Combined Effects of Water Deficit, Exogenous Ethylene Application and Root Symbioses on Trigonelline and ABA Accumulation in Fenugreek


 ,
,  ,
,  , and
, and 
Abstract
Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Planning
2.2. Fenugreek Sowing and Treatments
2.3. Assessment of Morphometric Parameters and Symbiosis Development
2.4. Trigonelline and ABA Quantification
2.5. Statistical Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oksman-Caldentey, K.M.; Inzé, D. Plant cell factories in the post-genomic era: New ways to produce designer secondary metabolites. Trends Plant Sci. 2004, 9, 433–440. [Google Scholar] [CrossRef]
- Jimenez-Garcia, S.N.; Vazquez-Cruz, M.A.; Guevara-Gonzalez, R.G.; Torres-Pacheco, I.; Cruz-Hernandez, A.; Feregrino-Perez, A.A. Current approaches for enhanced expression of secondary metabolites as bioactive compounds in plants for agronomic and human health purposes–a review. Pol. J. Food Nutr. Sci. 2013, 63, 67–78. [Google Scholar] [CrossRef]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef]
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef] [PubMed]
- Pistelli, L.; Tonelli, M.; Pellegrini, E.; Cotrozzi, L.; Pucciariello, C.; Trivellini, A.; Lorenzini, G.; Nali, C. Accumulation of rosmarinic acid and behaviour of ROS processing systems in Melissa officinalis L. under heat stress. Ind. Crop Prod. 2019, 138, 111469. [Google Scholar] [CrossRef]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef] [PubMed]
- Puig, D.G.; Perez, M.L.; Fuster, M.D.; Ortuno, A.; Sabater, F.; Porras, I.; Lidón, A.G.; Del Rio, J.A. Effect of ethylene on naringin, narirutin and nootkatone accumulation in grapefruit. Planta Med. 1995, 61, 283–285. [Google Scholar] [CrossRef] [PubMed]
- Villarreal-García, D.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Plants as biofactories: Postharvest stress-induced accumulation of phenolic compounds and glucosinolates in broccoli subjected to wounding stress and exogenous phytohormones. Front. Plant Sci. 2016, 7, 45. [Google Scholar] [CrossRef]
- Kieber, J.J. The ethylene response pathway in Arabidopsis. Ann. Rev. Plant Biol. 1997, 48, 277–296. [Google Scholar] [CrossRef]
- Cho, G.H.; Kim, D.I.; Pedersen, H.; Chin, C.K. Ethephon enhancement of secondary metabolite synthesis in plant cell cultures. Biotechnol. Prog. 1988, 4, 184–188. [Google Scholar] [CrossRef]
