An Evaluation of Vegetation Health in and around Southern African National Parks during the 21st Century (2000–2016)

 ,
, 
 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
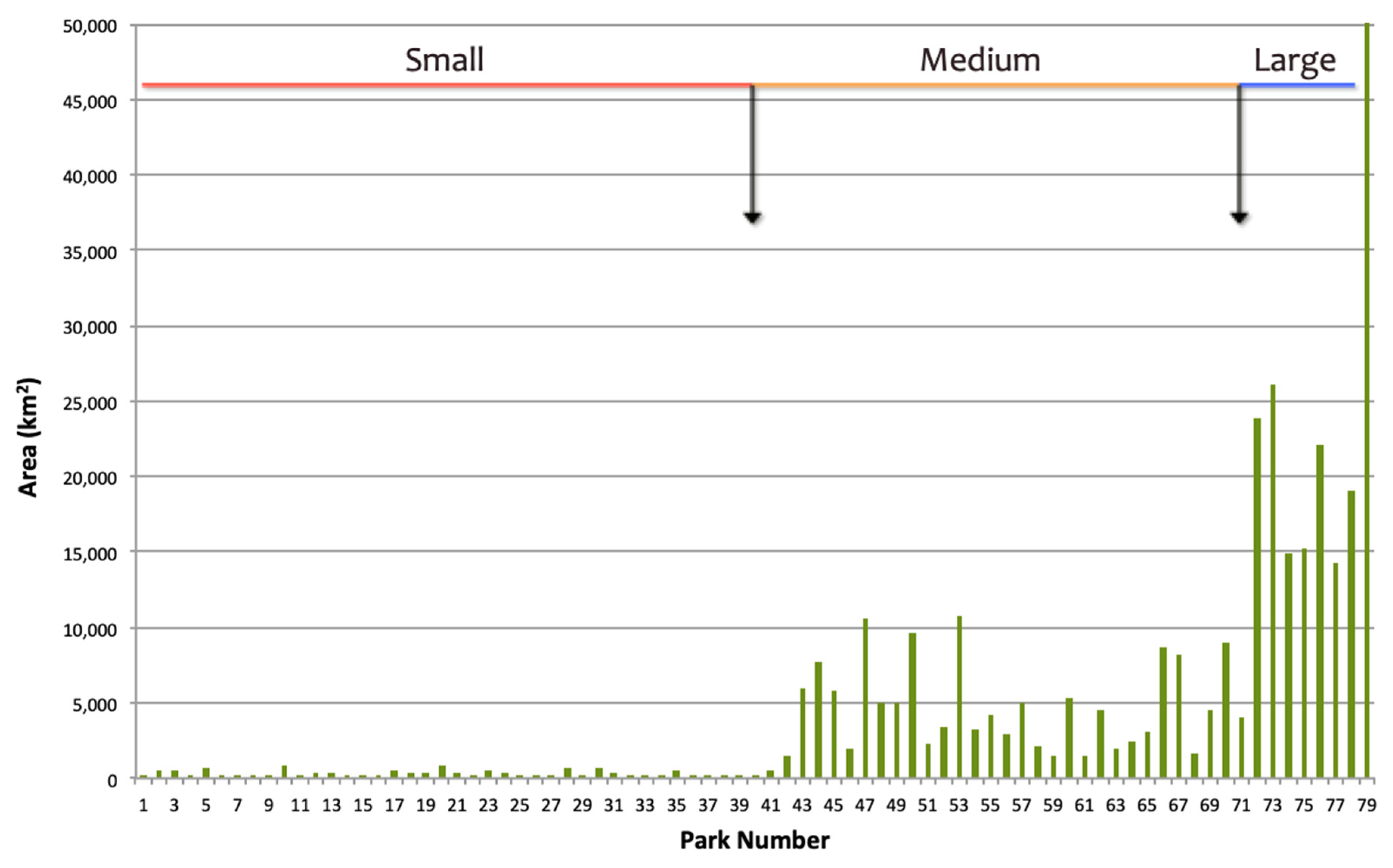
2.1. Study Area
2.2. Understanding Factors of Vegetation Cover: Stepwise Backward Regression
2.2.1. Model Selection
2.2.2. Model Variables
2.2.3. Vegetation Directional Persistence
3. Results
3.1. Stepwise Backward Regression
3.2. Vegetation Directional Persistence
4. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Park | Land Cover (Majority Pixels) |
|---|---|
| Small Parks | |
| Agulhas | Deciduous woodland |
| Augrabies Falls | Sparse grassland |
| Blue Lagoon | Deciduous shrubland with sparse trees |
| Bontebok | Deciduous woodland |
| Cangandala | Deciduous woodland |
| Cape Peninsula | Background |
| Chimanimani | Closed deciduous forest |
| Golden Gate (Qwa Qwa) | Open deciduous shrubland |
| Golden Gate Highlands | Open deciduous shrubland |
| Isangano | Deciduous woodland |
| Karoo | Open grassland with sparse shrubs |
| Kasanka | Deciduous woodland |
| Kazuma Pan | Croplands (>5 background %) |
| Knysna | Background |
| Lake Malawi | Deciduous woodland |
| Lengwe | Deciduous shrubland with sparse trees |
| Liwonde | Deciduous shrubland with sparse trees |
| Lochinvar | Deciduous shrubland with sparse trees |
| Luambe | Deciduous shrubland with sparse trees |
| Lusenga Plain | Closed deciduous forest |
| Mamili | Open deciduous shrubland |
| Mapungubwe | Open deciduous shrubland |
| Marakele | Deciduous woodland |
| Matobo (Matopos) | Deciduous woodland |
| Mosi-Oa-Tunya | Croplands (>5 background %) |
| Mountain Zebra | Open grassland with sparse shrubs |
| Mountain Zebra/Karoo | Open deciduous shrubland |
| Mudumu | Open deciduous shrubland |
| Mushandike | Deciduous woodland |
| Namaqua | Sparse grassland |
| Nyanga | Closed deciduous forest |
| Nyika (Zambia) | Closed deciduous forest |
| Table Mountain | Deciduous woodland |
| Tankwa | Sparse grassland |
| Tankwa Karoo | Sparse grassland |
| Tsitsikamma | Background |
| Vaalbos | Open grassland with sparse shrubs |
| Victoria Falls | Croplands (>5 background %) |
| West Coast | Open deciduous shrubland |
| Wilderness | Closed deciduous forest |
| Zambezi | Deciduous woodland |
| Medium Parks | |
| Addo Elephant | Closed evergreen lowland forest |
| Banhine | Croplands (>5 background %) |
| Bikuar | Deciduous woodland |
| Bwabwata | Open deciduous shrubland |
| Chizarira | Deciduous woodland |
| Chobe | Open deciduous shrubland |
| Gonarezhou | Deciduous woodland |
| Gorongosa | Closed deciduous forest |
