Plasma-Treated Flammulina velutipes-Derived Extract Showed Anticancer Potential in Human Breast Cancer Cells


 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Characterization of Gas Plasma
2.2. H2O2 Level, NOx Level, Temperature, and pH Measurement
2.3. Mushroom Cultivation and Plasma Application
2.4. Collection of Material
2.5. Mushroom Extract Preparation
2.6. Characterization of Mushroom Extract
2.6.1. FTIR and UV-Vis Analysis
2.6.2. Phenol Content
2.6.3. Flavonoid Content
2.7. DPPH Activity Assay
2.8. Cell Culture
2.9. Cytotoxicity of Extracts
2.10. Flow Cytometry Analysis
2.11. Primer Design and Quantitative Real-Time PCR
2.12. Confocal Microscopy for DNA Damage Assessment
2.13. Statistical Analysis
3. Results
3.1. Detection of RONS in Medium Exposed to Plasma
3.2. Phytochemical and Antioxidant Activities of Extracts Derived from Plasma-Exposed Mushrooms
3.3. Analysis of Active Constituents by FTIR and UV-Vis
3.4. Antiproliferative Activity of Plasma-Treated Mushroom-Derived Extracts in Breast Cancer Cells
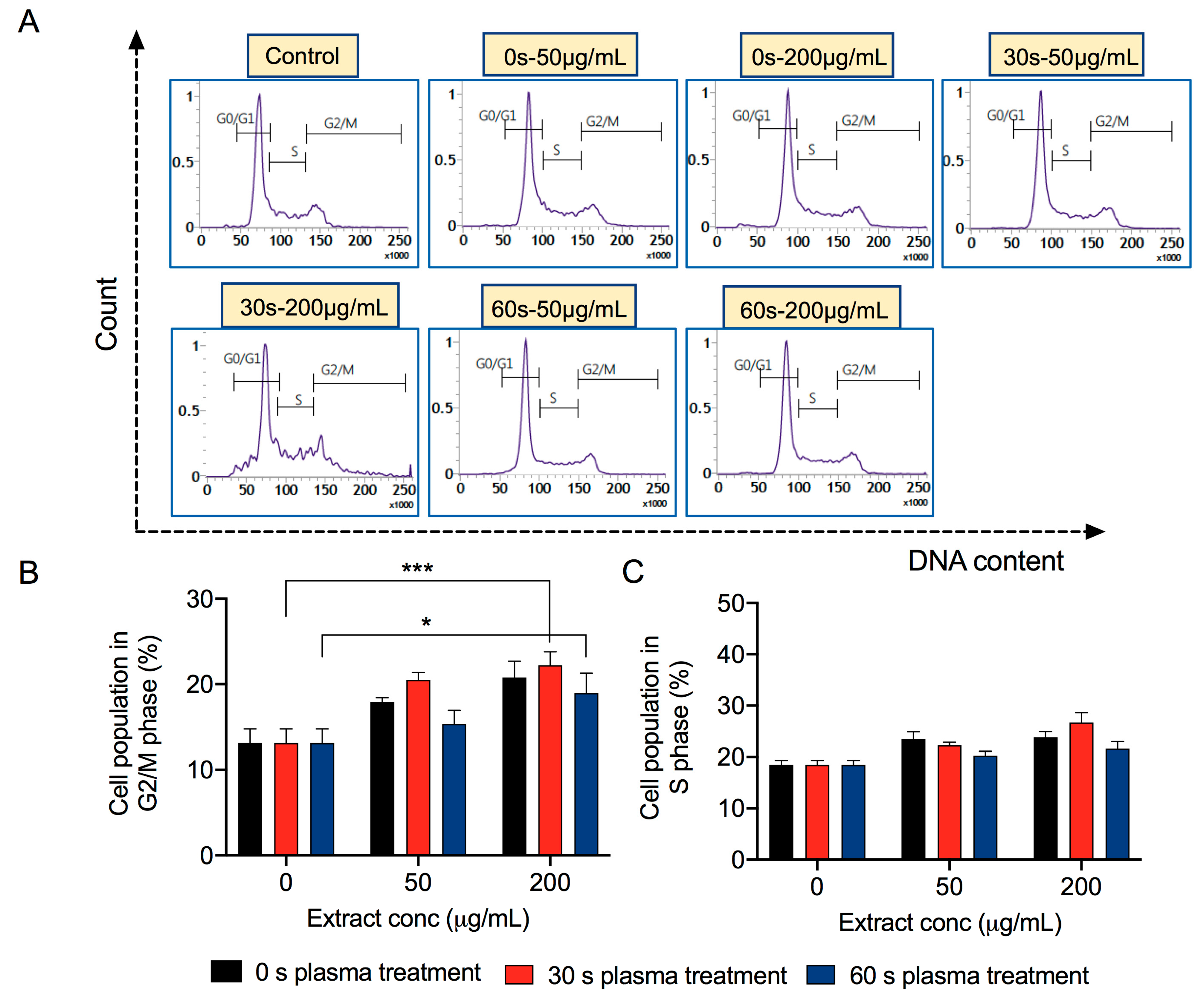
3.5. Extracts Derived from Plasma-Treated Mushrooms Induce Cancer Cell Death due to Cell Cycle Arrest
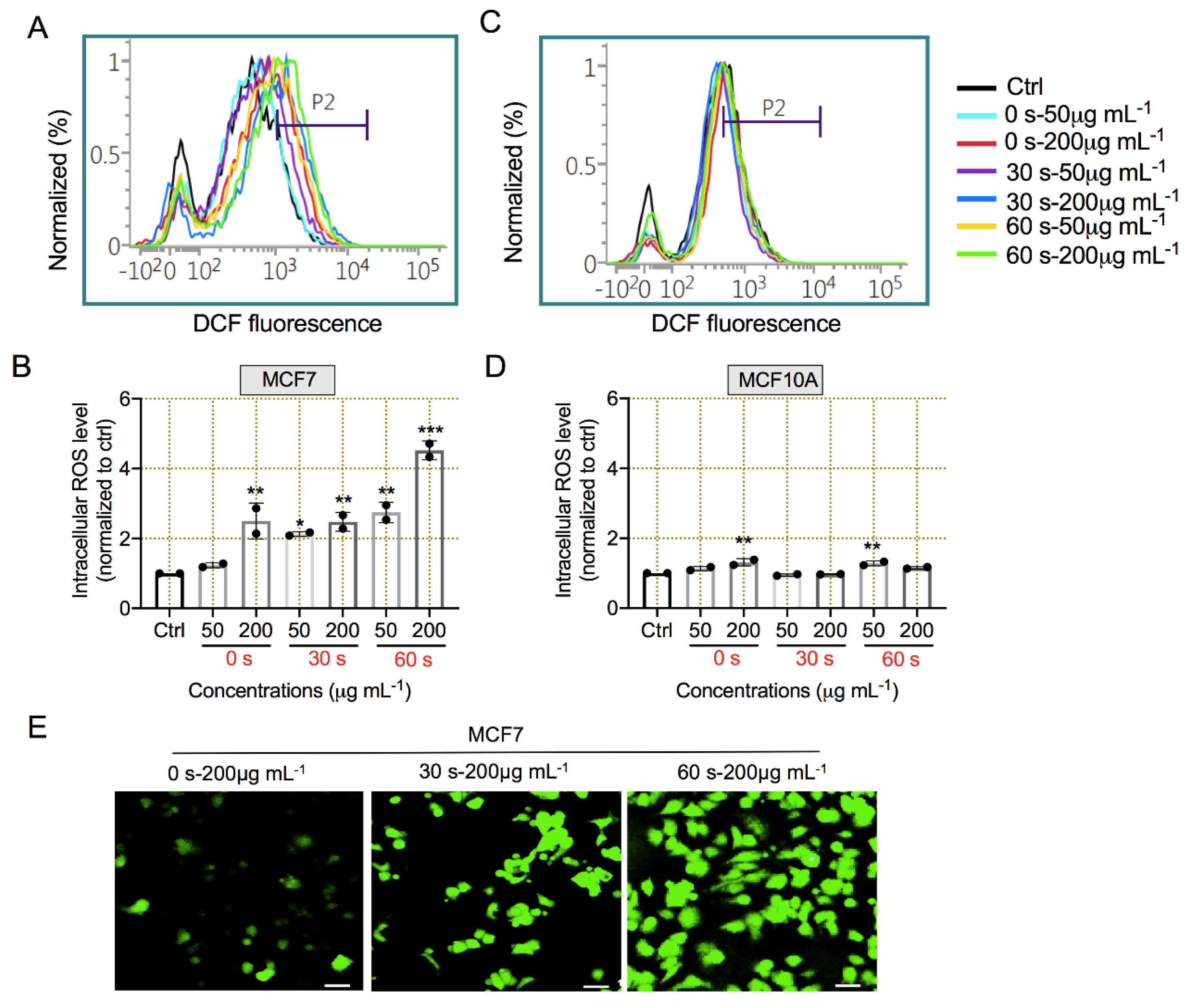
3.6. Plasma-Exposed Mushroom-Derived Extracts Induced ROS Generation for Apoptosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mitra, S.; Dash, R. Natural Products for the Management and Prevention of Breast Cancer. Evid. Based Complement. Altern. Med. 2018, 2018, 8324696. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Luo, X.; Xu, X.; Wei, W.; Yu, M.; Jiang, N.; Ye, L.; Yang, Z.; Fei, X. Antioxidant and immunomodulatory activities of a polysaccharide from Flammulina velutipes. J. Tradit. Chin. Med. 2014, 34, 733–740. [Google Scholar] [CrossRef]
- Ukaegbu, C.I.; Shah, S.R.; Hazrulrizawati, A.H.; Alara, O.R. Acetone extract of Flammulina velutipes caps: A promising source of antioxidant and anticancer agents. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 675–682. [Google Scholar] [CrossRef]
- Scholtz, V.; Šerá, B.; Khun, J.; Šerý, M.; Julák, J. Effects of Nonthermal Plasma on Wheat Grains and Products. J. Food Qual. 2019, 2019, 7917825. [Google Scholar] [CrossRef]
