Telomere Length as a Biomarker of Biological Aging in Shift Workers




Abstract
1. Introduction
2. Materials and Methods
2.1. Literature Search
2.2. Inclusion and Exclusion Criteria
2.3. Quality Assessment and Data Extraction
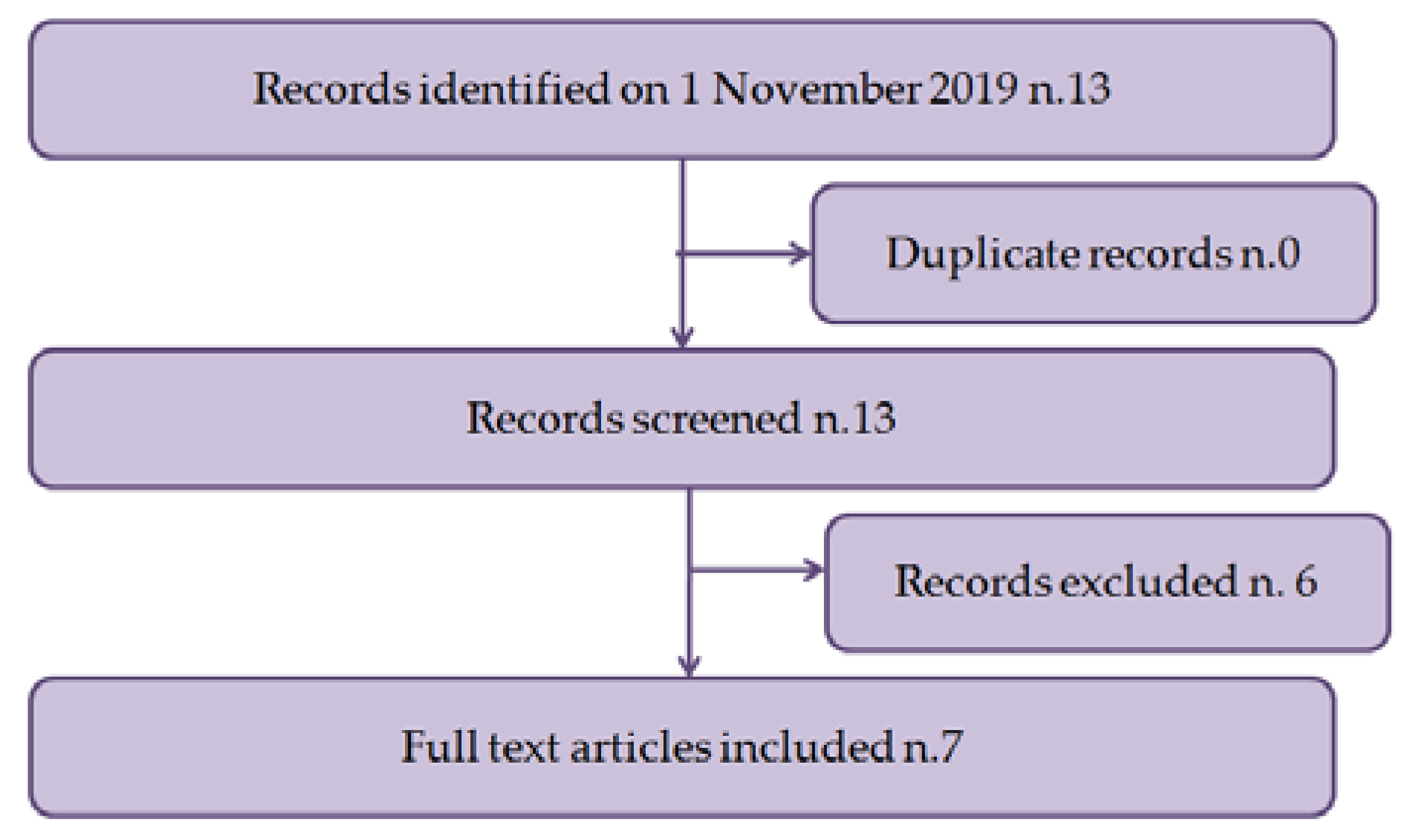
3. Results
3.1. Characteristics of Eligible Studies
3.2. Results of Eligible Studies
4. Discussion
5. Conclusions
Funding
Conflicts of Interest
References
- Wang, X.-S.; Armstrong, M.E.G.; Cairns, B.; Key, T.J.; Travis, R.C. Shift work and chronic disease: The epidemiological evidence. Occup. Med. 2011, 61, 78–89. [Google Scholar] [CrossRef]
- Åkerstedt, T. Shift work and disturbed sleep/wakefulness. Occup. Med. 2003, 53, 89–94. [Google Scholar] [CrossRef]
- Eurofound. Fifth European Working Conditions Survey; Eurofound: Dublin, UK, 2012.
