A High Manganese-Tolerant Pseudomonas sp. Strain Isolated from Metallurgical Waste Heap Can Be a Tool for Enhancing Manganese Removal from Contaminated Soil



Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Characterization
2.2. Isolation of Bacteria
2.3. Manganese Determination Using the Formaldoxime Method
2.4. Identification of Selected Strain
2.5. Determination of Minimal Inhibitory Concentration (MIC) of Selected Strains
2.6. Whole Cell Protein Extraction and Profiling by SDS/PAGE
2.7. Effects of Soil Concentration on Manganese Removal Efficiency
2.8. Bioaccumulation Experiments
2.9. Statistical Analysis
3. Results and Discussion
3.1. Selection of Manganese-Accumulating Strain
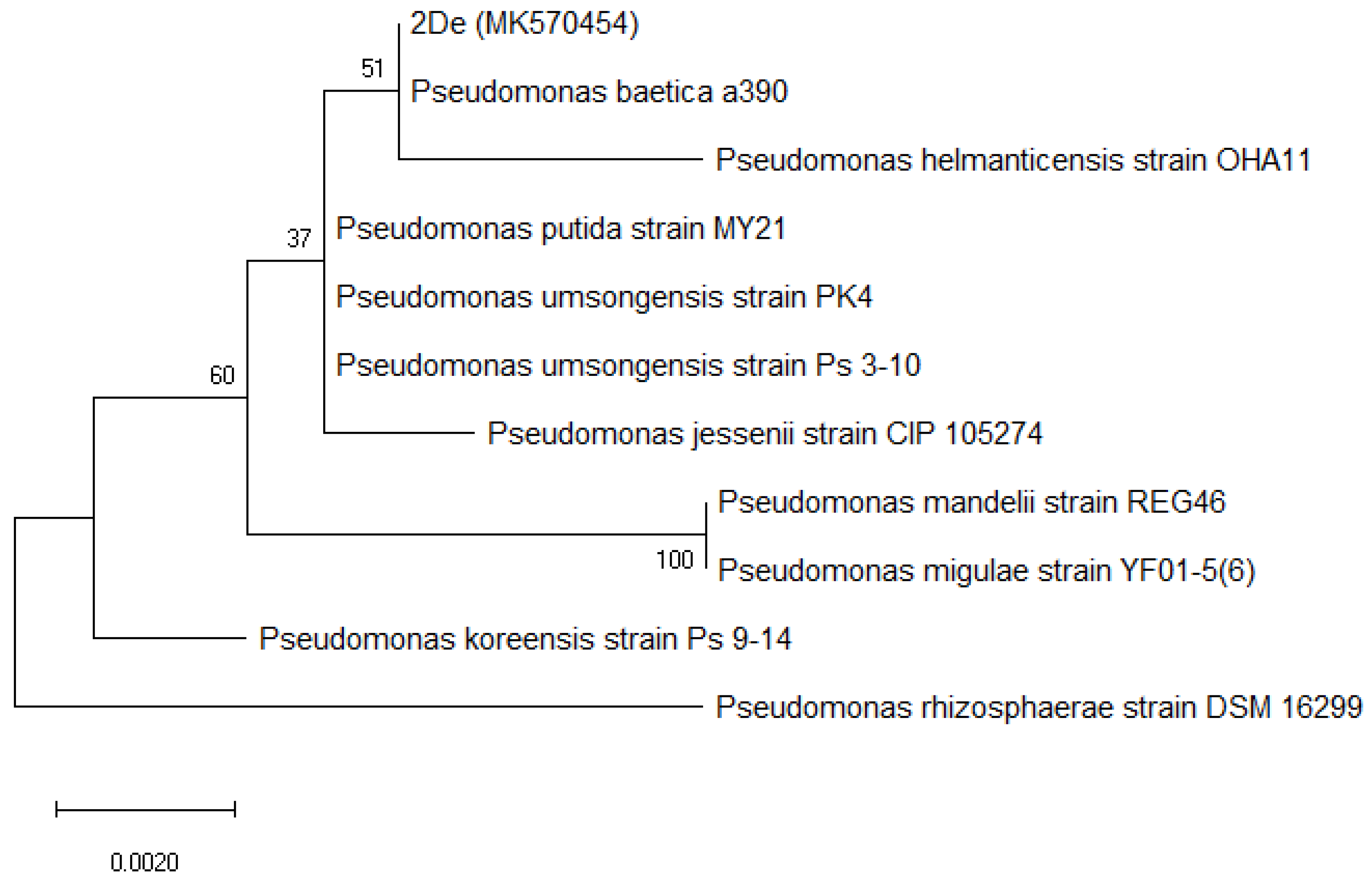
3.2. Phylogenic Analyses
3.3. Determination of MIC and Effects of Mn on Cellular Protein Expression
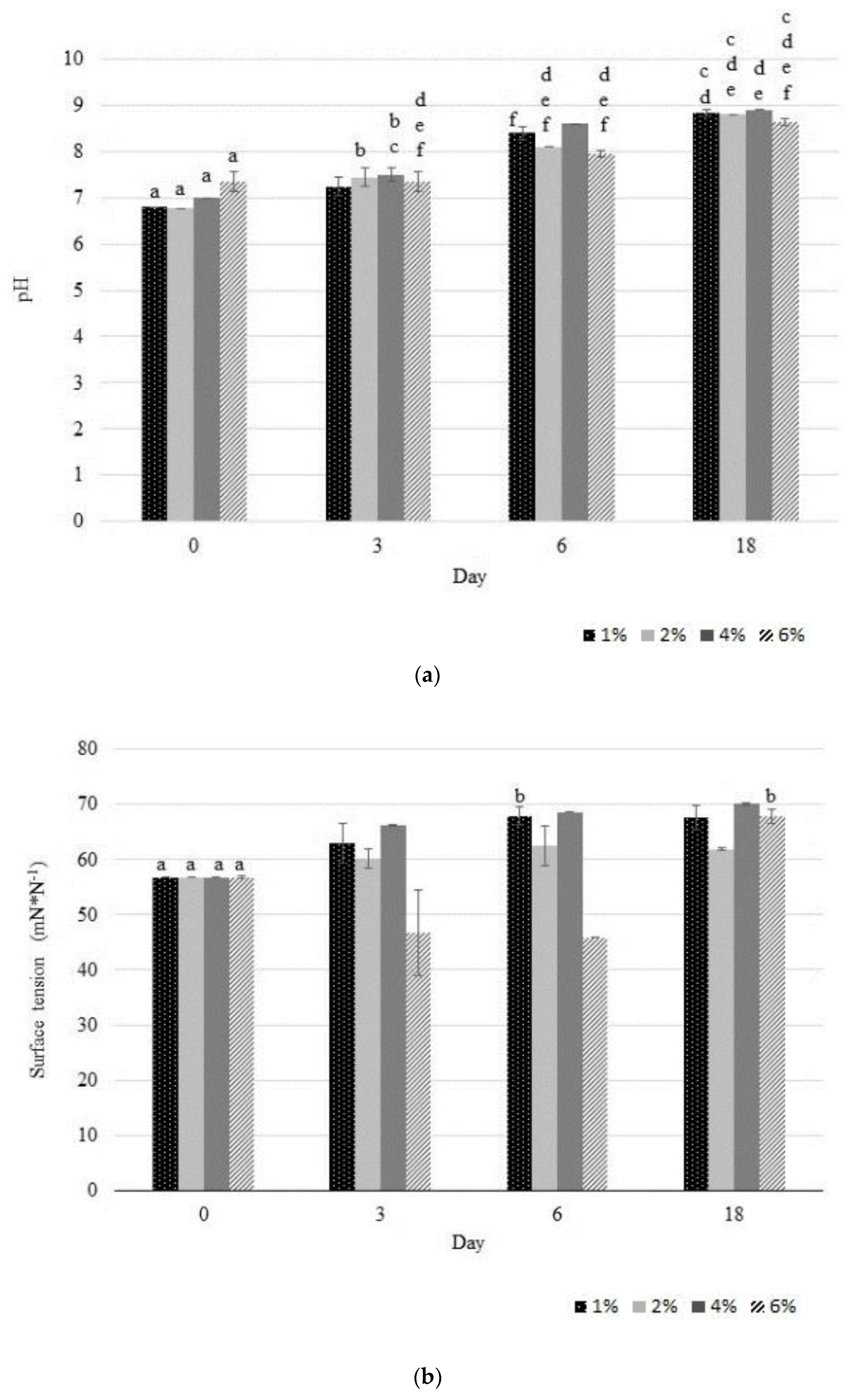
3.4. Removal Efficiency
3.5. Manganese Content in Bacterial Cells
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ghosh, S.; Mohanty, S.; Akcil, A.; Sukla, L.B.; Das, A.P. A greener approach for resource recycling: Manganese bioleaching. Chemosphere 2016, 154, 628–639. [Google Scholar] [CrossRef] [PubMed]
- Zeinert, R.; Martinez, E.; Schmitz, J.; Senn, K.; Usman, B.; Anantharaman, V.; Aravind, L.; Waters, L.S. Structure-function analysis of manganese exporter proteins across bacteria. J. Biol. Chem. 2018, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Juttukonda, L.J.; Skaar, E.P. Manganese homeostasis and utilization in pathogenic bacteria. Mol. Microbiol. 2015, 97, 216–228. [Google Scholar] [CrossRef] [PubMed]
- Pereira, F. Bioremediation of Contaminated Estuarine Sites. In Marine Pollution and Microbial Remediation; Naik, M.M., Dubey, S.K., Eds.; Springer: Singapore, 2016; ISBN 9789811010422. [Google Scholar]
- Ghosh, S.; Das, A.P. Bioleaching of manganese from mining waste residues using Acinetobacter sp. Geol. Ecol. Landsc. 2017, 1, 77–83. [Google Scholar] [CrossRef]
- Ghosh, S.; Bal, B.; Das, A.P. Enhancing Manganese Recovery from Low-Grade Ores by Using Mixed Culture of Indigenously Isolated Bacterial Strains. Geomicrobiol. J. 2018, 35, 242–246. [Google Scholar] [CrossRef]
