Abstract
Climate change has enhanced the occurrence of rural fires, since changes in the hydrological cycle have led to the occurrence of increasingly long and frequent periods of drought. This recurrence of rural fires in Portugal, in turn, has led to the successive elimination of vast areas traditionally occupied by native species or species of economic interest, which are being successively replaced by new species with invasive behavior. Among these, Acacia dealbata stands out for its dispersion capacity and for the area it has already occupied. In the present work, which reviews the evolution of forest cover over the last 18,000 years in the Serra da Estrela Natural Park, we intend to demonstrate that fire acts as a species selection agent and that it enhances the development of heliophile and pyrophyte species. For this purpose, an area of the municipality of Seia was selected, more specifically Casal do Rei, where the development of Acacia dealbata forests is monitored. In the end, it was concluded that, in fact, by analyzing the ages of the specimens present in these populations, fire acts as a selection agent by freeing up the space previously occupied by other species, opening the way for the growth of heliophiles and pyrophytes invasive species while enhancing their germination.
1. Introduction
Ecosystems are currently facing intense pressures related to changes driven by climate change [1,2,3,4]. However, species do not react in the same way to these adaptations, since not all respond equally to different stimuli, such as the increase in the average temperature of the air, decrease in the levels of humidity of the air, increase of the periods of drought, decrease of the precipitation, and alterations in the cycle of the seasons, among others [5,6,7,8]. This inability to react or difficulty in reacting to new conditions leads to a change in the ecosystem balance over time, and in this way, ecosystems evolve according to the new conditions, which is often the reason why traditional species are replaced by others that are more adapted to these new conditions [9,10,11].
Usually, it is in these scenarios that some species find the opportunity to become predominant, especially when exotic species which may acquire invasive behavior are present in ecosystems [12,13]. In this way, these species will occupy the niches left vacant by the native species, often becoming dominant, relegating native species to a secondary role in the occupation of physical space and changing the balance in the inter-species relationships [14,15]. Obviously, this type of change will cause imbalances at all levels and in all relationships within the ecosystem, leading to a development that favors the needs of the new dominant species [16].
Fire is an element that has always been present in the Mediterranean landscape, being an integral part of the phenomena that shaped both the landscape and the species that developed and evolved in this environment [17,18,19]. However, as a consequence of climate change, there has been an increase in the occurrence of extreme climatic phenomena, including profound changes in the hydrological cycle and the subsequent occurrence of periods of more frequent and prolonged drought [20,21]. This factor, associated with the presence of a set of exotic species of a pyrophyte characteristics, leads to an artificial selection of species by fire, which will contribute to the replacement of traditional indigenous species, since pyrophyte species take advantage of the openings created by fire to quickly develop and conquer the space previously occupied by native species [22,23].
This fast growth of exotic species, in relation to native species, is decisive for progressively replacing the vegetation cover. In the case of forest cover, this substitution of native species by exotic pyrophyte species, in addition to the losses associated with biodiversity and with the relationships previously established in the ecosystem, may be associated with the destruction of production crops, such as, for example, Eucalyptus globulus plantations for the pulp industry, or Pinus pinaster for the wood pellet industry, since, due to the rapid growth they present nd the aggressiveness with which occupy the physical space, would enter into direct competition with these species already installed, leading to their development being affected and to subsequent losses in productivity [24,25,26,27,28,29]. In Portugal, one of the species that stands out the most for its invasive behavior and for the proliferation that has occurred in recent years is Acacia dealbata, which has been occupying more and more area from year to year, forming dense forests where no other species can survive; such is the density that this species can develop and present [30]. At present, again under the influence of changes in the climate, and with the triggering of associated phenomena, such as the occurrence of long periods of drought and a subsequent increase in the recurrence of rural fires, the forest cover is also changing [31,32,33]. This is causing an artificial selection of species, where pyrophyte species, mostly exotic, such as Acacia dealbata, gain an advantage and take up more and more area at the expense of native species.
This study presents a review on the sequence of the development of forest cover in the region during a period that comprises the last 18,000 years, during which there was a stabilization of the different forest species, followed by the presentation of a case study in which the developments in the forest cover that occurred in the last two dozen years are analyzed. It is assumed that the increase in the recurrence of rural fires in the region causes changes in the structure of the forest cover through the selection of species by fire, namely one species in particular, Acacia dealbata, which stands out for the area that it already occupies. An analysis of the occurrence of rural fires in the region and the impact on the current state of forest cover was made using the information available in databases, namely those at Instituto Português do Mar e da Atmosfera (IPMA) for climatological data and by Sistema Nacional Informação Geográfica (SNIG) for georeferenced data on occurrences of rural fires. Information made available by the press on rural fires in the region was also collected, as was information from interviews with the inhabitants of Cabeça and Casal do Rei, about the perception of the evolution of the climate in the region and on the severity of rural fires that occurred in the past. Some of the methods currently used to control Acacia dealbata are also presented.
2. Geophysical and Biophysical Framework of the Area under Study
2.1. Location and Limits
The area selected for this study is located on a mountain slope in front of the village of Cabeça and the village of Casal do Rei. Both locations belong to the Union of Parishes of Vide and Cabeça in the municipality of Seia and the district of Guarda. Figure 1 shows the location of the target area and its regional and national context.
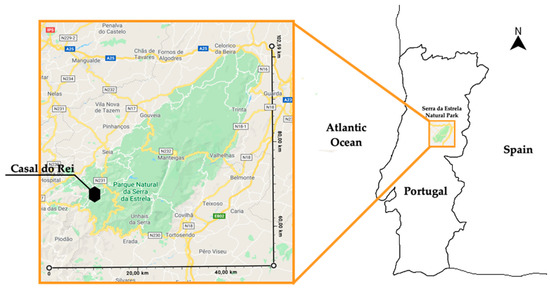
Figure 1.
