Retinal Drug Delivery: Rethinking Outcomes for the Efficient Replication of Retinal Behavior

 ,
,  , , ,
, , ,  , , ,
, , , 
Abstract
1. Introduction
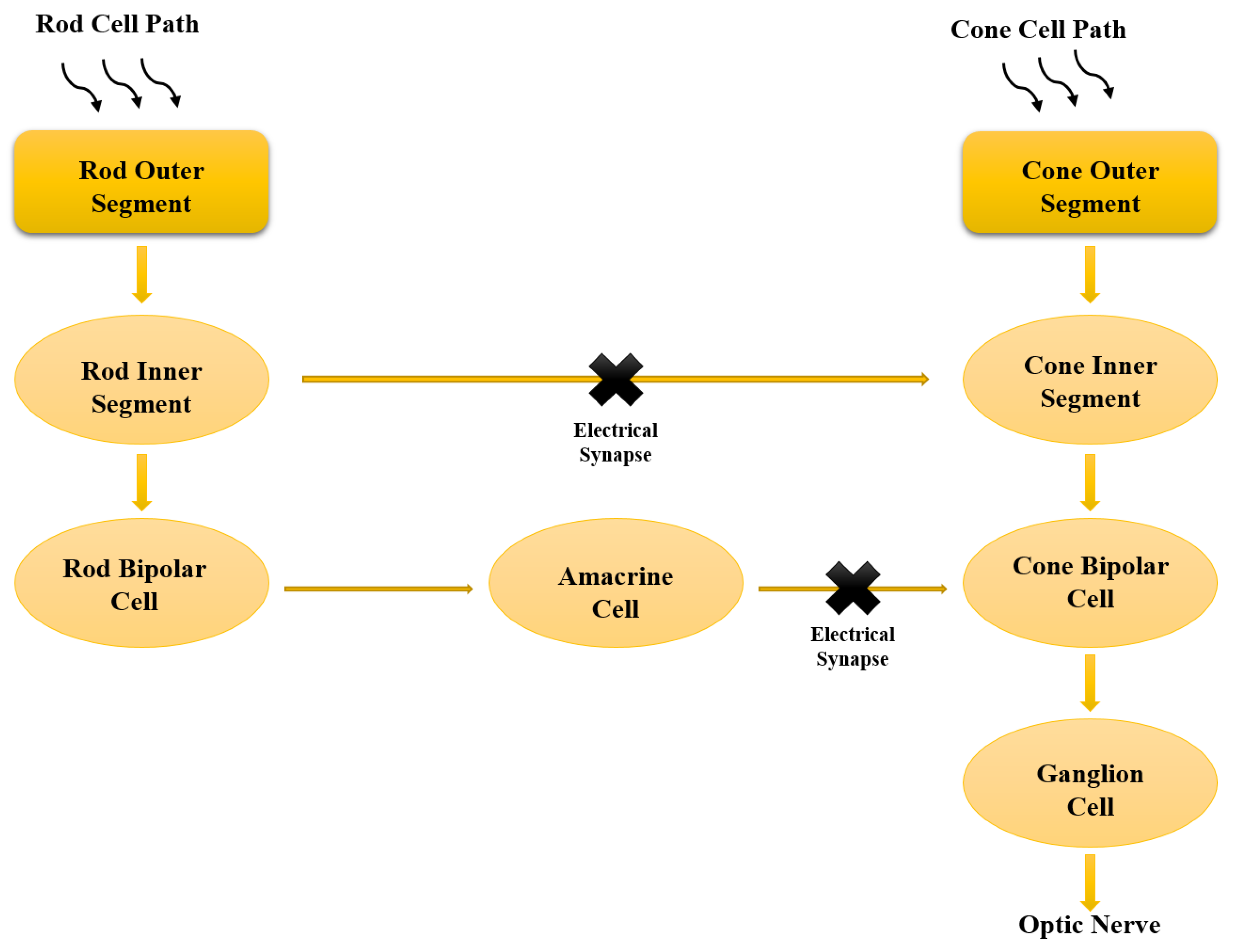
2. Biological Signalling Process of Retina
3. Computational Modeling of the Retina
4. Metadata Analysis
4.1. Virtual Pharmacokinetic 3D Model of the Human Eye by Kotha et al. Based on the FEM Using Comsol Multiphysics Software
4.2. Simulation Platform for Neural Computation in the Retina by Eshraghian et al.
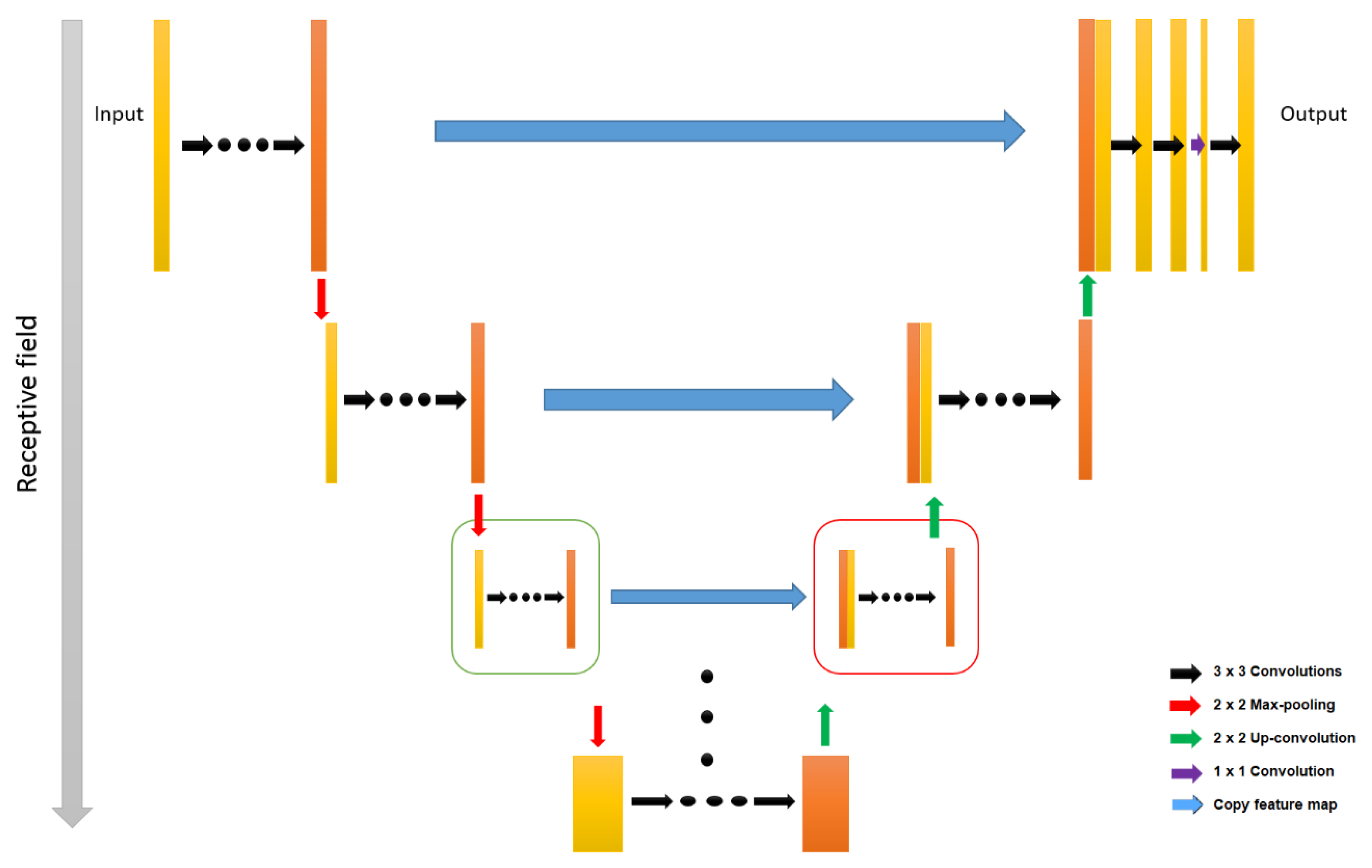
4.3. CNN Approach for Retina Segmentation in OCT Volumes
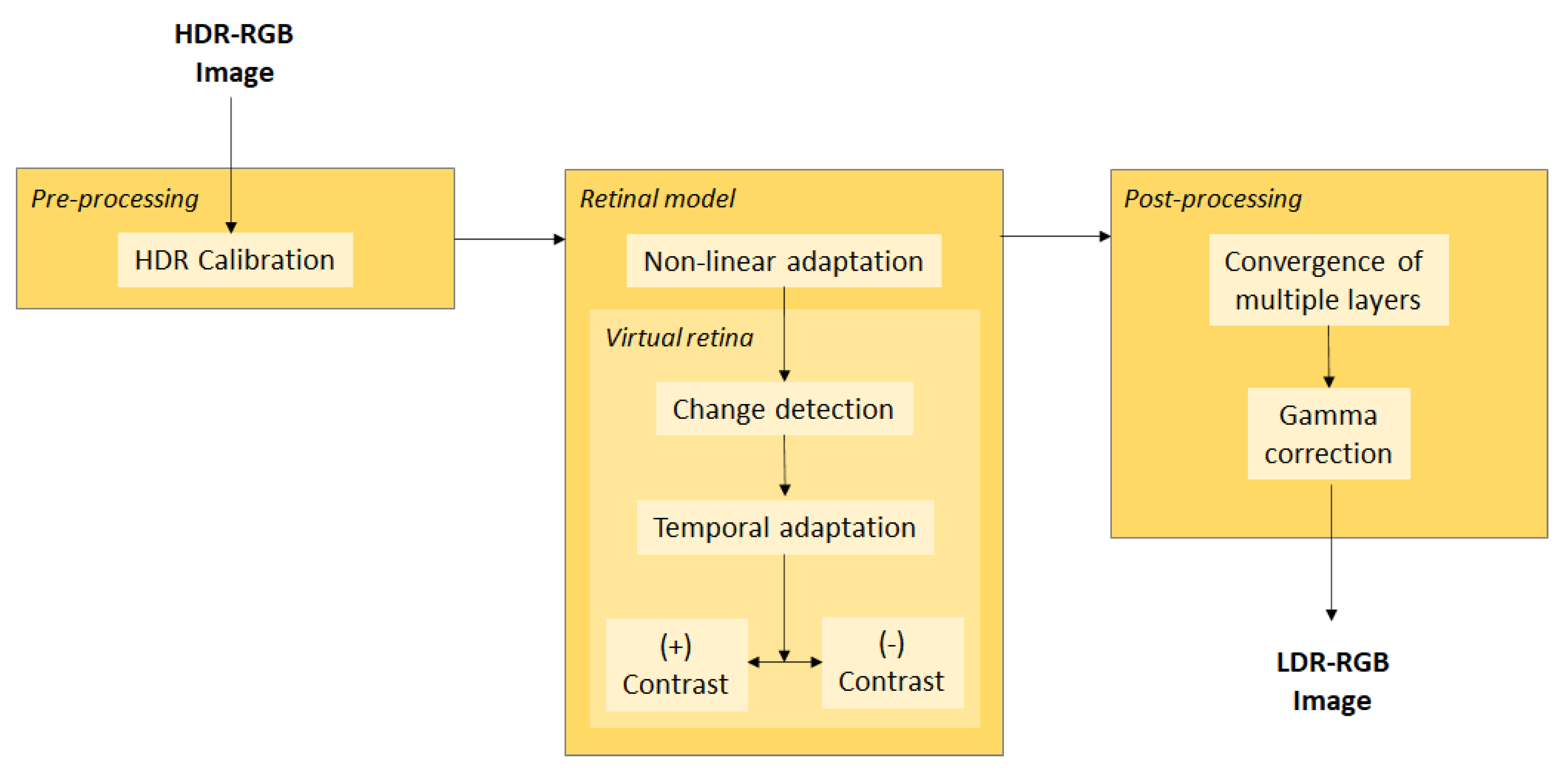
4.4. Tone Method Operators Supported by a Synergistic Model of the Retina
4.5. Adversarial Retinal Image Synthesis
4.6. Computational Model of Electrical Stimulation of the Retina
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BBM | Bruch basement membrane |
| CGC | Contrast gain control |
| CNN | Convolutional neural network |
| ECs | Choroid endothelial cells |
| FEM | Finite element method |
| HAO | Hardware adaptative optics |
| HDR | High dynamic range |
| HH | Hodgkin–Huxley |
| LDR | Low dynamic range |
| mRNA | Messenger RNA |
| oBRB | Outer blood–retina barrier |
| OCT | Optical coherence tomography |
| ODEs | Ordinary differential equations |
| OPL | Outer plexiform layer |
| OR | Outer retina |
| OS | Outer segments |
| PR | Photoreceptor |
| RPE | Retinal pigment epithelium |
| SACs | Starburst amacrine cells |
| SHP | Slow hyperpolarization current |
| TJs | Tight junctions |
| TMOs | Tone mapping operators |
| VEGF | Vascular endothelial growth factor |
| VLSI | Very large scale integrated |
| VR | Virtual Retina |
References
- Fangueiro, J.F.; Veiga, F.; Silva, A.M.; Souto, E.B. Ocular Drug Delivery—New Strategies for Targeting Anterior and Posterior Segments of the Eye. Curr. Pharm. Des. 2016, 22, 1135–1146. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Lopez, E.; Egea, M.A.; Davis, B.M.; Guo, L.; Espina, M.; Silva, A.M.; Calpena, A.C.; Souto, E.M.B.; Ravindran, N.; Ettcheto, M.; et al. Memantine-Loaded PEGylated Biodegradable Nanoparticles for the Treatment of Glaucoma. Small 2018, 14, 1701808. [Google Scholar] [CrossRef]
- Souto, E.B.; Dias-Ferreira, J.; Lopez-Machado, A.; Ettcheto, M.; Cano, A.; Camins Espuny, A.; Espina, M.; Garcia, M.L.; Sanchez-Lopez, E. Advanced Formulation Approaches for Ocular Drug Delivery: State-Of-The-Art and Recent Patents. Pharmaceutics 2019, 11, 460. [Google Scholar] [CrossRef] [PubMed]
- Querques, L.; Parravano, M.; Borrelli, E.; Chiaravalloti, A.; Tedeschi, M.; Sacconi, R.; Zucchiatti, I.; Bandello, F.; Querques, G. Anatomical and functional changes in neovascular AMD in remission: Comparison of fibrocellular and fibrovascular phenotypes. Br. J. Ophthalmol 2020, 104, 47–52. [Google Scholar] [CrossRef]
- Cho, H.J.; Kim, K.; Lim, S.H.; Kang, D.H.; Kim, J.W. Retinal pigment epithelial atrophy after anti-vascular endothelial growth factor therapy for polypoidal choroidal vasculopathy. Br. J. Ophthalmol. 2020, 10, 1136. [Google Scholar] [CrossRef] [PubMed]
- Hoon, M.; Okawa, H.; Della Santina, L.; Wong, R.O.L. Functional architecture of the retina: Development and disease. Prog. Retin. Eye Res. 2014, 42, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Palczewski, K. Chemistry and biology of the initial steps in vision: The Friedenwald lecture. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6651–6672. [Google Scholar] [CrossRef] [PubMed]
- Strauss, O. The Retinal Pigment Epithelium in Visual Function. Physiol. Rev. 2005, 85, 845–881. [Google Scholar] [CrossRef]
- Kevany, B.M.; Palczewski, K. Phagocytosis of retinal rod and cone photoreceptors. Physiology 2010, 25, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Cartwright, R.A.; Schwartz, R.S.; Merry, A.L.; Howell, M.M. The importance of selection in the evolution of blindness in cavefish. BMC Evol. Biol. 2017, 17, 45. [Google Scholar] [CrossRef]