- Balestrini, R.; Lumini, E. Focus on mycorrhizal symbioses. Appl. Soil Ecol. 2019, 123, 299–304. [Google Scholar] [CrossRef]
- Fester, T.; Hause, G. Accumulation of reactive oxygen species in arbuscular mycorrhizal roots. Mycorrhiza 2005, 15, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Lohse, S.; Schliemann, W.; Ammer, C.; Kopka, J.; Strack, D.; Fester, T. Organization and metabolism of plastids and mitochondria in arbuscular mycorrhizal roots of Medicago truncatula. Plant Physiol. 2005, 139, 329–340. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schliemann, W.; Ammer, C.; Strack, D. Metabolite profiling of mycorrhizal roots of Medicago truncatula. Phytochemistry 2008, 69, 112–146. [Google Scholar] [CrossRef] [PubMed]
- Sbrana, C.; Avio, L.; Giovannetti, M. Beneficial mycorrhizal symbionts affecting the production of health promoting phytochemicals. Electrophoresis 2014, 35, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Wipf, D.; Mongelard, G.; van Tuinen, D.; Gutierrez, L.; Casieri, L. Transcriptional responses of Medicago truncatula upon sulfur deficiency stress and arbuscular mycorrhizal symbiosis. Front. Plant Sci. 2014, 5, 680. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, R.; Müller, C. Leaf metabolome in arbuscular mycorrhizal symbiosis. Curr. Opin. Plant Biol. 2015, 26, 120–126. [Google Scholar] [CrossRef]
- Cervantes-Gámez, R.G.; Bueno-Ibarra, M.A.; Cruz-Mendívil, A.; Calderón-Vázquez, C.L.; Ramírez-Douriet, C.M.; Maldonado-Mendoza, I.E.; Villalobos-López, M.A.; Valdez-Ortíz, A.; López-Meyer, M. Arbuscular mycorrhizal symbiosis-induced expression changes in Solanum lycopersicum leaves revealed by RNA-seq analysis. Plant Mol. Biol. Rep. 2016, 34, 89–102. [Google Scholar] [CrossRef]
- Berruti, A.; Lumini, E.; Balestrini, R.; Bianciotto, V. Arbuscular mycorrhizal fungi as natural biofertilizers: let’s benefit from past successes. Front. Microbiol. 2016, 6, 1559. [Google Scholar] [CrossRef]
- Balestrini, R.; Chitarra, W.; Antoniou, C.; Ruocco, M.; Fotopoulos, V. Improvement of plant performance under water deficit with the employment of biological and chemical priming agents. J. Agric. Sci. 2018, 156, 680–688. [Google Scholar] [CrossRef]
