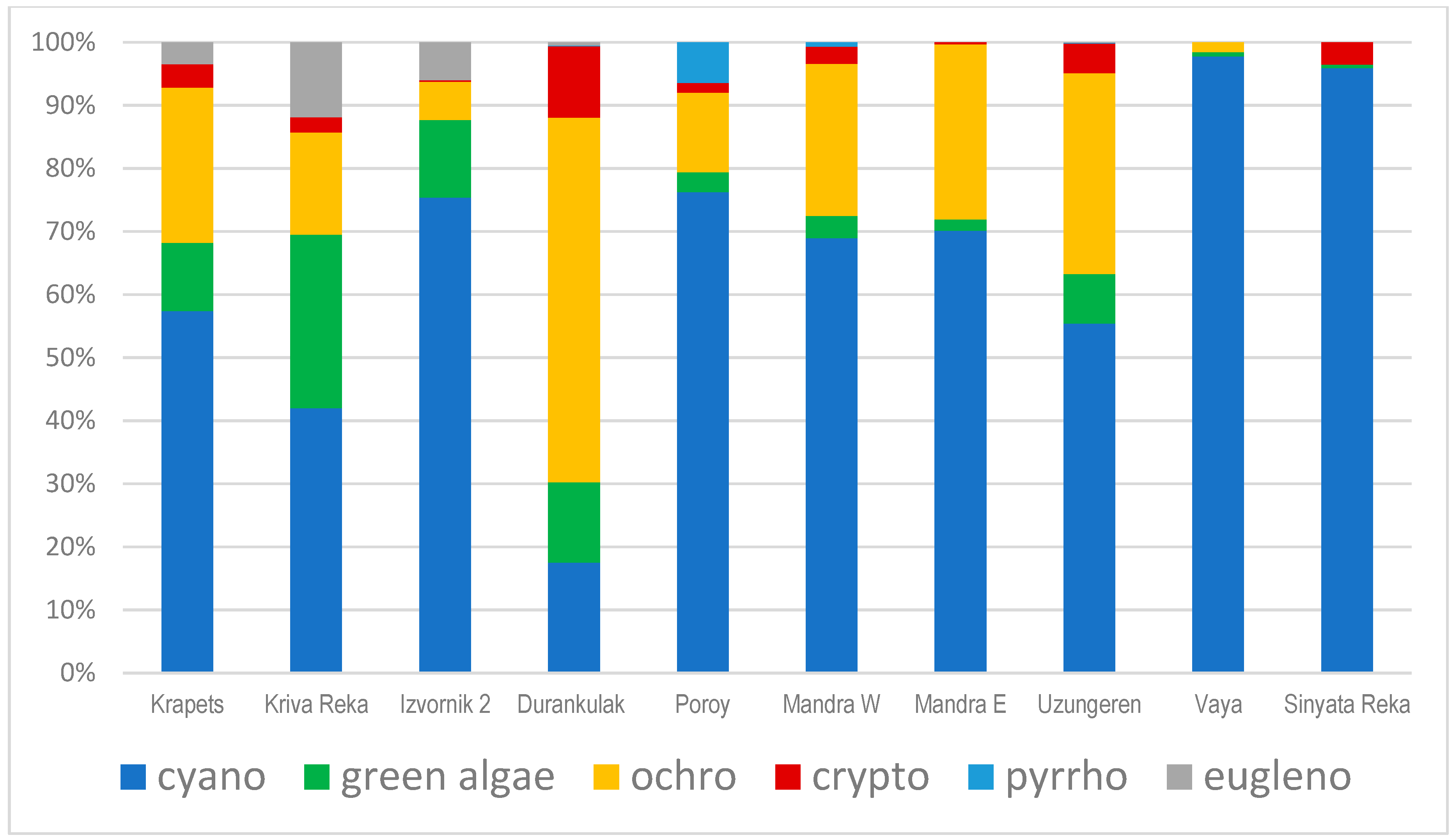
3.1. Phytoplankton Species Composition and Abundance, Obtained by Light Microscopy (LM)
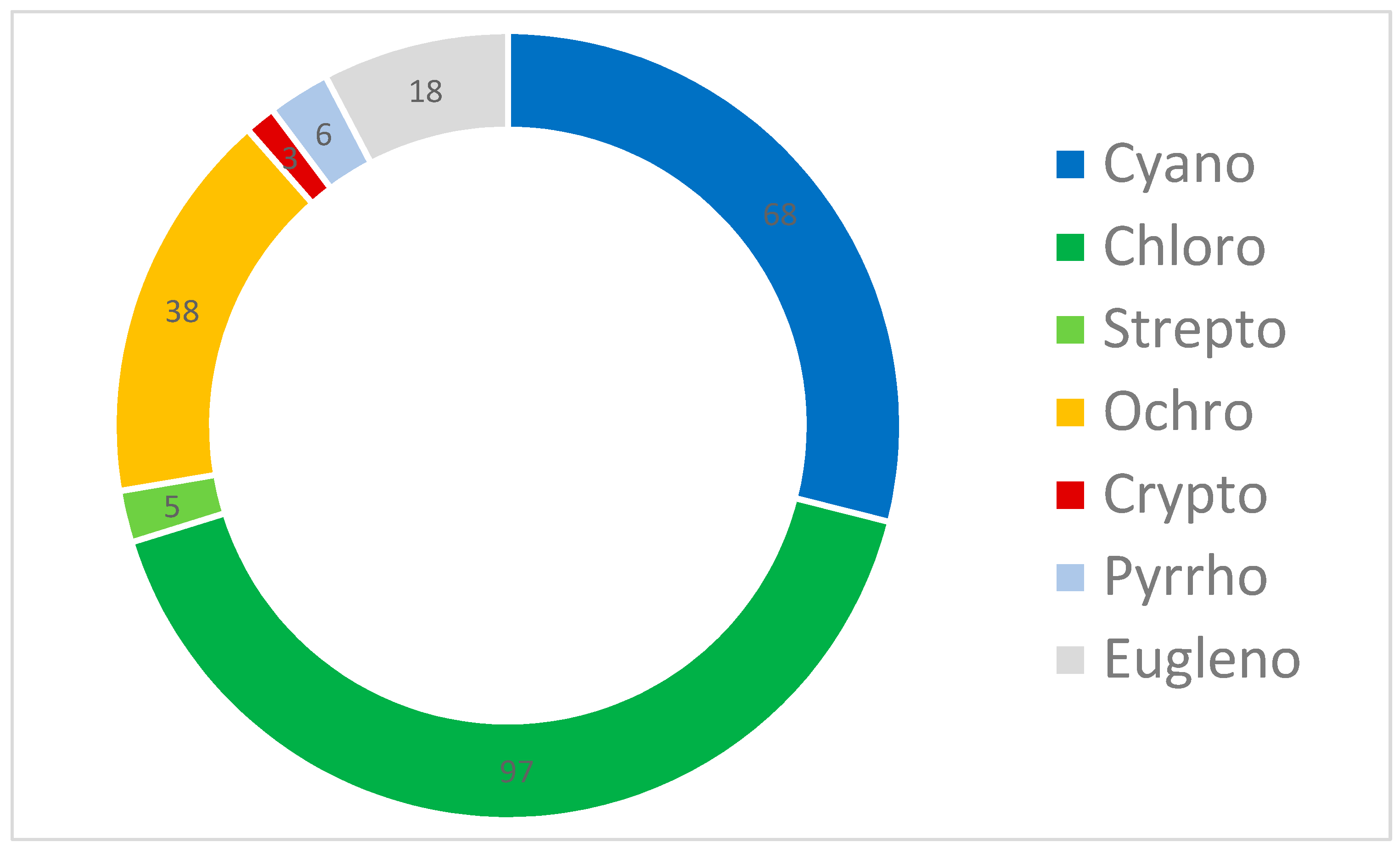
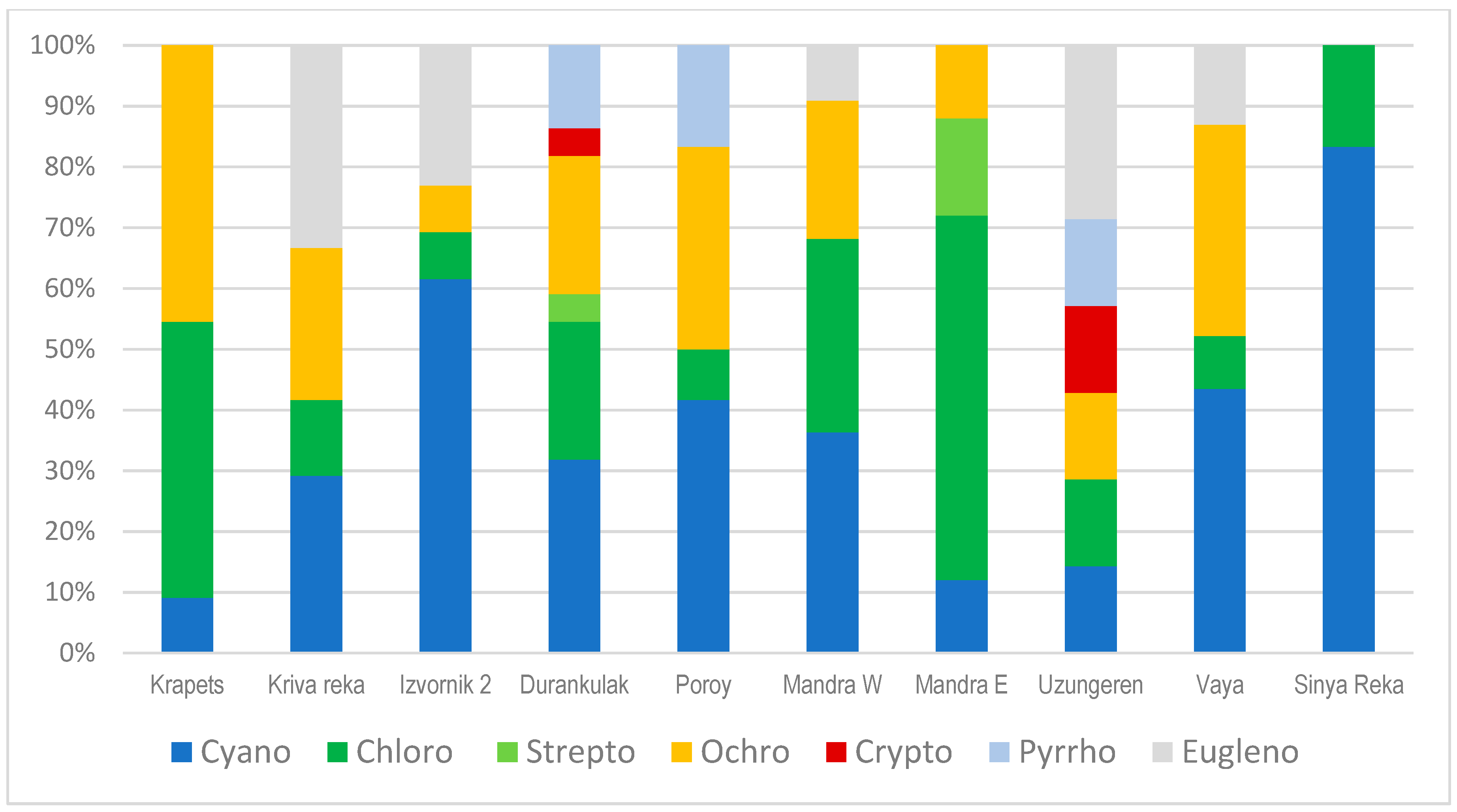
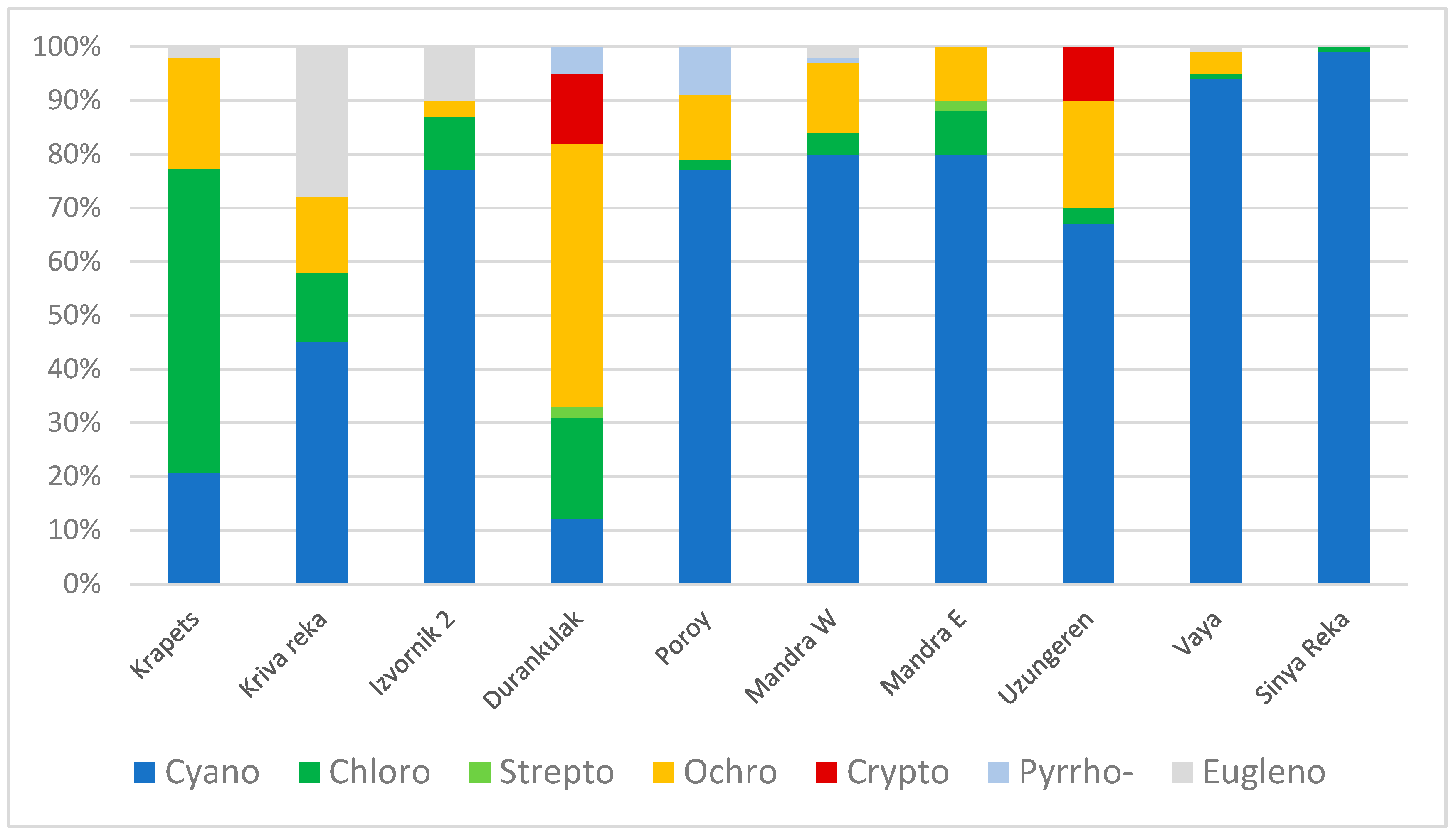
3.2. Results from Marker Pigment Analysis
3.3. Results from Toxin Analysis
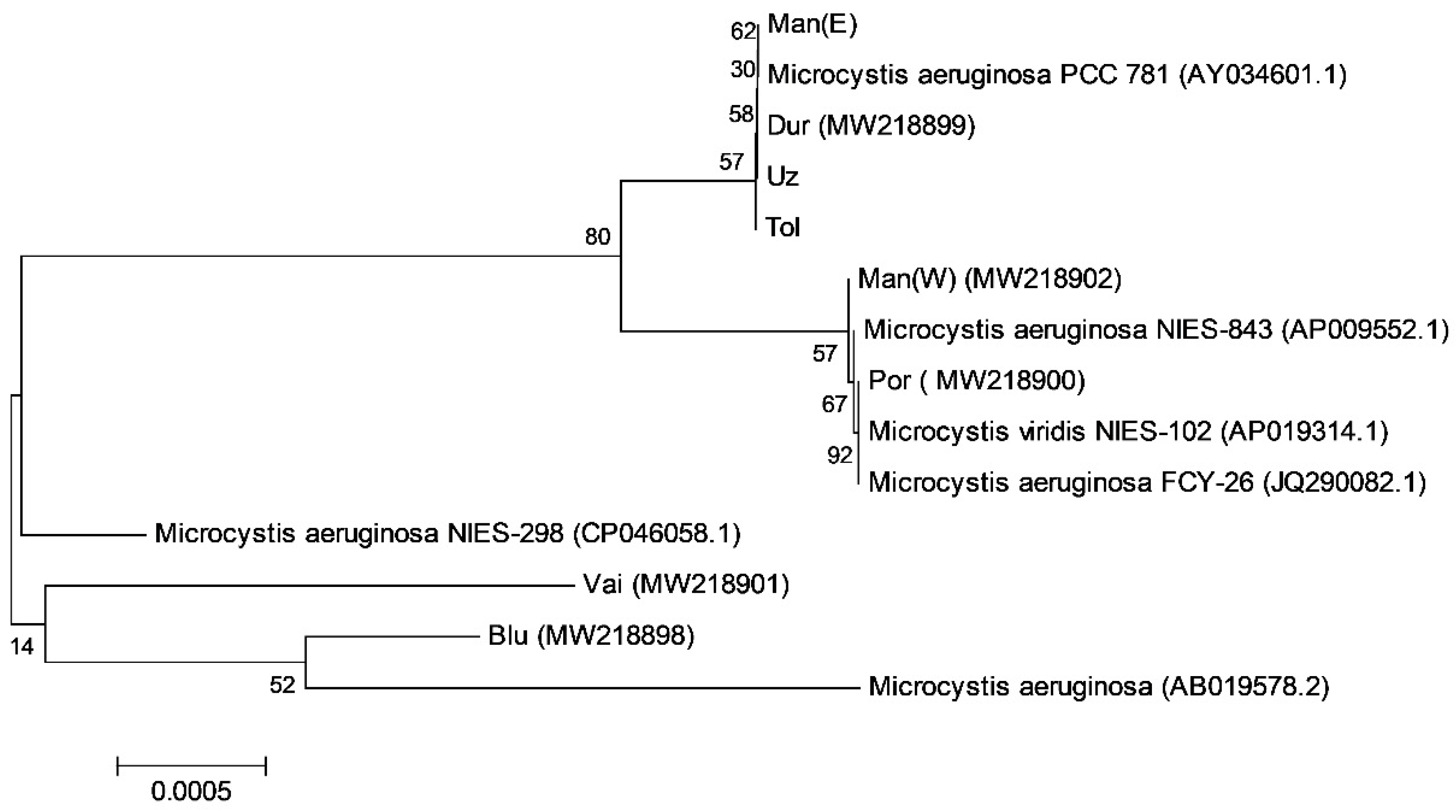
3.4. Results from PCR Analysis for Microcystin-Producing Strains
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anderson, D. HABs in a changing world: A perspective on harmful algal blooms, their impacts, and research and management in a dynamic era of climactic and environmental change. Harmful Algae 2014, 2012, 3–17. [Google Scholar]
- Svirčev, Z.; Lalić, D.; Savić, G.B.; Tokodi, N.; Backović, D.D.; Chen, L.; Meriluoto, J.; Codd, G.A. Global geographical and historical overview of cyanotoxin distribution and cyanobacterial poisonings. Arch. Toxicol. 2019, 93, 2429–2481. [Google Scholar] [CrossRef] [PubMed]
- Mitrovic, S.M.; Kobayashi, T.; Roelke, D.L. Cyanobacteria in inland waters: New monitoring, reporting, modelling and ecological research. Mar. Freshw. Res. 2020, 71, i–iv. [Google Scholar] [CrossRef]
- Meriluoto, J.; Blaha, L.; Bojadzija, G.; Bormans, M.; Brient, L.; Codd, G.A.; Drobac, D.; Faassen, E.J.; Fastner, J.; Anastasia, H.; et al. Toxic cyanobacteria and cyanotoxins in European waters—Recent progress achieved through the CYANOCOST Action and challenges for further research. Adv. Oceanogr. Limnol. 2017, 8, 161–178. [Google Scholar] [CrossRef]