- Venugopal, R.; Engineering, M. Solid Waste Processing for Industrial Utilization—A Few Case Studies. In Proceedings of the XI International Seminar on Mineral Processing Technology (MPT-2010), Abhilash, India, 15–17 December 2010; pp. 1075–1083. [Google Scholar]
- Kim, S.I.; Jang, Y.S.; Han, S.H.; Choi, M.J.; Go, E.H.; Cheon, Y.P.; Lee, J.S.; Lee, S.H. Effect of Manganese Exposure on the Reproductive Organs in Immature Female Rats. Dev. Reprod. 2012, 295, 295–300. [Google Scholar] [CrossRef]
- Santamaria, A.B.; Sulsky, S.I. Risk assessment of an essential element: Manganese. J. Toxicol. Environ. Health Part A Curr. Issues 2010, 73, 128–155. [Google Scholar] [CrossRef]
- Sis, H.; Chander, S. Reagents used in the flotation of phosphate ores: A critical review. Miner. Eng. 2003, 16, 577–585. [Google Scholar] [CrossRef]
- Veglio’, F.; Beolchini, F. Removal of metals by biosorption: A review. Hydrometallurgy 1997, 44, 301–316. [Google Scholar] [CrossRef]
- Behera, S.K.; Mulaba-Bafubiandi, A.F. Advances in microbial leaching processes for nickel extraction from lateritic minerals—A review. Korean J. Chem. Eng. 2015, 32, 1447–1454. [Google Scholar] [CrossRef]
- Sadeghabad, M.S.; Bahaloo-Horeh, N.; Mousavi, S.M. Using bacterial culture supernatant for extraction of manganese and zinc from waste alkaline button-cell batteries. Hydrometallurgy 2019, 188, 81–91. [Google Scholar] [CrossRef]
- Sheng, X.F.; Xia, J.J.; Jiang, C.Y.; He, L.Y.; Qian, M. Characterization of heavy metal-resistant endophytic bacteria from rape (Brassica napus) roots and their potential in promoting the growth and lead accumulation of rape. Environ. Pollut. 2008, 156, 1164–1170. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.C.; Cheng, C.L.; Han, Y.L.; Chen, B.Y.; Chang, J.S. Recovery of high-value metals from geothermal sites by biosorption and bioaccumulation. Bioresour. Technol. 2014, 160, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Dhankhar, R.; Hooda, A. Fungal biosorption-an alternative to meet the challenges of heavy metal pollution in aqueous solutions. Environ. Technol. 2011, 32, 467–491. [Google Scholar] [CrossRef]
- Yoo, K.; Shin, S.M.; Yang, D.H.; Sohn, J.S. Biological treatment of wastewater produced during recycling of spent lithium primary battery. Miner. Eng. 2010, 23, 219–224. [Google Scholar] [CrossRef]