- Gärtner, J.; Rosa, R.R.; Roach, G.; Kubo, T.; Takahashi, M. Working Time Society consensus statements: Regulatory approaches to reduce risks associated with shift work—A global comparison. Ind. Health 2019, 57, 245–263. [Google Scholar] [CrossRef]
- IARC Monographs Vol 124 group. Carcinogenicity of night shift work. Lancet Oncol. 2019, 20, 1058–1059. [Google Scholar] [CrossRef]
- Åkerstedt, T.; Nordin, M.; Alfredsson, L.; Westerholm, P.; Kecklund, G. Sleep and sleepiness: Impact of entering or leaving shiftwork—A prospective study. Chrono Int. 2010, 27, 987–996. [Google Scholar] [CrossRef]
- Härmä, M.; Ilmarinen, J.E. Towards the 24-hour society--new approaches for aging shift workers? Scand. J. Work. Environ. Health 1999, 25, 610–615. [Google Scholar] [CrossRef] [PubMed]
- Özdemir, P.G.; Selvi, Y.; Özkol, H.; Aydin, A.; Tülüce, Y.; Boysan, M.; Beşiroǧlu, L. The influence of shift work on cognitive functions and oxidative stress. Psychiatr. Res. 2013, 210, 1219–1225. [Google Scholar] [CrossRef] [PubMed]
- Machi, M.S.; Staum, M.; Callaway, C.W.; Moore, C.; Jeong, K.; Suyama, J.; Patterson, P.; Hostler, D. The Relationship Between Shift Work, Sleep, and Cognition in Career Emergency Physicians. Acad. Emerg. Med. 2012, 19, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, T.; Yaffe, K.; Ancoli-Israel, S.; Schneider, J.L.; Cauley, J.A.; Hillier, T.A.; Fink, H.A.; Stone, K.L. Poor sleep is associated with impaired cognitive function in older women: The study of osteoporotic fractures. J. Gerontol. Ser. A Biol. Sci. Med Sci. 2006, 61, 405–410. [Google Scholar] [CrossRef]
- Devore, E.E.; Grodstein, F.; Schernhammer, E. Shift work and cognition in the Nurses’ Health Study. Am. J. Epidemiol. 2013, 178, 1296–1300. [Google Scholar] [CrossRef]
- Bokenberger, K.; Ström, P.; Dahl Aslan, A.K.; Åkerstedt, T.; Pedersen, N.L. Shift work and cognitive aging: A longitudinal study. Scand. J. Work Environ. Health 2017, 43, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Bass, J.; Takahashi, J.S. Circadian Integration of Metabolism and Energetics. Science 2010, 330, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Copertaro, A.; Bracci, M. Working against the biological clock: A review for the Occupational Physician. Ind. Health 2019, 57, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Valladares, M.; Obregón, A.M.; Chaput, J.-P. Association between genetic variants of the clock gene and obesity and sleep duration. J. Physiol. Biochem. 2015, 71, 855–860. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.J.; Welsh, D. Cellular Circadian Clocks in Mood Disorders. J. Biol. Rhythm. 2012, 27, 339–352. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, F.C.; Rao, A.; Maguire, A. Circadian molecular clocks and cancer. Cancer Lett. 2014, 342, 9–18. [Google Scholar] [CrossRef]
- Antoch, M.P.; Gorbacheva, V.Y.; Vykhovanets, O.; Toshkov, I.A.; Kondratov, R.V.; Kondratova, A.A.; Lee, C.; Nikitin, A.Y. Disruption of the circadian clock due to the Clock mutation has discrete effects on aging and carcinogenesis. Cell Cycle 2008, 7, 1197–1204. [Google Scholar] [CrossRef]