- Jankowiak, J.T.; Hattenrath-Lehmann, B.J.; Kramer, M.L.; Gobler, C.J. Deciphering the effects of nitrogen, phosphorus, and temperature on cyanobacterial bloom intensification, diversity, and toxicity in western Lake Erie. Limnol. Oceanogr. 2019, 64, 1347–1370. [Google Scholar] [CrossRef]
- Janssen, E.M.L. Cyanobacterial peptides beyond microcystins—A review on co-occurrence, toxicity and challenges for risk assessment. Water Res. 2019, 151, 488–499. [Google Scholar] [CrossRef]
- Ghaffar, S.; Stevenson, R.J.; Khan, Z. Effect of phosphorus stress on Microcystis aeruginosa growth and phosphorus uptake. PLoS ONE 2017, 12, e0174349. [Google Scholar] [CrossRef]
- Xiao, M.; Li, M.; Reynolds, C.S. Colony formation in the cyanobacterium Microcystis. Biol. Rev. 2018, 93, 1399–1420. [Google Scholar] [CrossRef]
- Radkova, M.; Stefanova, K.; Uzunov, B.; Gärtner, G.; Stoyneva-Gärtner, M. Morphological and molecular identification of microcystin-producing cyanobacteria in nine shallow Bulgarian water bodies. Toxins 2020, 12, 39. [Google Scholar] [CrossRef]
- Scherer, P.I.; Raeder, U.; Geist, J.; Zwirglmaier, K. Influence of temperature, mixing, and addition of microcystin-LR on microcystin gene expression in Microcystis aeruginosa. MicrobiologyOpen 2017, 6, e00393. [Google Scholar] [CrossRef]
