Metabolic Syndrome-Preventive Peptides Derived from Milk Proteins and Their Presence in Cheeses: A Review



Abstract

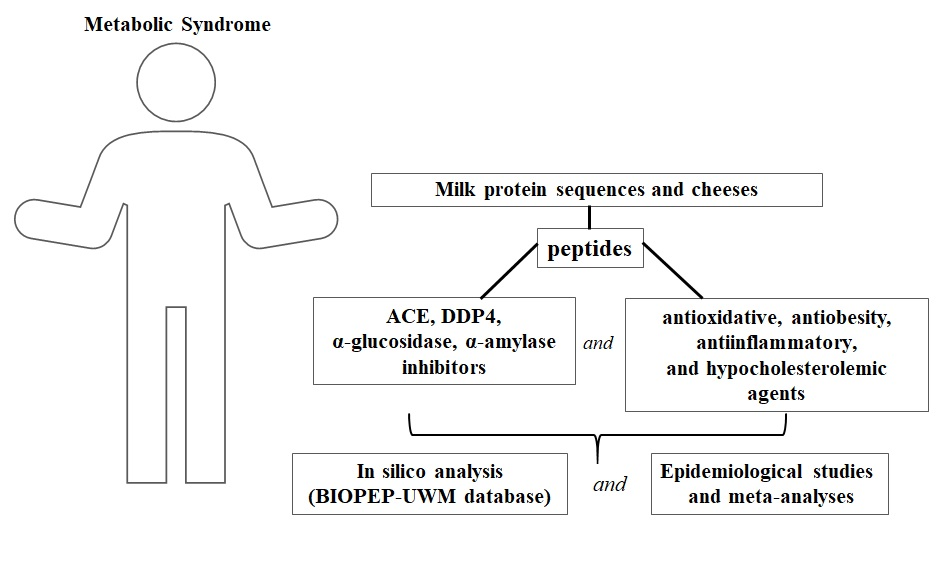
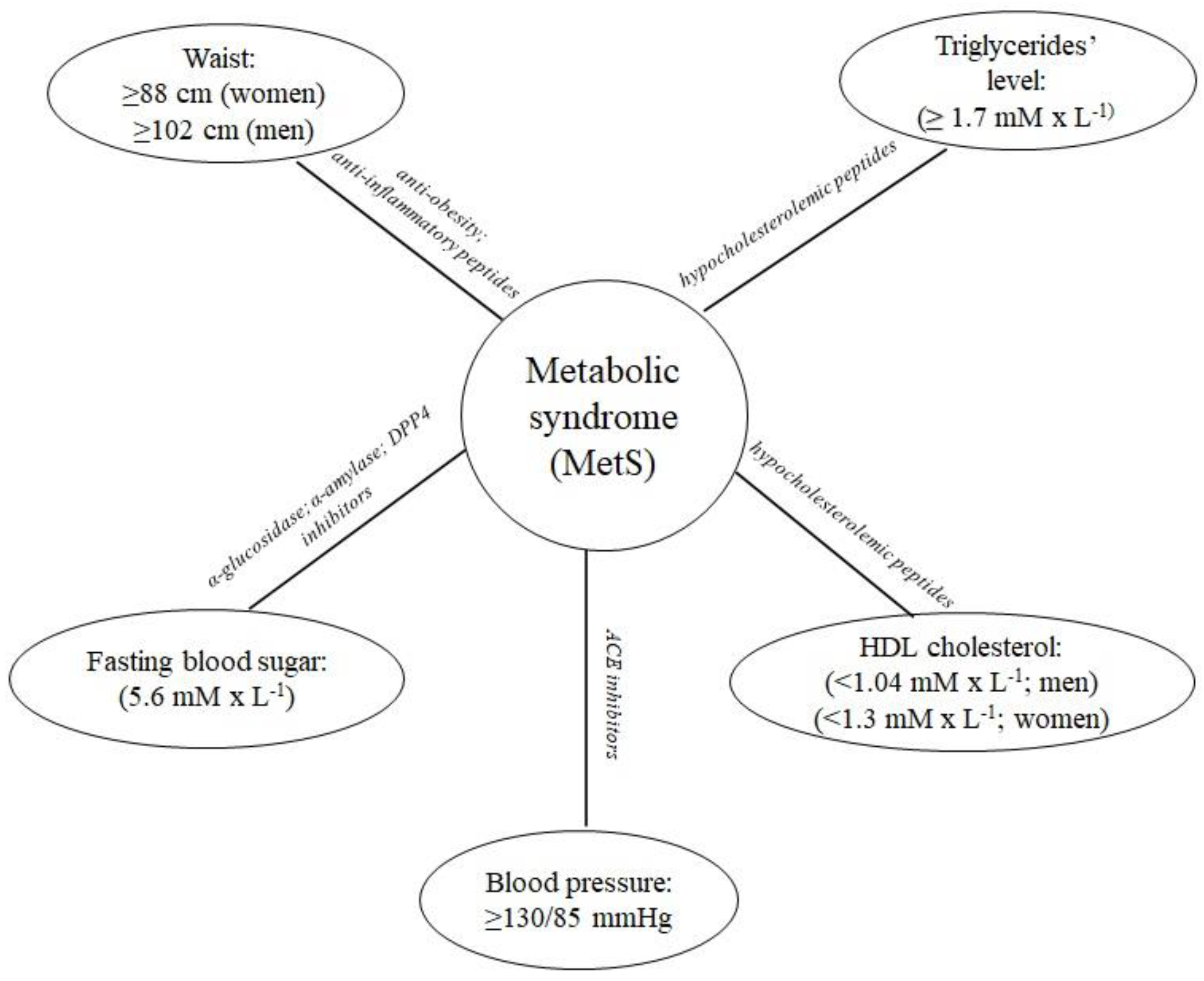
1. Introduction
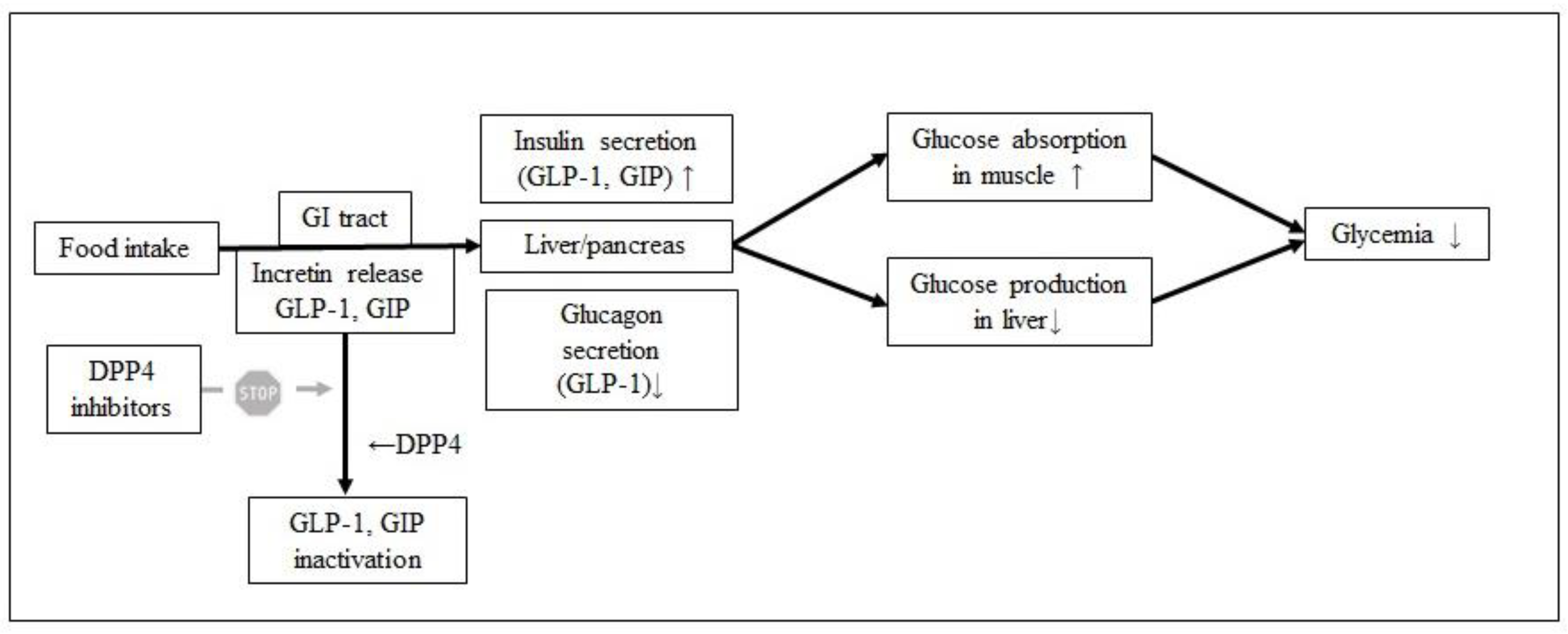
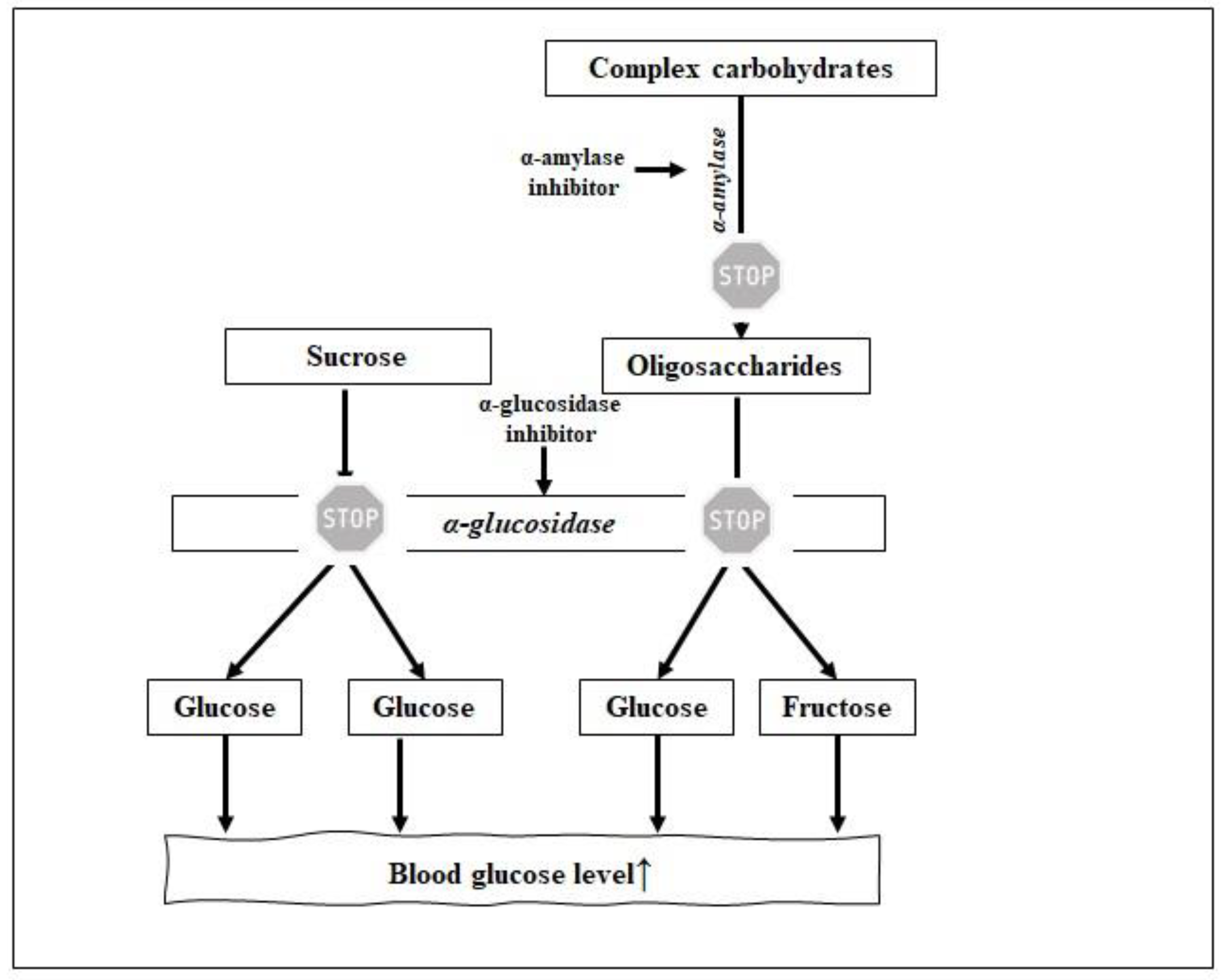
2. Antidiabetic Agents
3. Peptides with Antihypertensive Activity
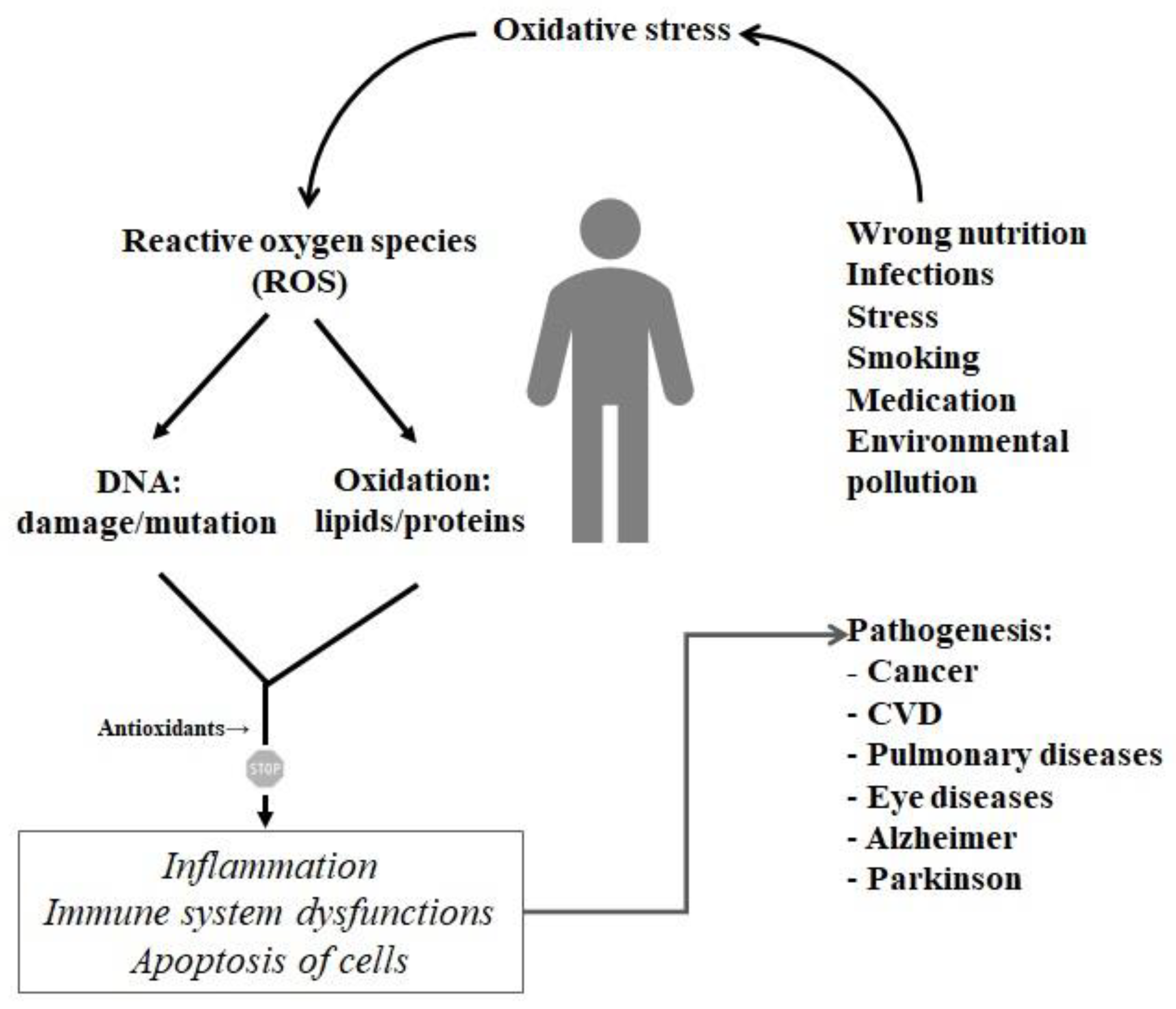
4. Antioxidative Peptides
5. Hypocholesterolemic Peptides
6. Antiobesity and Anti-Inflammatory Peptides
7. Multi-Activity of Peptides in MetS Prevention: A Screening Using BIOPEP-UWM Database
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACE | Angiotensin-converting enzyme (EC 3.4.15.1) |
| ApoE | ndogenous apoprotein E |
| BHA | butylated hydroxyanisole |
| BIOPEP-UWM | database of bioactive peptide sequences [68] |
| Boc | tert-butyloxycarbonyl |
| CHD | coronary heart disease |
| CVD | cardiovascular disease |
| DPP4 | dipeptidyl peptidase IV (EC 3.4.14.5) |
| DPPH | 2,2-diphenyl-1-picryl-hydrazyl) |
| Fmoc | fluorenylmethyloxycarbonyl |
| GIP | glucose-dependent insulinotropic polypeptide |
| GLP-1 | glucagon-like peptide 1 |
| IP3 | inositol 1,4,5-triphosphate |
| LC-MS/MS | Liquid Chromatography and Tandem Mass Spectrometry |
| LDL cholesterol | low density lipoprotein cholesterol |
| MAPK | mitogen-activated protein kinase |
| MetS | metabolic syndrome |
| NHANES | Health and Nutrition Examination Survey |
| NO | nitric oxide |
| NTR2 | neurotensin receptor 2 |
| ORAC-FL | Oxygen Radical Absorbance Capacity with Fluorescein |
| RAAS | renin-angiotensin-aldosterone system |
| RAS | renin-angiotensin system |
| ROS | reactive oxygen species |
| RR | relative risk |
| SBP | systolic blood pressure |
| SHR | spontaneously hypertensive rats |
| T2D | type 2 diabetes mellitus |
| t-BHQ | tert-butyl hydroquinone |
| VLDL cholesterol | very low density lipoprotein cholesterol |
| WSE | water-soluble cheese extracts |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BIOPEP-UWM ID | Activity | Sequence | Protein | Location |
|---|---|---|---|---|
| 7590/8637 | 1* | AA | κ-casein | (86–87) |
| 8758 | 12 | AE | αS1-casein | (77–78), (131–132) |
| αS2-casein | (77–78) | |||
| 7583/8759 | 1 | AF | β-casein | (189–190) |
| 8193 | 9 | AI | αS2-casein | (42–43) |
| κ-casein | (128–129) | |||
| 8559 | 12 | AL | αS1-casein | (13–14) |
| αS2-casein | (13–14), (96–97), (190–191) | |||
| κ-casein | (13–14), (70–71) | |||
| 3177/7584 | 1 | AP | αS1-casein | (41–42), (191–192) |
| β-casein | (103–104) | |||
| 7742 | 9 | AR | αS1-casein | (15–16) |
| κ-casein | (117–118) | |||
| 8762 | 12 | AS | κ-casein | (147–148) |
| 8763 | 12 | AT | αS2-casein | (80–81) |
| κ-casein | (165–166) | |||
| 8764/8951 | 1 | AV | αS1-casein | (11–12) |
| αS2-casein | (11–12), (131–132) | |||
| β-casein | (177–178) | |||
| κ-casein | (87–88), (159–160), (189–190) | |||
| 7543/8460/8695 | 7 | AW | αS1-casein | (178–179) |
| 3563/7866/8765 | 7 | AY | αS1-casein | (158–159), (173–174) |
| 7606 | 9 | DA | αS1-casein | (172–173), (190–191) |
| 9075 | 9 | DM | αS2-casein | (155–156) |
| β-casein | (184–185) | |||
| 8768 | 12 | DQ | αS1-casein | (66–67) |
| αS2-casein | (125–126) | |||
| 7623 | 9 | EA | αS1-casein | (76-77) |
| β-casein | (100–101) | |||
| κ-casein | (158–159) | |||
| 7622/8770 | 1 | EG | αS1-casein | (140–141) |
| 8771 | 12 | EH | αS2-casein | (20–21) |
| 7826/8772 | 1 | EI | αS1-casein | (85–86), (125–126) |
| αS2-casein | (99–100) | |||
| β-casein | (11–12) | |||
| κ-casein | (139–140), (179–180) | |||
| 7840/8558 | 1 | EK | αS1-casein | (50–51), (207–208) |
| αS2-casein | (38–39), (172–173) | |||
| β-casein | (31–32) | |||
| κ-casein | (33–34) | |||
| 7888 | 11 | EL | αS1-casein | (54–55), (156–157), (163–164) |
| β-casein | (2–3), (5–6), (44–45) | |||
| 8529 | 12 | EP | αS1-casein | (148–149) |
| β-casein | (117–118), (195–196) | |||
| κ-casein | (150–151) | |||
| 8773 | 12 | ES | αS1-casein | (62–63), (78–79) |
| αS2-casein | (27–28), (75–76), (157–158) | |||
| β-casein | (14–15), (21–22) | |||
| κ-casein | (161–162), (175–176) | |||
| 8774 | 12 | ET | αS2-casein | (33–34) |
| 7828/8775 | 1 | EV | αS1-casein | (29–30), (45–46) |
| αS2-casein | (57–58), (78–79), (83–84), (160–161) | |||
| β-casein | (91–92) | |||
| κ-casein | (172–173) | |||
| 7752/8777 | 1 | EY | αS2-casein | (66–67) |
| 3176 | 12 | FA | αS2-casein | (189–190) |
| β-casein | (52–53) | |||
| 9342 | 11 | FC | αS2-casein | (54–55) |
| 7605 | 9 | FG | αS1-casein | (47–48) |
| 8555 | 12 | FL | αS2-casein | (178–179) |
| β-casein | (87–88), (190–191) | |||
| κ-casein | (6–7), (18–19), (76–77) | |||
| 3502/8506 | 1 | FP | αS1-casein | (43–44) |
| αS2-casein | (107–108) | |||
| β-casein | (62–63), (111–112), (157–158), (205–206) | |||
| 8779/9076 | 1 | FQ | β-casein | (33–34) |
| 7592/8780 | 1 | FR | αS1-casein | (165–166) |
| 3556 | 9 | FY | αS1-casein | (160–161), (168–169) |
| αS2-casein | (103–104) | |||
| 7598/8524 | 1 | GA | αS1-casein | (177–178) |
| κ-casein | (20–21) | |||
| 7615/8781 | 1 | GE | β-casein | (10–11) |
| κ-casein | (149–150) | |||
| 7596/8785 | 1 | GI | αS1-casein | (141–142) |
| 7611 | 9 | GK | αS1-casein | (48–49) |
| 7599/8561 | 1 | GL | αS1-casein | (25–26) |
| κ-casein | (60–61) | |||
| 3169/7512 | 1 | GP | αS2-casein | (117–118) |
| β-casein | (64–65), (199–200), (203–204) | |||
| 7607 | 9 | GS | αS1-casein | (60–61), (202–203) |
| αS2-casein | (70–71) | |||
| 7612 | 9 | GT | αS1-casein | (185–186) |
| 7608/8786 | 1 | GV | αS1-casein | (152–153) |
| β-casein | (94–95) | |||
| 3532/8788 | 1 | GY | αS1-casein | (108–109) |
| 3184 | 12 | HA | αS1-casein | (143–144) |
| 8793 | 12 | HI | αS1-casein | (95–96) |
| 7844 | 9 | HK | β-casein | (106–107) |
| 3317/7602/8557 | 7 | HL | β-casein | (134–135) |
| κ-casein | (123–124) | |||
| 7842/8520 | 1 | HP | αS1-casein | (19–20) |
| κ-casein | (119–120), (121–122) | |||
| β-casein | (50–51) | |||
| 8795 | 12 | HS | αS1-casein | (136–137) |
| 8797 | 12 | HV | αS2-casein | (21–22) |
| 3494/8799 | 1 | HY | αS2-casein | (92–93) |
| 7562/8525 | 1 | IA | κ-casein | (43–44), (146–147) |
| 7827 | 9 | IE | β-casein | (30–31) |
| κ-casein | (174–175) | |||
| 7593 | 9 | IF | αS2-casein | (5–6) |
| 7595 | 9 | IG | αS1-casein | (59-60), (151-152), (201-202) |
| αS2-casein | (69–70) | |||
| 8800 | 12 | IH | αS1-casein | (142–143) |
| β-casein | (49–50) | |||
| 8801 | 12 | II | αS2-casein | (29–30) |