- Pedone-Bonfim, M.V.L.; da Silva, F.S.B.; Maia, L.C. Production of secondary metabolites by mycorrhizal plants with medicinal or nutritional potential. Acta Physiol. Plant. 2015, 37, 27. [Google Scholar] [CrossRef]
- Flammang, A.M.; Cifone, M.A.; Erexson, G.L.; Stankowski Jr, L.F. Genotoxicity testing of a fenugreek extract. Food Chem. Toxicol. 2004, 42, 1769–1775. [Google Scholar] [CrossRef] [PubMed]
- Altuntaş, E.; Özgöz, E.; Taşer, Ö.F. Some physical properties of fenugreek (Trigonella foenum-graecum L.) seeds. J. Food Eng. 2005, 71, 37–43. [Google Scholar] [CrossRef]
- Acharya, S.N.; Thomas, J.E.; Basu, S.K. Fenugreek: An “old world” crop for the “new world”. Biodiversity 2006, 7, 27–30. [Google Scholar] [CrossRef]
- Ashihara, H.; Ludwig, I.A.; Katahira, R.; Yokota, T.; Fujimura, T.; Crozier, A. Trigonelline and related nicotinic acid metabolites: Occurrence, biosynthesis, taxonomic considerations, and their roles in planta and in human health. Phytochem. Rev. 2015, 14, 765–798. [Google Scholar] [CrossRef]
- Minorsky, P.V. Trigonelline: A diverse regulator in plants. Plant Physiol. 2002, 128, 7–8. [Google Scholar] [CrossRef]
- Raheleh, A.; Hasanloo, T.; Khosroshahli, M. Evaluation of trigonelline production in Trigonella foenum-graecum hairy root cultures of two Iranian masses. POJ 2011, 4, 408–412. [Google Scholar]
- Qaderi, A.; Akbari, Z.; Kalateh-jari, S.; Fatehi, F.; Tolyat, M.; Jalali Moghadam, M.; Naghdi Badi, H. Improving trigonelline production in hairy root culture of fenugreek (Trigonella foenum-graecum). J. Med. Plant 2016, 3, 73–80. [Google Scholar]
- Dadrasan, M.; Chaichi, M.R.; Pourbabaee, A.A.; Yazdani, D.; Keshavarz-Afshar, R. Deficit irrigation and biological fertilizer influence on yield and trigonelline production of fenugreek. Ind. Crop Prod. 2015, 77, 156–162. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Mesure du taux de mycorhization VA d’un système radiculaire. Recherche de méthode d’estimation ayant une signification fonctionnelle. In Physiological and genetical aspects of mycorrhizae. In Proceedings of the 1st European Symposium on Mycorrhizae, Dijon, France, 1–5 July 1985; pp. 217–221. [Google Scholar]
- Foo, E. Plant hormones play common and divergent roles in nodulation and arbuscular mycorrhizal symbioses. In The Model Legume Medicago truncatula; Jhon Wiley & Sons, Inc.: Hoboken, NJ, USA, 2020; pp. 753–765. [Google Scholar]
