1. Introduction
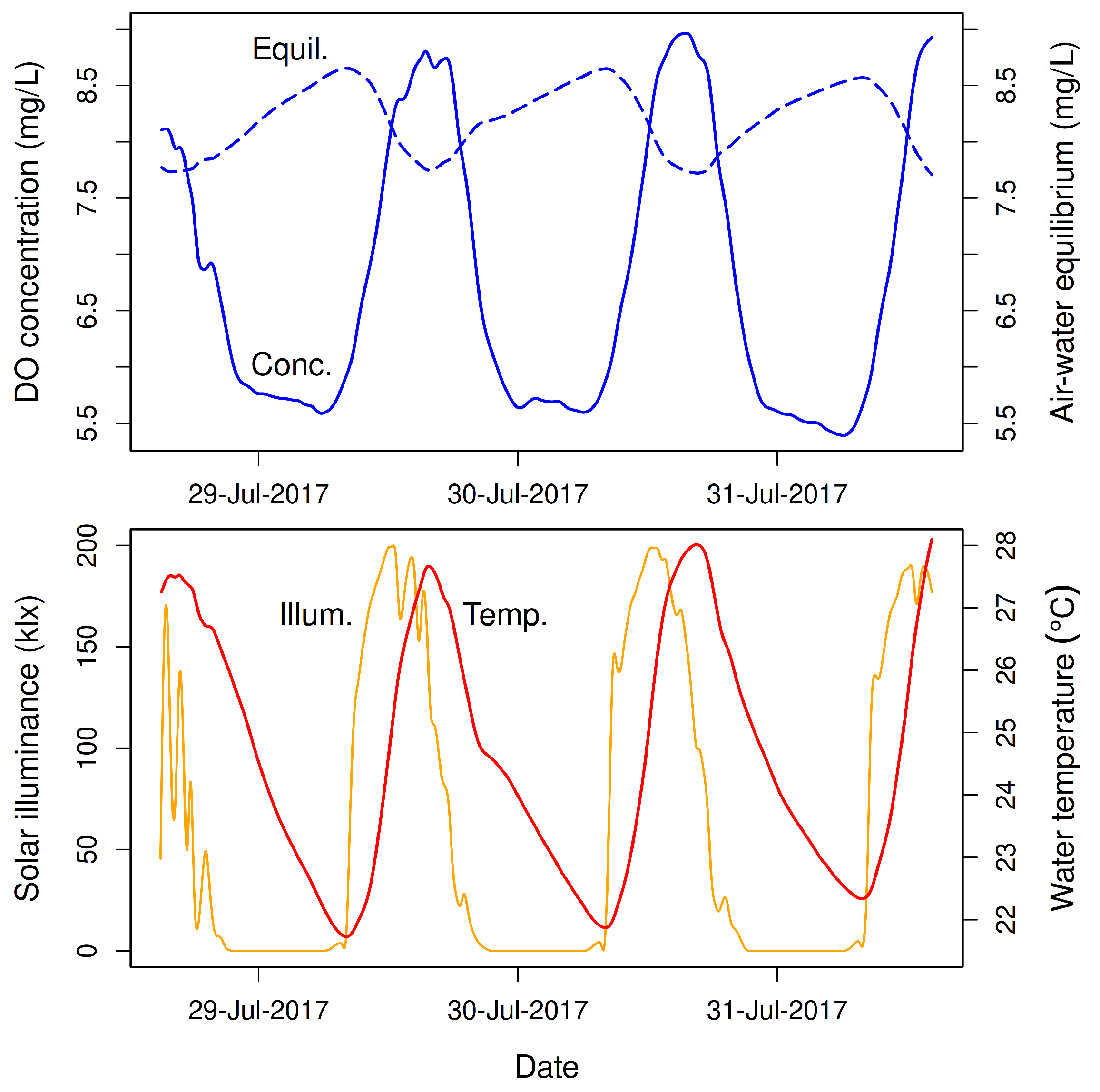
One of the most striking and predictable phenomena in healthy streams is the daily cycle of dissolved oxygen (DO) concentration that occurs during warm months of the year (
Figure 1). This cycle is caused mainly by the combined metabolism of aerobic bacteria, eukaryotic algae, and submersed aquatic macrophytes. During daytime, oxygenic photosynthesis by photoautotrophs produces oxygen at a higher rate than it is consumed by aerobic respiration of photoautotrophs and heterotrophs (
Figure 2). The DO concentration in the stream therefore increases sharply, often exceeding the gas-exchange equilibrium (“saturation”). During nightime, aerobic respiration by photoautotrophs and heterotrophs in the absence of photosynthesis causes a rapid decline in DO concentration, driving it well below the gas-exchange equilibrium. And so, as Odum and Hoskin [
1] put it, streams “like great creatures breathe in and out” over each diel cycle.