| β-casein | (207–208) | |||
| 8802/9079 | 1 | IL | αS1-casein | (5–6) |
| κ-casein | (11–12), (94–95) | |||
| 8804 | 12 | IN | αS2-casein | (43–44), (100–101) |
| β-casein | (26–27) | |||
| κ-casein | (72–73), (143–144), (180–181) | |||
| 7581/8501 | 1 | IP | αS1-casein | (197–198) |
| αS2-casein | (216–217) | |||
| β-casein | (66-67), (74-75) | |||
| κ-casein | (47–48), (129–130), (140–141) | |||
| 8805 | 12 | IQ | αS1-casein | (96–97) |
| αS2-casein | (209–210) | |||
| β-casein | (187–188) | |||
| κ-casein | (49–50) | |||
| 3258/8215/8806 | 7 | IR | κ-casein | (30–31) |
| 3174/7743 | 1 | KA | αS2-casein | (95–96), (203–204) |
| β-casein | (176–177) | |||
| 8134 | 11 | KD | αS1-casein | (57–58) |
| κ-casein | (34–35) | |||
| 7841/8808 | 1 | KE | αS1-casein | (49–50), (98–99), (139–140), (147–148) |
| αS2-casein | (47–48), (56–57) | |||
| β-casein | (99–100), (107–108) | |||
| 7692/8809 | 1 | KF | αS2-casein | (2–3), (106–107), (188–189) |
| β-casein | (32–33) | |||
| 8811 | 12 | KH | αS1-casein | (18–19), (22–23), (94–95) |
| αS2-casein | (91–92) | |||
| β-casein | (105–106) | |||
| 8812 | 12 | KI | αS2-casein | (85–86), (181–182) |
| β-casein | (29–30), (48–49) | |||
| κ-casein | (42–43) | |||
| 8813 | 12 | KK | αS1-casein | (117–118) |
| αS2-casein | (151–152), (164–165), (180–81) | |||
| β-casein | (28–29) | |||
| κ-casein | (132–133) | |||
| 7693 | 9 | KL | αS1-casein | (2–3) |
| αS2-casein | (167–168) | |||
| 7810/8218/8519 | 7 | KP | αS2-casein | (206–207) |
| κ-casein | (67–68), (84–85) | |||
| 7586/8814 | 1 | KR | αS2-casein | (128–129) |
| 8815 | 12 | KS | κ-casein | (3–4), (107–108) |
| 8816 | 12 | KT | αS1-casein | (208–209) |
| αS2-casein | (152–153), (165–166), (196–197), (212–213) | |||
| κ-casein | (137–138) | |||
| 8817 | 12 | KV | αS1-casein | (51–52), (120–121) |
| αS2-casein | (214–215) | |||
| β-casein | (97–98), (169–170) | |||
| 7691/8819 | 1 | KY | αS1-casein | (118–119) |
| β-casein | (113–114) | |||
| κ-casein | (45–46) | |||
| 3175/7585 | 1 | LA | αS1-casein | (14–15), (157–158) |
| αS2-casein | (10–11), (14–15) | |||
| κ-casein | (12–13) | |||
| 3551 | 9 | LF | αS1-casein | (164–165) |
| 7619 | 9 | LG | αS1-casein | (107–108), (184–185) |
| β-casein | (198–199) | |||
| κ-casein | (19–20) | |||
| 3305/8820 | 6 | LH | αS1-casein | (135–136) |
| β-casein | (133–134) | |||
| 8821 | 12 | LI | αS1-casein | (4–5) |
| κ-casein | (71–72) | |||
| 8217 | 11 | LK | αS1-casein | (116–117) |
| αS2-casein | (179–180), (195–196) | |||
| 3182 | 12 | LL | αS1-casein | (3–4), (35–36), (113–114) |
| αS2-casein | (9–10) | |||
| β-casein | (139–140), (191–192) | |||
| 7832/8823 | 1 | LN | αS1-casein | (31–32) |
| αS2-casein | (97–98), (121–122), (138–139), (176–177) | |||
| β-casein | (6–7) | |||
| κ-casein | (61–62) | |||
| 3180 | 12 | LP | αS1-casein | (26–27) |
| αS2-casein | (191–192) | |||
| β-casein | (70–71), (135–136), (137–138), (151–152), (171–172) | |||
| κ-casein | (16–17), (77–78) | |||
| 7831 | 9 | LQ | αS2-casein | (111–112) |
| β-casein | (45–46), (88–89), (140–141) | |||
| κ-casein | (95–96) | |||
| 9213 | 9 | LR | αS1-casein | (36–37), (114–115) |
| 8824 | 12 | LT | αS1-casein | (6–7) |
| αS2-casein | (168–169) | |||
| β-casein | (77–78), (125–126), (127–128) | |||
| κ-casein | (14–15) | |||
| 8825 | 12 | LV | αS1-casein | (9–10) |
| β-casein | (58–59) | |||
| κ-casein | (7–8) | |||
| 3389/8462/8688 | 7 | LW | αS1-casein | (213–214) |
| 3381/7872 | 2 | LY | αS2-casein | (114–115) |
| β-casein | (192–193) | |||
| 3173 | 12 | MA | αS2-casein | (41–42) |
| β-casein | (102–103) | |||
| κ-casein | (116–117), (127–128) | |||
| 7839/8826 | 1 | ME | αS1-casein | (69–70), (75–76) |
| αS2-casein | (19–20), (156–157) | |||
| 3385/8827 | 1 | MF | β-casein | (156–157) |
| 7609/8828 | 1 | MG | β-casein | (93–94) |
| 8829 | 12 | MH | β-casein | (144–145) |
| 8830 | 12 | MI | αS1-casein | (150–151) |
| 8831 | 12 | MK | αS1-casein | (1–2), (138–139) |
| αS2-casein | (1–2), (205–206) | |||
| κ-casein | (2–3) | |||
| 8833/9085/9086 | 7 | MM | κ-casein | (1–2) |
| 3171 | 12 | MP | αS1-casein | (211–212) |
| β-casein | (109–110), (185–186) | |||
| 8839 | 12 | NA | αS2-casein | (61–62), (130–131) |
| 8841 | 12 | NE | αS1-casein | (32–33), (53–54) |
| αS2-casein | (63–64), (98–99) | |||
| 7683/8842 | 1 | NF | αS2-casein | (177–178) |
| 7698 | 9 | NK | β-casein | (27–28) |
| 8845 | 12 | NL | αS1-casein | (34–35) |
| αS2-casein | (49–50) | |||
| β-casein | (132–133) | |||
| 8846 | 12 | NM | αS2-casein | (40–41) |
| 8847 | 12 | NN | κ-casein | (73–74) |
| 8530 | 12 | NP | αS1-casein | (199–200) |
| αS2-casein | (44–45), (122–123) | |||
| 8848 | 12 | NQ | αS1-casein | (154–155) |
| αS2-casein | (101–102) | |||
| κ-casein | (25–26), (74–75), (134–135) | |||
| 8849 | 12 | NR | αS2-casein | (139–140), (174–175) |
| 8850 | 12 | NT | αS2-casein | (17–18) |
| κ-casein | (102–103), (144–145), (181–182) | |||
| 8851 | 12 | NV | β-casein | (7–8) |
| 7682/8853 | 1 | NY | κ-casein | (62–63) |
| 3179 | 12 | PA | κ-casein | (85–86), (91–92), (105–106) |
| 8854 | 12 | PF | αS1-casein | (42–43) |
| β-casein | (51–52), (61–62), (86–87), (110–111), (118–119), (204–205) | |||
| κ-casein | (17–18) | |||
| 7625/8855 | 1 | PG | β-casein | (9–10), (63–64) |
| 7843/8856 | 1 | PH | β-casein | (147–148) |
| κ-casein | (120–121), (122–123) | |||
| 8857 | 12 | PI | αS1-casein | (20–21), (200–201) |
| αS2-casein | (118–119), (133–134) | |||
| β-casein | (65–66), (186–187), (206–207) | |||
| κ-casein | (29–30), (48–49) | |||
| 8858 | 12 | PK | αS1-casein | (17–18) |
| αS2-casein | (211–212) | |||
| β-casein | (104–105), (112–113) | |||
| κ-casein | (131–132) | |||
| 7513/8638 | 1 | PL | αS1-casein | (183–184), (212–213) |
| β-casein | (76–77), (136–137), (138–139), (150–151) | |||
| 8859 | 12 | PM | αS1-casein | (149–150) |
| 8860 | 12 | PN | αS1-casein | (88–89), (128–129), (198–199) |
| β-casein | (67–68) | |||
| 3170/7836 | 1 | PP | β-casein | (75–76), (85–86), (152–153), (158–159) |
| κ-casein | (130–131), (177–178) | |||
| 7837/8861 | 1 | PQ | αS1-casein | (27–28), (122–123) |
| αS2-casein | (108–109), (192–193) | |||
| β-casein | (71–72), (159–160), (174–175), (181–182) | |||
| 8862 | 12 | PS | αS1-casein | (102–103), (175–176), (192–193) |
| αS2-casein | (45–46) | |||
| κ-casein | (57–58) | |||
| 7833/8863 | 1 | PT | αS2-casein | (136–137) |
| β-casein | (153–154) | |||
| κ-casein | (113–114), (141–142), (151–152), (155–156) | |||