- Rosch, J.W.; Gao, G.; Ridout, G.; Wang, Y.D.; Tuomanen, E.I. Role of the manganese efflux system mntE for signalling and pathogenesis in Streptococcus pneumoniae. Mol. Microbiol. 2009, 72, 12–25. [Google Scholar] [CrossRef]
- Veyrier, F.J.; Boneca, I.G.; Cellier, M.F.; Taha, M.K. A novel metal transporter mediating manganese export (mntx) regulates the mn to fe intracellular ratio and Neisseria meningitidis virulence. PLoS Pathog. 2011, 7. [Google Scholar] [CrossRef]
- Waters, L.S.; Sandoval, M.; Storz, G. The Escherichia coli MntR miniregulon includes genes encoding a small protein and an efflux pump required for manganese homeostasis. J. Bacteriol. 2011, 193, 5887–5897. [Google Scholar] [CrossRef]
- Yuan, W.; Cheng, J.; Huang, H.; Xiong, S.; Gao, J.; Zhang, J.; Feng, S. Optimization of cadmium biosorption by Shewanella putrefaciens using a Box-Behnken design. Ecotoxicol. Environ. Saf. 2019, 175, 138–147. [Google Scholar] [CrossRef]
- Endo, R.; Aoyagi, H. Adsorption preference for divalent metal ions by Lactobacillus casei JCM1134. Appl. Microbiol. Biotechnol. 2018, 102, 6155–6162. [Google Scholar] [CrossRef]
- Kluczka, J.; ZoŁotajkin, M.; Ciba, J. Speciation of aluminium in the water and bottom sediment of fish-breeding ponds. Arch. Environ. Prot. 2012, 38, 83–96. [Google Scholar] [CrossRef]
- Smoliński, A.; Zołotajkin, M.; Ciba, J.; Dydo, P.; Kluczka, J. PLS-EP algorithm to predict aluminum content in soils of Beskid Mountains region. Chemosphere 2009, 76, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Kalembkiewicz, J.; Sitarz-Palczak, E.; Zapała, L. A study of the chemical forms or species of manganese found in coal fly ash and soil. Microchem. J. 2008, 90, 37–43. [Google Scholar] [CrossRef]
- Volkov, A.I.; Ossipov, K.B.; Seregin, A.N.; Zhdanov, P.A.; Seregina, I.F.; Bolshov, M.A. Determination of degree of oxidation and forms of manganese compounds in the Ulu-Telyak oxidized ore. Inorg. Mater. 2015, 51, 1394–1403. [Google Scholar] [CrossRef]
- Pacwa-Pł>ociniczak, M.; Płaza, G.A.; Paliwoda, A.; Piotrowska-Seget, Z. Characterization of hydrocarbon-degrading and biosurfactant-producing Pseudomonas sp. P-1 strain as a potential tool for bioremediation of petroleum-contaminated soil. Env. Sci. Pollut. Res. 2014, 9385–9395. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Lim, J.M.; Kwon, S.J.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef]