Location of the target area and its regional and national context.
The Acacia dealbata groves considered in this study, as already mentioned, are visible from the road that connects Casal do Rei to Cabeça, located on the opposite slope, on the other side of the valley, where the Loriga stream runs, as can be seen in Figure 2.

Figure 2.
Acacia dealbata groves on the slope opposite the road from Casal do Rei to Cabeça. Acacia groves are identifiable by their very characteristic greenish color.
2.2. Hydrography
The Serra da Estrela Natural Park covers two hydrographic basins, the Tagus River and the Mondego River [34]. Three important Portuguese rivers begin in the Serra da Estrela Natural Park—Mondego, Zêzere, and Alva [35]. The Mondego River is the largest Portuguese river with a hydrographic basin entirely in national territory [36]. The Alva River is one of the most important tributaries of the Mondego River, which has its source in Serra da Estrela, located near Sabugueiro, in the Rossim Valley area, at approximately 1500 m elevation [37]. The Zêzere River is the largest tributary of the Tagus River in Portugal, beginning in the middle of the glacier valley in the heart of the Serra da Estrela, at Covão d’Ametade (at about 1900 m elevation). In its upper course, upstream of Manteigas, it traverses a typical glacier valley with a U-section [38]. In the central area of the massif, there are several lagoons of glacial origin, some of which have been used for the construction of multi-purpose projects, with emphasis on hydroelectric power plants. The area under study is located on the left bank of the Loriga stream, one of the tributaries of the Alva River, which is formed by the union of the Nave stream and the São Bento stream [39].
2.3. Geomorphology
The geomorphology that characterizes the region essentially derives from tectonic displacements, which lifted the mountain from the surrounding plateaus and pushed it towards the Northeast [35]. The cliffs that limit it are fault cliffs with a relatively long evolution, which must have given rise to the mountain [40]. However, the great gaps that are observed in Serra da Estrela are not only due to tectonic movements, but also due to the deep indentations of the rivers, induced by the rising of the mountain itself from the marginal plateaus [41]. The forms of the relief can have very different origins. Some depend on the nature of the rocks, and others are related to much colder climates than the current one, particularly when the temperature dropped by at least 10 °C around twenty thousand years ago, giving rise to vast glaciers in the highest part of Serra da Estrela [39]. These climates left remarkable geomorphological testimonies, translated as U-shaped valleys, glacial circuses, lagoons, moray deposits, and erratic blocks, constituting the main originality of the physical landscape of the Natural Park [42].
2.4. Geology
The geology of the Serra da Estrela is dominated by the occurrence of hercynitic granitic rocks, which intruded the pre-Cambrian and Cambrian metasediments that constitute the grauvachic schist complex [43]. These rocks have a varied mineralogical composition, from granodiorites to leucogranites [44]. The area under study is essentially made up of schist terrain and, to a lesser extent, fluvial sedimentary deposits. The schists belong to the Malpica do Tejo formation and are between 500 and 650 million years old [45]. In these rocks, there are some quartz veins of reduced thickness and extension [46]. The river deposits, of recent age, are found along the bed of the Loriga stream, have a very reduced width and thickness, and are mainly composed of blocks and rolled pebbles (of different lithologies and dimensions) and sands [47].
2.5. Pedology
The soils in the area selected for the present study are mostly lithosols, which evolved into cambisols in some places where terraces were built, which were used and still are used in some places for agro-silvo-pastoral activity [48,49]. In the places where the activity stopped, the bushes and the forest slowly occupied the space, and these terraces are currently covered with spontaneous vegetation [50].
2.6. Climatology
A data search was then carried out on the meteorological stations geographically closest to the target area. Data were available on the website of IPMA (Instituto Português do Mar e da Atmosfera), which included Viseu, Guarda, and Castelo Branco, since the total absence of thermopluviometric stations in the target zone made it impossible to characterize the current climate in the locality, and only a comparison with these stations located nearby was possible. However, it should be noted that there can be significant differences in the conditions observed, mainly due to the different elevations at which these thermopluviometric stations are in relation to the target zone. Casal do Rei is about 42 km from Viseu, 49 km from Guarda, and 58 km from Castelo Branco.
For values corresponding to Casal do Rei, METEOBLUE (https://www.meteoblue.com) was used and the data modeled are shown in Table 1 and Figure 3. In fact, as can be seen, using the values modeled for Casal do Rei, although minimum negative minimum temperatures have always occurred during the winter months, it was observed and verified that these occurrences have decreased, although there is a dispersion of these occurrences for months other than the usual ones. It is also verified that, although periods with minimum temperatures have decreased and temperatures have risen, some phenomena that can be considered extreme occur, in which the minimum temperatures can fall below the values considered normal, and often outside the usual months. It is also verified that in the summer months, the maximum temperatures have been successively showing higher values, complemented by a decrease in the values of precipitation, which in recent years has been observed to be nonexistent, increasing the probability of the occurrence of rural fires as conditions become ideal for their proliferation.

Table 1.
Summary table of data collected on the website of IPMA (Instituto Português do Mar e da Atmosfera) for the three selected stations, namely, Viseu, Guarda, and Castelo Branco, due to the proximity to the target area, and of the data modeled for Casal do Rei using the platform METEOBLUE. Taverage represents the average temperature, Tmax represents the maximum temperature, Tmin represents the minimum temperature, and Paverage represents the average precipitation.
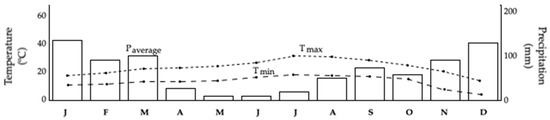
Figure 3.