| 8864 | 12 | PV | β-casein | (81–82), (115–116), (172–173), (196–197), (200–201) |
| κ-casein | (68–69) | |||
| 8190/8865 | 6 | PW | αS2-casein | (123–124), (207–208) |
| 8866 | 12 | PY | αS2-casein | (217–218) |
| β-casein | (179–180) | |||
| κ-casein | (78–79), (80–81) | |||
| 8867 | 12 | QA | αS1-casein | (67–68) |
| β-casein | (188–189) | |||
| κ-casein | (110–111) | |||
| 8868 | 12 | QD | β-casein | (46–47) |
| κ-casein | (135–136) | |||
| 8869 | 12 | QE | αS1-casein | (28–29), (155–156) |
| αS2-casein | (32–33), (37–38) | |||
| β-casein | (194–195) | |||
| κ-casein | (22–23), (26–27) | |||
| 8870 | 12 | QF | αS1-casein | (167–168) |
| αS2-casein | (102–103) | |||
| κ-casein | (75–76) | |||
| 7617/8871 | 1 | QG | αS1-casein | (24–25) |
| αS2-casein | (116–117) | |||
| 8872 | 12 | QH | αS2-casein | (200–201) |
| 8873 | 12 | QI | κ-casein | (93–94) |
| 7680 | 9 | QK | αS1-casein | (93–94), (97–98), (146–147) |
| αS2-casein | (94–95), (105–106), (187–188), (202–203) | |||
| β-casein | (175–176) | |||
| κ-casein | (66–67) | |||
| 8874 | 12 | QL | αS1-casein | (112–113), (123–124), (170–171) |
| αS2-casein | (142–143) | |||
| 8875 | 12 | QN | β-casein | (72–73) |
| κ-casein | (24–25) | |||
| 8532 | 12 | QP | αS2-casein | (210–211) |
| β-casein | (89–90), (146–147), (149–150) | |||
| κ-casein | (28–29), (112–113) | |||
| 8876 | 12 | αS1-casein | (145–146) | |
| β-casein | (38–39), (39–40) | |||
| κ-casein | (65–66) | |||
| 8877 | 12 | QS | β-casein | (34–35), (56–57), (123–124), (141–142), (160–161), (167–168) |
| 8878 | 12 | QT | β-casein | (40–41), (54–55), (79–80) |
| 8879 | 12 | QV | αS2-casein | (126–127) |
| κ-casein | (98–99), (184–185) | |||
| 8880 | 12 | QW | κ-casein | (96–97) |
| 8881 | 12 | QY | αS1-casein | (187–188) |
| αS2-casein | (109–110), (112–113), (193–194) | |||
| κ-casein | (50–51) | |||
| 3489 | 9 | RF | αS1-casein | (37–38) |
| κ-casein | (37–38) | |||
| 8882/9173 | 1 | RG | β-casein | (202–203) |
| 8883 | 12 | RH | κ-casein | (118–119) |
| 8884 | 12 | RI | β-casein | (25–26) |
| 3257/8886 | 1 | RL | αS1-casein | (115–116), (134–135) |
| αS2-casein | (175–176) | |||
| 8888 | 12 | RN | αS2-casein | (60–61), (129–130) |
| 7582/8518 | 1 | RP | αS1-casein | (16–17) |
| 3380 | 9 | RY | αS1-casein | (105–106) |
| αS2-casein | (185–186), (220–221) | |||
| κ-casein | (55–56) | |||
| 7685/8891 | 1 | SF | αS1-casein | (193–194) |
| κ-casein | (4–5), (125–126) | |||
| 7618 | 9 | SG | αS1-casein | (176–177) |
| κ-casein | (148–149) | |||
| 8893 | 12 | SI | αS1-casein | (79–80) |
| αS2-casein | (28–29), (68–69) | |||
| β-casein | (22–23) | |||
| 8894 | 12 | SK | αS1-casein | (56–57) |
| αS2-casein | (46–47), (150–151) | |||
| β-casein | (96–97), (168–169) | |||
| 8560 | 12 | SL | β-casein | (15–16), (57–58), (69–70), (124–125), (164–165) |
| 8505 | 12 | SP | κ-casein | (90–91), (170–171), (176–177) |
| 9184 | 9 | ST | αS1-casein | (63–64) |
| αS2-casein | (52–53), (144–145), (158–159) | |||
| κ-casein | (153–154), (162–163), (187–188) | |||
| 8895 | 12 | SV | αS1-casein | (90–91) |
| β-casein | (161–162) | |||
| 8896 | 12 | SW | β-casein | (142–143) |
| 7684/8897 | 1 | SY | κ-casein | (58–59) |
| 8531 | 12 | TA | κ-casein | (188–189) |
| 8898 | 12 | TD | αS1-casein | (189–190) |
| β-casein | (128–129) | |||
| 7830/8899 | 1 | TE | αS1-casein | (64–65) |
| αS2-casein | (81–82), (159–160), (169–170) | |||
| β-casein | (41–42), (120–121) | |||
| κ-casein | (138–-139), (157–158) | |||
| 8185/8900 | 1 | TF | αS2-casein | (53–54) |
| 8903 | 12 | TI | κ-casein | (10–11), (142–143), (145–146) |
| 8904 | 12 | TK | αS2-casein | (163–164), (166–167), (213–214) |
| 8905 | 12 | TL | αS2-casein | (137–138) |
| β-casein | (126–127) | |||
| κ-casein | (15–16), (166–167) | |||
| 8906 | 12 | TM | αS1-casein | (210–211) |
| αS2-casein | (18–19) | |||
| κ-casein | (115–116) | |||
| 8503/9073 | 1 | TP | αS2-casein | (135–136) |
| β-casein | (80–81) | |||
| κ-casein | (154–155) | |||
| 7834/8908 | 1 | TQ | αS1-casein | (186–187) |
| β-casein | (55–56), (78–79) | |||
| 8909 | 12 | TR | β-casein | (24–25) |
| 8910 | 12 | TS | αS2-casein | (145–146) |
| κ-casein | (152–153), (186–187) | |||
| 8911 | 12 | TT | αS1-casein | (209–210) |
| κ-casein | (114–115), (156–157) | |||
| 8912 | 12 | TV | αS2-casein | (87–88), (153–154), (197–198) |
| β-casein | (154–155) | |||
| κ-casein | (103–104), (163–164), (182–183) | |||
| 8219/8914 | 6 | TY | αS2-casein | (34–35) |
| 3172 | 12 | VA | αS1-casein | (10–11), (12–13), (40–41) |
| αS2-casein | (12–13), (79–80) | |||
| κ-casein | (69–70), (164–165) | |||
| 8915 | 12 | VD | αS2-casein | (88–89), (154–155) |
| 7829/8916 | 1 | VE | αS1-casein | (91–92) |
| β-casein | (13–14), (116–117), (130–131) | |||
| κ-casein | (160–161) | |||
| 3384/8917 | 1 | VF | αS1-casein | (46–47) |
| αS2-casein | (161–162) | |||
| 8920 | 12 | VI | αS2-casein | (215–216) |
| κ-casein | (173–174) | |||
| 7558/8921 | 1 | VK | αS2-casein | (84–85), (127–128) |
| β-casein | (98–99) | |||
| 8922 | 12 | VL | αS1-casein | (30–31) |
| αS2-casein | (120–121) | |||
| β-casein | (162–163), (170–171), (197–198) | |||
| κ-casein | (52–53), (99–100) | |||
| 8923 | 12 | VM | β-casein | (92–93), (155–156) |
| 8924 | 12 | VN | αS1-casein | (52–53), (153–154) |
| 3181/7587 | 1 | VP | αS1-casein | (87–88), (101–102), (121–122), (127–128), (182–183) |
| αS2-casein | (132–133) | |||
| β-casein | (8–9), (84–85), (173–174), (178–179) | |||
| κ-casein | (104–105) | |||
| 8925 | 12 | VQ | κ-casein | (183–184) |
| 7628/8594 | 1 | VR | αS2-casein | (59–60), (219–220) |
| β-casein | (201–202) | |||
| κ-casein | (88–89) | |||
| 8926 | 12 | VS | αS2-casein | (22–23) |
| β-casein | (95–96) | |||
| 8927 | 12 | VT | κ-casein | (9–10), (185–186) |
| 3183 | 12 | VV | αS2-casein | (58–59) |
| β-casein | (82–83), (83–84) | |||
| κ-casein | (8–9) | |||
| 3492/8224/8929 | 7 | VY | αS2-casein | (198–199) |
| β-casein | (59–60) | |||
| 8930 | 12 | WD | αS2-casein | (124–125) |
| 8679 | 12 | WI | αS2-casein | (208–209) |
| 8682/9090 | 1 | WM | β-casein | (143–144) |
| 8678 | 12 | WQ | κ-casein | (97–98) |
| 7898/8683 | 6 | WY | αS1-casein | (179–180) |
| 7589/8932 | 1 | YA | κ-casein | (82–83) |
| 8935 | 12 | YF | αS1-casein | (159–160) |
| 3553/8936 | 1 | YG | κ-casein | (59–60) |
| 8938 | 12 | YI | κ-casein | (46–47) |
| 7697/8939 | 1 | YK | αS1-casein | (119–120) |
| αS2-casein | (35–36) | |||
| 8940 | 12 | YL | αS1-casein | (106–107), (109–110) |