- Sanchez-Lopez, E.; Egea, M.A.; Cano, A.; Espina, M.; Calpena, A.C.; Ettcheto, M.; Camins, A.; Souto, E.B.; Silva, A.M.; Garcia, M.L. PEGylated PLGA nanospheres optimized by design of experiments for ocular administration of dexibuprofen-in vitro, ex vivo and in vivo characterization. Colloids Surf. B Biointerfaces 2016, 145, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Fangueiro, J.F.; Calpena, A.C.; Clares, B.; Andreani, T.; Egea, M.A.; Veiga, F.J.; Garcia, M.L.; Silva, A.M.; Souto, E.B. Biopharmaceutical evaluation of epigallocatechin gallate-loaded cationic lipid nanoparticles (EGCG-LNs): In vivo, in vitro and ex vivo studies. Int J. Pharm. 2016, 502, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Abrego, G.; Alvarado, H.; Souto, E.B.; Guevara, B.; Bellowa, L.H.; Parra, A.; Calpena, A.; Garcia, M.L. Biopharmaceutical profile of pranoprofen-loaded PLGA nanoparticles containing hydrogels for ocular administration. Eur. J. Pharm. Biopharm. 2015, 95, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Mira, E.; Nikolic, S.; Calpena, A.C.; Egea, M.A.; Souto, E.B.; Garcia, M.L. Improved and safe transcorneal delivery of flurbiprofen by NLC and NLC-based hydrogels. J. Pharm. Sci. 2012, 101, 707–725. [Google Scholar] [CrossRef] [PubMed]
- Araujo, J.; Garcia, M.L.; Mallandrich, M.; Souto, E.B.; Calpena, A.C. Release profile and transscleral permeation of triamcinolone acetonide loaded nanostructured lipid carriers (TA-NLC): In vitro and ex vivo studies. Nanomedicine 2012, 8, 1034–1041. [Google Scholar] [CrossRef]
- Gonzalez-Mira, E.; Egea, M.A.; Souto, E.B.; Calpena, A.C.; Garcia, M.L. Optimizing flurbiprofen-loaded NLC by central composite factorial design for ocular delivery. Nanotechnology 2010, 22, 045101. [Google Scholar] [CrossRef]
- Araujo, J.; Nikolic, S.; Egea, M.A.; Souto, E.B.; Garcia, M.L. Nanostructured lipid carriers for triamcinolone acetonide delivery to the posterior segment of the eye. Colloids Surf. B Biointerfaces 2011, 88, 150–157. [Google Scholar] [CrossRef]
- Kotha, S.; Murtomäki, L. Virtual pharmacokinetic model of human eye. Math. Biosci. 2014, 253, 11–18. [Google Scholar] [CrossRef]
- Bassett, D.S.; Sporns, O. Network neuroscience. Nat. Neurosci. 2017, 20, 353–364. [Google Scholar] [CrossRef]
- Matzakos-Karvouniari, D.; Gil, L.; Orendorff, E.; Marre, O.; Picaud, S.; Cessac, B. A biophysical model explains the spontaneous bursting behavior in the developing retina. Sci. Rep. 2019, 9, 1859. [Google Scholar] [CrossRef]
- Zheng, J.; Lee, S.; Zhou, Z.J. A transient network of intrinsically bursting starburst cells underlies the generation of retinal waves. Nat. Neurosci. 2006, 9, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Joselevitch, C. Human retinal circuitry and physiology. Psychol. Neurosci. 2008, 1, 141–165. [Google Scholar] [CrossRef]
- Huang, P.-C.; Hsiao, Y.-T.; Kao, S.-Y.; Chen, C.-F.; Chen, Y.-C.; Chiang, C.-W.; Lee, C.-F.; Lu, J.-C.; Chern, Y.; Wang, C.-T. Adenosine A2A Receptor Up-Regulates Retinal Wave Frequency via Starburst Amacrine Cells in the Developing Rat Retina. PLoS ONE 2014, 9, e95090. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.W.; Chen, Y.C.; Lu, J.C.; Hsiao, Y.T.; Chang, C.W.; Huang, P.C.; Chang, Y.T.; Chang, P.Y.; Wang, C.T. Synaptotagmin I regulates patterned spontaneous activity in the developing rat retina via calcium binding to the C2AB domains. PLoS ONE 2012, 7, e47465. [Google Scholar] [CrossRef] [PubMed]
- Hanson, L.; Sethuramanujam, S.; deRosenroll, G.; Jain, V.; Awatramani, G.B. Retinal direction selectivity in the absence of asymmetric starburst amacrine cell responses. eLife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Maccione, A.; Hennig, M.H.; Gandolfo, M.; Muthmann, O.; van Coppenhagen, J.; Eglen, S.J.; Berdondini, L.; Sernagor, E. Following the ontogeny of retinal waves: Pan-retinal recordings of population dynamics in the neonatal mouse. J. Physiol. 2014, 592, 1545–1563. [Google Scholar] [CrossRef]
- Sun, Y.; Smith, L.E.H. Retinal Vasculature in Development and Diseases. Annu. Rev. Vis. Sci. 2018, 4, 101–122. [Google Scholar] [CrossRef]
- Kerschensteiner, D. Glutamatergic Retinal Waves. Front. Neural Circuits 2016, 10, 38. [Google Scholar] [CrossRef]
- Fried, S.I.; Münch, T.A.; Werblin, F.S. Mechanisms and circuitry underlying directional selectivity in the retina. Nature 2002, 420, 411–414. [Google Scholar] [CrossRef]