Air temperature and precipitation based on the information available in Table 1 for Casal do Rei using the values modeled with METEOBLUE (https://www.meteoblue.com). In the figure, the maximum temperature (Tmax), minimum temperature (Tmin), and precipitation are represented.
Despite the geographical proximity between the area under study and the Penhas da Saúde weather station, we decided not to use the data from this station, since in this specific case, the influence of elevation, especially during winter, can result in significantly different effects compared to the real conditions observed in Casal do Rei. Therefore, we opted for the use of data related to the weather stations of Viseu, Guarda, and Castelo Branco since they present conditions closer to the reality observed in Casal do Rei. For example, in conversations with permanent residents in Casal do Rei and in Cabeça, in response to the question about how often it snowed in both locations, everyone replied, “Now, almost every year, snow falls, but it only serves to kill the pests and rarely stays for long periods”. They also replied, “It used to be different,” and “Sometimes, we stayed closed in for weeks down here (Casal do Rei) because cars did not pass with the snow up there”, thus indicating certainty that snow did not accumulate at the bottom of the embedded valley that comprises the Loriga stream. This information is in line with the data that were collected on the website of IPMA (Instituto Português do Mar e da Atmosfera) (www.ipma.pt).
In general, the region has a Mediterranean climate, with two mild seasons, spring and autumn, separated by two extreme seasons, one hot and dry (summer), and the other cold and wet (winter) [51,52]. The patterns of precipitation and temperature are, in the case of the Loriga stream valley, further conditioned by the effect of the orientation of the slope and the surrounding relief [53]. Thus, the embedded valley and the exposure to the North determine that, in the area selected for the present study, relatively mild temperatures and high humidity are felt.
3. Evolution of Vegetation Cover
The oldest mountainous deposits in the entire central cordillera date back to the late glacial period (around 13,000 years ago) and are found only in Serra da Estrela, the most western part of the entire central mountain range [50,54].
According to the fossil records found, the genus Pinus is the most represented in the interior of Serra da Estrela, indicating the presence of pine forests (probably in areas of microthermal climate, forming both open and dense forest structures) at medium to low elevations [55,56]. However, as presented by Rubiales et al. (2020), pollen deposits indicate an abrupt change at the beginning of the Holocene (10,350 years ago) in which the genus Pinus makes way for other groups of hardwoods adapted to a mesophilic climate or even of a Mediterranean character, such as Quercus, or with hygrophilic–microthermal characteristics, such as some species of the genus Betula, or species such as Sorbus aucuparia or Frangula alnus (Figure 4) [55].
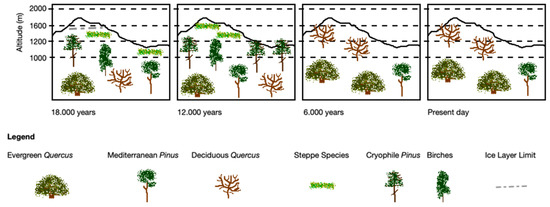
Figure 4.
Evolution of pine forests and other woody formations since the last glacial maximum in the different bands of Serra da Estrela (adapted from [55]).
In the innermost strips, already in Spain, the time records, although referring to a much shorter time period than what is available for Serra da Estrela, show a broadly similar pattern during the second half of the Holocene, where pines do not seem to thrive, most likely due to the decrease in the continentality factor [57]. The pollen sequences found, as well as limited plant remnants of larger dimensions, suggest that the genus Betula was the dominant and most stable genus [55,58].
The decrease in the population of Casal do Rei has direct impacts on landscape management, mainly due to the abandonment of agro-silvo-pastoral activities on the slopes surrounding the village [59]. The agricultural land, essentially formed of terraces with shale walls, disappeared among the spontaneous vegetation that was growing. In view of the anthropic action, the landscape is currently quite altered in relation to the potential natural vegetation [60]. In fact, in recent decades, there has been a strong focus on maritime pine and eucalyptus stands. The changes associated with the monoculture of these stands result in a worsening of the impact of fire and prevent the installation of several native species, especially those belonging to the most evolved stages of the vegetation cover. However, next to Casal do Rei, there are still some fragments of what the native forest of this landscape would be, albeit in very poor condition [60].
The spontaneous vegetation of the area, altered little by human activity, presents a great complexity and floristic diversity, constituting a residual mark of the potential natural vegetation of the Central region of the country, currently covered mainly by eucalyptus, pine forests, and bush and also by patches of acacias. The forest vegetation is characterized by a dense tree layer dominated by Prunus lusitanica, accompanied by Castanea sativa, Viburnum tinus, and Arbutus unedo. Figure 5 presents the current land cover, where the development of acacia groves can be seen.

Figure 5.
Current land cover in the study area.
Although the potential natural vegetation of the slopes of Casal do Rei belongs to the domain of Quercus robur subsp. broteroana, there are no well-constituted forests of this species. Instead, the current vegetation is formed by a set of heliophilous scrublands, lashed by recurrent fires that occur in increasingly shorter cycles, which prevents the installation of the slowest growing and most demanding species at ground level [61,62]. Other types of oak, such as Quercus pyrenaica, Quercus suber, and Quercus rotundifolia, also live in the areas with less water retention; the latter is only present on rocky outcrops. However, on the slopes exposed to the North and with high soil moisture, a relic community resists, dominated by Prunus lusitanica, which is accompanied by a set of pre-forest shrubs, such as Arbutus unedo, Viburnum tinus, and Rhamnus alaternus, among others, which need to be preserved. These communities are even recognized by the European Community, through the Habitats Directive (92/43/EEC), as a priority habitat for conservation (5230 × pt2) called “Laurus nobilis arborescent forests” [63].