| Kalahari Gemsbok | Sparse grassland |
| Kasungu | Deciduous woodland |
| Lavushi Manda | Closed deciduous forest |
| Limpopo | Closed deciduous forest |
| Liuwa Plain | Closed grassland |
| Lower Zambezi | Deciduous shrubland with sparse trees |
| Lukusuzi | Deciduous shrubland with sparse trees |
| Makgadikgadi Pans | Open grassland with sparse shrubs |
| Mana Pools | Croplands (>5 background %) |
| Matusadona | Deciduous woodland |
| Mupa | Croplands (>5 background %) |
| Mweru-Wantipa | Deciduous woodland |
| North Luangwa | Deciduous shrubland with sparse trees |
| Nsumbu | Deciduous woodland |
| Nxai Pan | Open deciduous shrubland |
| Nyika | Deciduous woodland |
| Quicama | Open grassland with sparse shrubs |
| Quirimbas | Deciduous woodland |
| Richtersveld | Sparse grassland |
| Sioma Ngwezi | Open deciduous shrubland |
| South Luangwa | Deciduous shrubland with sparse trees |
| Zinave | Deciduous woodland |
| Large parks | |
| Etosha | Open grassland |
| Gemsbok | Open grassland |
| Hwange | Deciduous woodland |
| Iona | Sandy desert and dunes |
| Kafue | Deciduous woodland |
| Kameia | Closed grassland |
| Kruger | Deciduous woodland |
| Namib-Naukluft | Stony desert |
References
- Chapin, F.S., III; Chapin, M.C.; Matson, P.A.; Vitousek, P. Principles of Terrestrial Ecosystem Ecology; Springer: Berlin/Heidelberg, Germany, 2011; ISBN 1-4419-9504-8. [Google Scholar]
- Vogel, M.; Strohbach, M. Monitoring of savanna degradation in Namibia using Landsat TM/ETM+ data. In Proceedings of the Geoscience and Remote Sensing Symposium, 2009 IEEE International, IGARSS 2009, Cape Town, South Africa, 12–17 July 2009; Volume 3, pp. 931–934. [Google Scholar]
- Hanan, N.; Lehmann, C. Tree-Grass Interactions in Savannas: Paradigms, Contradictions, and Conceptual Models; Taylor and Francis Group: Boca Raton, FL, USA, 2010. [Google Scholar]
- Scholes, R.J.; Walker, B.H. An African Savanna: Synthesis of the Nylsvley Study; Cambridge University Press: Cambridge, UK, 1993; ISBN 0-521-41971-9. [Google Scholar]
- Houghton, E.; Change, I.P. D’experts intergouvernemental sur l’évolution du climat W. In Climate Change 1995: The Science of Climate Change: Contribution of Working Group I to the Second Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 1996; ISBN 978-0-521-56436-6. [Google Scholar]
- Houghton, R.A.; Hackler, J.L.; Lawrence, K.T. The US Carbon Budget: Contributions from Land-Use Change. Science 1999, 285, 574–578. [Google Scholar] [CrossRef]
- Monserud, R.A.; Tchebakova, N.M.; Leemans, R. Global vegetation change predicted by the modified Budyko model. Clim. Chang. 1993, 25, 59–83. [Google Scholar] [CrossRef]