- Galbete, C.; Contreras, R.; Martínez, A.; Martínez-González, M.Á.; Guillen-Grima, F.; Marti, A. Physical Activity and Sex Modulate Obesity Risk Linked to 3111T/C Gene Variant of theCLOCKGene in an Elderly Population: The SUN Project. Chronobiol. Int. 2012, 29, 1397–1404. [Google Scholar] [CrossRef]
- Liu, F.; Chang, H.-C. Physiological links of circadian clock and biological clock of aging. Protein Cell 2017, 8, 477–488. [Google Scholar] [CrossRef]
- Froy, O. Circadian Rhythms, Aging, and Life Span in Mammals. Physiology 2011, 26, 225–235. [Google Scholar] [CrossRef]
- Gibson, E.M.; Williams, W.P.; Kriegsfeld, L.J. Aging in the circadian system: Considerations for health, disease prevention and longevity. Exp. Gerontol. 2008, 44, 51–56. [Google Scholar] [CrossRef]
- Levine, M.E. Modeling the Rate of Senescence: Can Estimated Biological Age Predict Mortality More Accurately Than Chronological Age? J. Gerontol. Ser. A Biol. Sci. Med Sci. 2013, 68, 667–674. [Google Scholar] [CrossRef]
- Belsky, D.W.; Caspi, A.; Houts, R.; Cohen, H.J.; Corcoran, D.L.; Danese, A.; Harrington, H.; Israel, S.; Levine, M.E.; Schaefer, J.; et al. Quantification of biological aging in young adults. Proc. Natl. Acad. Sci. USA 2015, 112, E4104–E4110. [Google Scholar] [CrossRef]
- White, A.J.; Kresovich, J.K.; Xu, Z.; Sandler, D.P.; Taylor, J.A. Shift work, DNA methylation and epigenetic age. Int. J. Epidemiol. 2019, 48, 1536–1544. [Google Scholar] [CrossRef]
- Chen, B.H.; Marioni, R.E.; Colicino, E.; Peters, M.J.; Ward-Caviness, C.K.; Tsai, P.-C.; Roetker, N.S.; Just, A.; Demerath, E.W.; Guan, W.; et al. DNA methylation-based measures of biological age: Meta-analysis predicting time to death. Aging 2016, 8, 1844–1865. [Google Scholar] [CrossRef]
- Belsky, D.W.; Moffitt, T.E.; Cohen, A.A.; Corcoran, D.L.; Levine, M.E.; Prinz, J.A.; Schaefer, J.; Sugden, K.; Williams, B.; Poulton, R.; et al. Eleven Telomere, Epigenetic Clock, and Biomarker-Composite Quantifications of Biological Aging: Do They Measure the Same Thing? Am. J. Epidemiol. 2018, 187, 1220–1230. [Google Scholar] [CrossRef]
- Ferrucci, L.; Levine, M.E.; Kuo, P.-L.; Simonsick, E.M. Time and the Metrics of Aging. Circ. Res. 2018, 123, 740–744. [Google Scholar] [CrossRef]
- Bracci, M.; Copertaro, A.; Ciarapica, V.; Barbaresi, M.; Esposito, S.; Albanesi, A.; Valentino, M.; Ledda, C.; Rapisarda, V.; Santarelli, L. NOCTURNIN Gene Diurnal Variation in Healthy Volunteers and Expression Levels in Shift Workers. BioMed Res. Int. 2019. [Google Scholar] [CrossRef]
- Bracci, M.; Manzella, N.; Copertaro, A.; Staffolani, S.; Strafella, E.; Barbaresi, M.; Copertaro, B.; Rapisarda, V.; Valentino, M.; Santarelli, L. Rotating-shift nurses after a day off: Peripheral clock gene expression, urinary melatonin, and serum 17-β-estradiol levels. Scand. J. Work Environ. Health 2014, 40, 295–304. [Google Scholar] [CrossRef]