- Dobrin, D.; Magureanu, M.; Mandache, N.B.; Ionita, M.-D. The effect of non-thermal plasma treatment on wheat germination and early growth. Innov. Food Sci. Emerg. 2015, 29, 255–260. [Google Scholar] [CrossRef]
- Bourke, P.; Ziuzina, D.; Boehm, D.; Cullen, P.J.; Keener, K. The potential of cold plasma for safe and sustainable food production. Trends Biotechnol. 2018, 36, 615–626. [Google Scholar] [CrossRef]
- Neyts, E.C. Plasma-Surface Interactions in Plasma Catalysis. Plasma Chem. 2016, 36, 185–212. [Google Scholar] [CrossRef]
- Hirst, A.M.; Frame, F.M.; Arya, M.; Maitland, N.J.; O’Connell, D. Low temperature plasmas as emerging cancer therapeutics: The state of play and thoughts for the future. Tumor Biol. 2016, 37, 7021–7031. [Google Scholar] [CrossRef]
- Laroussi, M. Plasma Medicine: A Brief Introduction. Plasma 2018, 1, 5. [Google Scholar] [CrossRef]
- Dong, Y.R.; Cheng, S.J.; Qi, G.H.; Yang, Z.P.; Yin, S.Y.; Chen, G.T. Antimicrobial and antioxidant activities of Flammulina velutipes polysaccharides and polysaccharide-iron(III) complex [corrected]. Carbohydr. Polym. 2017, 161, 26–32. [Google Scholar] [CrossRef]
- Wu, D.-m.; Duan, W.-q.; Liu, Y.; Cen, Y. Anti-inflammatory effect of the polysaccharides of Golden needle mushroom in burned rats. Int. J. Biol. Macromol. 2010, 46, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-H.; Hsieh, K.-Y.; Yeh, C.-H.; Tu, Y.-P.; Sheu, F. Oral administration of an Enoki mushroom protein FVE activates innate and adaptive immunity and induces anti-tumor activity against murine hepatocellular carcinoma. Int. Immunopharmacol. 2010, 10, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.H.; Leonard, J. In vitro effects on proliferation, apoptosis and colony inhibition in ER-dependent and ER-independent human breast cancer cells by selected mushroom species. Oncol. Rep. 2006, 15, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Leung, M.Y.; Fung, K.P.; Choy, Y.M. The isolation and characterization of an immunomodulatory and anti-tumor polysaccharide preparation from Flammulina velutipes. Immunopharmacology 1997, 35, 255–263. [Google Scholar] [CrossRef]
- Yi, C.; Zhong, H.; Tong, S.; Cao, X.; Firempong, C.K.; Liu, H.; Fu, M.; Yang, Y.; Feng, Y.; Zhang, H.; et al. Enhanced oral bioavailability of a sterol-loaded microemulsion formulation of Flammulina velutipes, a potential antitumor drug. Int. J. Nanomed. 2012, 7, 5067–5078. [Google Scholar]