| αS2-casein | (110–111), (113–114), (194–195), (221–222) | |||
| 3666/8521/9548 | 8 | YP | αS1-casein | (161–162), (174–175) |
| β-casein | (60–61), (114–115), (180–181) | |||
| κ-casein | (56–57), (79–80) | |||
| 8943 | 12 | YQ | αS1-casein | (169–170) |
| αS2-casein | (93–94), (104–105), (115–116), (186–187), (199–200) | |||
| β-casein | (193–194) | |||
| κ-casein | (64–65) | |||
| 8945 | 12 | YS | αS2-casein | (67–68) |
| 8696 | 12 | YT | αS1-casein | (188–189) |
| 8946/9077 | 1 | YV | αS1-casein | (181–182) |
| αS2-casein | (218–219) | |||
| κ-casein | (51–52) | |||
| 8948 | 12 | YY | αS1-casein | (180–181) |
| κ-casein | (63–64), (81–82) | |||
| 9042 | 9 | AFL | β-casein | (189–191) |
| 3597 | 9 | AIP | κ-casein | (128–130) |
| 9029 | 9 | ALP | αS2-casein | (190–192) |
| 3370 | 9 | AVP | αS2-casein | (131–133) |
| β-casein | (177–179) | |||
| 9359 | 11 | CLV | αS1-casein | (8–10) |
| 7823 | 9 | FAL | αS2-casein | (189–191) |
| 7560 | 9 | FFL | κ-casein | (5–7) |
| 3377 | 9 | FGK | αS1-casein | (47–49) |
| 7554 | 9 | GEP | κ-casein | (149–151) |
| 7509 | 9 | GLP | αS1-casein | (25–27) |
| 7545/9116 | 1 | GPV | β-casein | (199–201) |
| 3311 | 11 | HPH | κ-casein | (119–121), (121–123) |
| 9027 | 9 | HQG | αS1-casein | (23–25) |
| 7626 | 9 | IAK | κ-casein | (43–45) |
| 3167 | 12 | IPI | κ-casein | (47–49) |
| 3522 | 9 | IPP | β-casein | (74–76) |
| κ-casein | (129–131) | |||
| 7803 | 9 | IPY | αS2-casein | (216–218) |
| 8184/8693 | 1 | IQP | αS2-casein | (209–211) |
| 8149/8180 | 2 | IQY | κ-casein | (49–51) |
| 9365 | 11 | KKY | αS1-casein | (117–119) |
| 8133 | 11 | KVI | αS2-casein | (214–216) |
| 3558 | 9 | LAY | αS1-casein | (157–159) |
| 9031 | 9 | LEE | β-casein | (3–5) |
| 9206 | 9 | LFR | αS1-casein | (164–166) |
| 7508 | 9 | LGP | β-casein | (198–200) |
| 7995 | 11 | LHL | β-casein | (133–135) |
| 8000 | 11 | LHS | αS1-casein | (135–137) |
| 8484 | 11 | LLR | αS1-casein | (35–37), (113–115) |
| 8509 | 9 | LNF | αS2-casein | (176–178) |
| 3544 | 9 | LNP | αS2-casein | (121–123) |
| 9026 | 9 | LNY | κ-casein | (61–63) |
| 9040 | 9 | LPF | κ-casein | (16–18) |
| 8616 | 12 | LPL | β-casein | (135–137), (137–139) |
| 3391 | 9 | LPP | β-casein | (151–153) |
| 9339 | 12 | LPQ | αS1-casein | (26–28) |
| αS2-casein | (191–193) | |||
| β-casein | (70–72) | |||
| 3542/8689 | 1 | LQP | β-casein | (88–90) |
| 9163 | 11 | LTC | αS1-casein | (6–8) |
| 8402 | 9 | LVY | β-casein | (58–60) |
| 9241 | 9 | MAP | β-casein | (102–104) |
| 8958 | 9 | MKP | αS2-casein | (205–207) |
| 9363 | 11 | NEN | αS1-casein | (32–34) |
| 7965 | 11 | NYY | κ-casein | (62–64) |
| 8139 | 11 | PEL | αS1-casein | (162–164) |
| 8029 | 11 | PHL | κ-casein | (122–124) |
| 8032 | 11 | PHQ | β-casein | (147–149) |
| 3094/7510 | 5 | PLG | αS1-casein | (183–185) |
| 2664 | 9 | PLP | β-casein | (136–138), (150–152) |
| 3531 | 9 | PLW | αS1-casein | (212–214) |
| 7645 | 9 | PPK | κ-casein | (130–132) |
| 8652 | 12 | PPL | β-casein | (75–77) |
| 3373 | 9 | PQR | β-casein | (181–183) |
| 7559 | 9 | PSY | κ-casein | (57–59) |
| 8040 | 11 | PWD | αS2-casein | (123–125) |
| 8044 | 11 | PWI | αS2-casein | (207–209) |
| 3372 | 9 | PYP | β-casein | (179–181) |
| κ-casein | (78–80) | |||
| 3307 | 11 | PYY | κ-casein | (80–82) |
| 9146 | 9 | QGP | αS2-casein | (116–118) |
| 9543 | 9 | RYL | αS1-casein | (105–107) |
| αS2-casein | (220–222) | |||
| 8382/8474 | 2 | RYQ | αS2-casein | (185–187) |
| 7498 | 9 | TVY | αS2-casein | (197–199) |
| 3521 | 9 | VAP | αS1-casein | (40–42) |
| 7635 | 9 | VAV | αS1-casein | (10–12) |
| 2653 | 9 | VLP | β-casein | (170–172) |
| 8347 | 12 | VPL | αS1-casein | (182–184) |
| 3524 | 9 | VPP | β-casein | (84–86) |
| 9046 | 9 | VQV | κ-casein | (183–185) |
| 8384 | 9 | VRY | αS2-casein | (219–221) |
| 3505 | 9 | VYP | β-casein | (59–61) |
| 7962 | 11 | WYY | αS1-casein | (179–181) |
| 7963 | 11 | YFY | αS1-casein | (159–161) |
| 3973 | 9 | YGL | κ-casein | (59–61) |
| 7959 | 11 | YLY | αS2-casein | (113–115) |
| 2870 | 15 | YPF | β-casein | (60–62) |
| 8617/9550 | 3 | YPY | κ-casein | (79–81) |
| 8487 | 11 | YQL | αS1-casein | (169–171) |
| 8150 | 11 | YVL | κ-casein | (51–53) |
| 7634 | 9 | YVP | αS1-casein | (181–183) |
| 7939 | 11 | YYA | κ-casein | (81–83) |
| 7948 | 11 | YYQ | κ-casein | (63–65) |
| 7942 | 11 | YYV | αS1-casein | (180–182) |
| 8369 | 9 | AFLL | β-casein | (189–192) |
| 7668 | 9 | AIPP | κ-casein | (128–131) |
| 8380/8472 | 2 | AYPS | αS1-casein | (173–176) |
| 8591 | 12 | FLQP | β-casein | (87–90) |
| 7499 | 9 | FVAP | αS1-casein | (39–42) |
| 8489 | 11 | FYQL | αS1-casein | (168–171) |
| 8371/8467 | 2 | GTQY | αS1-casein | (185–188) |
| 3306 | 11 | HPHL | κ-casein | (121–124) |
| 7661 | 9 | LFRQ | αS1-casein | (164–167) |
| 2668 | 9 | LHLP | β-casein | (133–136) |
| 2665 | 9 | LPLP | β-casein | (135–138) |
| 3397 | 9 | LQSW | β-casein | (140–143) |
| 9236 | 9 | LRFF | αS1-casein | (36–39) |
| 3331 | 9 | LVYP | β-casein | (58–61) |
| 3374 | 9 | PFPE | αS1-casein | (42–45) |
| 7896 | 11 | PYPQ | β-casein | (179–182) |
| 8357 | 9 | RYLG | αS1-casein | (105–108) |
| 8378/8470 | 2 | RYPS | κ-casein | (55–58) |
| 8221 | 11 | TSTA | κ-casein | (186–189) |
| 8483 | 11 | TVYQ | αS2-casein | (197–200) |
| 8593 | 12 | VLGP | β-casein | (197–200) |
| 8309 | 9 | VRSP | κ-casein | (88–91) |
| 7801 | 9 | VRYL | αS2-casein | (219–222) |
| 8308 | 9 | VVPP | β-casein | (83–86) |
| 8592 | 12 | WIQP | αS2-casein | (208–211) |
| 8381/8473 | 2 | YAKP | κ-casein | (82–85) |
| 8485 | 11 | YGLN | κ-casein | (59–62) |
| 9420/9421 | 2 | YLGY | αS1-casein | (106–109) |
| 8482 | 11 | YLKT | αS2-casein | (194–197) |
| 8140 | 11 | YPEL | αS1-casein | (161–164) |
| 2868 | 15 | YPFP | β-casein | (60–63) |
| 3218/8615 | 4 | YPYY | κ-casein | (79–82) |
| 7878 | 11 | YQEP | β-casein | (193–196) |
| 8477 | 11 | YQLD | αS1-casein | (169–172) |
| 3399/7660 | 9 | AMKPW | αS2-casein | (204–208) |
| 9229 | 9 | ARHPH | κ-casein | (117–121) |
| 3371 | 9 | AVPYP | β-casein | (177–181) |
| 3519 | 9 | AYFYP | αS1-casein | (158–162) |
| 3497 | 9 | FFVAP | αS1-casein | (38–42) |
| 8368 | 9 | FPIIV | β-casein | (205–209) |
| 9378 | 9 | FPPQS | β-casein | (157–161) |
| 8141 | 11 | FYPEL | αS1-casein | (160–164) |
| 9375 | 9 | GLPQE | αS1-casein | (25–29) |
| 8478 | 11 | GYLEQ | αS1-casein | (108–112) |
| 2666 | 9 | HLPLP | β-casein | (134–138) |
| 8613 | 12 | IPIQY | κ-casein | (47–51) |
| 7627 | 9 | KDERF | κ-casein | (34–38) |