- Chitarra, W.; Pagliarani, C.; Maserti, B.; Lumini, E.; Siciliano, I.; Cascone, P.; Schubert, A.; Gambino, G.; Balestrini, R.; Guerrieri, E. Insights on the impact of arbuscular mycorrhizal symbiosis on tomato tolerance to water stress. Plant Physiol. 2016, 171, 1009–1023. [Google Scholar] [CrossRef]
- Volpe, V.; Chitarra, W.; Cascone, P.; Volpe, M.G.; Bartolini, P.; Moneti, G.; Pieraccini, G.; Di Serio, C.; Maserti, B.; Guerrieri, E.; et al. The association with two different arbuscular mycorrhizal fungi differently affects water stress tolerance in tomato. Front. Plant Sci. 2018, 9, 1480. [Google Scholar] [CrossRef] [PubMed]
- Alagna, F.; Balestrini, R.; Chitarra, W.; Marsico, A.D.; Nerva, L. Getting ready with the priming: Innovative weapons against biotic and abiotic crop enemies in a global changing scenario. In Priming-Mediated Stress and Cross-Stress Tolerance in Crop Plants; Academic Press: Cambridge, MA, USA, 2020; pp. 35–56. [Google Scholar]
- Yu, H.; Zhang, Y.; Xie, Y.; Wang, Y.; Duan, L.; Zhang, M.; Li, Z. Ethephon improved drought tolerance in maize seedlings by modulating cuticular wax biosynthesis and membrane stability. J. Plant Physiol. 2017, 214, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Barba-Espín, G.; Glied, S.; Crocoll, C.; Dzhanfezova, T.; Joernsgaard, B.; Okkels, F.; Lutken, H.; Muller, R. Foliar-applied ethephon enhances the content of anthocyanin of black carrot roots (Daucus carota ssp. Sativus var. atrorubens Alef.). BMC Plant Biol. 2017, 17, 70. [Google Scholar]
- Oncina, R.; Del Río, J.A.; Gomez, P.; Ortuno, A. Effect of ethylene on diosgenin accumulation in callus cultures of Trigonella foenum-graecum L. Food Chem. 2002, 76, 475–479. [Google Scholar] [CrossRef]
- Gomez, S.; Roy, S.K.; Pal, R.K. Primary processing of fenugreek (Trigonella foenum graecum L.)–An eco-friendly approach for convenience and quality. Plant Food Hum. Nutr. 2003, 58, 1–10. [Google Scholar] [CrossRef]
- Zamani, Z.; Amiri, H.; Ismaili, A. Improving drought stress tolerance in fenugreek (Trigonella foenum-graecum) by exogenous melatonin. Plant Biosyst. 2019. [Google Scholar] [CrossRef]