However, the recurrence of rural fire favors a vegetation cover dominated by heliophilous shrubs and, when there is a seed bank, an intense regeneration of maritime pine. Among the most frequent shrub communities are Cytisus scoparius, Cytisus grandiflorus, and Genista falcata, as well as the Ulex minor, Erica australis, Erica scoparia, and Erica umbellata that normally live on degraded or stony soils [64]. This dynamic becomes difficult to break with an aging population and with poor resources. In addition, the disappearance of the vegetation cover in the post-fire period favors the expansion of invasive alien species such as Acacia dealbata and Hackea sericea, which, together with depopulation, have all the conditions to thrive [65,66]. With each rural fire, new points of dissemination of these species emerge, spreading in a concentric manner, and thus gradually decreasing the area for native vegetation [65].
4. Impact of Fire on Vegetation Cover
In July 2005, a forest fire reached an area of 17,445 hectares. It severely affected a natural forest of 20 hectares located in Casal do Rei in the Serra da Estrela Natural Park. On 22 July 2005, the newspaper Diário de Notícias, in an article signed by journalist João Fonseca, reported the tragedy that devastated the region, using comments directly from those who experienced the events up close. For example, one of the interviewees stated that, “it can only be arson. Flames appear where and when you least expect them. The firefighters are still extinguishing one and soon another one appears, far or near to the villages, beside the road or in the most inaccessible places, but always where there is thicket and dense forest. This hell seems to have no end. The wonderful forest that was here has all burned.” Other witnesses declared that, “thousands and thousands of pine, eucalyptus, oak, and chestnut trees died, but also olive trees, vines, and gardens.” Many hectares were destroyed after another fire in this hillside of Serra da Estrela, namely in Senhora da Guia, Pedras Lavradas, Teixeira de Cima, Casegas, Casal do Rei, and Cabeça, mainly composed of eucalyptus and maritime pine, but also of a group of native species of great value.
The spontaneous vegetation of the area, little altered by human activity, constitutes a residual mark of the climatic vegetation of the center of the country, currently covered mainly by eucalyptus, pine forests, and scrub. This spontaneous forest, now circumscribed, is characterized by a dense tree layer dominated by Prunus lusitanica, Castanea sativa, Viburnum tinus, and Arbutus unedo. The floristic list of the site includes more than 200 vascular plants species, which corresponds to about a quarter of the total species identified in the region of Serra da Estrela, on an area that represents only 0.02% of the protected area. Among the all the species, Prunus lusitanica stands out because it is considered a relic of the Laurisilva type forests, which had high importance in Iberian Peninsula during the Tertiary period.
However, this event was not a unique case, since it has been repeated over time, as can be seen in the following figures where the burned areas over the period between 1990 and 2018 are presented. As can be seen in Figure 6, there is a high frequency in the occurrence of rural fires, indicated on more than 50% of the area of the geological map 1:50,000 (20B—Covilhã), which served as the basis for the projection of all burned areas over the past 28 years. The data were obtained through the website of the National Geographic Information System (https://snig.dgterritorio.gov.pt/), and were superimposed on the geological map 1:50,000 20-B (Covilhã) using WMS files with QGIS 3.6 open-source software, available through the website https://www.qgis.org/en/site/.

Figure 6.
Overlapping of the burnt areas over the period between 1990 and 2018.
In the particular case of the area between the villages of Cabeça and Casal do Rei, it can be seen that rural fires occurred in the period between 1990 and 1999, with the other situation occurring in the period between 2000 and 2008, more precisely in 2005, as previously mentioned. The area has not been struck again by rural fires since then, probably because the occurrences were close in time; a less intense fire occurred two years earlier on June 24, 2003 and consumed much of the vegetation on the slopes of the embedded valley where the Loriga stream runs, as reported in the online version of the newspaper Correio da Manhã.
This redundant occurrence of rural fires in the Serra da Estrela Natural Park led to a gradual destruction of the native forest, much of which is very difficult to recover. That is the case for the continental Laurisilva forest, where the development of small sanctuaries was resumed at great cost, while the rest of the territory is at the mercy of spontaneous and disorderly growth of maritime pines and eucalyptus, or even to a set of invasive species such as acacias, hakeas, or ailanthus.
In the case of acacias, the Acacia dealbata species stands out because it is the one that progresses the most in the area, already forming extensive patches, and appears to occupy all types of soils and habitats, including rising in elevation to areas not normally occupied by this type of species. In any case, as can be seen in Figure 7, Acacia dealbata already occupies significant areas on land adjacent to the target area of this study, which, despite everything, is not yet in a situation as critical as the one shown in the example presented in following figure.
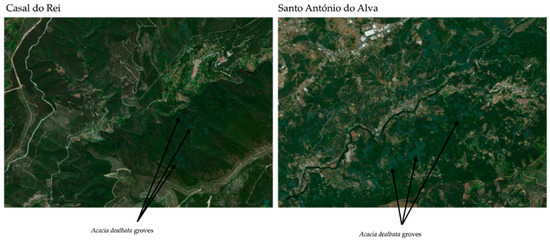
Figure 7.
Acacia dealbata groves. As can be seen in the figure on the right, in the Santo António do Alva region, located close to Casal do Rei and also greatly affected by rural fires, one can see a significant dispersion of Acacia dealbata groves, which are replacing eucalyptus and maritime pine areas. These acacia groves are easily identifiable through their shade of green, distinct from other species found in the region.
In fact, this species proliferated extensively on the slopes of the mountain, with particular emphasis on the area under analysis in the present work, forcing frequent control actions. These control actions aim to reduce the fuel load present in the forest and try to eradicate this invasive species, which is gaining more and more ground, especially after the occurrence of rural fires. Figure 8 shows the work done in one of these control actions.

Figure 8.
Cutting work and application of control techniques for invasive species. (a) Cutting; (b) peeling the acacias; and (c) application of glyphosate.