- Płociniczak, T.; Chodór, M.; Pacwa-Płociniczak, M.; Piotrowska-Seget, Z. Metal-tolerant endophytic bacteria associated with Silene vulgaris support the Cd and Zn phytoextraction in non-host plants. Chemosphere 2019, 219, 250–260. [Google Scholar] [CrossRef]
- Furmanczyk, E.M.; Kaminski, M.A.; Lipinski, L.; Dziembowski, A.; Sobczak, A. Pseudomonas laurylsulfatovorans sp. nov., sodium dodecyl sulfate degrading bacteria, isolated from the peaty soil of a wastewater treatment plant. Syst. Appl. Microbiol. 2018, 41, 348–354. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Tang, M.; Zhu, W.; Yu, J.; Fu, Y.; Fan, L.; Wei, G. Isolation and characterization of the heavy metal resistant bacteria CCNWRS33-2 isolated from root nodule of Lespedeza cuneata in gold mine tailings in China. J. Hazard. Mater. 2008, 162, 50–56. [Google Scholar] [CrossRef]
- Xuezheng, L.; Aiguo, G.; Haowen, C. Isolation and phylogenetic analysis of cultivable manganese bacteria in sediments from the Arctic Ocean. Acta Ecol. Sin. 2008, 28, 6364–6370. [Google Scholar] [CrossRef]
- Huang, H.; Zhao, Y.; Xu, Z.; Ding, Y.; Zhou, X.; Dong, M. A high Mn(II)-tolerance strain, Bacillus thuringiensis HM7, isolated from manganese ore and its biosorption characteristics. PeerJ 2020, 2020, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Wakeman, C.A.; Skaar, E.P. Metalloregulation of Gram-positive pathogen physiology. Curr. Opin. Microbiol. 2012, 15, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Estes, E.R.; Andeer, P.F.; Nordlund, D.; Wankel, S.D.; Hansel, C.M. Biogenic manganese oxides as reservoirs of organic carbon and proteins in terrestrial and marine environments. Geobiology 2017, 15, 158–172. [Google Scholar] [CrossRef]
- Robinson-Lora, M.A.; Brennan, R.A. Biosorption of manganese onto chitin and associated proteins during the treatment of mine impacted water. Chem. Eng. J. 2010, 162, 565–572. [Google Scholar] [CrossRef]
- Pereira, F.; Kerkar, S.; Krishnan, K.P. Bacterial response to dynamic metal concentrations in the surface sediments of a solar saltern (Goa, India). Environ. Monit. Assess. 2013, 3625–3636. [Google Scholar] [CrossRef]