- Agrahari, V.; Mandal, A.; Agrahari, V.; Trinh, H.M.; Joseph, M.; Ray, A.; Hadji, H.; Mitra, R.; Pal, D.; Mitra, A.K. A comprehensive insight on ocular pharmacokinetics. Drug Deliv. Transl. Res. 2016, 6, 735–754. [Google Scholar] [CrossRef]
- Labro, A.J.; Priest, M.F.; Lacroix, J.J.; Snyders, D.J.; Bezanilla, F. Kv3.1 uses a timely resurgent K+ current to secure action potential repolarization. Nat. Commun. 2015, 6, 10173. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.F.; Chakarova, C.F.; Abd El-Aziz, M.M.; Bhattacharya, S.S. Photoreceptor degeneration: Genetic and mechanistic dissection of a complex trait. Nat. Rev. Genet. 2010, 11, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Shemonski, N.D.; South, F.A.; Liu, Y.-Z.; Adie, S.G.; Scott Carney, P.; Boppart, S.A. Computational high-resolution optical imaging of the living human retina. Nat. Photonics 2015, 9, 440–443. [Google Scholar] [CrossRef] [PubMed]
- Semple, J.L.; Woolridge, N.; Lumsden, C.J. Review: In Vitro, in Vivo, in Silico: Computational Systems in Tissue Engineering and Regenerative Medicine. Tissue Eng. 2005, 11, 341–356. [Google Scholar] [CrossRef] [PubMed]
- Eshraghian, J.K.; Baek, S.; Kim, J.-H.; Iannella, N.; Cho, K.; Goo, Y.S.; Iu, H.H.C.; Kang, S.-M.; Eshraghian, K. Formulation and Implementation of Nonlinear Integral Equations to Model Neural Dynamics Within the Vertebrate Retina. Int. J. Neural Syst. 2018, 28, 1850004. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, H.; Maurer-Stroh, S.; Cheng, L. Synthesizing retinal and neuronal images with generative adversarial nets. Med. Image Anal. 2018, 49, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.; Galdran, A.; Meyer, M.I.; Niemeijer, M.; Abràmoff, M.; Mendonça, A.M.; Campilho, A. End-to-End Adversarial Retinal Image Synthesis. IEEE Trans. Med. Imaging 2018, 37, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Collins, D.L.; Zijdenbos, A.P.; Kollokian, V.; Sled, J.G.; Kabani, N.J.; Holmes, C.J.; Evans, A.C. Design and construction of a realistic digital brain phantom. IEEE Trans. Med. Imaging 1998, 17, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Hodneland, E.; Hanson, E.; Munthe-Kaas, A.Z.; Lundervold, A.; Nordbotten, J.M. Physical Models for Simulation and Reconstruction of Human Tissue Deformation Fields in Dynamic MRI. IEEE Trans. Biomed. Eng. 2016, 63, 2200–2210. [Google Scholar] [CrossRef]
- van Tulder, G.; de Bruijne, M. Why Does Synthesized Data Improve Multi-sequence Classification. In Medical Image Computing and Computer-Assisted Intervention—MICCAI 2015, Proceedings of the International Conference on Medical Image Computing and Computer-Assisted Intervention, Munich, Germany, 5–9 October 2015; Springer: Cham, Switzerland, 2015; pp. 531–538. [Google Scholar]
- Loizos, K.; Marc, R.; Humayun, M.; Anderson, J.R.; Jones, B.W.; Lazzi, G. Increasing Electrical Stimulation Efficacy in Degenerated Retina: Stimulus Waveform Design in a Multiscale Computational Model. IEEE Trans. Neural Syst. Rehabil. Eng. 2018, 26, 1111–1120. [Google Scholar] [CrossRef]
- Weiland, J.D.; Humayun, M.S. Retinal Prosthesis. IEEE Trans. Biomed. Eng. 2014, 61, 1412–1424. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, A.K.; Yeoh, J.; Dorn, J.D.; Caspi, A.; Wuyyuru, V.; McMahon, M.J.; Humayun, M.S.; Greenberg, R.J.; Dacruz, L. Factors Affecting Perceptual Threshold in Argus II Retinal Prosthesis Subjects. Transl. Vis. Sci. Technol. 2013, 2, 1. [Google Scholar] [CrossRef] [PubMed]
- Ho, A.C.; Humayun, M.S.; Dorn, J.D.; da Cruz, L.; Dagnelie, G.; Handa, J.; Barale, P.-O.; Sahel, J.-A.; Stanga, P.E.; Hafezi, F.; et al. Long-Term Results from an Epiretinal Prosthesis to Restore Sight to the Blind. Ophthalmology 2015, 122, 1547–1554. [Google Scholar] [CrossRef] [PubMed]
- Weitz, A.C.; Behrend, M.R.; Ahuja, A.K.; Christopher, P.; Wei, J.; Wuyyuru, V.; Patel, U.; Greenberg, R.J.; Humayun, M.S.; Chow, R.H.; et al. Interphase gap as a means to reduce electrical stimulation thresholds for epiretinal prostheses. J. Neural Eng. 2014, 11, 016007. [Google Scholar] [CrossRef]
- Hadjinicolaou, A.E.; Savage, C.O.; Apollo, N.V.; Garrett, D.J.; Cloherty, S.L.; Ibbotson, M.R.; Brien, B.J.O. Optimizing the Electrical Stimulation of Retinal Ganglion Cells. IEEE Trans. Neural Syst. Rehabil. Eng. 2015, 23, 169–178. [Google Scholar] [CrossRef]