- Davis, T.W.; Berry, D.L.; Boyer, G.L.; Gobler, C.J. The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae 2009, 8, 715–725. [Google Scholar] [CrossRef]
- Vézie, C.; Rapala, J.; Vaitomaa, J.; Seitsonen, J.; Sivonen, K. Effect of nitrogen and phosphorus on growth of toxic and nontoxic Microcystis strains and on intracellular microcystin concentrations. Microb. Ecol. 2002, 43, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Kaufononga, S.A.F.; Cary, S.C.; Hamilton, D.P. High levels of structural diversity observed in microcystins from Microcystis CAWBG11 and characterization of six new microcystin congeners. Mar. Drugs 2014, 12, 5372–5395. [Google Scholar] [CrossRef] [PubMed]
- Rantala, A.; Rajaniemi-Wacklin, P.; Lyra, C.; Lepisto, L.; Rintala, J.; Mankiewicz-Boczek, J.; Sivonen, K. Detection of microcystin-producing cyanobacteria in Finnish lakes with genus-specific microcystin synthetase gene E (mcyE) PCR and associations with environmental factors. Appl. Environ. Microbiol. 2006, 72, 6101–6110. [Google Scholar] [CrossRef] [PubMed]
- Hisbergues, M.; Christiansen, G.; Rouhiainen, L.; Sivonen, K.; Börner, T. PCR-based identification of microcystin-producing genotypes of different cyanobacterial genera. Arch. Microbiol. 