- Caser, M.; Chitarra, W.; D’Angiolillo, F.; Perrone, I.; Demasi, S.; Lovisolo, C.; Pistelli, L.; Pistelli, L.; Scariot, V. Drought stress adaptation modulates plant secondary metabolite in Salvia dolomitica Codd. Ind. Crop Prod. 2019, 129, 85–96. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.; De Bruin, S.; Luckerhoff, L.; Van Logtestijn, R.S.; Schlaeppi, K. A widespread plant-fungal-bacterial symbiosis promotes plant biodiversity, plant nutrition and seedling recruitment. ISME J. 2016, 10, 389–399. [Google Scholar] [CrossRef]
- Rojas-Andrade, R.; Cerda-García-Rojas, C.; Frías-Hernández, J.; Dendooven, L.; Olalde-Portugal, V.; Ramos-Valdivia, A. Changes in the concentration of trigonelline in a semi-arid leguminous plant (Prosopis laevigata) induced by an arbuscular mycorrhizal fungus during the presymbiotic phase. Mycorrhiza 2003, 13, 49–52. [Google Scholar] [CrossRef]
- Metwally, R.A.; Abdelhameed, R.E. Synergistic effect of arbuscular mycorrhizal fungi on growth and physiology of salt-stressed Trigonella foenum-graecum plants. Biocatal. Agric. Biotechnol. 2018, 16, 538–544. [Google Scholar] [CrossRef]
- Evelin, H.; Kapoor, R. Arbuscular mycorrhizal symbiosis modulates antioxidant response in salt-stressed Trigonella foenum-graecum plants. Mycorrhiza 2014, 24, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [PubMed]



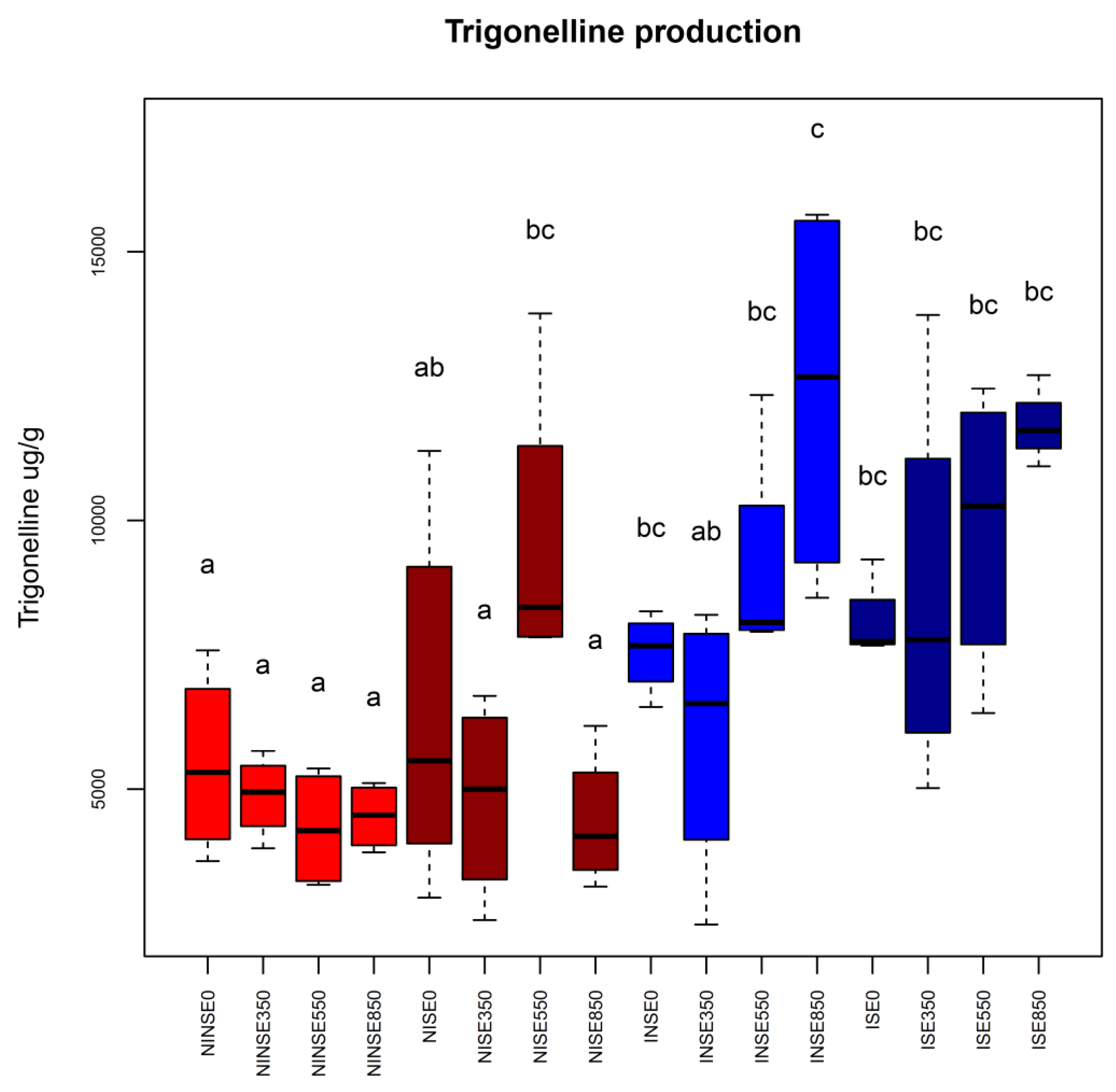{kind=link}
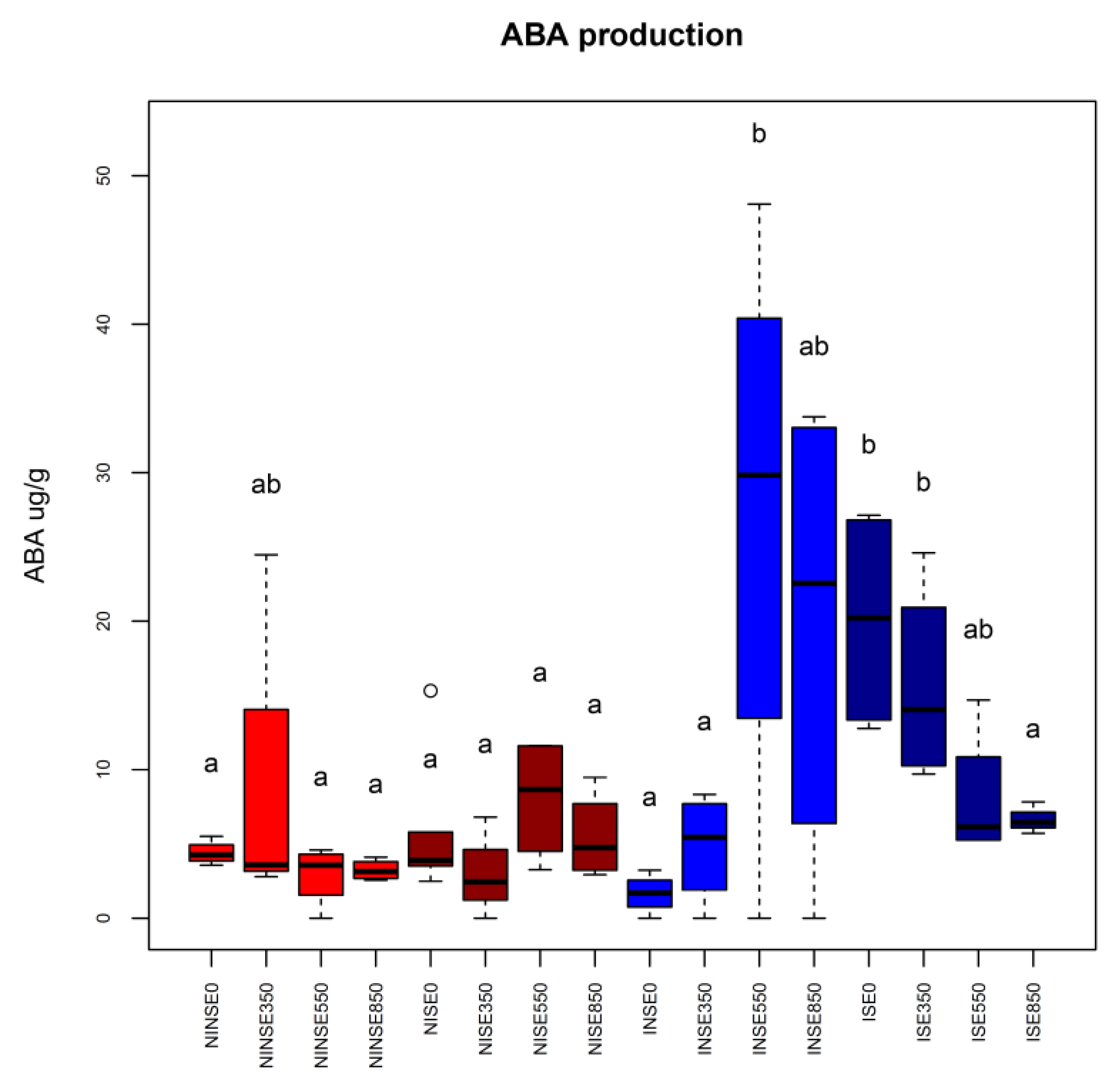{kind=link}
| Source of Variance | Stem h(cm) | Stem DW (g) | Root DW (g) | Fruit # | Fruit DW (g) |
|---|---|---|---|---|---|
| Stress | ns | ns | ns | ns | ns |
| Inoculation | ns | ns | ns | * | ns |
| Ethephon | ns | ns | ns | ns | ns |
| Stress*Inoculation | ns | ns | ns | ns | ns |
| Stress* Ethephon | ns | ns | ns | ns | ns |
| Inoculation* Ethephon | ns | ns | ns | ns | ns |
| Stress*Inoculation*Ethephon | ns | ns | ns | ns | ns |
| Stress | |||||
| NS | 16.51 ± 3.91 | 0.03 ± 0.02 | 0.03 ± 0.02 | 0.50 ± 0.46 | 0.02 ± 0.02 |
| S | 15.33 ± 2.12 | 0.03 ± 0.01 | 0.04 ± 0.02 | 0.50 ± 0.40 | 0.03 ± 0.05 |
| Inoculation | |||||
| NI | 16.73 ± 2.67 | 0.03 ± 0.01 | 0.03 ± 0.02 | 0.65 ± 0.53a | 0.04 ± 0.05 |
| I | 15.11 ± 3.47 | 0.03 ± 0.02 | 0.03 ± 0.02 | 0.34 ± 0.33b | 0.01 ± 0.02 |
| Ethephon | |||||
| E0 | 16.90 ± 3.65 | 0.04 ± 0.02 | 0.03 ± 0.01 | 0.63 ± 0.43 | 0.02 ± 0.03 |