- Chang, Y.C.; Hsiao, Y.M.; Wu, M.F.; Ou, C.C.; Lin, Y.W.; Lue, K.H.; Ko, J.L. Interruption of lung cancer cell migration and proliferation by fungal immunomodulatory protein FIP-fve from Flammulina velutipes. J. Agric. Food Chem. 2013, 61, 12044–12052. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144. [Google Scholar]
- Chandra, S.; Khan, S.; Avula, B.; Lata, H.; Yang, M.H.; ElSohly, M.A.; Khan, I.A. Assessment of Total Phenolic and Flavonoid Content, Antioxidant Properties, and Yield of Aeroponically and Conventionally Grown Leafy Vegetables and Fruit Crops: A Comparative Study. Evid. Based Complement. Altern. Med. 2014, 2014, 253875. [Google Scholar] [CrossRef]
- Chang, C.-C.; Yang, M.-H.; Wen, H.-M.; Chern, J.-C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.-E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Adhikari, M.; Adhikari, B.; Kaushik, N.; Lee, S.-J.; Kaushik, N.K.; Choi, E.H. Melanoma Growth Analysis in Blood Serum and Tissue Using Xenograft Model with Response to Cold Atmospheric Plasma Activated Medium. Appl. Sci. 2019, 9, 4227. [Google Scholar] [CrossRef]
- Baba, S.A.; Malik, S.A. Determination of total phenolic and flavonoid content, antimicrobial and antioxidant activity of a root extract of Arisaema jacquemontii Blume. J. Taibah Univ. Sci. 2015, 9, 449–454. [Google Scholar] [CrossRef]
- Shimoi, K.; Masuda, S.; Shen, B.; Furugori, M.; Kinae, N. Radioprotective effects of antioxidative plant flavonoids in mice. Mutat. Res. Fund. Mol. M 1996, 350, 153–161. [Google Scholar] [CrossRef]
- Alam, M.N.; Bristi, N.J.; Rafiquzzaman, M. Review on in vivo and in vitro methods evaluation of antioxidant activity. Saudi Pharm. J. 2013, 21, 143–152. [Google Scholar] [CrossRef]
- Engida, A.M.; Faika, S.; Nguyen-Thi, B.T.; Ju, Y.H. Analysis of major antioxidants from extracts of Myrmecodia pendans by UV/visible spectrophotometer, liquid chromatography/tandem mass spectrometry, and high-performance liquid chromatography/UV techniques. J. Food Drug Anal. 2015, 23, 303–309. [Google Scholar] [CrossRef]
- Zhao, M.; Yang, B.; Wang, J.; Li, B.; Jiang, Y. Identification of the major flavonoids from pericarp tissues of lychee fruit in relation to their antioxidant activities. Food Chem. 2006, 98, 539–544. [Google Scholar] [CrossRef]