2003, 180, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Lyon-Colbert, A.; Su, S.; Cude, C. A systematic literature review for evidence of Aphanizomenon flos-aquae toxigenicity in recreational waters and toxicity of dietary supplements: 2000–2017. Toxins 2018, 10, 254. [Google Scholar] [CrossRef]
- Panou, M.; Zervou, S.-K.; Kaloudis, T.; Hiskia, A.; Gkelis, S. A Greek Cylindrospermopsis raciborskii strain: Missing link in tropic invader’s phylogeography tale. Harmful Algae 2018, 80, 96–106. [Google Scholar] [CrossRef]
- Michev, T.; Stoyneva, M. (Eds.) Inventory of Bulgarian Wetlands and Their Biodiversity; Elsi-M: Sofia, Bulgaria, 2007. [Google Scholar]
- Stoyneva-Gärtner, M.P.; Descy, J.-P.; Latli, A.; Uzunov, B.; Pavlova, V.; Bratanova, Z.; Babica, P.; Maršálek, B.; Meriluoto, J.; Spoof, L. Assessment of cyanoprokaryote blooms and of cyanotoxins in Bulgaria in a 15-years period (2000–2015). Adv. Oceanogr. Limnol. 2017, 8, 131–152. [Google Scholar] [CrossRef][Green Version]
- Stoyneva-Gärtner, M.; Uzunov, B.; Dimitrova, P.; Pavlova, V. Algal toxins—New risk factors for national security in Bulgaria. In Proceedings of the Actual Problems of the Security, Veliko Turnovo, Bulgaria, 26–27 October 2017; Publishing House Complex of NVU “Vasil Levski”: Veliko Turnovo, Bulgaria, 2017; pp. 271–281, Electronic Publication. ISSN 2367-7473 (In Bulgarian, English Summary). [Google Scholar]