In this control action, several methods were used for the control of Acacia dealbata, namely cutting and subsequent application of glyphosate, which is a systemic broad-spectrum herbicide that has shown some effectiveness with this species. However, this chemical product also presents environmental and contamination problems, so its use is being avoided on a large scale. For this reason, the use of techniques that are less aggressive to the environment, such as peeling, has been an alternative that shows interesting success rates, despite the time of performance being quite long.
During the same procedure, slices were cut from the base of the tree trunks, so that their age could be determined by counting the growth rings. From this, it was possible to verify that the oldest trees corresponded to the period following the fires that occurred in the 1990–1998 interval. It was noted that there is a hiatus in the occurrence of trees, in addition to a few specimens precisely from the period immediately after the last fire in 1998, that only reappeared after 2005—that is, the fire that occurred in 1998 eliminated the cover of pine, eucalyptus, and other native hardwoods, leaving space for the growth of acacias, which once again had an open path by another fire in 2005, which was the last one to hit the area under study, although others have occurred in the vicinity.
5. Conclusions
The Casal do Rei region, located in the Serra da Estrela Natural Park, presents a problem common to many other regions of the country due to the proliferation of invasive species such as Acacia dealbata, which already stands out for its large number of occurrences and area, and also other species such as Acacia melanoxylon, Acacia longifolia, and Hackea sericea. The development of the region’s forest cover, which developed over thousands of years as a result of successive adaptations to the climate and its transformations, gave rise to a set of habitats occupied by native species, among which are the so-called “Laurus nobilis arborescent forests”, which is a priority habitat for conservation (5230×pt2) recognized by the European Community through the Habitats Directive (92/43/EEC). Despite successive changes in the typology of vegetation cover, observed mainly throughout the 20th century with the intensive planting of species such as Pinus pinaster, and more recently of Eucalyptus globulus, extensive patches of species of the genus Quercus continued to persist, but also patches of other species that were on the edge of water courses or in niches in more protected areas.
Currently, especially with the increasingly frequent occurrence of rural fires that have swept over almost the entire area of the Natural Park in the last 30 years, the destruction of the native or traditional forest cover has given rise to the emergence in force of invasive species, which are slowly occupying the territory, replacing the remaining species. Thus, there is a marked decrease in biodiversity, while, due to the large amount of dense biomass that these species are able to develop, the risk of occurrence of rural fires increases. This situation creates a cycle of species selection by fire, since the new fires open space for these species, specifically, heliophiles and pyrophytes, to take advantage of every opportunity to conquer new spaces.
Control programs for invasive species are fundamental for the maintenance of ecosystems, and all methods are valid to achieve this. However, given the increasing number of invasive species, as well as the area occupied, it is urgent to develop integrated plans for eradication/control in order to avoid dispersion to areas not yet reached through the creation of rigid and intensive protocols in order to create permanent pressure on the populations of invasive species to reduce their vigor and dispersion capacity.
Author Contributions
Conceptualization, L.J.R.N. and C.I.R.M.; methodology, L.J.R.N., N.M.C.A.R., and C.J.P.G.; validation, L.J.R.N., M.A.M.R., and C.J.P.G.; formal analysis, L.J.R.N., C.I.R.M., and M.A.M.R.; investigation, L.J.R.N., M.A.M.R., C.I.R.M., C.J.P.G., and N.M.C.A.R.; resources, L.J.R.N.; data curation, L.J.R.N., M.A.M.R., and C.I.R.M.; writing—original draft preparation, L.J.R.N., M.A.M.R., and C.I.R.M.; writing—review and editing, L.J.R.N., M.A.M.R., C.I.R.M., C.J.P.G., and N.M.C.A.R.; supervision, N.M.C.A.R. and C.J.P.G.; project administration, N.M.C.A.R. and C.J.P.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors would like to thank the group of forest sappers from Manteigas for their support in opening access to the site under study, and the União de Freguesias de Vide e Cabeça for their support during the entire investigation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Xu, Z.; Jiang, Y.; Zhou, G. Nitrogen cycles in terrestrial ecosystems: Climate change impacts and mitigation. Environ. Rev. 2016, 24, 132–143. [Google Scholar] [CrossRef]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef] [PubMed]