- White, C.; Wilkinson, S.C.; Gadd, G.M. The role of microorganisms in biosorption of toxic metals and radionuclides. Int. Biodeterior. Biodegrad. 1995, 35, 17–40. [Google Scholar] [CrossRef]
- Zeng, X.; Tang, J.; Xue-duan, L.I.U.; Pei, J. Isolation, identification and characterization of cadmium-resistant Pseudomonas aeruginosa strain E 1. J. Cent. South Univ. Technol. 2009, 416–421. [Google Scholar] [CrossRef]
- Li, F.; Wang, W.; Li, C.; Zhu, R.; Ge, F.; Zheng, Y.; Tang, Y. Self-mediated pH changes in culture medium affecting biosorption and biomineralization of Cd2+ by Bacillus cereus Cd01. J. Hazard. Mater. 2018, 358, 178–186. [Google Scholar] [CrossRef]
- Limcharoensuk, T.; Sooksawat, N.; Sumarnrote, A.; Awutpet, T.; Kruatrachue, M.; Pokethitiyook, P.; Auesukaree, C. Bioaccumulation and biosorption of Cd2+ and Zn2+ by bacteria isolated from a zinc mine in Thailand. Ecotoxicol. Environ. Saf. 2015, 122, 322–330. [Google Scholar] [CrossRef]
- Calderón, M.E.; Elena, M.; Buitrón, G. Biosorption of Cd, Cr, Mn and Pb from aqueus solution by Bacillus sp. strains isolated from industrail waste activate sludge. TIP Rev.Esp.Cienc.Quím.Biol. 2016, 19, 5–14. [Google Scholar] [CrossRef]
- Pacheco, P.H.; Gil, R.A.; Cerutti, S.E.; Smichowski, P.; Martinez, L.D. Biosorption: A new rise for elemental solid phase extraction methods. Talanta 2011, 85, 2290–2300. [Google Scholar] [CrossRef] [PubMed]
- Javanbakht, V.; Alavi, S.A.; Zilouei, H. Mechanisms of heavy metal removal using microorganisms as biosorbent. Water Sci. Technol. 2014, 69, 1775–1787. [Google Scholar] [CrossRef] [PubMed]
- Boyanov, M.; Kelly, K.M.; Bunker, B.A.; Fein, J.B.; Fowle, D.A. Adsorption of cadmium to Bacillus subtilis bacterial cell walls: A pH-dependent X-ray absorption fine structure spectroscopy study. Geochim. Cosmochim. Acta 2003, 67, 3299–3311. [Google Scholar] [CrossRef]
- Anjum, F.; Shahid, M.; Akcil, A. Hydrometallurgy Biohydrometallurgy techniques of low grade ores: A review on black shale. Hydrometallurgy 2012, 117, 1–12. [Google Scholar] [CrossRef]
- Vázquez-Ortega, A.; Fein, J.B. Thermodynamic modeling of Mn (II) adsorption onto manganese oxidizing bacteria. Chem. Geol. 2017, 464, 147–154. [Google Scholar] [CrossRef]