- Fried, S.I.; Hsueh, H.A.; Werblin, F.S. A Method for Generating Precise Temporal Patterns of Retinal Spiking Using Prosthetic Stimulation. J. Neurophysiol. 2006, 95, 970–978. [Google Scholar] [CrossRef]
- Tomita, M.; Hashimoto, K.; Takahashi, K.; Shimizu, T.S.; Matsuzaki, Y.; Miyoshi, F.; Saito, K.; Tanida, S.; Yugi, K.; Venter, J.C.; et al. E-CELL: Software environment for whole-cell simulation. Bioinformatics 1999, 15, 72–84. [Google Scholar] [CrossRef]
- Becker, S.A.; Feist, A.M.; Mo, M.L.; Hannum, G.; Palsson, B.Ø.; Herrgard, M.J. Quantitative prediction of cellular metabolism with constraint-based models: The COBRA Toolbox. Nat. Protoc. 2007, 2, 727–738. [Google Scholar] [CrossRef]
- Song, Z.; Postma, M.; Billings, S.A.; Coca, D.; Hardie, R.C.; Juusola, M. Stochastic, adaptive sampling of information by microvilli in fly photoreceptors. Curr. Biol. 2012, 22, 1371–1380. [Google Scholar] [CrossRef]
- Song, Z.; Juusola, M. Refractory sampling links efficiency and costs of sensory encoding to stimulus statistics. J. Neurosci. 2014, 34, 7216–7237. [Google Scholar] [CrossRef]
- Juusola, M.; Dau, A.; Song, Z.; Solanki, N.; Rien, D.; Jaciuch, D.; Dongre, S.A.; Blanchard, F.; de Polavieja, G.G.; Hardie, R.C.; et al. Microsaccadic sampling of moving image information provides Drosophila hyperacute vision. eLife 2017, 6, e26117. [Google Scholar] [CrossRef] [PubMed]
- Medathati, N.V.K.; Neumann, H.; Masson, G.S.; Kornprobst, P. Bio-inspired computer vision: Towards a synergistic approach of artificial and biological vision. Comput. Vis. Image Underst. 2016, 150, 1–30. [Google Scholar] [CrossRef]
- Gariano, R.F.; Gardner, T.W. Retinal angiogenesis in development and disease. Nature 2005, 438, 960–966. [Google Scholar] [CrossRef] [PubMed]
- Benedicto, I.; Lehmann, G.L.; Ginsberg, M.; Nolan, D.J.; Bareja, R.; Elemento, O.; Salfati, Z.; Alam, N.M.; Prusky, G.T.; Llanos, P.; et al. Concerted regulation of retinal pigment epithelium basement membrane and barrier function by angiocrine factors. Nat. Commun. 2017, 8, 15374. [Google Scholar] [CrossRef]
- Booij, J.C.; Baas, D.C.; Beisekeeva, J.; Gorgels, T.G.M.F.; Bergen, A.A.B. The dynamic nature of Bruch's membrane. Prog. Retin. Eye Res. 2010, 29, 1–18. [Google Scholar] [CrossRef]
- Rizzolo, L.J. Development and role of tight junctions in the retinal pigment epithelium. Int. Rev. Cytol. 2007, 258, 195–234. [Google Scholar]
- Caceres, P.S.; Benedicto, I.; Lehmann, G.L.; Rodriguez-Boulan, E.J. Directional Fluid Transport across Organ-Blood Barriers: Physiology and Cell Biology. Cold Spring Harb. Perspect. Biol. 2017, 9, a027847. [Google Scholar] [CrossRef]
- Rafii, S.; Butler, J.M.; Ding, B.-S. Angiocrine functions of organ-specific endothelial cells. Nature 2016, 529, 316–325. [Google Scholar] [CrossRef]
- Connolly, S.E.; Hores, T.A.; Smith, L.E.H.; D'Amore, P.A. Characterization of vascular development in the mouse retina. Microvasc. Res. 1988, 36, 275–290. [Google Scholar] [CrossRef]
- Qazi, Y.; Maddula, S.; Ambati, B.K. Mediators of ocular angiogenesis. J Genet 2009, 88, 495–515. [Google Scholar] [CrossRef]
- Gariano, R.F. Cellular mechanisms in retinal vascular development. Prog. Retin. Eye Res. 2003, 22, 295–306. [Google Scholar] [CrossRef]
- Flower, R.W.; McLeod, D.S.; Lutty, G.A.; Goldberg, B.; Wajer, S.D. Postnatal retinal vascular development of the puppy. Investig. Ophthalmol. Vis. Sci. 1985, 26, 957–968. [Google Scholar]
- Chan-Ling, T.; Halasz, P.; Stone, J. Development of retinal vasculature in the cat: Processes and mechanisms. Curr. Eye Res. 1990, 9, 459–478. [Google Scholar] [CrossRef]
- Michaelson, I.C. The mode of development of the vascular system of the retina, with some observations on its significance for certain retinal diseases. Trans. Ophthalmol. Soc. UK 1948, 68, 137–180. [Google Scholar]
- Stalmans, I.; Ng, Y.-S.; Rohan, R.; Fruttiger, M.; Bouché, A.; Yuce, A.; Fujisawa, H.; Hermans, B.; Shani, M.; Jansen, S.; et al. Arteriolar and venular patterning in retinas of mice selectively expressing VEGF isoforms. J. Clin. Investig. 2002, 109, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Claxton, S.; Fruttiger, M. Role of arteries in oxygen induced vaso-obliteration. Exp. Eye Res. 2003, 77, 305–311. [Google Scholar] [CrossRef]