- Ojima, D.S.; Valentine, D.W.; Mosier, A.R.; Parton, W.J.; Schimel, D.S. Effect of land use change on methane oxidation in temperate forest and grassland soils. Chemosphere 1993, 26, 675–685. [Google Scholar] [CrossRef]
- Scholes, R.J.; Archer, S.R. Tree-grass interactions in savannas. Annu. Rev. Ecol. Syst. 1997, 28, 517–544. [Google Scholar] [CrossRef]
- Scholes, R.J.; Hall, D.O. The carbon budget of tropical savannas, woodlands, and grasslands. SCOPE Sci. Comm. Probl. Environ. Int. Counc. Sci. Unions 1996, 56, 69–100. [Google Scholar]
- Andela, N.; Liu, Y.Y.; van Dijk, A.I.J.M.; de Jeu, R.A.M.; McVicar, T.R. Global changes in dryland vegetation dynamics (1988–2008) assessed by satellite remote sensing: Comparing a new passive microwave vegetation density record with reflective greenness data. Biogeosciences 2013, 10, 6657–6676. [Google Scholar] [CrossRef]
- Campo-Bescós, M.A.; Muñoz-Carpena, R.; Southworth, J.; Zhu, L.; Waylen, P.R.; Bunting, E. Combined spatial and temporal effects of environmental controls on long-term monthly NDVI in the southern Africa Savanna. Remote Sens. 2013, 5, 6513–6538. [Google Scholar] [CrossRef]
- Lehmann, C.E.R.; Anderson, T.M.; Sankaran, M.; Higgins, S.I.; Archibald, S.; Hoffmann, W.A.; Hanan, N.P.; Williams, R.J.; Fensham, R.J.; Felfili, J.; et al. Savanna Vegetation-Fire-Climate Relationships Differ among Continents. Science 2014, 343, 548–552. [Google Scholar] [CrossRef]
- Sankaran, M.; Hanan, N.P.; Scholes, R.J.; Ratnam, J.; Augustine, D.J.; Cade, B.S.; Gignoux, J.; Higgins, S.I.; Le Roux, X.; Ludwig, F.; et al. Determinants of woody cover in African savannas. Nature 2005, 438, 846–849. [Google Scholar] [CrossRef]
- Wells, M.; Bradon, K. People and Parks: Linking Protected Area Management with Local Communities; World Bank: Washington, DC, USA, 1992. [Google Scholar]
- Southworth, J.; Zhu, L.; Bunting, E.; Ryan, S.J.; Herrero, H.; Waylen, P.R.; Hill, M.J. Changes in vegetation persistence across global savanna landscapes, 1982–2010. J. Land Use Sci. 2016, 11, 7–32. [Google Scholar] [CrossRef]
- Carlson, T.N.; Ripley, D.A. On the relation between NDVI, fractional vegetation cover, and leaf area index. Remote Sens. Environ. 1997, 62, 241–252. [Google Scholar] [CrossRef]
- Bégué, A.; Vintrou, E.; Ruelland, D.; Claden, M.; Dessay, N. Can a 25-year trend in Soudano-Sahelian vegetation dynamics be interpreted in terms of land use change? A remote sensing approach. Glob. Environ. Chang. 2011, 21, 413–420. [Google Scholar] [CrossRef]