- Lin, H.-W.; Hsieh, M.-J.; Yeh, C.-B.; Hsueh, K.-C.; Hsieh, Y.-H.; Yang, S.-F. Coronarin D induces apoptotic cell death through the JNK pathway in human hepatocellular carcinoma. Environ. Toxicol. 2018, 33, 946–954. [Google Scholar] [CrossRef]
- Ryu, S.; Lim, W.; Bazer, F.W.; Song, G. Chrysin induces death of prostate cancer cells by inducing ROS and ER stress. J. Cell. Physiol. 2017, 232, 3786–3797. [Google Scholar] [CrossRef]
- Zhang, S.; He, Y.; Tong, Q.; Chen, Q.; Wu, X.; Huang, W. Deltonin induces apoptosis in MDA-MB-231 human breast cancer cells via reactive oxygen species-mediated mitochondrial dysfunction and ERK/AKT signaling pathways. Mol. Med. Rep. 2013, 7, 1038–1044. [Google Scholar] [CrossRef]
- Leibowitz, B.; Yu, J. Mitochondrial signaling in cell death via the Bcl-2 family. Cancer Biol. Ther. 2010, 9, 417–422. [Google Scholar] [CrossRef]
- Youle, R.J.; Strasser, A. The BCL-2 protein family: Opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Goyal, A. Recent developments in mushrooms as anti-cancer therapeutics: A review. 3 Biotech 2012, 2, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ayeka, P.A. Potential of Mushroom Compounds as Immunomodulators in Cancer Immunotherapy: A Review. Evid. Based Complement. Altern. Med. 2018, 2018, 7271509. [Google Scholar] [CrossRef] [PubMed]
- Blagodatski, A.; Yatsunskaya, M.; Mikhailova, V.; Tiasto, V.; Kagansky, A.; Katanaev, V.L. Medicinal mushrooms as an attractive new source of natural compounds for future cancer therapy. Oncotarget 2018, 9, 29259–29274. [Google Scholar] [CrossRef]
- Pérez-Pizá, M.C.; Prevosto, L.; Grijalba, P.E.; Zilli, C.G.; Cejas, E.; Mancinelli, B.; Balestrasse, K.B. Improvement of growth and yield of soybean plants through the application of non-thermal plasmas to seeds with different health status. Heliyon 2019, 5, e01495. [Google Scholar] [CrossRef]
- Randeniya, L.K.; de Groot, G.J.J.B. Non-Thermal Plasma Treatment of Agricultural Seeds for Stimulation of Germination, Removal of Surface Contamination and Other Benefits: A Review. Plasma Process. Polym. 2015, 12, 608–623. [Google Scholar] [CrossRef]
- Dampier, K.; Hudson, E.A.; Howells, L.M.; Manson, M.M.; Walker, R.A.; Gescher, A. Differences between human breast cell lines in susceptibility towards growth inhibition by genistein. Br. J. Cancer 2001, 85, 618–624. [Google Scholar] [CrossRef]