- Das, P.; Mukherjee, S.; Sen, R. Bioresource Technology Biosurfactant of marine origin exhibiting heavy metal remediation properties. Bioresour. Technol. 2009, 100, 4887–4890. [Google Scholar] [CrossRef]
- Pacwa-Płociniczak, M.; Płaza, G.A.; Piotrowska-Seget, Z.; Cameotra, S.S. Environmental applications of biosurfactants: Recent advances. Int. J. Mol. Sci. 2011, 12, 633–654. [Google Scholar] [CrossRef]
- Reynier, N.; Blais, J.; Mercier, G. Optimization of arsenic and pentachlorophenol removal from soil using an experimental design methodology. J. Soils Sediments 2013, 13, 1189–1200. [Google Scholar] [CrossRef]
- Yang, L.; Cheng, Q.; Lin, L.; Su, W.; Luan, T. Contributions of Abiotic and Biotic Processes to the Aerobic Removal of Phenolic Endocrine-Disrupting Chemicals in a Simulated Estuarine Aquatic Environment. Environ. Sci. Technol. 2016, 50, 4324–4334. [Google Scholar] [CrossRef]
- Singh, A.K.; Cameotra, S.S. Efficiency of lipopeptide biosurfactants in removal of petroleum hydrocarbons and heavy metals from contaminated soil. Environ. Sci. Pollut. Res. 2013, 20, 7367–7376. [Google Scholar] [CrossRef]
- Dahrazma, B.; Mulligan, C.N. Investigation of the removal of heavy metals from sediments using rhamnolipid in a continuous flow configuration. Chemosphere 2007, 69, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Aşçi, Y.; Nurbaş, M.; Saǧ Açikel, Y. A comparative study for the sorption of Cd(II) by K-feldspar and sepiolite as soil components, and the recovery of Cd(II) using rhamnolipid biosurfactant. J. Environ. Manage. 2008, 88, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Juwarkar, A.A.; Nair, A.; Dubey, K.V.; Singh, S.K.; Devotta, S. Biosurfactant technology for remediation of cadmium and lead contaminated soils. Chemosphere 2007, 68, 1996–2002. [Google Scholar] [CrossRef] [PubMed]
- Keren, N.; Kidd, M.J.; Penner-Hahn, J.E.; Pakrasi, H.B. A light-dependent mechanism for massive accumulation of manganese in the photosynthetic bacterium Synechocystis sp. PCC 6803. Biochemistry 2002, 41, 15085–15092. [Google Scholar] [CrossRef] [PubMed]