- Alon, T.; Hemo, I.; Itin, A.; Pe’er, J.; Stone, J.; Keshet, E. Vascular endothelial growth factor acts as a survival factor for newly formed retinal vessels and has implications for retinopathy of prematurity. Nat. Med. 1995, 1, 1024–1028. [Google Scholar] [CrossRef] [PubMed]
- Curatola, A.M.; Moscatelli, D.; Norris, A.; Hendricks-Munoz, K. Retinal blood vessels develop in response to local VEGF-A signals in the absence of blood flow. Exp. Eye Res. 2005, 81, 147–158. [Google Scholar] [CrossRef]
- Shih, S.-C.; Ju, M.; Liu, N.; Smith, L.E.H. Selective stimulation of VEGFR-1 prevents oxygen-induced retinal vascular degeneration in retinopathy of prematurity. J. Clin. Investig. 2003, 112, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Gariano, R.F.; Hu, D.; Helms, J. Expression of angiogenesis-related genes during retinal development. Gene Expr. Patterns 2006, 6, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, H.; Golding, M.; Fruttiger, M.; Ruhrberg, C.; Lundkvist, A.; Abramsson, A.; Jeltsch, M.; Mitchell, C.; Alitalo, K.; Shima, D.; et al. VEGF guides angiogenic sprouting utilizing endothelial tip cell filopodia. J. Cell Biol. 2003, 161, 1163–1177. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P.; Tessier-Lavigne, M. Common mechanisms of nerve and blood vessel wiring. Nature 2005, 436, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Feeney, S.A.; Simpson, D.A.C.; Gardiner, T.A.; Boyle, C.; Jamison, P.; Stitt, A.W. Role of Vascular Endothelial Growth Factor and Placental Growth Factors During Retinal Vascular Development and Hyaloid Regression. Investig. Ophthalmol. Vis. Sci. 2003, 44, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Sarlos, S.; Rizkalla, B.; Moravski, C.J.; Cao, Z.; Cooper, M.E.; Wilkinson-Berka, J.L. Retinal angiogenesis is mediated by an interaction between the angiotensin type 2 receptor, VEGF, and angiopoietin. Am. J. Pathol. 2003, 163, 879–887. [Google Scholar] [CrossRef][Green Version]
- Dawson, D.W.; Volpert, O.V.; Gillis, P.; Crawford, S.E.; Xu, H.-J.; Benedict, W.; Bouck, N.P. Pigment Epithelium-Derived Factor: A Potent Inhibitor of Angiogenesis. Science 1999, 285, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Walpole, J.; Mac Gabhann, F.; Peirce, S.M.; Chappell, J.C. Agent-based computational model of retinal angiogenesis simulates microvascular network morphology as a function of pericyte coverage. Microcirculation 2017, 24, e12393. [Google Scholar] [CrossRef] [PubMed]
- Eshraghian, J.K.; Baek, S.; Cho, K.; Iannella, N.; Kim, J.; Goo, Y.S.; Iu, H.H.C.; Fernando, T.; Eshraghian, K. Modelling and analysis of signal flow platform implementation into retinal cell pathway. In Proceedings of the 2016 IEEE Asia Pacific Conference on Circuits and Systems (APCCAS), Jeju, Korea, 25–28 October 2016; pp. 491–494. [Google Scholar]
- Draghici, S. Neural networks in analog hardware—Design and implementation issues. Int. J. Neural Syst. 2000, 10, 19–42. [Google Scholar] [CrossRef]
- Tojo, K. A Pharmacokinetic Model for Ocular Drug Delivery. Chem. Pharm. Bull. 2004, 52, 1290–1294. [Google Scholar] [CrossRef][Green Version]
- Tojo, K.J.; Ohtori, A. Pharmacokinetic model of intravitreal drug injection. Math. Biosci. 1994, 123, 59–75. [Google Scholar] [CrossRef]
- Wyatt, H.J. Modelling Transport in the Anterior Segment of the Eye. Optom. Vis. Sci. 2004, 81, 272–282. [Google Scholar] [CrossRef]
- Balachandran, R.K.; Barocas, V.H. Computer Modeling of Drug Delivery to the Posterior Eye: Effect of Active Transport and Loss to Choroidal Blood Flow. Pharm. Res. 2008, 25, 2685–2696. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, A.; Vishnampet, R. Effect of choroidal blood flow on transscleral retinal drug delivery using a porous medium model. Int. J. Heat Mass Transf. 2012, 55, 5665–5672. [Google Scholar] [CrossRef]
- Friedrich, S.; Cheng, Y.-L.; Saville, B. Finite element modeling of drug distribution in the vitreous humor of the rabbit eye. Ann. Biomed. Eng. 1997, 25, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Haghjou, N.; Abdekhodaie, M.J.; Cheng, Y.L.; Saadatmand, M. Computer modeling of drug distribution after intravitreal administration. World Acad. Sci. Eng. Technol. 2011, 77, 706–716. [Google Scholar]