- Stoyneva-Gärtner, M.P.; Uzunov, B.A.; Dimitrova, P. Pilot assessment of cyanotoxins as potential risk factors for cancer in Bulgaria. BioDiscovery 2017, 20, e20501. [Google Scholar] [CrossRef][Green Version]
- Descy, J.-P.; Stoyneva-Gärtner, M.P.; Uzunov, B.A.; Dimitrova, P.H.; Pavlova, V.T.; Gärtner, G. Studies on cyanoprokaryotes of the water bodies along the Bulgarian Black Sea Coast (1890–2017): A review, with special reference to new, rare and harmful taxa. Acta Zool. Bulgar. Suppl. 2018, 11, 43–52. [Google Scholar]
- Stoyneva-Gärtner, M.P.; Uzunov, B.A.; Descy, J.-P.; Gärtner, G.; Draganova, P.H.; Borisova, C.I.; Pavlova, V.; Mitreva, M. Pilot application of drone-observations and pigment marker detection by HPLC in the studies of CyanoHABs in Bulgarian inland waters. Mar. Freshw. Res. 2019, 71, 606–616. [Google Scholar] [CrossRef]
- Andersen, R.A. Algal Culturing Techniques; Elsevier Academic Press: Burlington, NJ, USA, 2005. [Google Scholar]
- Uzunov, B.; Stoyneva, M.; Mancheva, A.; Gärtner, G. ACUS—The new collection of living aeroterrestrial algae of the University of Sofia ‘St Kliment Ohridski’. In Proceedings of the VII National Botanical Conference, Sofia, Bulgaria, 29–30 September 2011; Petrova, A., Ed.; Bulgarian Botanical Society: Sofia, Bulgaria, 2012; pp. 271–274. [Google Scholar]
- AlgaeBase. Available online: http://www.algaebase.org/ (accessed on 28 October 2020).