- Hu, H.; Zhang, Z.; Lei, Z.; Yang, Y.; Sugiura, N. Comparative study of antioxidant activity and antiproliferative effect of hot water and ethanol extracts from the mushroom Inonotus obliquus. J. Biosci. Bioeng. 2009, 107, 42–48. [Google Scholar] [CrossRef]
- Jeong, J.W.; Jin, C.Y.; Park, C.; Hong, S.H.; Kim, G.Y.; Jeong, Y.K.; Lee, J.D.; Yoo, Y.H.; Choi, Y.H. Induction of apoptosis by cordycepin via reactive oxygen species generation in human leukemia cells. Toxicol. In Vitro 2011, 25, 817–824. [Google Scholar] [CrossRef]
- Sznarkowska, A.; Kostecka, A.; Meller, K.; Bielawski, K.P. Inhibition of cancer antioxidant defense by natural compounds. Oncotarget 2017, 8, 15996. [Google Scholar] [CrossRef]
- Stepanic, V.; Gasparovic, A.C.; Troselj, K.G.; Amic, D.; Zarkovic, N. Selected attributes of polyphenols in targeting oxidative stress in cancer. Curr. Top. Med. Chem. 2015, 15, 496–509. [Google Scholar] [CrossRef] [PubMed]
- Chian, S.; Li, Y.-Y.; Wang, X.-J.; Tang, X.-W. Luteolin sensitizes two oxaliplatin-resistant colorectal cancer cell lines to chemotherapeutic drugs via inhibition of the Nrf2 pathway. Asian Pac. J. Cancer Prev. 2014, 15, 2911–2916. [Google Scholar] [CrossRef] [PubMed]
- Chien, S.Y.; Wu, Y.C.; Chung, J.G.; Yang, J.S.; Lu, H.F.; Tsou, M.F.; Wood, W.G.; Kuo, S.J.; Chen, D.R. Quercetin-induced apoptosis acts through mitochondrial- and caspase-3-dependent pathways in human breast cancer MDA-MB-231 cells. Hum. Exp. Toxicol. 2009, 28, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.D.; Elias, R.J. The antioxidant and pro-oxidant activities of green tea polyphenols: A role in cancer prevention. Arch. Biochem. Biophys. 2010, 501, 65–72. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, G.; Tang, X.; Chen, H. Polysaccharide lentinan extracted from the stipe of Lentinus edodes mushroom exerts anticancer activities through the transcriptional regulation of cell cycle progression and metastatic markers in human colon cancer cells. Faseb. J. 2017, 31, lb391. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence (5′-3′) | Size (bp) |
|---|---|---|
| ACTIN-forward | GGC ATC CTC ACC CTG AAG TA | 82 |
| ACTIN-reverse | AGG TGT GGT GCC AGA TTT TC | |