- Cho, K.; Baek, S.; Cho, S.W.; Kim, J.H.; Goo, Y.S.; Eshraghian, J.K.; Iannella, N.; Eshraghian, K. Signal Flow Platform for Mapping and Simulation of Vertebrate Retina for Sensor Systems. IEEE Sens. J. 2016, 16, 5856–5866. [Google Scholar] [CrossRef]
- Loizos, K.; Lazzi, G.; Lauritzen, J.S.; Anderson, J.; Jones, B.W.; Marc, R. A multi-scale computational model for the study of retinal prosthetic stimulation. In Proceedings of the 2014 36th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Chicago, IL, USA, 26–30 August 2014. [Google Scholar]
- Venhuizen, F.G.; van Ginneken, B.; Liefers, B.; van Grinsven, M.J.J.P.; Fauser, S.; Hoyng, C.; Theelen, T.; Sánchez, C.I. Robust total retina thickness segmentation in optical coherence tomography images using convolutional neural networks. Biomed. Opt. Express 2017, 8, 3292–3316. [Google Scholar] [CrossRef]
- Debevec, P.; Malik, J. Recovering High Dynamic Range Radiance Maps from Photographs. In Proceedings of the 24th International Conference on Computer Graphics and Interactive Techniques, Los Angeles, CA, USA, 3–8 August 1997; pp. 369–378. [Google Scholar] [CrossRef]
- Reinhard, E.; Devlin, K. Dynamic range reduction inspired by photoreceptor physiology. IEEE Trans. Vis. Comput. Graph. 2005, 11, 13–24. [Google Scholar] [CrossRef]
- Benzi, M.; Escobar, M.J.; Kornprobst, P. A bio-inspired synergistic virtual retina model for tone mapping. Comput. Vis. Image Underst. 2018, 168, 21–36. [Google Scholar] [CrossRef]
- Wohrer, A.; Kornprobst, P. Virtual Retina: A biological retina model and simulator, with contrast gain control. J. Comput. Neurosci. 2009, 26, 219–249. [Google Scholar] [CrossRef]
- Rahman, Z.U.; Jobson, D.; Woodell, G. Multiscale retinex for color rendition and dynamic range compression. In Applications of Digital Image Processing XIX; SPIE: Bellingham, WA, USA, 1996; Volume 2847. [Google Scholar]
- Land, E.H.; McCann, J.J. Lightness and Retinex Theory. J. Opt. Soc. Am. 1971, 61, 1–11. [Google Scholar] [CrossRef]
- Benoit, A.; Alleysson, D.; Herault, J.; Le Callet, P. Spatio-temporal tone mapping operator based on a retina model. In International Workshop on Computational Color Imaging; Springer: Berlin, Germany, 2009. [Google Scholar]
- Benoit, A.; Caplier, A.; Durette, B.; Herault, J. Using Human Visual System modeling for bio-inspired low level image processing. Comput. Vis. Image Underst. 2010, 114, 758–773. [Google Scholar] [CrossRef]
- Herault, J. Color Processing in the Retina. In Vision: Images, Signals and Neural Networks; Progress in Neural Processing: Volume 19; World Scientific: Singapore; London, UK; Hackensack, NJ, USA, 2010; pp. 141–172. [Google Scholar]
- Zhang, X.-S.; Gao, S.-B.; Li, C.-Y.; Li, Y.-J. A Retina Inspired Model for Enhancing Visibility of Hazy Images. Front. Comput. Neurosci. 2015, 9, 151. [Google Scholar] [CrossRef] [PubMed]
- Hennig, M.H.; Adams, C.; Willshaw, D.; Sernagor, E. Early-stage waves in the retinal network emerge close to a critical state transition between local and global functional connectivity. J. Neurosci. 2009, 29, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- O’Hearn, T.M.; Sadda, S.R.; Weiland, J.D.; Maia, M.; Margalit, E.; Humayun, M.S. Electrical stimulation in normal and retinal degeneration (rd1) isolated mouse retina. Vis. Res. 2006, 46, 3198–3204. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Humayun, M.S.; Weiland, J.D.; Chen, S.-J.; Margalit, E.; Piyathaisere, D.V.; de Juan, E. Comparison of Electrical Stimulation Thresholds in Normal and Retinal Degenerated Mouse Retina. Jpn. J. Ophthalmol. 2004, 48, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.J. Activation of ganglion cells in wild-type and P23H rat retinas with a small subretinal electrode. Exp. Eye Res. 2012, 99, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.J.; Rizzo, J.F. Activation of ganglion cells in wild-type andrd1mouse retinas with monophasic and biphasic current pulses. J. Neural Eng. 2009, 6, 35004. [Google Scholar] [CrossRef]
- Jensen, R.J.; Rizzo, J.F. Activation of retinal ganglion cells in wild-type and rd1 mice through electrical stimulation of the retinal neural network. Vis. Res. 2008, 48, 1562–1568. [Google Scholar] [CrossRef]