- Geitler, L. Cyanophyceae. In Rabenhorst's Kryptogamenflora von Deutschland, Österreich und der Schweiz, 2nd ed.; Rabenhorst, L., Ed.; Akademische Verlagsgesellschaft: Leipzig, Germany, 1932; Volume 14, pp. 1–1196. [Google Scholar]
- Geitler, L. Schizophyta: Klasse Schizophyceae. In Die natürlichen Pflanzenfamilien, Zweite Auflage; Engler, A., Prantl, K., Eds.; Wilhelm Engelmann: Leipzig, Germany, 1942; Volume 1b, pp. 1–232. [Google Scholar]
- Gollerbakh, M.M.; Kossinskaya, E.K.; Polyanskiy, V.I. Manual of Freshwater Algae of the USSR. Volume 2. Blue-Green Algae; Sovetskaya Nauka: Moscow, Russia, 1953. [Google Scholar]
- Starmach, K. Cyanophyta-Sinice. Glaucophyta-Glaukofity; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1966. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota. Teil 1/Part 1: Chroococcales. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Spektrum Akademischer Verlag: Heidelberg, Germany, 2008; Volume 19/1, pp. 1–556. [Google Scholar]
- Komárek, J.; Komárková, J. Review of the European Microcystis morphospecies (Cyanoprokaryotes) from nature. Czech Phycol. 2002, 2, 1–24. [Google Scholar]
- Šejnohová, L.; Maršálek, B. Microcystis. In Ecology of Cyanobacteria II. Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 195–228. [Google Scholar]
- CyanoDB 2.0. Available online: http://www.cyanodb.cz (accessed on 29 October 2020).
- Catherine, Q.; Wood, S.; Echenigue-Subiabre, I.; Heath, M.; Villeneuve, A.; Humbert, J.-F. A review of current knowledge on toxic benthic freshwater cyanobacteria—Ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar] [CrossRef]
- Bernard, C.; Ballot, A.; Thomazeau, S.; Maloufi, S.; Furey, A.; Mankiewicz-Boczek, I.; Pawlik-Skowrońska, B.; Capelli, C.; Salmazo, N. Appendix 2. Cyanobacteria associated with the production of cyanotoxins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, J., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 501–525. [Google Scholar]
- Rott, E. Some results from phytoplankton counting intercalibration. Schweiz. Z. Hydrol. 1981, 43, 34–62. [Google Scholar] [CrossRef]
- Stoyneva, M.P.; Descy, J.-P.; Vyverman, W. Green algae in Lake Tanganyika: Is morphological variation a response to seasonal changes? Hydrobiologia 2007, 578, 7–16. [Google Scholar] [CrossRef]
- Descy, J.P. SOP5: Estimation of cyanobacteria biomass by marker pigment analysis. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, J., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 343–349. [Google Scholar]
- Mackey, M.D.; Mackey, D.J.; Higgins, H.W.; Wright, S.W. CHEMTAX—A program for estimating class abundances from chemical markers: Application to HPLC measurements of phytoplankton. Mar. Ecol. Prog. Ser. 1996, 144, 265–283. [Google Scholar] [CrossRef]
- Sarmento, H.; Descy, J.-P. Use of marker pigments and functional groups for assessing the status of phytoplankton assemblages in lakes. J. Appl. Phycol. 2008, 20, 1001–1011. [Google Scholar] [CrossRef]
- Van Wichelen, J.; Van Gremberghe, I.; Vanormelingen, P.; Debeer, A.-E.; Leporcq, B.; Menzel, D.; Codd, G.A.; Descy, J.-P.; Vyverman, W. Strong effects of amoebae grazing on the biomass and genetic structure of a Microcystis bloom (Cyanobacteria). Environ. Microbiol. 2010, 12, 2797–2813. [Google Scholar] [CrossRef]
- International Standard Organization. ISO 20179:2005. Water Quality—Determination of Microcystins—Method Using Solid Phase Extraction (SPE) and High Performance Liquid Chromatography (HPLC) with Ultraviolet (UV) Detection. Available online: https://www.iso.org/standard/34098.html (accessed on 18 December 2020).