| E350 | 14.71 ± 2.60 | 0.02 ± 0.01 | 0.04 ± 0.02 | 0.56 ± 0.52 | 0.05 ± 0.08 |
| E550 | 15.91 ± 3.35 | 0.04 ± 0.02 | 0.02 ± 0.02 | 0.50 ± 0.40 | 0.02 ± 0.02 |
| E850 | 16.09 ± 2.45 | 0.03 ± 0.02 | 0.04 ± 0.02 | 0.31 ± 0.38 | 0.01 ± 0.01 |
| Stress*Inoculation | |||||
| NINS | 17.16 ± 3.70 | 0.04 ± 0.01 | 0.04 ± 0.02 | 0.62 ± 0.54 | 0.02 ± 0.02 |
| NIS | 16.31 ± 1.40 | 0.04 ± 0.01 | 0.03 ± 0.02 | 0.68 ± 0.53 | 0.05 ± 0.08 |
| INS | 15.88 ± 4.11 | 0.04 ± 0.02 | 0.03 ± 0.02 | 0.38 ± 0.40 | 0.01 ± 0.02 |
| IS | 14.34 ± 2.84 | 0.03 ± 0.01 | 0.04 ± 0.02 | 0.31 ± 0.27 | 0.01 ± 0.02 |
| Stress*Ethephon | |||||
| NSE0 | 18.69 ± 5.14 | 0.05 ± 0.02 | 0.03 ± 0.01 | 0.50 ± 0.58 | 0.02 ± 0.03 |
| NSE350 | 14.57 ± 3.28 | 0.03 ± 0.01 | 0.04 ± 0.01 | 0.50 ± 0.50 | 0.02 ± 0.02 |
| NSE550 | 15.57 ± 4.34 | 0.03 ± 0.02 | 0.02 ± 0.02 | 0.75 ± 0.50 | 0.03 ± 0.03 |
| NSE850 | 17.25 ± 2.86 | 0.04 ± 0.02 | 0.04 ± 0.03 | 0.25 ± 0.29 | 0.01 ± 0.01 |
| SE0 | 15.12 ± 2.16 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.75 ± 0.29 | 0.03 ± 0.03 |
| SE350 | 15.01 ± 1.92 | 0.03 ± 0.01 | 0.04 ± 0.02 | 0.63 ± 0.54 | 0.09 ± 0.15 |
| SE550 | 16.25 ± 2.36 | 0.04 ± 0.02 | 0.03 ± 0.02 | 0.25 ± 0.29 | 0.01 ± 0.01 |
| SE850 | 14.94 ± 2.04 | 0.04 ± 0.01 | 0.04 ± 0.02 | 0.38 ± 0.48 | 0.01 ± 0.01 |
| Inoculation*Ethephon | |||||
| NIE0 | 17.32 ± 3.79 | 0.05 ± 0.02 | 0.03 ± 0.01 | 0.75 ± 0.29 | 0.02 ± 0.02 |
| NIE350 | 15.44 ± 1.61 | 0.03 ± 0.01 | 0.04 ± 0.01 | 0.62 ± 0.54 | 0.09 ± 0.15 |
| NIE550 | 16.50 ± 2.51 | 0.04 ± 0.01 | 0.03 ± 0.02 | 0.63 ± 0.54 | 0.02 ± 0.03 |
| NIE850 | 17.69 ± 2.28 | 0.04 ± 0.02 | 0.03 ± 0.02 | 0.63 ± 0.77 | 0.02 ± 0.02 |
| IE0 | 16.50 ± 3.52 | 0.04 ± 0.01 | 0.03 ± 0.01 | 0.50 ± 0.58 | 0.03 ± 0.04 |
| IE350 | 14.13 ± 3.59 | 0.025 ± 0.01 | 0.04 ± 0.02 | 0.50 ± 0.50 | 0.02 ± 0.02 |
| IE550 | 15.32 ± 4.20 | 0.03 ± 0.03 | 0.02 ± 0.02 | 0.38 ± 0.25 | 0.01 ± 0.01 |
| IE850 | 14.5 ± 2.61 | 0.03 ± 0.01 | 0.05 ± 0.02 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Stress*Inoculation*Ethephon | |||||
| NINSE0 | 19.63 ± 6.63 | 0.05 ± 0.02 | 0.04 ± 0.01 | 0.50 ± 0.58 | 0.01 ± 0.02 |
| NINS350 | 15.25 ± 1.66 | 0.03 ± 0.00 | 0.04 ± 0.01 | 0.75 ± 0.50 | 0.03 ± 0.02 |
| NINS550 | 15.75 ± 3.57 | 0.03 ± 0.01 | 0.03 ± 0.02 | 0.75 ± 0.50 | 0.03 ± 0.03 |
| NINSE850 | 18.00 ± 2.92 | 0.04 ± 0.02 | 0.03 ± 0.03 | 0.50 ± 0.58 | 0.01 ± 0.02 |