- Sekirnjak, C.; Hulse, C.; Jepson, L.H.; Hottowy, P.; Sher, A.; Dabrowski, W.; Litke, A.M.; Chichilnisky, E.J. Loss of responses to visual but not electrical stimulation in ganglion cells of rats with severe photoreceptor degeneration. J. Neurophysiol. 2009, 102, 3260–3269. [Google Scholar] [CrossRef]
- Sekirnjak, C.; Jepson, L.H.; Hottowy, P.; Sher, A.; Dabrowski, W.; Litke, A.M.; Chichilnisky, E.J. Changes in physiological properties of rat ganglion cells during retinal degeneration. J. Neurophysiol. 2011, 105, 2560–2571. [Google Scholar] [CrossRef]
- Weitz, A.C.; Nanduri, D.; Behrend, M.R.; Gonzalez-Calle, A.; Greenberg, R.J.; Humayun, M.S.; Chow, R.H.; Weiland, J.D. Improving the spatial resolution of epiretinal implants by increasing stimulus pulse duration. Sci. Transl. Med. 2015, 7, 318ra203. [Google Scholar] [CrossRef] [PubMed]
- Jepson, L.H.; Hottowy, P.; Mathieson, K.; Gunning, D.E.; Dabrowski, W.; Litke, A.M.; Chichilnisky, E.J. Focal electrical stimulation of major ganglion cell types in the primate retina for the design of visual prostheses. J. Neurosci. 2013, 33, 7194–7205. [Google Scholar] [CrossRef] [PubMed]
- Abramian, M.; Lovell, N.H.; Morley, J.W.; Suaning, G.J.; Dokos, S. Activation and inhibition of retinal ganglion cells in response to epiretinal electrical stimulation: A computational modelling study. J. Neural Eng. 2014, 12, 16002. [Google Scholar] [CrossRef] [PubMed]
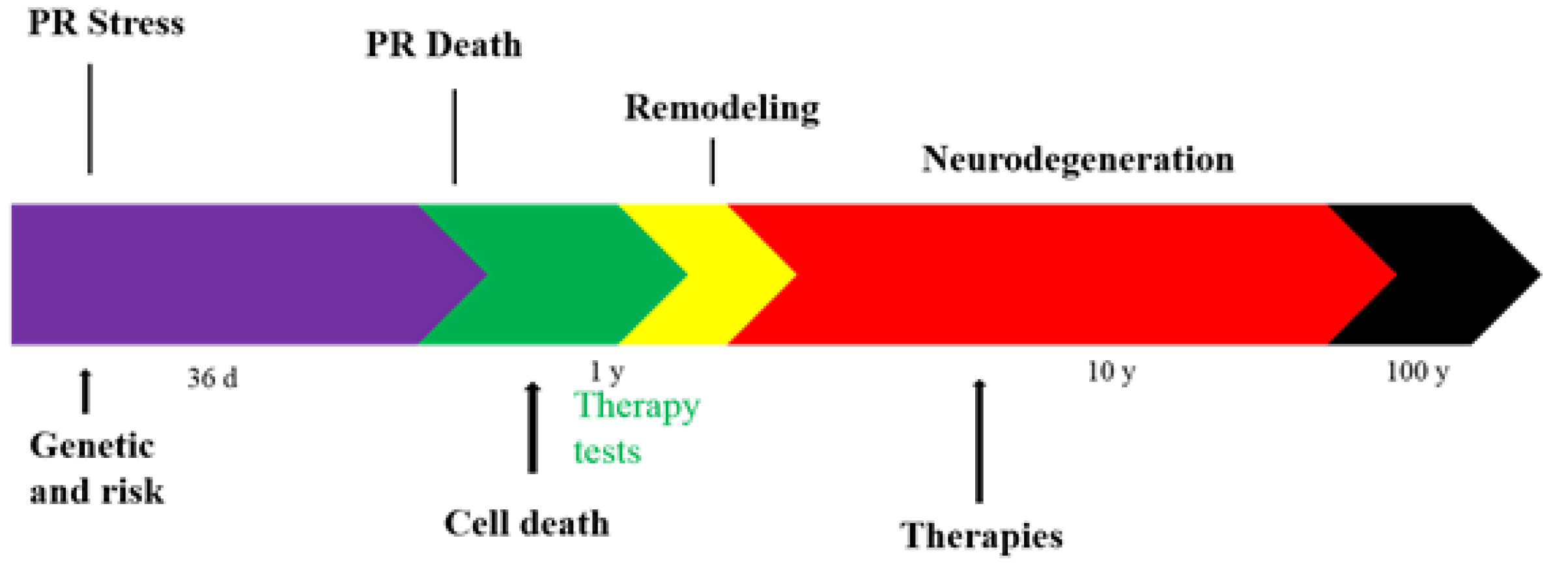
- Jones, B.W.; Watt, C.B.; Frederick, J.M.; Baehr, W.; Chen, C.-K.; Levine, E.M.; Milam, A.H.; Lavail, M.M.; Marc, R.E. Retinal remodeling triggered by photoreceptor degenerations. J. Comp. Neurol. 2003, 464, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.W.; Pfeiffer, R.L.; Ferrell, W.D.; Watt, C.B.; Tucker, J.; Marc, R.E. Retinal Remodeling and Metabolic Alterations in Human AMD. Front. Cell. Neurosci. 2016, 10, 103. [Google Scholar] [CrossRef] [PubMed]
- Trenholm, S.; Awatramani, G.B. Origins of spontaneous activity in the degenerating retina. Front. Cell. Neurosci. 2015, 9, 277. [Google Scholar] [CrossRef] [PubMed]
- Borowska, J.; Trenholm, S.; Awatramani, G.B. An Intrinsic Neural Oscillator in the Degenerating Mouse Retina. J. Neurosci. 2011, 31, 5000–5012. [Google Scholar] [CrossRef]
- Toychiev, A.H.; Ivanova, E.; Yee, C.W.; Sagdullaev, B.T. Block of gap junctions eliminates aberrant activity and restores light responses during retinal degeneration. J. Neurosci. 2013, 33, 13972–13977. [Google Scholar] [CrossRef]
- Choi, H.; Zhang, L.; Cembrowski, M.S.; Sabottke, C.F.; Markowitz, A.L.; Butts, D.A.; Kath, W.L.; Singer, J.H.; Riecke, H. Intrinsic bursting of AII amacrine cells underlies oscillations in the rd1 mouse retina. J. Neurophysiol. 2014, 112, 1491–1504. [Google Scholar] [CrossRef]
- Cho, A.; Ratliff, C.; Sampath, A.; Weiland, J. Changes in ganglion cell physiology during retinal degeneration influence excitability by prosthetic electrodes. J. Neural Eng. 2016, 13, 025001. [Google Scholar] [CrossRef]
- Goo, Y.S.; Park, D.J.; Ahn, J.R.; Senok, S. Spontaneous oscillatory rhythms in the degenerating mouse retina modulate retinal ganglion cell responses to electrical stimulation. Front. Cell. Neurosci. 2016, 9. [Google Scholar] [CrossRef] [PubMed]
- Park, D.J.; Senok, S.S.; Goo, Y.S. Degeneration stage-specific response pattern of retinal ganglion cell spikes in rd10 mouse retina. In Proceedings of the 2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Milan, Italy, 25–29 August 2015; pp. 3351–3354. [Google Scholar]
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500–544. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Cañada, P.; Morillas, C.; Pino, B.; Ros, E.; Pelayo, F. A Computational Framework for Realistic Retina Modeling. Int. J. Neural Syst. 2016, 26, 1650030. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Farhat, N.H. The double queue method: A numerical method for integrate-and-fire neuron networks. Neural Netw. 2001, 14, 921–932. [Google Scholar] [CrossRef]
- Momiji, H.; Bharath, A.A.; Hankins, M.W.; Kennard, C. Numerical study of short-term afterimages and associate properties in foveal vision. Vis. Res. 2006, 46, 365–381. [Google Scholar] [CrossRef][Green Version]