| 9552 | 9 | KFPQY | αS2-casein | (106–110) |
| 8618 | 12 | LPLPL | β-casein | (135–139) |
| 7631/8614 | 1 | LPYPY | κ-casein | (77–81) |
| 2671 | 9 | NLHLP | β-casein | (132–136) |
| 8157 | 9 | PGPIP | β-casein | (63–67) |
| 9242 | 9 | PLPLL | β-casein | (136–140) |
| 9243 | 9 | PPEIN | κ-casein | (177–181) |
| 8373/8469 | 2 | RHPHP | κ-casein | (118–122) |
| 9244 | 9 | RINKK | β-casein | (25–29) |
| 8481 | 11 | RLKKY | αS1-casein | (115–119) |
| 8358 | 9 | RYLGY | αS1-casein | (105–109) |
| 8375 | 9 | RYQKF | αS2-casein | (185–189) |
| 9246 | 9 | SLPQN | β-casein | (69–73) |
| 3332 | 9 | SLVYP | β-casein | (57–61) |
| 8379/8471 | 2 | SRYPS | κ-casein | (54–58) |
| 7485 | 9 | TKVIP | αS2-casein | (213–217) |
| 8278/9556 | 6 | VPYPQ | β-casein | (178–182) |
| 8370/8466 | 2 | YAKPA | κ-casein | (82–86) |
| 8480 | 11 | YPELF | αS1-casein | (161–165) |
| 3263 | 13 | YPFPG | β-casein | (60–64) |
| 8486 | 11 | YQKFP | αS2-casein | (104–108) |
| 8372/8468 | 2 | ARHPHP | κ-casein | (117–122) |
| 3501 | 9 | AYFYPE | αS1-casein | (158–163) |
| 7493 | 9 | DKIHPF | β-casein | (47–52) |
| 7629 | 9 | EKDERF | κ-casein | (33–38) |
| 3511 | 9 | EMPFPK | β-casein | (108–113) |
| 7480 | 9 | FALPQY | αS2-casein | (189–194) |
| 7487 | 9 | KVLPVP | β-casein | (169–174) |
| 3513 | 9 | LAYFYP | αS1-casein | (157–162) |
| 2667 | 9 | LHLPLP | β-casein | (133–138) |
| 8179 | 9 | LKKISQ | αS2-casein | (179–184) |
| 9238 | 9 | LVYPFP | β-casein | (58–63) |
| 8152/8178 | 2 | PYVRYL | αS2-casein | (217–222) |
| 3426 | 9 | QSLVYP | β-casein | (56–61) |
| 8479 | 11 | RDMPIQ | β-casein | (183–188) |
| 3128 | 14 | RYLGYL | αS1-casein | (105–110) |
| 3217 | 14 | SRYPSY | κ-casein | (54–59) |
| 3127/3530 | 5 | TTMPLW | αS1-casein | (209–214) |
| 9248 | 9 | VLSRYP | κ-casein | (52–57) |
| 8658 | 12 | VPITPT | αS2-casein | (132–137) |
| 7481 | 9 | VTSTAV | κ-casein | (185–190) |
| 9251 | 9 | VVVPPF | β-casein | (82–87) |
| 3504 | 9 | VYPFPG | β-casein | (59–64) |
| 7887/8376 | 2 | YFYPEL | αS1-casein | (159–164) |
| 7630 | 9 | YIPIQY | κ-casein | (46–51) |
| 3500 | 9 | YKVPQL | αS1-casein | (119–124) |
| 3265 | 15 | YLGYLE | αS1-casein | (106–111) |
| 9252 | 12 | YPVEPF | β-casein | (114–119) |
| 7662/9253 | 9 | YQEPVL | β-casein | (193–198) |
| 8383 | 9 | YYQQKP | κ-casein | (63–68) |
| 3480/7875 | 2 | AVPYPQR | β-casein | (177–183) |
| 8359 | 9 | AYFYPEL | αS1-casein | (158–164) |
| 7496 | 9 | FPEVFGK | αS1-casein | (43–49) |
| 8659 | 12 | FPGPIPN | β-casein | (62–68) |
| 7659 | 9 | FPQYLQY | αS2-casein | (107–113) |
| 8181 | 9 | FSDKIAK | κ-casein | (39–45) |
| 8159 | 9 | GPFPIIV | β-casein | (203–209) |
| 8285 | 11 | IPIQYVL | κ-casein | (47–53) |
| 9373 | 9 | KEDVPSE | αS1-casein | (98–104) |
| 9234 | 9 | KKYKVPQ | αS1-casein | (117–123) |
| 3498 | 9 | KVLPVPQ | β-casein | (169–175) |
| 7565 | 9 | LHLPLPL | β-casein | (133–139) |
| 8657 | 12 | LPQNIPP | β-casein | (70–76) |
| 8155 | 9 | LTLTDVE | β-casein | (125–131) |
| 2669 | 9 | NLHLPLP | β-casein | (132–138) |
| 9058 | 12 | PQNIPPL | β-casein | (71–77) |
| 3473 | 13 | RYLGYLE | αS1-casein | (105–111) |
| 8476 | 11 | TIASGEP | κ-casein | (145–151) |
| 3333 | 9 | TQSLVYP | β-casein | (55–61) |
| 7877 | 11 | VLPVPQK | β-casein | (170–176) |
| 8475 | 11 | VLSRYPS | κ-casein | (52–58) |
| 9249 | 12 | VPITPTL | αS2-casein | (132–138) |
| 9250 | 9 | VPSERYL | αS1-casein | (101–107) |
| 3262/7665 | 5 | YPFPGPI | β-casein | (60–66) |
| 3214 | 15 | YPSYGLN | κ-casein | (56–62) |
| 9254 | 9 | YQKFPQY | αS2-casein | (104–110) |
| 8377 | 9 | AYFYPELF | αS1-casein | (158–165) |
| 3512 | 9 | DAYPSGAW | αS1-casein | (172–179) |
| 9539 | 9 | EKVNELSK | αS1-casein | (50–57) |
| 2672 | 9 | ENLHLPLP | β-casein | (131–138) |
| 7658/8151 | 2 | FALPQYLK | αS2-casein | (189–196) |
| 7876 | 11 | KVLPVPQK | β-casein | (169–176) |
| 9235 | 12 | LPQNIPPL | β-casein | (70–77) |
| 3398 | 9 | MKPWIQPK | αS2-casein | (205–212) |
| 7802 | 9 | NMAINPSK | αS2-casein | (40–47) |
| 9168/9273 | 9 | PFPEVFGK | αS1-casein | (42–49) |
| 3334 | 9 | QTQSLVYP | β-casein | (54–61) |
| 8160 | 9 | RGPFPIIV | β-casein | (202–209) |
| 7796 | 11 | VKEAMAPK | β-casein | (98–105) |
| 7492 | 9 | VYPFPGPI | β-casein | (59–66) |
| 2869/7501 | 5 | YPFPGPIP | β-casein | (60–67) |
| 7879 | 11 | YQEPVLGP | β-casein | (193–200) |
| 7804 | 9 | ALNEINQFY | αS2-casein | (96–104) |
| 3395/7497 | 9 | AMKPWIQPK | αS2-casein | (204–212) |
| 3335 | 9 | AQTQSLVYP | β-casein | (53–61) |
| 8660 | 12 | IPPLTQTPV | β-casein | (74–82) |
| 9380 | 9 | LGPVRGPFP | β-casein | (198–206) |
| 7494 | 9 | LNVPGEIVE | β-casein | (6–14) |
| 8396 | 9 | QSWMHQPHQ | β-casein | (141–149) |
| 7483 | 9 | RPKHPIKHQ | αS1-casein | (16–24) |
| 9376 | 9 | VEQKHIQKE | αS1-casein | (91–99) |
| 7566 | 9 | VRGPFPIIV | β-casein | (201–209) |
| 7486 | 9 | YPFPGPIPN | β-casein | (60–68) |
| 8156 | 9 | YPQRDMPIQ | β-casein | (180–188) |
| 8397 | 9 | YYAKPAAVR | κ-casein | (81–89) |
| 3336 | 9 | FAQTQSLVYP | β-casein | (52–61) |
| 8169/8286 | 2 | GPVRGPFPII | β-casein | (199–208) |
| 9231 | 12 | INNQFLPYPY | κ-casein | (72–81) |
| 7495 | 9 | NIPPLTQTPV | β-casein | (73–82) |
| 3216/3495 | 5 | YIPIQYVLSR | κ-casein | (46–55) |
| 7805 | 9 | ALNEINQFYQK | αS2-casein | (96–106) |
| 7797 | 11 | ARHPHPHLSFM | κ-casein | (117–127) |
| 9237 | 9 | LTQTPVVVPPF | β-casein | (77–87) |
| 3503 | 9 | TPVVVPPFLQP | β-casein | (80–90) |
| 3251 | 13 | YPFPGPIPNSL | β-casein | (60–70) |
| 9377 | 9 | EIVPNSAEERLH | αS1-casein | (125–136) |
| 7800 | 9 | FFVAPFPEVFGK | αS1-casein | (38–49) |
| 3337 | 9 | HPFAQTQSLVYP | β-casein | (50–61) |
| 3338 | 9 | IHPFAQTQSLVYP | β-casein | (49–61) |
| 9379 | 9 | QEPVLGPVRGPFP | β-casein | (194–206) |
| 3339 | 9 | KIHPFAQTQSLVYP | β-casein | (48–61) |
| 9240 | 9 | LVYPFPGPIPNSLPQN | β-casein | (58–73) |
| 7564 | 9 | LVYPFPGPIPNSLPQNIPP | β-casein | (58–76) |
References
- Karadag, A.S.; Ozlu, E.; Lavery, M.J. Cutaneous manifestations of diabetes mellitus and the metabolic syndrome. Clin. Dermatol. 2018, 36, 89–93. [Google Scholar] [CrossRef]
- Rochlani, Y.; Pothineni, N.V.; Kovelamudi, S.; Mehta, J.L. Metabolic syndrome: Pathophysiology, management, and modulation by natural compounds. Ther. Adv. Cardiovasc. Dis. 2017, 11, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.X.; Chaudhary, N.; Akinyemiju, T. Metabolic syndrome prevalence by race/ethnicity and sex in the united states, national health and nutrition examination survey, 1988–2012. Prev. Chronic Dis. 2017, 14, 1–16. [Google Scholar] [CrossRef]