- Manzella, N.; Bracci, M.; Strafella, E.; Staffolani, S.; Ciarapica, V.; Copertaro, A.; Rapisarda, V.; Ledda, C.; Amati, M.; Valentino, M.; et al. Circadian Modulation of 8-Oxoguanine DNA Damage Repair. Sci. Rep. 2015, 5, 13752. [Google Scholar] [CrossRef]
- Barbadoro, P.; Santarelli, L.; Croce, N.; Bracci, M.; Vincitorio, D.; Prospero, E.; Minelli, A. Rotating Shift-Work as an Independent Risk Factor for Overweight Italian Workers: A Cross-Sectional Study. PLoS ONE 2013, 8, e63289. [Google Scholar] [CrossRef]
- Bracci, M.; Ciarapica, V.; Zabaleta, M.E.; Tartaglione, M.F.; Pirozzi, S.; Giuliani, L.; Piva, F.; Valentino, M.; Ledda, C.; Rapisarda, V.; et al. BRCA1 and BRCA2 Gene Expression: Diurnal Variability and Influence of Shift Work. Cancers 2019, 11, 1146. [Google Scholar] [CrossRef]
- Bracci, M.; Ciarapica, V.; Copertaro, A.; Barbaresi, M.; Manzella, N.; Tomasetti, M.; Gaetani, S.; Monaco, F.; Amati, M.; Valentino, M.; et al. Peripheral Skin Temperature and Circadian Biological Clock in Shift Nurses after a Day off. Int. J. Mol. Sci. 2016, 17, 623. [Google Scholar] [CrossRef]
- Bracci, M.; Copertaro, A.; Manzella, N.; Staffolani, S.; Strafella, E.; Nocchi, L.; Barbaresi, M.; Copertaro, B.; Rapisarda, V.; Valentino, M.; et al. Influence of night-shift and napping at work on urinary melatonin, 17- ß -estradiol and clock gene expression in pre-menopausal nurses. J. Biol. Regul. Homeost. Agents 2013, 27, 267–274. [Google Scholar]
- Copertaro, A.; Bracci, M.; Barbaresi, M.; Santarelli, L. Assessment of cardiovascular risk in shift healthcare workers. Eur. J. Cardiovasc. Prev. Rehabil. 2008, 15, 224–229. [Google Scholar] [CrossRef]
- Copertaro, A.; Bracci, M.; Gesuita, R.; Carle, F.; Amati, M.; Baldassari, M.; Mocchegiani, E.; Santarelli, L. Influence of shift-work on selected immune variables in nurses. Ind. Health 2011, 49, 597–604. [Google Scholar] [CrossRef]
- Ledda, C.; Cinà, D.; Matera, S.; Mucci, N.; Bracci, M.; Rapisarda, V. High HOMA-IR Index in Healthcare Shift Workers. Medicina 2019, 55, 186. [Google Scholar] [CrossRef]
- Starkweather, A.R.; Alhaeeri, A.A.; Montpetit, A.; Brumelle, J.; Filler, K.; Montpetit, M.; Mohanraj, L.; Lyon, D.E.; Jackson-Cook, C.K. An integrative review of factors associated with telomere length and implications for biobehavioral research. Nurs. Res. 2013, 63, 36–50. [Google Scholar] [CrossRef]
- Armstrong, L.; Lako, M.; Van Herpe, I.; Evans, J.; Saretzki, G.; Hole, N. A role for nucleoprotein Zap3 in the reduction of telomerase activity during embryonic stem cell differentiation. Mech. Dev. 2004, 121, 1509–1522. [Google Scholar] [CrossRef]
- McKerlie, M.; Zhu, X.-D. Cyclin B-dependent kinase 1 regulates human TRF1 to modulate the resolution of sister telomeres. Nat. Commun. 2011, 2, 371. [Google Scholar] [CrossRef]
- Parks, C.G.; DeRoo, L.A.; Miller, D.B.; McCanlies, E.C.; Cawthon, R.M.; Sandler, D.P. Employment and work schedule are related to telomere length in women. Occup. Environ. Med. 2011, 68, 582–589. [Google Scholar] [CrossRef]