- Momiji, H.; Hankins, M.W.; Bharath, A.A.; Kennard, C. A numerical study of red–green colour opponent properties in the primate retina. Eur. J. Neurosci. 2007, 25, 1155–1165. [Google Scholar] [CrossRef]
- Radisic, M.; Deen, W.; Langer, R.; Vunjak-Novakovic, G. Mathematical model of oxygen distribution in engineered cardiac tissue with parallel channel array perfused with culture medium containing oxygen carriers. Am. J. Physiol. Circ. Physiol. 2005, 288, H1278. [Google Scholar] [CrossRef]
- Kushner, J.I.V.; Deen, W.; Blankschtein, D.; Langer, R. First-principles, structure-based transdermal transport model to evaluate lipid partition and diffusion coefficients of hydrophobic permeants solely from stratum corneum permeation experiments. J. Pharm. Sci. 2007, 96, 3236–3251. [Google Scholar] [CrossRef]
- Trucco, G.; Liberali, V. Analog Design Issues for Mixed-Signal CMOS Integrated Circuits. In Advances in Analog Circuits; Tlelo-Cuautle, E., Ed.; InTechOpen: London, UK, 2011. [Google Scholar]
- Waldstein, S.M.; Gerendas, B.S.; Montuoro, A.; Simader, C.; Schmidt-Erfurth, U. Quantitative comparison of macular segmentation performance using identical retinal regions across multiple spectral-domain optical coherence tomography instruments. Br. J. Ophthalmol. 2015, 99, 794–800. [Google Scholar] [CrossRef]
- Ronneberger, O.; Fischer, P.; Brox, T. U-net: Convolutional networks for biomedical image segmentation. Lect. Notes Comput. Sci. 2015, 9351, 234–241. [Google Scholar]
- Loizos, K.; RamRakhyani, A.K.; Anderson, J.; Marc, R.; Lazzi, G. On the computation of a retina resistivity profile for applications in multi-scale modeling of electrical stimulation and absorption. Phys. Med. Biol. 2016, 61, 4491–4505. [Google Scholar] [CrossRef] [PubMed]
- Cela, C.J.; Lee, R.C.; Lazzi, G. Modeling Cellular Lysis in Skeletal Muscle Due to Electric Shock. IEEE Trans. Biomed. Eng. 2011, 58, 1286–1293. [Google Scholar] [CrossRef] [PubMed]
- Marc, R.E.; Jones, B.W.; Watt, C.B.; Anderson, J.R.; Sigulinsky, C.; Lauritzen, S. Retinal connectomics: Towards complete, accurate networks. Prog. Retin. Eye Res. 2013, 37, 141–162. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Synaptic Transmission | Main Action | Mediated By | Effect |
|---|---|---|---|
| Gap junctions | Activity-dependent regulation and plasticity | Electrical synapse coding ON–OFF directionally selective ganglion cells | Dynamic interactions between electrical and chemical synapses |
| Acetylcholine | To held under a current clamp (Ih = 0) when a voltage is pulsed from −70 mV to 15 mV. | Transient network neuron with an ECl (−70 mV) manipulation, and A2AR up-regulate SACs’ presynaptic waves. | Elicits retinal waves |
| Glutamate | Ultra-fast response of photoreceptors synapse | P8 and P22 retinas | Restores bursting activity |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souto, E.B.; Sanchez-Lopez, E.; Campos, J.R.; da Ana, R.; Espina, M.; Garcia, M.L.; Severino, P.; Batain, F.; F. R. Alves, T.; Crescencio, K.M.M.; et al. Retinal Drug Delivery: Rethinking Outcomes for the Efficient Replication of Retinal Behavior. Appl. Sci. 2020, 10, 4258. https://doi.org/10.3390/app10124258
Souto EB, Sanchez-Lopez E, Campos JR, da Ana R, Espina M, Garcia ML, Severino P, Batain F, F. R. Alves T, Crescencio KMM, et al. Retinal Drug Delivery: Rethinking Outcomes for the Efficient Replication of Retinal Behavior. Applied Sciences. 2020; 10(12):4258. https://doi.org/10.3390/app10124258
Chicago/Turabian StyleSouto, Eliana B., Elena Sanchez-Lopez, Joana R. Campos, Raquel da Ana, Marta Espina, Maria L. Garcia, Patrícia Severino, Fernando Batain, Thais F. R. Alves, Kessi M. M. Crescencio, and et al. 2020. "Retinal Drug Delivery: Rethinking Outcomes for the Efficient Replication of Retinal Behavior" Applied Sciences 10, no. 12: 4258. https://doi.org/10.3390/app10124258
APA StyleSouto, E. B., Sanchez-Lopez, E., Campos, J. R., da Ana, R., Espina, M., Garcia, M. L., Severino, P., Batain, F., F. R. Alves, T., Crescencio, K. M. M., Souto, S. B., & V. Chaud, M. (2020). Retinal Drug Delivery: Rethinking Outcomes for the Efficient Replication of Retinal Behavior. Applied Sciences, 10(12), 4258. https://doi.org/10.3390/app10124258