- Huang, P.L. A comprehensive definition for metabolic syndrome. Dis. Model. Mech. 2009, 2, 231–237. [Google Scholar] [CrossRef]
- Iwaniak, A.; Darewicz, M.; Minkiewicz, P. Peptides Derived from Foods as Supportive Diet Components in the Prevention of Metabolic Syndrome. Compr. Rev. Food Sci. Food Saf. 2018, 17, 63–81. [Google Scholar] [CrossRef]
- González-Ortega, O.; López-Limón, A.R.; Morales-Domínguez, J.F.; Soria-Guerra, R.E. Production and purification of recombinant hypocholesterolemic peptides. Biotechnol. Lett. 2015, 37, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.P.; Vij, S.; Hati, S. Functional significance of bioactive peptides derived from soybean. Peptides 2014, 54, 171–179. [Google Scholar] [CrossRef]
- Yuan, L.; Shu, W.; Shen, Y.; Jin, W.; Gao, R. Inflammation and food-derived anti-inflammatory peptides. Biomed. Res. Sci. 2019, 1, 1001. [Google Scholar]
- Xu, Q.; Yan, X.; Zhang, Y.; Wu, J. Current understanding of transport and bioavailability of bioactive peptides derived from dairy proteins: A review. Int. J. Food Sci. Technol. 2019, 54, 1930–1941. [Google Scholar] [CrossRef]
- Dehghan, M.; Mente, A.; Rangarajan, S.; Sheridan, P.; Mohan, V.; Iqbal, R.; Gupta, R.; Lear, S.; Wentzel-Viljoen, E.; Avezum, A.; et al. Association of dairy intake with cardiovascular disease and mortality in 21 countries from five continents (PURE): A prospective cohort study. Lancet 2018, 392, 2288–2297. [Google Scholar] [CrossRef]
- Abedini, M.; Falahi, E.; Roosta, S. Dairy product consumption and the metabolic syndrome. Diabetes Metab. Syndr. Clin. Res. Rev. 2015, 9, 34–37. [Google Scholar] [CrossRef] [PubMed]
- López-Expósito, I.; Amigo, L.; Recio, I. A mini-review on health and nutritional aspects of cheese with a focus on bioactive peptides. Dairy Sci. Technol. 2012, 92, 419–438. [Google Scholar] [CrossRef]
- Darewicz, M.; Dziuba, B.; Minkiewicz, P.; Dziuba, J. The Preventive Potential of Milk and Colostrum Proteins and Protein Fragments. Food Rev. Int. 2011, 27, 357–388. [Google Scholar] [CrossRef]
- Skwarek, A.; Darewicz, M.; Borawska-Dziadkiewicz, J. Ripened cheese as a source of bioactive peptides. Biotechnol. Food Sci. 2018, 82, 49–60. [Google Scholar]
- McIntosh, C.H.S.; Widenmaier, S.; Kim, S. Chapter 15 Glucose-Dependent Insulinotropic Polypeptide (Gastric Inhibitory Polypeptide; GIP). Vitam. Horm. 2009, 80, 409–471. [Google Scholar] [CrossRef]
- Mkele, G. Dipeptidyl peptidase-4 inhibitors: Their role in the management of type 2 diabetes. S. Afr. Fam. Pr. 2013, 55, 508–510. [Google Scholar] [CrossRef]
- Uenishi, H.; Kabuki, T.; Seto, Y.; Serizawa, A.; Nakajima, H. Isolation and identification of casein-derived dipeptidyl-peptidase 4 (DPP-4)-inhibitory peptide LPQNIPPL from gouda-type cheese and its effect on plasma glucose in rats. Int. Dairy J. 2012, 22, 24–30. [Google Scholar] [CrossRef]
- Patil, P.; Mandal, S.; Tomar, S.K.; Anand, S. Food protein-derived bioactive peptides in management of type 2 diabetes. Eur. J. Nutr. 2015, 54, 863–880. [Google Scholar] [CrossRef]
- Sartorius, T.; Weidner, A.; Dharsono, T.; Boulier, A.; Wilhelm, M.; Schön, C. Postprandial Effects of a Proprietary Milk Protein Hydrolysate Containing Bioactive Peptides in Prediabetic Subjects. Nutrients 2019, 11, 1700. [Google Scholar] [CrossRef]
- Grom, L.C.; Rocha, R.S.; Balthazar, C.F.; Guimarães, J.T.; Coutinho, N.M.; Barros, C.P.; Pimentel, T.C.; Venâncio, E.L.; Collopy Junior, I.; Maciel, P.M.C.; et al. Postprandial glycemia in healthy subjects: Which probiotic dairy food is more adequate? J. Dairy Sci. 2020, 103, 1110–1119. [Google Scholar] [CrossRef]
- Hutchison, A.T.; Piscitelli, D.; Horowitz, M.; Jones, K.L.; Clifton, P.M.; Standfield, S.; Hausken, T.; Feinle-Bisset, C.; Luscombe-Marsh, N.D. Acute load-dependent effects of oral whey protein on gastric emptying, gut hormone release, glycemia, appetite, and energy intake in healthy men. Am. J. Clin. Nutr. 2015, 102, 1574–1584. [Google Scholar] [CrossRef] [PubMed]
- Drouin-Chartier, J.-P.; Li, Y.; Ardisson Korat, A.V.; Ding, M.; Lamarche, B.; Manson, J.E.; Rimm, E.B.; Willett, W.C.; Hu, F.B. Changes in dairy product consumption and risk of type 2 diabetes: Results from 3 large prospective cohorts of US men and women. Am. J. Clin. Nutr. 2019, 110, 1201–1212. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Ning, N.; Wang, C.; Wang, Y.; Li, Q.; Meng, Z.; Liu, Y.; Li, Q. Dairy Products Consumption and Risk of Type 2 Diabetes: Systematic Review and Dose-Response Meta-Analysis. PLoS ONE 2013, 8, e73965. [Google Scholar] [CrossRef]
- Aune, D.; Norat, T.; Romundstad, P.; Vatten, L.J. Dairy products and the risk of type 2 diabetes: A systematic review and dose-response meta-analysis of cohort studies. Am. J. Clin. Nutr. 2013, 98, 1066–1083. [Google Scholar] [CrossRef] [PubMed]
- Hruby, A.; Ma, J.; Rogers, G.; Meigs, J.B.; Jacques, P.F. Associations of Dairy Intake with Incident Prediabetes or Diabetes in Middle-Aged Adults Vary by Both Dairy Type and Glycemic Status. J. Nutr. 2017, 147, 1764–1775. [Google Scholar] [CrossRef] [PubMed]
- Iwaniak, A.; Minkiewicz, P.; Darewicz, M. Food-Originating ACE Inhibitors, Including Antihypertensive Peptides, as Preventive Food Components in Blood Pressure Reduction. Compr. Rev. Food Sci. Food Saf. 2014, 13, 114–134. [Google Scholar] [CrossRef]
- Hrynkiewicz, M.; Iwaniak, A.; Bucholska, J.; Minkiewicz, P.; Darewicz, M. Structure-activity prediction of ACE inhibitory/bitter dipeptides - a chemometric approach based on stepwise regression. Molecules 2019, 24, 950. [Google Scholar] [CrossRef] [PubMed]
- Beltrán-Barrientos, L.M.; Hernández-Mendoza, A.; Torres-Llanez, M.J.; González-Córdova, A.F.; Vallejo-Córdoba, B. Inivited review: Fermented milk as antihypertensive functional food. J. Dairy Sci. 2016, 99, 499–4110. [Google Scholar] [CrossRef]
- Bütikofer, U.; Meyer, J.; Sieber, R.; Wechsler, D. Quantification of the angiotensin-converting enzyme-inhibiting tripeptides Val-Pro-Pro and Ile-Pro-Pro in hard, semi-hard and soft cheeses. Int. Dairy J. 2007, 17, 968–975. [Google Scholar] [CrossRef]
- Pihlanto, A. Lactic fermentation and bioactive peptides. In Lactic Acid Bacteria. R & D for Food, Health and Livestock Purposes; Kongo, J.M., Ed.; IntechOpen: London, UK, 2013; pp. 309–332. [Google Scholar] [CrossRef]