- Fadel, M.; Hassanein, N.M.; Elshafei, M.M.; Mostafa, A.H.; Ahmed, M.A.; Khater, H.M. Biosorption of manganese from groundwater by biomass of Saccharomyces cerevisiae. HBRC J. 2017, 13, 106–113. [Google Scholar] [CrossRef]
- Utgikar, V.P.; Harmon, S.M.; Chaudhary, N.; Tabak, H.H.; Govind, R.; Haines, J.R. Inhibition of sulfate-reducing bacteria by metal sulfide formation in bioremediation of acid mine drainage. Environ. Toxicol. 2002, 17, 40–48. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metal | Concentration (ppm) |
|---|---|
| Pb | 1000 < c < 5000 |
| K | 1000 < c < 5000 |
| Zn | 1000 < c < 5000 |
| Si | 1000 < c < 5000 |
| Au | 100 < c < 500 |
| Sr | 500 < c < 1000 |
| S | 500 < c < 1000 |
| Bacterial Strain | Decrease in Mn Concentration (%) |
|---|---|
| Control | 1.20 ± 0.53 f |
| 2Ge | 60.12 ± 1.22 abc |
| 2De | 71.07 ± 1.22 e |
| 2Dm | 12.32 ± 2.43 d |
| 2Dl | 65.47 ± 1.22 ae |
| 2D10 | 54.93 ± 2.43 c |
| 2Gh | 62.04 ± 1.22 ab |
| 2Dj | 61.68 ± 2.43 ab |
| 2Gm | 11.40 ± 3.65 d |
| 2Gi | 57.88 ± 1.22 bc |
| 2D12 | 64.84 ± 1.22 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noszczyńska, M.; Łakomy, K.; Nowacki, K.; Piotrowska-Seget, Z. A High Manganese-Tolerant Pseudomonas sp. Strain Isolated from Metallurgical Waste Heap Can Be a Tool for Enhancing Manganese Removal from Contaminated Soil. Appl. Sci. 2020, 10, 5717. https://doi.org/10.3390/app10165717
Noszczyńska M, Łakomy K, Nowacki K, Piotrowska-Seget Z. A High Manganese-Tolerant Pseudomonas sp. Strain Isolated from Metallurgical Waste Heap Can Be a Tool for Enhancing Manganese Removal from Contaminated Soil. Applied Sciences. 2020; 10(16):5717. https://doi.org/10.3390/app10165717
Chicago/Turabian StyleNoszczyńska, Magdalena, Karolina Łakomy, Krzysztof Nowacki, and Zofia Piotrowska-Seget. 2020. "A High Manganese-Tolerant Pseudomonas sp. Strain Isolated from Metallurgical Waste Heap Can Be a Tool for Enhancing Manganese Removal from Contaminated Soil" Applied Sciences 10, no. 16: 5717. https://doi.org/10.3390/app10165717
APA StyleNoszczyńska, M., Łakomy, K., Nowacki, K., & Piotrowska-Seget, Z. (2020). A High Manganese-Tolerant Pseudomonas sp. Strain Isolated from Metallurgical Waste Heap Can Be a Tool for Enhancing Manganese Removal from Contaminated Soil. Applied Sciences, 10(16), 5717. https://doi.org/10.3390/app10165717