- Hiskia, A.; Spoof, L.; Kaloudis, T.; Meriluoto, J. Determination of cyanotoxins by High-Performance Liquid Chromatography with Photodiode Array. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, J., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 205–211. [Google Scholar]
- Heck, K.; Alvarenga, D.O.; Shishido, T.K.; Varani, A.M.; Dorr, F.A.; Pinto, E.; Rouhiainen, L.; Jokela, J.; Sivonen, K.; Fiore, M.F. Biosynthesis of microcystin hepatotoxins in the cyanobacterial genus Fischerella. Toxicon 2017, 141, 43–50. [Google Scholar] [CrossRef]
- Humbert, J.-F. Molecular tools for the detection of toxigenic Cyanobacteria in natural ecosystems. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, J., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 280–283. [Google Scholar]
- Pacheco, A.B.F.; Guedes, I.A.; Azevedo, S.M.F.O. Is qPCR a reliable indicator of cyanotoxin risk in freshwater? Toxins 2016, 8, 172. [Google Scholar] [CrossRef]
- Vaitomaa, J.; Rantala, A.; Halinen, K.; Rouhiainen, L.; Tallberg, P.; Mokelke, L.; Sivonen, K. Quantitative real-time PCR for determination of microcystin synthetase E copy numbers for Microcystis and Anabaena in lakes. Appl. Environ. Microbiol. 2003, 69, 7289–7297. [Google Scholar] [CrossRef] [PubMed]
- Moffitt, M.C.; Neilan, B.A. Characterization of the nodularin synthetase gene cluster and proposed theory of the evolution of cyanobacterial hepatotoxins. Appl. Environ. Microb. 2004, 70, 6353–6362. [Google Scholar] [CrossRef] [PubMed]
- Mankiewicz-Boczek, J.; Izydorczyk, K.; Romanowska-Duda, Z.; Jurczak, T.; Stefaniak, K.; Kokocinski, M. Detection and monitoring toxigenicity of cyanobacteria by application of molecular methods. Environ. Toxicol. 2006, 21, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Nonneman, D.; Zimba, P.V. A PCR-based test to assess the potential for microcystin occurrence in Channel catfish production ponds. J. Phycol. 2002, 38, 230–233. [Google Scholar] [CrossRef]
- Jungblut, A.-D.; Neilan, B.A. Molecular identification and evolution of the cyclic peptide hepatotoxins, microcystin and nodularin synthetase genes in three orders of cyanobacteria. Arch. Microbiol. 2006, 185, 107–114. [Google Scholar] [CrossRef]
- Foulds, I.V.; Granacki, A.; Xiao, C.; Krull, U.J.; Castle, A.; Horgen, P.A. Quantification of microcystin-producing cyanobacteria and E. coli in water by 5′-nuclease PCR. J. Appl. Microbiol. 2002, 93, 825–834. [Google Scholar] [CrossRef]
- Basic Local Alignment Search Tool (BLAST). Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 24 October 2019).
- National Centre for Biotechnology Information (NCBI). Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 24 October 2019).
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Reynolds, C.S. The Ecology of Phytoplankton; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Whitton, B.A. (Ed.) Ecology of Cyanobacteria II: Their Diversity in Space and Time, 2nd ed.; Springer: Dordrecht, the Netherlands, 2012. [Google Scholar]
- Chorus, I.; Bartram, J. (Eds.) Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; E & FN Spon: London, UK, 1999. [Google Scholar]
- Tang, X.; Krausfeldt, L.E.; Shao, K.; LeCleir, G.R.; Stough, J.M.A.; Gao, G.; Boyer, G.L.; Zhang, Y.; Paerl, H.W.; Qin, B.; et al. Seasonal Gene Expression and the ecophysiological implications of toxic Microcystis aeruginosa blooms in Lake Taihu. Environ. Sci. Technol. 2018, 52, 11049–11059. [Google Scholar] [CrossRef]
- Vasconcelos, V. Global changes and the new challenges in the research on cyanotoxin risk evaluation. Limnetica 2015, 34, 149–158. [Google Scholar]
- Komárek, J. A polyphasic approach for the taxonomy of cyanobacteria: Principles and applications. Eur. J. Phycol. 2016, 51, 346–353. [Google Scholar] [CrossRef]
- Köker, L.; Akçaalan, R.; Albay, M.; Neilan, B.A. Molecular detection of hepatotoxic cyanobacteria in inland water bodies of the Marmara Region, Turkey. Adv. Oceanogr. Limnol. 2017, 8, 52–60. [Google Scholar] [CrossRef]