- Von Zglinicki, T. Oxidative stress shortens telomeres. Trends Biochem. Sci. 2002, 27, 339–344. [Google Scholar] [CrossRef]
- Gilchrest, B.A.; Eller, M.S. The Tale of the Telomere: Implications for Prevention and Treatment of Skin Cancers. J. Investig. Dermatol. Symp. Proc. 2005, 10, 124–130. [Google Scholar] [CrossRef]
- Sharpless, N.E.; DePinho, R.A. Telomeres, stem cells, senescence, and cancer. J. Clin. Investig. 2004, 113, 160–168. [Google Scholar] [CrossRef]
- Houben, J.M.; Moonen, H.J.; Van Schooten, F.J.; Hageman, G.J. Telomere length assessment: Biomarker of chronic oxidative stress? Free. Radic. Biol. Med. 2008, 44, 235–246. [Google Scholar] [CrossRef]
- Kawanishi, S.; Oikawa, S. Mechanism of Telomere Shortening by Oxidative Stress. Ann. N. Y. Acad. Sci. 2004, 1019, 278–284. [Google Scholar] [CrossRef]
- Sitte, N.; Saretzki, G.; Von Zglinicki, T. Accelerated telomere shortening in fibroblasts after extended periods of confluency. Free. Radic. Biol. Med. 1998, 24, 885–893. [Google Scholar] [CrossRef]
- Oikawa, S.; Kawanishi, S. Site-specific DNA damage at GGG sequence by oxidative stress may accelerate telomere shortening. FEBS Lett. 1999, 453, 365–368. [Google Scholar] [CrossRef]
- Oikawa, S.; Tada-Oikawa, S.; Kawanishi, S. Site-specific DNA damage at the GGG sequence by UVA involves acceleration of telomere shortening. Biochemistry 2001, 40, 4763–4768. [Google Scholar] [CrossRef]
- Petersen, S.; Saretzki, G.; Von Zglinicki, T. Preferential Accumulation of Single-Stranded Regions in Telomeres of Human Fibroblasts. Exp. Cell Res. 1998, 239, 152–160. [Google Scholar] [CrossRef]
- Kruk, P.A.; Rampino, N.J.; Bohr, V.A. DNA damage and repair in telomeres: Relation to aging. Proc. Natl. Acad. Sci. USA 1995, 92, 258–262. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Altman, D.; Antes, G.; Atkins, D.; Barbour, V.; Barrowman, N.; Berlin, J.A.; et al. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6. [Google Scholar] [CrossRef]
- Liang, G.; Schernhammer, E.; Qi, L.; Gao, X.; De Vivo, I.; Han, J. Associations between Rotating Night Shifts, Sleep Duration, and Telomere Length in Women. PLoS ONE 2011, 6, e23462. [Google Scholar] [CrossRef]
- Chen, W.-D.; Wen, M.-S.; Shie, S.-S.; Lo, Y.-L.; Wo, H.-T.; Wang, C.-C.; Hsieh, I.-C.; Lee, T.-H.; Wang, C.-Y. The circadian rhythm controls telomeres and telomerase activity. Biochem. Biophys. Res. Commun. 2014, 451, 408–414. [Google Scholar] [CrossRef]
- Pavanello, S.; Stendardo, M.; Mastrangelo, G.; Bonci, M.; Bottazzi, B.; Campisi, M.; Nardini, M.; Leone, R.; Mantovani, A.; Boschetto, P. Inflammatory Long Pentraxin 3 is Associated with Leukocyte Telomere Length in Night-Shift Workers. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Erdem, J.S.; Notø, H.Ø.; Skare, Ø.; Lie, J.S.; Petersen-Øverleir, M.; Reszka, E.; Pepłońska, B.; Zienolddiny, S. Mechanisms of breast cancer risk in shift workers: Association of telomere shortening with the duration and intensity of night work. Cancer Med. 2017, 6, 1988–1997. [Google Scholar] [CrossRef]