| BAX-forward | AAG AAG CTG AGC GAG TGT CTC | 186 |
| BAX-reverse | GCT GGC AAA GTA GAA AAG GGC | |
| BCL2-forward | TTC TTT GAG TTC GGT GGG GTC | 208 |
| BCL2-reverse | TCT TCA GAG ACA GCC AGG AGA | |
| BCL-XL-forward | TCA GTG AGT GAG CAG GTG TT | 170 |
| BCL-XL-reverse | GGC CTC AGT CCT GTT CTC TT | |
| TP53-forward | GCC CCT CCT CAG CAT CTT ATC | 246 |
| TP53-reverse | AAA GCT GTT CCG TCC CAG TAG | |
| H2AX-forward | CAA CAA GAA GAC GCG AAT CA | 223 |
| H2AX-reverse | CGG GCC CTC TTA GTA CTC CT | |
| Casp3-forward | ATG TCG ATG CAG CAA ACC TC | 173 |
| Casp3-reverse | TCC TTC TTC ACC ATG GCT CA | |
| Casp7-forward | AGA AGC TGT TAC CAC ACC CA | 185 |
| Casp7-reverse | ACT CCA TCT CAG TCA GTG GC | |
| Casp8-forward | CCC AAA TCA ACA AGA GCC TGC | 219 |
| Casp8-reverse | TCA GAC AGT ATC CCC GAG GTT | |
| CytC-forward | CAA CAC CTC TCA CAT CTT ACA GA | 185 |
| CytC-reverse | GGC AAA AGC ACC ATT CCC AA | |
| PUMA-forward | CCC ACC ACC ATC TCA GGA AA | 185 |
| PUMA-reverse | GTG GTC ACG TTT GGC TCA TT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mitra, S.; Bhartiya, P.; Kaushik, N.; Nhat Nguyen, L.; Wahab, R.; Bekeschus, S.; Choi, E.H.; Kaushik, N.K. Plasma-Treated Flammulina velutipes-Derived Extract Showed Anticancer Potential in Human Breast Cancer Cells. Appl. Sci. 2020, 10, 8395. https://doi.org/10.3390/app10238395
Mitra S, Bhartiya P, Kaushik N, Nhat Nguyen L, Wahab R, Bekeschus S, Choi EH, Kaushik NK. Plasma-Treated Flammulina velutipes-Derived Extract Showed Anticancer Potential in Human Breast Cancer Cells. Applied Sciences. 2020; 10(23):8395. https://doi.org/10.3390/app10238395
Chicago/Turabian StyleMitra, Sarmistha, Pradeep Bhartiya, Neha Kaushik, Linh Nhat Nguyen, Rizwan Wahab, Sander Bekeschus, Eun Ha Choi, and Nagendra Kumar Kaushik. 2020. "Plasma-Treated Flammulina velutipes-Derived Extract Showed Anticancer Potential in Human Breast Cancer Cells" Applied Sciences 10, no. 23: 8395. https://doi.org/10.3390/app10238395
APA StyleMitra, S., Bhartiya, P., Kaushik, N., Nhat Nguyen, L., Wahab, R., Bekeschus, S., Choi, E. H., & Kaushik, N. K. (2020). Plasma-Treated Flammulina velutipes-Derived Extract Showed Anticancer Potential in Human Breast Cancer Cells. Applied Sciences, 10(23), 8395. https://doi.org/10.3390/app10238395