| NISE0 | 15.00 ± 0.94 | 0.04 ± 0.01 | 0.02 ± 0.01 | 1.00 ± 0.00 | 0.03 ± 0.01 |
| NSE350 | 15.63 ± 1.55 | 0.03 ± 0.01 | 0.04 ± 0.02 | 0.50 ± 0.58 | 0.15 ± 0.27 |
| NISE550 | 17.25 ± 1.44 | 0.04 ± 0.01 | 0.02 ± 0.02 | 0.50 ± 0.58 | 0.01 ± 0.02 |
| NISE850 | 17.38 ± 1.65 | 0.04 ± 0.01 | 0.02 ± 0.02 | 0.75 ± 0.96 | 0.02 ± 0.02 |
| INSE0 | 17.75 ± 3.66 | 0.05 ± 0.02 | 0.03 ± 0.01 | 0.50 ± 0.58 | 0.02 ± 0.03 |
| INSE350 | 13.88 ± 4.89 | 0.03 ± 0.01 | 0.04 ± 0.01 | 0.25 ± 0.50 | 0.01 ± 0.01 |
| INSE550 | 15.38 ± 5.11 | 0.03 ± 0.03 | 0.02 ± 0.01 | 0.75 ± 0.50 | 0.02 ± 0.02 |
| INSE850 | 16.50 ± 2.80 | 0.03 ± 0.01 | 0.05 ± 0.03 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| ISE0 | 15.25 ± 3.38 | 0.03 ± 0.01 | 0.04 ± 0.01 | 0.50 ± 0.58 | 0.03 ± 0.04 |
| ISE350 | 14.38 ± 2.29 | 0.02 ± 0.01 | 0.04 ± 0.02 | 0.75 ± 0.50 | 0.02 ± 0.02 |
| ISE550 | 15.25 ± 3.28 | 0.04 ± 0.02 | 0.03 ± 0.02 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| ISETH850 | 12.50 ± 2.42 | 0.03 ± 0.01 | 0.05 ± 0.01 | 0.00 ± 0.00 | 0.00 ± 0.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irankhah, S.; Sillo, F.; Nerva, L.; Ganjeali, A.; Balestrini, R.; Chitarra, W. Combined Effects of Water Deficit, Exogenous Ethylene Application and Root Symbioses on Trigonelline and ABA Accumulation in Fenugreek. Appl. Sci. 2020, 10, 2338. https://doi.org/10.3390/app10072338
Irankhah S, Sillo F, Nerva L, Ganjeali A, Balestrini R, Chitarra W. Combined Effects of Water Deficit, Exogenous Ethylene Application and Root Symbioses on Trigonelline and ABA Accumulation in Fenugreek. Applied Sciences. 2020; 10(7):2338. https://doi.org/10.3390/app10072338
Chicago/Turabian StyleIrankhah, Simin, Fabiano Sillo, Luca Nerva, Ali Ganjeali, Raffaella Balestrini, and Walter Chitarra. 2020. "Combined Effects of Water Deficit, Exogenous Ethylene Application and Root Symbioses on Trigonelline and ABA Accumulation in Fenugreek" Applied Sciences 10, no. 7: 2338. https://doi.org/10.3390/app10072338
APA StyleIrankhah, S., Sillo, F., Nerva, L., Ganjeali, A., Balestrini, R., & Chitarra, W. (2020). Combined Effects of Water Deficit, Exogenous Ethylene Application and Root Symbioses on Trigonelline and ABA Accumulation in Fenugreek. Applied Sciences, 10(7), 2338. https://doi.org/10.3390/app10072338