- Catullo, R.A.; Llewelyn, J.; Phillips, B.L.; Moritz, C.C. The potential for rapid evolution under anthropogenic climate change. Curr. Biol. 2019, 29, R996–R1007. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.D.; Friess, D.A.; Day, R.H.; MacKenzie, R.A. Impacts of climate change on mangrove ecosystems: A region by region overview. Ecosyst. Health Sustain. 2016, 2, e01211. [Google Scholar] [CrossRef]
- Saxe, H.; Cannell, M.G.; Johnsen, Ø.; Ryan, M.G.; Vourlitis, G. Tree and forest functioning in response to global warming. New Phytol. 2001, 149, 369–399. [Google Scholar] [CrossRef]
- Funk, C.; Harrison, L.; Alexander, L.; Peterson, P.; Behrangi, A.; Husak, G. Exploring trends in wet-season precipitation and drought indices in wet, humid and dry regions. Environ. Res. Lett. 2019, 14, 115002. [Google Scholar] [CrossRef]
- Ciceu, A.; Popa, I.; Leca, S.; Pitar, D.; Chivulescu, S.; Badea, O. Climate change effects on tree growth from Romanian forest monitoring Level II plots. Sci. Total Environ. 2020, 698, 134129. [Google Scholar] [CrossRef]
- Runting, R.K.; Bryan, B.A.; Dee, L.E.; Maseyk, F.J.; Mandle, L.; Hamel, P.; Wilson, K.A.; Yetka, K.; Possingham, H.P.; Rhodes, J.R. Incorporating climate change into ecosystem service assessments and decisions: A review. Glob. Chang. Biol. 2017, 23, 28–41. [Google Scholar] [CrossRef]
- Williamson, C.E.; Neale, P.J.; Hylander, S.; Rose, K.C.; Figueroa, F.L.; Robinson, S.A.; Häder, D.-P.; Wängberg, S.-Å.; Worrest, R.C. The interactive effects of stratospheric ozone depletion, UV radiation, and climate change on aquatic ecosystems. Photochem. Photobiol. Sci. 2019, 18, 717–746. [Google Scholar] [CrossRef]
- Greaver, T.; Clark, C.; Compton, J.; Vallano, D.; Talhelm, A.; Weaver, C.; Band, L.; Baron, J.S.; Davidson, E.; Tague, C. Key ecological responses to nitrogen are altered by climate change. Nat. Clim. Chang. 2016, 6, 836–843. [Google Scholar] [CrossRef]
- Thom, D.; Rammer, W.; Seidl, R. Disturbances catalyze the adaptation of forest ecosystems to changing climate conditions. Glob. Chang. Biol. 2017, 23, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Macfadyen, S.; McDonald, G.; Hill, M.P. From species distributions to climate change adaptation: Knowledge gaps in managing invertebrate pests in broad-acre grain crops. Agric. Ecosyst. Environ. 2018, 253, 208–219. [Google Scholar] [CrossRef]
- Millington, R.; Cox, P.M.; Moore, J.R.; Yvon-Durocher, G. Modelling ecosystem adaptation and dangerous rates of global warming. Emerg. Top. Life Sci. 2019, 3, 221–231. [Google Scholar]
- Prober, S.M.; Potts, B.M.; Bailey, T.; Byrne, M.; Dillon, S.; Harrison, P.A.; Hoffmann, A.A.; Jordan, R.; McLean, E.H.; Steane, D.A. Climate adaptation and ecological restoration in eucalypts. Proc. R. Soc. Vic. 2016, 128, 40–53. [Google Scholar] [CrossRef]
- Razgour, O.; Forester, B.; Taggart, J.B.; Bekaert, M.; Juste, J.; Ibáñez, C.; Puechmaille, S.J.; Novella-Fernandez, R.; Alberdi, A.; Manel, S. Considering adaptive genetic variation in climate change vulnerability assessment reduces species range loss projections. Proc. Natl. Acad. Sci. USA 2019, 116, 10418–10423. [Google Scholar] [CrossRef]
- Batllori, E.; De Cáceres, M.; Brotons, L.; Ackerly, D.D.; Moritz, M.A.; Lloret, F. Compound fire-drought regimes promote ecosystem transitions in Mediterranean ecosystems. J. Ecol. 2019, 107, 1187–1198. [Google Scholar] [CrossRef]
- Paraskevopoulou, A.T.; Nektarios, P.A.; Kotsiris, G. Post-fire attitudes and perceptions of people towards the landscape character and development in the rural Peloponnese, a case study of the traditional village of Leontari, Arcadia, Greece. J. Environ. Manag. 2019, 241, 567–574. [Google Scholar] [CrossRef]
- Wolpert, F.; Quintas-Soriano, C.; Plieninger, T. Exploring land-use histories of tree-crop landscapes: A cross-site comparison in the Mediterranean Basin. Sustain. Sci. 2020, 1–17. [Google Scholar] [CrossRef]
- Batllori, E.; De Cáceres, M.; Brotons, L.; Ackerly, D.D.; Moritz, M.A.; Lloret, F. Cumulative effects of fire and drought in Mediterranean ecosystems. Ecosphere 2017, 8, e01906. [Google Scholar] [CrossRef]
- Bazan, G.; Barba, A.C.; Rotolo, A.; Marino, P. Geobotanical approach to detect land-use change of a Mediterranean landscape: A case study in Central-Western Sicily. GeoJournal 2019, 84, 795–811. [Google Scholar] [CrossRef]
- Ojeda, F.; Midgley, J.; Pauw, A.; Lavola, A.; Casimiro-Soriguer, R.; Hattas, D.; Segarra-Moragues, J.G.; Julkunen-Tiitto, R. Flower colour divergence is associated with post-fire regeneration dimorphism in the fynbos heath Erica coccinea subsp. coccinea (Ericaceae). Evol. Ecol. 2019, 33, 345–367. [Google Scholar] [CrossRef]
- Kane, J.M.; Varner, J.M.; Saunders, M.R. Resurrecting the Lost Flames of American Chestnut. Ecosystems 2019, 22, 995–1006. [Google Scholar] [CrossRef]
- Soler, M.; Úbeda, X. Evaluation of fire severity via analysis of photosynthetic pigments: Oak, eucalyptus and cork oak leaves in a Mediterranean forest. J. Environ. Manag. 2018, 206, 65–68. [Google Scholar] [CrossRef]