- Chmelar, C.; Jörres, R.A.; Kronseder, A.; Müller, A.; Nowak, D.; Weigl, M. Associations Between Age, Psychosocial Work Conditions, Occupational Well-Being, and Telomere Length in Geriatric Care Professionals. J. Occup. Environ. Med. 2017, 59, 949–955. [Google Scholar] [CrossRef]
- Pavanello, S.; Stendardo, M.; Mastrangelo, G.; Casillo, V.; Nardini, M.; Mutti, A.; Campisi, M.; Andreoli, R.; Boschetto, P. Higher Number of Night Shifts Associates with Good Perception of Work Capacity and Optimal Lung Function but Correlates with Increased Oxidative Damage and Telomere Attrition. BioMed Res. Int. 2019. [Google Scholar] [CrossRef]
- Carugno, M.; Maggioni, C.; Crespi, E.; Bonzini, M.; Cuocina, S.; Dioni, L.; Tarantini, L.; Consonni, D.; Ferrari, L.; Pesatori, A.C. Night Shift Work, DNA Methylation and Telomere Length: An Investigation on Hospital Female Nurses. Int. J. Environ. Res. Public Health 2019, 16, 2292. [Google Scholar] [CrossRef]
- Welsh, D.K.; Takahashi, J.S.; Kay, S.A. Suprachiasmatic nucleus: Cell autonomy and network properties. Annu. Rev. Physiol. 2009, 72, 551–577. [Google Scholar] [CrossRef]
- Hofman, M.; Swaab, D.F. Living by the clock: The circadian pacemaker in older people. Ageing Res. Rev. 2006, 5, 33–51. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, S.; Straume, M.; Tei, H.; Sakaki, Y.; Menaker, M.; Block, G.D. Effects of aging on central and peripheral mammalian clocks. Proc. Natl. Acad. Sci. USA 2002, 99, 10801–10806. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.F.; Czeisler, C.A. Age-related change in the relationship between circadian period, circadian phase, and diurnal preference in humans. Neurosci. Lett. 2002, 318, 117–120. [Google Scholar] [CrossRef]
- Valentinuzzi, V.S.; Scarbrough, K.; Takahashi, J.S.; Turek, F.W. Effects of aging on the circadian rhythm of wheel-running activity in C57BL/6 mice. Am. J. Physiol. Content 1997, 273, R1957–R1964. [Google Scholar] [CrossRef]
- Weinert, D. Age-dependent changes of the circadian system. Chronobiol. Int. 2000, 17, 261–283. [Google Scholar] [CrossRef]
- Yoon, I.-Y.; Kripke, D.F.; Elliott, J.A.; Youngstedt, S.D.; Rex, K.M.; Hauger, R.L. Age-related changes of circadian rhythms and sleep-wake cycles. J. Am. Geriatr. Soc. 2003, 51, 1085–1091. [Google Scholar] [CrossRef]
- Kondratov, R.V.; Antoch, M.P. Circadian proteins in the regulation of cell cycle and genotoxic stress responses. Trends Cell Biol. 2007, 17, 311–317. [Google Scholar] [CrossRef]
- Zhu, Y.; Stevens, R.G.; Hoffman, A.E.; Tjønneland, A.; Vogel, U.; Zheng, T.; Hansen, J. Epigenetic impact of long-term shiftwork: Pilot evidence from circadian genes and whole-genome methylation analysis. Chronobiol. Int. 2011, 28, 852–861. [Google Scholar] [CrossRef]
- Bollati, V.; Baccarelli, A.A.; Sartori, S.; Tarantini, L.; Motta, V.; Rota, F.; Costa, G. Epigenetic effects of shiftwork on blood DNA methylation. Chronobiol. Int. 2010, 27, 1093–1094. [Google Scholar] [CrossRef]