- Jiang, Z.; Huete, A.R.; Chen, J.; Chen, Y.; Li, J.; Yan, G.; Zhang, X. Analysis of NDVI and scaled difference vegetation index retrievals of vegetation fraction. Remote Sens. Environ. 2006, 101, 366–378. [Google Scholar] [CrossRef]
- Lambin, E.F.; Ehrlich, D. Land-cover changes in sub-saharan Africa (1982–1991): Application of a change index based on remotely sensed surface temperature and vegetation indices at a continental scale. Remote Sens. Environ. 1997, 61, 181–200. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Wang, J.; Rich, P.M.; Price, K.P.; Kettle, W.D. Relations between NDVI, Grassland Production, and Crop Yield in the Central Great Plains. Geocarto Int. 2005, 20, 5–11. [Google Scholar] [CrossRef]
- Waylen, P.; Southworth, J.; Gibbes, C.; Tsai, H. Time Series Analysis of Land Cover Change: Developing Statistical Tools to Determine Significance of Land Cover Changes in Persistence Analyses. Remote Sens. 2014, 6, 4473–4497. [Google Scholar] [CrossRef]
- Zhu, Z.; Piao, S.; Myneni, R.B.; Huang, M.; Zeng, Z.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A.; et al. Greening of the Earth and its drivers. Nat. Clim. Chang. 2016, 6, 791–795. [Google Scholar] [CrossRef]
- De Jong, R.; de Bruin, S.; de Wit, A.; Schaepman, M.E.; Dent, D.L. Analysis of monotonic greening and browning trends from global NDVI time-series. Remote Sens. Environ. 2011, 115, 692–702. [Google Scholar] [CrossRef]
- Olsson, L.; Eklundh, L.; Ardö, J. A recent greening of the Sahel—Trends, patterns and potential causes. J. Arid Environ. 2005, 63, 556–566. [Google Scholar] [CrossRef]
- Zhang, K.; Kimball, J.S.; Nemani, R.R.; Running, S.W.; Hong, Y.; Gourley, J.J.; Yu, Z. Vegetation Greening and Climate Change Promote Multidecadal Rises of Global Land Evapotranspiration. Sci. Rep. 2015, 5, 15956. [Google Scholar] [CrossRef] [PubMed]
- Campo-Bescós, M.A.; Muñoz-Carpena, R.; Kaplan, D.A.; Southworth, J.; Zhu, L.; Waylen, P.R. Beyond Precipitation: Physiographic Gradients Dictate the Relative Importance of Environmental Drivers on Savanna Vegetation. PLoS ONE 2013, 8, e72348. [Google Scholar] [CrossRef] [PubMed]
- Staver, A.C.; Archibald, S.; Levin, S. Tree cover in sub-Saharan Africa: Rainfall and fire constrain forest and savanna as alternative stable states. Ecology 2011, 92, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Bunting, E.L.; Fullman, T.; Kiker, G.; Southworth, J. Utilization of the SAVANNA model to analyze future patterns of vegetation cover in Kruger National Park under changing climate. Ecol. Model. 2016, 342, 147–160. [Google Scholar] [CrossRef]
- Verweij, R.J.T.; Higgins, S.I.; Bond, W.J.; February, E.C. Water sourcing by trees in a mesic savanna: Responses to severing deep and shallow roots. Environ. Exp. Bot. 2011, 74, 229–236. [Google Scholar] [CrossRef]