- Sieber, R.; Bütikofer, U.; Egger, C.; Portmann, R.; Walther, B.; Wechsler, D. ACE inhibitory activity and ACE-inhibiting peptides. Dairy Sci. Technol. 2010, 90, 47–73. [Google Scholar] [CrossRef]
- Haque, E.; Chand, R. Antihypertensive and Antimicrobial Bioactive Peptides from Milk Proteins. Eur. Food Res. Technol. 2008, 227, 7–15. [Google Scholar] [CrossRef]
- De Noni, I.; Cattaneo, S. Occurrence of β-casomorphins 5 and 7 in commercial dairy products and in their digests following in vitro simulated gastro-intestinal digestion. Food Chem. 2010, 119, 560–566. [Google Scholar] [CrossRef]
- Gómez-Ruiz, J.Á.; Ramos, M.; Recio, I. Angiotensin-converting enzyme-inhibitory peptides in Manchego cheeses manufactured with different starter cultures. Int. Dairy J. 2002, 12, 697–706. [Google Scholar] [CrossRef]
- Pripp, A.H.; Isaksson, T.; Stepaniak, L.; Sørhaug, T. Quantitative structure-activity relationship modelling of ACE-inhibitory peptides derived from milk proteins. Eur. Food Res. Technol. 2004, 219, 579–583. [Google Scholar] [CrossRef]
- Gómez-Ruiz, J.Á.; Taborda, G.; Amigo, L.; Recio, I.; Ramos, M. Identification of ACE-inhibitory peptides in different Spanish cheeses by tandem mass spectrometry. Eur. Food Res. Technol. 2006, 223, 595–601. [Google Scholar] [CrossRef]
- Gómez-Ruiz, J.A.; Recio, I.; Belloque, J. ACE-Inhibitory Activity and Structural Properties of Peptide Asp-Lys-Ile-His-Pro [β-CN f(47−51)]. Study of the Peptide Forms Synthesized by Different Methods. J. Agric. Food Chem. 2004, 52, 6315–6319. [Google Scholar] [CrossRef]
- Tonouchi, H.; Suzuki, M.; Uchida, M.; Oda, M. Antihypertensive effect of an angiotensin converting enzyme inhibitory peptide from enzyme modified cheese. J. Dairy Sci. 2008, 75, 284–290. [Google Scholar] [CrossRef]
- Nilsen, R.; Pripp, A.H.; Høstmark, A.T.; Haug, A.; Skeie, S. Short communication: Is consumption of a cheese rich in angiotensin-converting enzyme-inhibiting peptides, such as the Norwegian cheese Gamalost, associated with reduced blood pressure? J. Dairy Sci. 2014, 97, 2662–2668. [Google Scholar] [CrossRef]
- Crippa, G.; Zabzuni, D.; Bravi, E.; Piva, G.; De Noni, I.; Bighi, E.; Rossi, F. Randomized, double blind placebo-controlled pilot study of the antihypertensive effects of Grana Padano D.O.P. cheese consumption in mild—Moderate hypertensive subjects. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7573–7581. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-C.; Wang, Y.; Tong, X.; Szeto, I.M.Y.; Smit, G.; Li, Z.-N.; Qin, L.-Q. Cheese consumption and risk of cardiovascular disease: A meta-analysis of prospective studies. Eur. J. Nutr. 2017, 56, 2565–2575. [Google Scholar] [CrossRef]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef]
- Gupta, A.; Mann, B.; Kumar, R.; Sangwan, R.B. Antioxidant activity of Cheddar cheeses at different stages of ripening. Int. J. Dairy Technol. 2009, 62, 339–347. [Google Scholar] [CrossRef]
- Gupta, A.K.; Mann, B.; Kumar, R.; Sangwan, R. Identification of antioxidant peptides in cheddar cheese made with adjunct culture Lactobacillus casei ssp. casei 300. Milchwissenschaft 2010, 65, 396–399. [Google Scholar]
- Pritchard, S.R.; Phillips, M.; Kailasapathy, K. Identification of bioactive peptides in commercial Australian organic cheddar cheeses. Aust. J. Dairy Technol. 2010, 65, 170–173. [Google Scholar] [CrossRef]
- Hernández-Galán, L.; Cardador-Martínez, A.; López-del-Castillo, M.; Picque, D.; Spinnler, H.E.; Martín del Campo, S.T. Antioxidant and angiotensin-converting enzyme inhibitory activity in fresh goat cheese prepared without starter culture: A preliminary study. CyTA J. Food 2016, 15, 1–9. [Google Scholar] [CrossRef]
- Timón, M.L.; Andrés, A.; Otte, J.; Petrón, M.J. Antioxidant peptides (<3 kDa) identified on hard cow milk cheese with rennet from different origin. Food Res. Int. 2019, 120, 643–649. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Chemometric analysis of the amino acid requirements of antioxidant food protein hydrolysates. Int. J. Mol. Sci. 2011, 12, 3148–3161. [Google Scholar] [CrossRef]
- Timón, M.L.; Parra, V.; Otte, J.; Broncano, J.M.; Petrón, M.J. Identification of radical scavenging peptides (<3 kDa) from Burgos-type cheese. LWT-Food Sci. Technol. 2014, 57, 359–365. [Google Scholar] [CrossRef]
- Cox, R.A.; García-Palmieri, M.R. Cholesterol, Triglycerides, and Associated Lipoproteins. In Clinical Methods: The History, Physical, and Laboratory Examinations, 3rd ed.; Walker, H.K., Hall, W.D., Hurst, J.W., Eds.; Butterworths: Boston, MA, USA, 1990; Chapter 31; pp. 153–160. [Google Scholar]
- Contarini, G.; Povolo, M. Phospholipids in milk fat: Composition, biological and technological significance, and analytical strategies. Int. J. Mol. Sci. 2013, 14, 2808–2831. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Rouvinen-Watt, K. The Role of Food Peptides in Lipid Metabolism during Dyslipidemia and Associated Health Conditions. Int. J. Mol. Sci. 2015, 16, 9303–9313. [Google Scholar] [CrossRef]
- Hsieh, C.-C.; Hernández-Ledesma, B.; Fernández-Tomé, S.; Weinborn, V.; Barile, D.; de Moura Bell, J.M.L.N. Milk Proteins, Peptides, and Oligosaccharides: Effects against the 21st Century Disorders. Biomed Res. Int. 2015, 2015, 146840. [Google Scholar] [CrossRef]
- Nagaoka, S. Structure-function properties of hypolipidemic peptides. J. Food Biochem. 2019, 43, e12539. [Google Scholar] [CrossRef]
- Wakasa, Y.; Tamakoshi, C.; Ohno, T.; Hirose, S.; Goto, T.; Nagaoka, S.; Takaiwa, F. The Hypocholesterolemic Activity of Transgenic Rice Seed Accumulating Lactostatin, a Bioactive Peptide Derived from Bovine Milk β-Lactoglobulin. J. Agric. Food Chem. 2011, 59, 3845–3850. [Google Scholar] [CrossRef] [PubMed]
- Cabanos, C.; Ekyo, A.; Amari, Y.; Kato, N.; Kuroda, M.; Nagaoka, S.; Takaiwa, F.; Utsumi, S.; Maruyama, N. High-level production of lactostatin, a hypocholesterolemic peptide, in transgenic rice using soybean A1aB1b as carrier. Transgenic Res. 2013, 22, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, R.; Ohinata, K.; Yoshikawa, M. β-Lactotensin and neurotensin rapidly reduce serum cholesterol via NT2 receptor. Peptides 2003, 24, 1955–1961. [Google Scholar] [CrossRef]