- Burns, A.; Brayne, C.; Folstein, M. Mini-mental state: A practical method for grading the cognitive state of patients for the clinician. M. folstein, S. folstein and P. McHugh, journal of psychiatric research. Int. J. Geriatr. Psychiatry 1998, 13, 285–294. [Google Scholar] [CrossRef]
- Hofer, S.; Flaherty, B.P.; Hoffman, L. Cross-Sectional Analysis of Time-Dependent Data: Mean-Induced Association in Age-Heterogeneous Samples and an Alternative Method Based on Sequential Narrow Age-Cohort Samples. Multivar. Behav. Res. 2006, 41, 165–187. [Google Scholar] [CrossRef]
- Hofer, S.M.; Sliwinski, M.J. Understanding Ageing. An evaluation of research designs for assessing the interdependence of ageing-related changes. Gerontology 2001, 47, 341–352. [Google Scholar] [CrossRef]
- Blackburn, E.; Epel, E.; Lin, J. Human telomere biology: A contributory and interactive factor in aging, disease risks, and protection. Science 2015, 350, 1193–1198. [Google Scholar] [CrossRef]
- Armanios, M.; Blackburn, E.H. The telomere syndromes. Nat. Rev. Genet. 2012, 13, 693–704. [Google Scholar] [CrossRef]
- Glousker, G.; Touzot, F.; Revy, P.; Tzfati, Y.; Savage, S.A. Unraveling the pathogenesis of Hoyeraal-Hreidarsson syndrome, a complex telomere biology disorder. Br. J. Haematol. 2015, 170, 457–471. [Google Scholar] [CrossRef]
- Sahin, E.; Colla, S.; Liesa, M.; Moslehi, J.; Muller, F.; Guo, M.; Cooper, M.; Kotton, D.N.; Fabian, A.J.; Walkley, C.R.; et al. Telomere dysfunction induces metabolic and mitochondrial compromise. Nature 2011, 470, 359–365. [Google Scholar] [CrossRef]
- Bracci, M.; Zabaleta, M.E.; Tartaglione, M.F.; Ledda, C.; Rapisarda, V.; Santarelli, L. Exosomal miR-92a Concentration in the Serum of Shift Workers. Appl. Sci. 2020, 10, 430. [Google Scholar] [CrossRef]
- Butler, R.N.; Sprott, R.; Warner, H.; Bland, J.; Feuers, R.; Forster, M.; Fillit, H.; Mitchell Harman, S.; Hewitt, M.; Hyman, M.; et al. Biomarkers of aging: From primitive organisms to humans. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2004, 59, 560–567. [Google Scholar] [CrossRef]
- Warner, H.R. Current status of efforts to measure and modulate the biological rate of aging. J. Gerontol. Ser. A Biol. Sci. Med Sci. 2004, 59, 692–696. [Google Scholar] [CrossRef]


{kind=link}
| Reference | Type of Workers | Study Design and Setting | n.* | Age | Country | Intervention/Outcome | Biological Sample Analyzed | Analysis Method | Results |
|---|---|---|---|---|---|---|---|---|---|
| Liang et al. 2011 [54] | Female nurses | Nested case-control study from The Nurses’ Health Study | 2409 | n.d. | U.S.A. | Correlations between years of rotating night shifts, self-reported sleep duration, and telomere length | Peripheral blood leukocytes | Real-Time PCR | Sleep duration was positively correlated with telomere length, in particular in women under 50 years. Workers with longer duration of rotating night shifts seem to have short telomere length, (not statistically significant). |