- Matusick, G.; Ruthrof, K.X.; Fontaine, J.B.; Hardy, G.E.S.J. Eucalyptus forest shows low structural resistance and resilience to climate change-type drought. J. Veg. Sci. 2016, 27, 493–503. [Google Scholar] [CrossRef]
- Franco-Manchón, I.; Salo, K.; Oria-de-Rueda, J.A.; Bonet, J.A.; Martín-Pinto, P. Are wildfires a threat to fungi in European pinus forests? A case study of Boreal and Mediterranean forests. Forests 2019, 10, 309. [Google Scholar] [CrossRef]
- Gea-Izquierdo, G.; Férriz, M.; García-Garrido, S.; Aguín, O.; Elvira-Recuenco, M.; Hernandez-Escribano, L.; Martin-Benito, D.; Raposo, R. Synergistic abiotic and biotic stressors explain widespread decline of Pinus pinaster in a mixed forest. Sci. Total Environ. 2019, 685, 963–975. [Google Scholar] [CrossRef]
- Dodet, M.; Collet, C. When should exotic forest plantation tree species be considered as an invasive threat and how should we treat them? Biol. Invasions 2012, 14, 1765–1778. [Google Scholar] [CrossRef]
- Moser, W.K.; Barnard, E.L.; Billings, R.F.; Crocker, S.J.; Dix, M.E.; Gray, A.N.; Ice, G.G.; Kim, M.-S.; Reid, R.; Rodman, S.U. Impacts of nonnative invasive species on US forests and recommendations for policy and management. J. For. 2009, 107, 320–327. [Google Scholar]
- Martins, F.; Alegria, C.; Gil, A. Mapping invasive alien Acacia dealbata Link using ASTER multispectral imagery: A case study in central-eastern of Portugal. For. Syst. 2016, 25, 13. [Google Scholar] [CrossRef]
- Coumou, D.; Rahmstorf, S. A decade of weather extremes. Nat. Clim. Chang. 2012, 2, 491–496. [Google Scholar] [CrossRef]
- Alexander, L.V.; Zhang, X.; Peterson, T.C.; Caesar, J.; Gleason, B.; Klein Tank, A.; Haylock, M.; Collins, D.; Trewin, B.; Rahimzadeh, F. Global observed changes in daily climate extremes of temperature and precipitation. J. Geophys. Res. Space Phys. 2006, 111. [Google Scholar] [CrossRef]
- Kharin, V.V.; Zwiers, F.W.; Zhang, X.; Hegerl, G.C. Changes in temperature and precipitation extremes in the IPCC ensemble of global coupled model simulations. J. Clim. 2007, 20, 1419–1444. [Google Scholar] [CrossRef]
- Feio, M.; Norris, R.; Graça, M.; Nichols, S. Water quality assessment of Portuguese streams: Regional or national predictive models? Ecol. Indic. 2009, 9, 791–806. [Google Scholar] [CrossRef]
- Migoń, P.; Vieira, G. Granite geomorphology and its geological controls, Serra da Estrela, Portugal. Geomorphology 2014, 226, 1–14. [Google Scholar] [CrossRef]
- Martins, A.N. Sustainable Design, Cultural Landscapes and Heritage Parks; the Case of the Mondego River. Sustain. Dev. 2016, 24, 330–343. [Google Scholar] [CrossRef]
- Ferreira, V.; Graça, M.A.; Feio, M.J.; Mieiro, C. Water quality in the Mondego river basin: Pollution and habitat heterogeneity. Limnetica 2004, 23, 295–306. [Google Scholar]
- Martins, A.A.; Cunha, P.P.; Rosina, P.; Osterbeek, L.; Cura, S.; Grimaldi, S.; Gomes, J.; Buylaert, J.-P.; Murray, A.S.; Matos, J. Geoarchaeology of Pleistocene open-air sites in the Vila Nova da Barquinha-Santa Cita area (Lower Tejo River basin, central Portugal). Proc. Geol. Assoc. 2010, 121, 128–140. [Google Scholar] [CrossRef]
- Mora, C. A synthetic map of the climatopes of the Serra da Estrela (Portugal). J. Maps 2010, 6, 591–608. [Google Scholar] [CrossRef]
- Vieira, G. Combined numerical and geomorphological reconstruction of the Serra da Estrela plateau icefield, Portugal. Geomorphology 2008, 97, 190–207. [Google Scholar] [CrossRef]
- Vieira, G.T.; Mora, C.; Ramos, M. Ground temperature regimes and geomorphological implications in a Mediterranean mountain (Serra da Estrela, Portugal). Geomorphology 2003, 52, 57–72. [Google Scholar] [CrossRef]
- Palacios, D.; Andrés, N.; Marcos, J.; Vázquez-Selem, L. Maximum glacial advance and deglaciation of the Pinar Valley (Sierra de Gredos, Central Spain) and its significance in the Mediterranean context. Geomorphology 2012, 177, 51–61. [Google Scholar] [CrossRef]
- Marques, J.E.; Marques, J.M.; Chaminé, H.I.; Carreira, P.M.; Fonseca, P.E.; Santos, F.A.M.; Moura, R.; Samper, J.; Pisani, B.; Teixeira, J. Conceptualizing a mountain hydrogeologic system by using an integrated groundwater assessment (Serra da Estrela, Central Portugal): A review. Geosci. J. 2013, 17, 371–386. [Google Scholar] [CrossRef]
- Sant’Ovaia, H.; Olivier, P.; Ferreira, N.; Noronha, F.; Leblanc, D. Magmatic structures and kinematics emplacement of the Variscan granites from Central Portugal (Serra da Estrela and Castro Daire areas). J. Struct. Geol. 2010, 32, 1450–1465. [Google Scholar] [CrossRef]
- Sá, A.A.; Gutiérrez-Marco, J.C.; Meireles, C.; García-Bellido, D.C.; Rábano, I. A revised correlation of Lower Ordovician sedimentary rocks in the Central Iberian Zone (Portugal and Spain). In STRATI 2013; Springer: Berlin/Heidelberg, Germany, 2014; pp. 441–445. [Google Scholar]
- Meireles, C.; Castro, P.; Ferreira, N. Evidências cartográficas, litoestratigráficas e estruturais sobre a presença de discordância cadomiana intra Grupo das Beiras. Comunicações Geológicas 2014, 101, 495–498. [Google Scholar]