- Gkelis, S.; Zaoutsos, N. Cyanotoxin occurrence and potentially toxin producing cyanobacteria in freshwaters of Greece: A multi-disciplinary approach. Toxicon 2014, 78, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yasuno, M.; Sugaya, Y.; Kaya, K.; Watanabe, M.M. Variations in the toxicity of Microcystis species to Moina macrocopa. Phycol. Res. 2006, 46, 31–36. [Google Scholar] [CrossRef]
- Christophoridis, C.; Zervou, S.-K.; Manolidi, K.; Katsiapi, M.; Moustaka-Gouni, M.; Kaloudis, T.; Triantis, T.M.; Hiskia, A. Occurrence and diversity of cyanotoxins in Greek lakes. Sci. Rep. 2018, 8, 17877. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Waterbody | Alt | Latitude | Longitude | WT | pH | SD | CN | TD | DO | TP | TN |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Res. Krapets (IBW2000) | 410 | 43°04.0316′ | 24°52.3835′ | 28.7 | 8.26 | 5.0 | 870 | 564 | 7.74 | 0.1 | 1 |
| Res. Kriva Reka (IBW3071) | 133 | 43°22.6573′ | 27°10.9807′ | 23.7 | 8.38 | 0.3 | 662 | 428 | 6.24 | 1.0 | 9 |
| Res. Izvornik 2 (IBW3082) | 255 | 43°27.3838′ | 27°21.111′ | 24.5 | 9.44 | 0.15 | 389 | 253 | 13.26 | 8.96 | 4.8 |
| Lake Durankulak (IBW0216) | 4 | 43°40.5355′ | 28°33.0806′ | 26.7 | 8.91 | 0.6 | 1048 | 680 | 6.04 | 0.33 | 0.63 |
| Res. Poroy (IBW3038) | 43 | 42°43.3403′ | 27°37.5255′ | 27.5 | 8.05 | 0.4 | 644 | 416 | 7.6 | 0.10 | 0.31 |
| Res. Mandra (IBW1720) - West | 7 | 42°24.0295′ | 27°19.1194′ | 25.88 | 7.9 | 0.45 | 676 | 436 | 7.93 | 0.66 | 0.46 |
| - East | 8 | 42°25.9303′ | 27°26.7652′ | 27.2 | 8.46 | 0.45 | 578 | 375 | 7.87 | 1.5 | 1.8 |
| Lake Uzungeren (IBW0710) | −3 | 42°26.1551′ | 27°27.2235′ | 27.6 | 8.45 | 0.45 | 1748 | 1132 | 9.7 | 0.40 | 0.28 |
| Lake Vaya (IBW0191) | −2 | 42°30.5940′ | 27°22.075′ | 27.9 | 9.22 | 0.15 | 490 | 17 | 7.69 | 0.50 | 0.26 |
| Res. Sinyata Reka (IBW1793) | 317 | 42°28.1518′ | 24°42.0159′ | 28.2 | 10.39 | 0.4 | 490 | 317 | 14.76 | 1.0 | 0.23 |
| Waterbody | Microcystis aeruginosa | Microcystis wesenbergii |
|---|---|---|
| Reservoir Izvornik 2 | <0.5% | <0.5% |
| Lake Durankulak | n.d. | <1% |
| Reservoir Poroy | n.d. | <0.5% |
| Reservoir Mandra West | <1% | <0.5% |
| Reservoir Mandra East | <5% | <0.5% |
| Reservoir Sinyata Reka | <0.5% | <1% |
| Waterbody | Sampling Date | MC–RR, –YR, –LR |
|---|---|---|
| Reservoir Krapets | 14 August 2019 | <LOD |
| Reservoir Kriva Reka | 16 August 2020 | <LOD |
| Reservoir Izvornik 2 | 16 August 2019 | <LOD |
| Lake Durankulak | 16 August 2019 | LR 0.15 µg L−1 |
| Reservoir Poroy | 17 August 2019 | <LOD |
| Reservoir Mandra (site West) | 18 August 2019 | LR 0.24 µg L−1 |
| Reservoir Mandra (site East) | 19 August 2019 | LR 0.46 µg L−1 |
| Lake Uzungeren | 19 August 2019 | <LOD |
| Lake Vaya | 19 August 2019 | <LOD |
| Reservoir Sinyata Reka | 20 August 2019 | <LOD |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoyneva-Gärtner, M.; Stefanova, K.; Descy, J.-P.; Uzunov, B.; Radkova, M.; Pavlova, V.; Mitreva, M.; Gärtner, G. Microcystis aeruginosa and M. wesenbergii Were the Primary Planktonic Microcystin Producers in Several Bulgarian Waterbodies (August 2019). Appl. Sci. 2021, 11, 357. https://doi.org/10.3390/app11010357
Stoyneva-Gärtner M, Stefanova K, Descy J-P, Uzunov B, Radkova M, Pavlova V, Mitreva M, Gärtner G. Microcystis aeruginosa and M. wesenbergii Were the Primary Planktonic Microcystin Producers in Several Bulgarian Waterbodies (August 2019). Applied Sciences. 2021; 11(1):357. https://doi.org/10.3390/app11010357
Chicago/Turabian StyleStoyneva-Gärtner, Maya, Katerina Stefanova, Jean-Pierre Descy, Blagoy Uzunov, Mariana Radkova, Vera Pavlova, Mariya Mitreva, and Georg Gärtner. 2021. "Microcystis aeruginosa and M. wesenbergii Were the Primary Planktonic Microcystin Producers in Several Bulgarian Waterbodies (August 2019)" Applied Sciences 11, no. 1: 357. https://doi.org/10.3390/app11010357
APA StyleStoyneva-Gärtner, M., Stefanova, K., Descy, J.-P., Uzunov, B., Radkova, M., Pavlova, V., Mitreva, M., & Gärtner, G. (2021). Microcystis aeruginosa and M. wesenbergii Were the Primary Planktonic Microcystin Producers in Several Bulgarian Waterbodies (August 2019). Applied Sciences, 11(1), 357. https://doi.org/10.3390/app11010357