| Chen et al., 2014 [55] | Physicians | Case-control study | 14 physicians working in the emergency department and 13 physicians working in the non-emergency department without night duties | n.d. | Taiwan | SW/Telomere length | Peripheral blood leukocytes | Quantitative fluorescence in situ hybridization | Circadian rhythm of telomerase activity and telomerase reverse transcriptase is involved in human leukocytes of emergency department workers. |
| Pavanello et al., 2017 [56] | Nurses | Cross-sectional study | 84 day workers and 71 night shift workers | 46.7 ± 5.3 | Italy | Alteration of inflammation, found by plasmatic long pentraxin 3, and biological aging, measured by leukocyte telomere length, induced by NSW | Peripheral blood leukocytes | Real-Time PCR | Plasmatic long pentraxin 3, is associated with leukocyte telomere length induced by SW. |
| Samulin Erdem et al., 2017 [57] | Female nurses | Nested case-control study from The Norwegian Nurses Cohort | 563 shift nurses and 619 non shift nurses | 54.47 ± 7.70 | Norway | Breast cancer/SW and Telomere length | Saliva | Real-Time PCR | Telomere shortening is correlated with the duration and intensity of SW and may be a risk factor for breast cancer risk among female shift workers. |
| Chmelar et al., 2017 [58] | Geriatric care professionals | Cross-sectional study | 141 | 44.14 ± 12.35 | Germany | Age, psychosocial work conditions, occupational well-being/telomere length | Peripheral blood leukocytes | Real-Time PCR | Health care supportive psychosocial in workplace have a moderate correlation with influence on the association among SW, aging and telomere length. |
| Pavanello et al., 2019 [59] | Nurses | Cross-sectional study | 155 | n.d. | Italy | SW, perception of work capacity, lung function/oxidative damage and telomere length. | Peripheral blood leukocytes | Real-Time PCR | Association between SW and telomere erosion. |
| Carugno et al., 2019 [60] | Female nurses | Case-control study | 46 | 35.9 ± 5.4 | Italy | Correlations between night shift work and molecular alterations probably connected to a higher carcinogenic risk | Peripheral blood leukocytes | Real-Time PCR | Reduced telomere length in female nurses in SW < 12 years, while a decrease if employed in SW ≥ 12 years. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ledda, C.; Loreto, C.; Rapisarda, V. Telomere Length as a Biomarker of Biological Aging in Shift Workers. Appl. Sci. 2020, 10, 2764. https://doi.org/10.3390/app10082764
Ledda C, Loreto C, Rapisarda V. Telomere Length as a Biomarker of Biological Aging in Shift Workers. Applied Sciences. 2020; 10(8):2764. https://doi.org/10.3390/app10082764
Chicago/Turabian StyleLedda, Caterina, Carla Loreto, and Venerando Rapisarda. 2020. "Telomere Length as a Biomarker of Biological Aging in Shift Workers" Applied Sciences 10, no. 8: 2764. https://doi.org/10.3390/app10082764
APA StyleLedda, C., Loreto, C., & Rapisarda, V. (2020). Telomere Length as a Biomarker of Biological Aging in Shift Workers. Applied Sciences, 10(8), 2764. https://doi.org/10.3390/app10082764