- Marques, J.E.; Samper, J.; Pisani, B.; Alvares, D.; Carvalho, J.M.; Chaminé, H.I.; Marques, J.; Vieira, G.; Mora, C.; Borges, F.S. Evaluation of water resources in a high-mountain basin in Serra da Estrela, Central Portugal, using a semi-distributed hydrological model. Environ. Earth Sci. 2011, 62, 1219–1234. [Google Scholar] [CrossRef]
- Marques, J.E.; Duarte, J.M.; Constantino, A.T.; Martins, A.A.; Aguiar, C.; Rocha, F.T.; Inácio, M.; Marques, J.M.; Chaminé, H.I.; Teixeira, J. Vadose zone characterisation of a hydrogeologic system in a mountain region: Serra da Estrela case study (Central Portugal). In Aquifer Systems Management: Darcy’s Legacy in a World of Impending Water Shortage: Selected Papers on Hydrogeology; CRC Press: Boca Raton, FL, USA, 2007; p. 207. [Google Scholar]
- Connor, S.E.; Araújo, J.; van der Knaap, W.O.; van Leeuwen, J.F. A long-term perspective on biomass burning in the Serra da Estrela, Portugal. Quat. Sci. Rev. 2012, 55, 114–124. [Google Scholar] [CrossRef]
- Van der Knaap, W.; Van Leeuwen, J. Holocene vegetation succession and degradation as responses to climatic change and human activity in the Serra de Estrela, Portugal. Rev. Palaeobot. Palynol. 1995, 89, 153–211. [Google Scholar] [CrossRef]
- Meireles, C.; Pinto-Gomes, C.; Cano, E. Approach to climatophilous vegetation series of Serra da Estrela (Portugal) Approche des séries de végétation climatophiles de la Serra da Estrela (Portugal). Acta Bot. Gall. 2012, 159, 283–287. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Belmonte, D.; Cantó, P.; Fernández-González, F.; Fuente, V.; Moreno, J.; Sánchez-Mata, D.; GARCÍA SANCHO, L. Piornales, enebrales y pinares oromediterráneos (Pino-Cytision oromediterranei) en el Sistema Central. Lazaroa 1987, 7, 93–124. [Google Scholar]
- Ramos, C.; Ramos-Pereira, A. Landscapes of Portugal: Paleogeographic Evolution, Tectonics and Geomorphology. In Landscapes and Landforms of Portugal; Springer: Berlin/Heidelberg, Germany, 2020; pp. 3–31. [Google Scholar]
- Mugica, F.F.; Antón, M.G.; Ollero, H.S. Vegetation dynamics and human impact in the Sierra de Guadarrama, Central System, Spain. Holocene 1998, 8, 69–82. [Google Scholar] [CrossRef]
- Rubiales, J.M.; García-Amorena, I.; Hernández, L.; Génova, M.; Martínez, F.; Manzaneque, F.G.; Morla, C. Late Quaternary dynamics of pinewoods in the Iberian Mountains. Quat. Sci. Rev. 2010, 162, 476–491. [Google Scholar] [CrossRef]
- Rubiales, J.; García-Amorena, I.; Génova, M.; Manzaneque, F.G.; Morla, C. The Holocene history of highland pine forests in a submediterranean mountain: The case of Gredos mountain range (Iberian Central range, Spain). Quat. Sci. Rev. 2007, 26, 1759–1770. [Google Scholar] [CrossRef]
- Figueiral, I.; Carcaillet, C. A review of Late Pleistocene and Holocene biogeography of highland Mediterranean pines (Pinus type sylvestris) in Portugal, based on wood charcoal. Quat. Sci. Rev. 2005, 24, 2466–2476. [Google Scholar] [CrossRef]
- Rubiales, J.M.; Génova, M. Late Holocene pinewoods persistence in the Gredos Mountains (central Spain) inferred from extensive megafossil evidence. Quat. Rev. 2015, 84, 12–20. [Google Scholar] [CrossRef]
- Nunes, A.N. Regional variability and driving forces behind forest fires in Portugal an overview of the last three decades (1980–2009). Appl. Geogr. 2012, 34, 576–586. [Google Scholar] [CrossRef]
- Nunes, L.J.R.; Raposo, M.; Meireles, C.I.R.; Gomes, C.J.P.; Nuno, M.C.; Almeida, R. The propagation of invasive species as a consequence of rural Fires: A case study in Casal do Rei (Seia). In Proceedings of the IV PhD Students Meeting in Environmental and Agriculture, Évora, Portugal, 11–12 November 2019; p. 62. [Google Scholar]
- Ferreira-Leite, F.; Lourenço, L.; Bento-Gonçalves, A. Large forest fires in mainland Portugal, brief characterization. Méditerranée. Revue géographique des pays méditerranéens/J. Mediterr. Geogr. 2013, 121, 53–65. [Google Scholar] [CrossRef]
- Nunes, A. Incêndios florestais no Parque Natural da Serra da Estrela: Quadro físico-geográfico da sua ocorrência. Cadernos de Geografia 2001, 20, 93–112. [Google Scholar] [CrossRef]
- Directive, H. Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Union 1992, 206, 7–50. [Google Scholar]
- Fernández-García, V.; Fulé, P.Z.; Marcos, E.; Calvo, L. The role of fire frequency and severity on the regeneration of Mediterranean serotinous pines under different environmental conditions. For. Ecol. Manag. 2019, 444, 59–68. [Google Scholar] [CrossRef]
- Jansen, J.; Diemont, H. Implementing Natura 2000 in farmed landscapes: The Serra da Estrela, Portugal. Ekológia 2011, 30, 199–215. [Google Scholar] [CrossRef]
- Chirino, E.; Vilagrosa, A.; Cortina, J.; Valdecantos, A.; Fuentes, D.; Trubat, R.; Luis, V.C.; Puértolas, J.; Bautista, S.; Baeza, J. Ecological restoration in degraded drylands: The need to improve the seedling quality and site conditions in the field. In Forest Management; Nova Publisher: New York, NY, USA, 2009; pp. 85–158. [Google Scholar]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).