- Campbell, A.; Child, G. The impact of man on the environment of Botswana. Botsw. Notes Rec. 1971, 3, 91–110. [Google Scholar]
- Ringrose, S.; Matheson, W.; Wolski, P.; Huntsman-Mapila, P. Vegetation cover trends along the Botswana Kalahari transect. J. Arid Environ. 2003, 54, 297–317. [Google Scholar] [CrossRef]
- Nagy, L.; Grabherr, G. The Biology of Alpine Habitats; OUP Oxford: Oxford, UK, 2009; ISBN 978-0-19-856703-5. [Google Scholar]
- Campbell, B.M. The Miombo in Transition: Woodlands and Welfare in Africa; CIFOR: Bogor, Indonesia, 1996; ISBN 978-979-8764-07-3. [Google Scholar]
- Gambiza, J. A Primer on Savanna Ecology; Institute of Environmental Studies, University of Zimbabwe: Harare, Zimbabwe, 2001. [Google Scholar]
- Newmark, W.D. Insularization of Tanzanian Parks and the Local Extinction of Large Mammals. Conserv. Biol. 1996, 10, 1549–1556. [Google Scholar] [CrossRef]
- DeFries, R.; Hansen, A.; Turner, B.L.; Reid, R.; Liu, J. Land Use Change around Protected Areas: Management to Balance Human Needs and Ecological Function. Ecol. Appl. 2007, 17, 1031–1038. [Google Scholar] [CrossRef]
- Saura, S.; Bertzky, B.; Bastin, L.; Battistella, L.; Mandrici, A.; Dubois, G. Protected area connectivity: Shortfalls in global targets and country-level priorities. Biol. Conserv. 2018, 219, 53–67. [Google Scholar] [CrossRef]
- Protected Planet Protected Planet: World Database on Protected Areas. Available online: https://www.protectedplanet.net/ (accessed on 23 February 2020).
- Global Historical Climatology Network (GHCN)|National Centers for Environmental Information (NCEI) Formerly Known as National Climatic Data Center (NCDC). Available online: https://www.ncdc.noaa.gov/data-access/land-based-station-data/land-based-datasets/global-historical-climatology-network-ghcn (accessed on 28 February 2020).
- Friedl, M.A.; Sulla-Menashe, D.; Tan, B.; Schneider, A.; Ramankutty, N.; Sibley, A.; Huang, X. MODIS Collection 5 global land cover: Algorithm refinements and characterization of new datasets. Remote Sens. Environ. 2010, 114, 168–182. [Google Scholar] [CrossRef]
- Home—Peace Parks Foundation. Available online: https://www.peaceparks.org/ (accessed on 24 September 2019).
- Hocking, R.R. A Biometrics Invited Paper. The Analysis and Selection of Variables in Linear Regression. Biometrics 1976, 32, 1–49. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Measuring Vegetation (NDVI & EVI). Available online: https://earthobservatory.nasa.gov/features/MeasuringVegetation (accessed on 24 September 2019).