- Keller, U. Dietary Proteins in Obesity and in Diabetes. Int. J. Vitam. Nutr. Res. 2011, 81, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Campos-Nonato, I.; Hernandez, L.; Barquera, S. Effect of a High-Protein Diet versus Standard-Protein Diet on Weight Loss and Biomarkers of Metabolic Syndrome: A Randomized Clinical Trial. Obes Facts. 2017, 10, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, K.; Cheung, B.W.; Schröder, H. The possible role of food-derived peptides in reducing the risk of cardiovascular disease. J. Nutr. Biochem. 2008, 19, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Bhat, Z.F.; Kumar, S.; Bhat, H.F. Bioactive peptides of animal origin: A review. J. Food Sci. Technol. 2015, 52, 5377–5392. [Google Scholar] [CrossRef] [PubMed]
- Kondrashina, A.; Brodkorb, A.; Giblin, L. Dairy-derived peptides in satiety. J. Funct. Foods 2020, 66, 103801. [Google Scholar] [CrossRef]
- Monteiro, R.; Azevedo, I. Chronic inflammation in obesity and the metabolic syndrome. Mediat. Inflamm. 2010, 2010, 289645. [Google Scholar] [CrossRef]
- Zielińska, E.; Baraniak, B.; Karaś, M. Antioxidant and Anti-Inflammatory Activities of Hydrolysates and Peptide Fractions Obtained by Enzymatic Hydrolysis of Selected Heat-Treated Edible Insects. Nutrients 2017, 9, 970. [Google Scholar] [CrossRef]
- Marcone, S.; Belton, O.; FitzGerald, D.J. Milk-derived bioactive peptides and their health-promoting effects: A potential role in atherosclerosis. Br. J. Clin. Pharmacol. 2017, 83, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Rafiq, S.; Huma, N.; Rakariyatham, K.; Hussain, I.; Gulzar, N.; Hayat, I. Anti-inflammatory and anticancer activities of water-soluble peptide extracts of buffalo and cow milk Cheddar cheeses. Int. J. Dairy Technol. 2017, 71, 432–438. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef] [PubMed]
- Iwaniak, A.; Darewicz, M.; Mogut, D.; Minkiewicz, P. Elucidation of the role of in silico methodologies in approaches to studying bioactive peptides derived from foods. J. Funct. Foods 2019, 61, 103486. [Google Scholar] [CrossRef]





| Cheese | VPP | IPP | IC50 |
|---|---|---|---|
| (mg × kg−1) | (mg × kg−1) | (mg cheese × mL−1) | |
| Hard cheeses | |||
| Emmental: | |||
| cave-aged, aged 12 months | 100.1 | 23.5 | 7.1 ± 0.2 |
| reserve, aged 8 months | 68.6 | 13.2 | 9.9 ± 0.2 |
| organic, aged 4 months | 153.7 | 35.8 | 7.5 ± 1.8 |
| classic, aged 4 months | 42.6 | 4.6 | 10.5 ± 0.6 |
| Gouda old | 97.7 | 90.1 | 2.0 ± 0.3 |
| Gruyere AOC: | |||
| vieux, aged 12 months | 13.7 | 7.9 | 29.4 ± 16.8 |
| salé, aged 10 months | 68.7 | 20.4 | 14.2 ± 0.9 |
| mi-salé, aged 8 months | 22.7 | 14.9 | 18.7 ± 6.1 |
| doux, aged 5 months | 53.0 | 9.3 | 14.9 ± 0.7 |
| Hobelkäse from the Bernese Oberland | *224.1 | 95.4 | 2.6 ± 0.2 |
| Parmino | 2.0 | 0.8 | 28.6 ± 4.2 |
| Sbrinz | 28.0 | 9.4 | 16.4 ± 5.5 |
| Semi-hard cheeses | |||
| Appenzeller: | |||
| aged 7–10 months (1/4 fat) | 94.4 | 64.9 | 4.2 ± 1.6 |
| extra, aged 6–7 months | 29.3 | 14.5 | 16.8 ± 1.1 |
| surchoix, aged 4 months | 26.7 | 17.8 | 8.0 ± 1.0 |
| aged 3 months (1/4 fat) | 56.0 | 10.5 | 7.1 ± 1.9 |
| classic, aged 3 months | 22.0 | 8.5 | 17.6 ± 7.6 |
| Edam | 1.0 | 0.1 | 13.3 ± 0.4 |
| Küsnachter | 25.7 | 18.0 | 19.6 ± 2.4 |
| Mountain cheese Graubünden: | |||
| ¼ fat | 32.0 | 22.7 | 4.8 ± 0.0 |
| ½ fat | 10.4 | 7.1 | 8.9 ± 4.1 |
| full fat | 25.1 | 4.3 | 9.2 ± 1.4 |
| Raclette: | |||
| from raw milk | 12.6 | 2.6 | 8.8 ± 0.0 |
| from pasteurized milk | 1.0 | 0.1 | 9.0 ± 0.3 |
| St. Paulin | 5.2 | 1.0 | 23.5 ± 6.3 |
| Tête de Moine | 98.0 | 15.5 | 7.1 ± 1.9 |
| Tilsit: | |||
| from raw milk | 120.2 | 30.0 | 4.4 ± 0.2 |
| from pasteurized milk | 1.2 | 0.1 | 18.0 ± 6.3 |
| Vacherin fribourgeois | 62.1 | 46.7 | 5.0 ± 0.3 |
| Wangener Geissmutschli (goat cheese) | 33.2 | 0.0 | 7.9 ± 0.5 |
| Winzerkäse | 97.7 | 27.1 | 9.9 ± 1.8 |
| Soft cheeses | |||
| Brie | 0.4 | 0.3 | 25.9 ± 3.0 |
| Camembert | 0.2 | 0.2 | Very weak |
| Mozarella: | |||
| from buffalo milk | 0.1 | 0.0 | - |
| from cow milk | 0.0 | 0.0 | - |
| Tomme vaudoise | 0.0 | 0.0 | 14.9 ± 3.7 |
| Vacherin Mont d’Or | 0.2 | 0.1 | 21.4 ± 9.6 |
| Cheese | Concentration of YPFPGPI (mg × kg−1) | |
|---|---|---|
| Before Simulated Digestion | After Simulated Digestion | |
| Brie | 0.15 | 2.07 ± 0.09 |
| Caprino | 0 | 1.51 ± 0.09 |
| Cheddar | 0.11 | 15.22 ± 0.13 |
| Fontina | 0.04 | 10.43 ± 0.12 |
| Gorgonzola | 0.01 | 5.41 ± 0.08 |
| Gouda | 0.10 | 21.77 ± 0.12 |
| Grana Padano (aged 25 months) | 0 | 11.06 ± 0.08 |
| Grana Padano (aged 17 months) | 0 | 12.55 ± 0.09 |
| Grana Padano (aged 10 months) | 0 | 8.79 ± 0.04 |
| Taleggio | 0 | 7.15 ± 0.07 |
| Cheese | Peptide | IC50 (µM) | Precursor | Reference |
| Manchego, Roncal, Idiazábal, Cabrales, goat | DKIHP | 113.1 | β-casein | [36] |
| DKIHPF | 2419.4 | |||
| Goat | VRGP | 120.9 | ||
| Manchego | PFP | 144.4 | various fragments | [36] |
| FP | 1215.7 | |||
| PP | 2284.7 | |||
| KKYNVPQL | 77.2 | αs1-casein | [30] | |
| VRYL | 24.1 | |||
| Manchego, Roncal, Idiazabal, Goat | QP | 598.1 | various fragments | [36] |
| Roncal | PKHP | 709.1 | αS1- casein |
| Dual and Triple Activity of Casein-Derived Peptides | Number of Peptides Encrypted in the All Casein Sequences |
|---|---|
| ACE and DPP4 inhibitor | 66 |
| ACE inhibitor and antioxidative | 17 |
| DPP4 and α-glucosidase inhibitor | 1 |
| DPP4 inhibitor and opioid | 1 |
| ACE inhibitor and opioid | 5 |
| DPP4 inhibitor and antioxidative | 5 |
| ACE, DPP4 inhibitor, and antioxidative | 8 |
| ACE, DPP4, and α-glucosidase inhibitor | 1 |
| Total | 104 |
| Peptides Showing one Bioactivity | |
| ACE inhibitor | 171 |
| α-glucosidase inhibitor | 0 |
| antioxidative | 52 |
| DPP4 inhibitor | 127 |
| opioid agonist | 3 |
| opioid antagonist | 2 |
| opioid | 4 |
| Total | 359 |
| Total Number of Bioactive Sequences | 463 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iwaniak, A.; Mogut, D. Metabolic Syndrome-Preventive Peptides Derived from Milk Proteins and Their Presence in Cheeses: A Review. Appl. Sci. 2020, 10, 2772. https://doi.org/10.3390/app10082772
Iwaniak A, Mogut D. Metabolic Syndrome-Preventive Peptides Derived from Milk Proteins and Their Presence in Cheeses: A Review. Applied Sciences. 2020; 10(8):2772. https://doi.org/10.3390/app10082772
Chicago/Turabian StyleIwaniak, Anna, and Damir Mogut. 2020. "Metabolic Syndrome-Preventive Peptides Derived from Milk Proteins and Their Presence in Cheeses: A Review" Applied Sciences 10, no. 8: 2772. https://doi.org/10.3390/app10082772
APA StyleIwaniak, A., & Mogut, D. (2020). Metabolic Syndrome-Preventive Peptides Derived from Milk Proteins and Their Presence in Cheeses: A Review. Applied Sciences, 10(8), 2772. https://doi.org/10.3390/app10082772