- Thematic Mapping Engine—Thematicmapping.org. Available online: http://thematicmapping.org/engine/ (accessed on 24 September 2019).
- WorldPop. Available online: https://www.worldpop.org/ (accessed on 24 September 2019).
- Ensslin, A.; Rutten, G.; Pommer, U.; Zimmermann, R.; Hemp, A.; Fischer, M. Effects of elevation and land use on the biomass of trees, shrubs and herbs at Mount Kilimanjaro. Ecosphere 2015, 6, 1–15. [Google Scholar] [CrossRef]
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The Shuttle Radar Topography Mission. Rev. Geophys. 2007, 45. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human Domination of Earth’s Ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef]
- Barnes, M.D.; Glew, L.; Wyborn, C.; Craigie, I.D. Prevent perverse outcomes from global protected area policy. Nat. Ecol. Evol. 2018, 2, 759–762. [Google Scholar] [CrossRef]
- Rodrigues, A.S.L.; Andelman, S.J.; Bakarr, M.I.; Boitani, L.; Brooks, T.M.; Cowling, R.M.; Fishpool, L.D.C.; da Fonseca, G.A.B.; Gaston, K.J.; Hoffmann, M.; et al. Effectiveness of the global protected area network in representing species diversity. Nature 2004, 428, 640–643. [Google Scholar] [CrossRef]
- Herrero, H.; Waylen, P.; Southworth, J.; Khatami, R.; Yang, D.; Child, B. A Healthy Park Needs Healthy Vegetation: The Story of Gorongosa National Park in the 21st Century. Remote Sens. 2020, 12, 476. [Google Scholar] [CrossRef]
- Herrero, H.V.; Southworth, J.; Bunting, E. Utilizing Multiple Lines of Evidence to Determine Landscape Degradation within Protected Area Landscapes: A Case Study of Chobe National Park, Botswana from 1982 to 2011. Remote Sens. 2016, 8, 623. [Google Scholar] [CrossRef]
- Child, B. Zimbabwe’s CAMPFIRE programme: Using the high value of wildlife recreation to revolutionize natural resource management in communal areas. Commonw. For. Rev. 1993, 72, 284–296. [Google Scholar]






| Area/Time | R2 | Best Model | Intercept | Akaike’s Criterion |
|---|---|---|---|---|
| Parks DJF 2000–2004 | 0.68 | y~X2 + X4 + X5 + X7 | 2.346 × 10−1 | −331.36 |
| Buffers DJF 2000–2004 | 0.62 | y~X2 + X4 + X5 + X6 + X7 | −4.158 × 10−2 | −276.89 |
| Parks DJF 2005–2008 | 0.67 | y~X3 + X4 + X5 + X7 | 1.813 × 10−1 | −326.71 |
| Buffers DJF 2005–2008 | 0.62 | y~X2 + X4 + X5 + X6 + X7 | −3.596 × 10−2 | −275.83 |
| Parks DJF 2009–2012 | 0.66 | y~X4 + X5 + X7 | 1.656 × 10−1 | −325.82 |
| Buffers DJF 2009–2012 | 0.61 | y~X2 + X4 + X5 + X6 + X7 | −2.330 × 10−2 | −272.69 |
| Parks DJF 2013–2016 | 0.65 | y~X2 + X3 + X4 + X5 + X7 | 2.502 × 10−1 | −317.62 |
| Buffers DJF 2013–2016 | 0.62 | y~X2 + X4 + X5 + X6 + X7 | −3.360 × 10−2 | −277.28 |
| Area/Time | R2 | Best Model | Intercept | Akaike’s Criterion |
|---|---|---|---|---|
| Parks MAM 2000–2004 | 0.69 | y~X2 + X4 + X5 + X6 + X7 | 3.383 × 10−1 | −350.74 |
| Buffers MAM 2000–2004 | 0.61 | y~X2 + X4 + X5 + X7 | 3.079 × 10−2 | −293.31 |
| Parks MAM 2005–2008 | 0.69 | y~X2 + X4 + X5 + X6 + X7 | 3.004 × 10−1 | −352.89 |
| Buffers MAM 2005–2008 | 0.62 | y~X2 + X4 + X5 + X7 | 3.756 × 10−2 | −297.45 |
| Parks MAM 2009–2012 | 0.67 | y~X2 + X4 + X5 + X7 | 3.108 × 10−1 | −348.22 |
| Buffers MAM 2009–2012 | 0.62 | y~X2 + X4 + X5 + X7 | 3.090 × 10−2 | −293.39 |
| Parks MAM 2013–2016 | 0.69 | y~X2 + X4 + X5 + X6 + X7 | 3.363 × 10−1 | −343.49 |
| Buffers MAM 2013–2016 | 0.62 | y~X2 + X4 + X5 + X7 | 3.547 × 10−2 | −292.97 |
| Area/Time | Country | Land Cover | Area (km2) | MAP (mm) | Mean Population | Mean Elevation | Transfrontier (Y/N) |
|---|---|---|---|---|---|---|---|
| Parks DJF 2000–2004 | + **** | − * | |||||
| Buffers DJF 2000–2004 | * | + **** | − **** | + * | + *** | ||
| Parks DJF 2005–2008 | − * | + **** | − * | ||||
| Buffers DJF 2005–2008 | * | + **** | − **** | + *** | |||
| Parks DJF 2009–2012 | + **** | + *** | |||||
| Buffers DJF 2009–2012 | * | + **** | − **** | + *** | |||
| Parks DJF 2013–2016 | + **** | + *** | |||||
| Buffers DJF 2013–2016 | + **** | − **** | + * | + *** | |||
| Variable Significance: 0 = **** 0.001 = *** 0.01 = ** 0.05 = * 1 = none | |||||||
| Area/Time | Country | Land Cover | Area (km2) | MAP (mm) | Mean Population | Mean Elevation | Transfrontier (Y/N) |
|---|---|---|---|---|---|---|---|
| Parks MAM 2000–2004 | ** | + **** | − ** | − ** | + ** | ||
| Buffers MAM 2000–2004 | ** | + **** | − **** | + ** | |||
| Parks MAM 2005–2008 | * | + **** | + ** | ||||
| Buffers MAM 2005–2008 | ** | + **** | − **** | + ** | |||
| Parks MAM 2009–2012 | ** | + **** | + ** | ||||
| Buffers MAM 2009–2012 | *** | + **** | − **** | + ** | |||
| Parks MAM 2013–2016 | ** | + **** | − * | − * | + ** | ||
| Buffers MAM 2013–2016 | ** | + **** | − **** | + *** | |||
| Variable Significance: 0 = **** 0.001 = *** 0.01 = ** 0.05 = * 1 = none | |||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrero, H.; Southworth, J.; Muir, C.; Khatami, R.; Bunting, E.; Child, B. An Evaluation of Vegetation Health in and around Southern African National Parks during the 21st Century (2000–2016). Appl. Sci. 2020, 10, 2366. https://doi.org/10.3390/app10072366
Herrero H, Southworth J, Muir C, Khatami R, Bunting E, Child B. An Evaluation of Vegetation Health in and around Southern African National Parks during the 21st Century (2000–2016). Applied Sciences. 2020; 10(7):2366. https://doi.org/10.3390/app10072366
Chicago/Turabian StyleHerrero, Hannah, Jane Southworth, Carly Muir, Reza Khatami, Erin Bunting, and Brian Child. 2020. "An Evaluation of Vegetation Health in and around Southern African National Parks during the 21st Century (2000–2016)" Applied Sciences 10, no. 7: 2366. https://doi.org/10.3390/app10072366
APA StyleHerrero, H., Southworth, J., Muir, C., Khatami, R., Bunting, E., & Child, B. (2020). An Evaluation of Vegetation Health in and around Southern African National Parks during the 21st Century (2000–2016). Applied Sciences, 10(7), 2366. https://doi.org/10.3390/app10072366