The pronounced diel cycle of DO concentration in a stream, and the associated cycle of solar radiation that drives it, are the basis for a common method of estimating the rates of oxygen production and consumption by stream communities. This method employs times series of DO concentration, water temperature, and solar radiation, along with a process-based model of DO dynamics, to estimate rates of oxygenic photosynthesis and aerobic respiration. These rates are the main components of stream metabolism.
Various difficulties in the estimation process are created by the facts that stream water is continually flowing, and stream habitat and community structure typically exhibit marked longitudinal heterogeneity. In particular, these stream properties complicate the process of determining which stream reach and associated community a measured metabolic rate applies to. As flowing water enters a given stream reach, its DO concentration mainly reflects habitat and community structure upstream of the focal reach. As the water flows through the focal reach, its DO concentration changes and approaches a value characteristic of habitat and community structure in the focal reach. Thus, there is a transition zone in the initial portion of the focal reach, within which the DO concentration changes from being characteristic of the upstream reach to being characteristic of the focal reach. In order to obtain metabolism estimates that reflect the habitat and community structure of the focal reach, it is necessary to know the approximate length of this transition zone and to base the estimates on time series of DO concentration, solar radiation, and other predictor variables acquired downstream of the transition zone but still within the focal reach. A simple traditional estimate of transition-zone length has been widely employed by stream ecologists since the early 2000s (see
Section 3), but we will argue that it fails to account for most of the key processes responsible for changes in DO concentration within the transition zone and therefore is inadequate.
The main goal of the present paper is to propose a new practical estimate of the length of the DO transition zone. In order to achieve this goal, it is necessary to extend the classical model of stream DO dynamics in several ways. The problem we address is of fundamental importance in stream ecology, but some of the empirical and theoretical background information necessary for understanding the problem and the approaches we will use to solve it is sufficiently specialized that many readers may be unfamiliar with it. Therefore, to make our paper accessible to as wide an audience as possible, we include a significant amount of background information that is necessary for understanding the model extensions we propose and their biological rationale.
The paper is organized as follows.
Section 2 presents background empirical information on stream metabolism that is required for understanding the remainder of the paper; it can be skipped or skimmed by readers who are already familiar with this information.
Section 3 presents the traditional estimate of DO transition-zone length.
Section 4 presents an intuitive argument that reveals the main processes responsible for the changes in DO concentration that occur after water flows into a focal stream reach, only one of which is accounted for by the traditional estimate of transition-zone length.
Section 5 outlines the classical model of stream DO dynamics and can be skipped or skimmed by readers who are already familiar with this model.
Section 6 presents an extension of the classical model of stream DO dynamics, which distinguishes three types of DO in a focal stream reach (old DO that entered the stream upstream of the focal reach, new DO that entered the focal reach from the atmosphere, and new DO that was produced within the focal reach by photosynthesis), then uses the extended model to derive a new estimate of transition-zone length that accounts for all major processes responsible for changes in DO concentration within the transition zone.
Section 7 assesses and compares the traditional and proposed estimates of transition-zone length with exact values computed using a more-realistic model of DO dynamics that includes an energy balance and transport model for water temperature, whose dynamics drive temporal variation in temperature-dependent process rates (photosynthesis, respiration, and exchange of oxygen with the atmosphere); this section can be skipped on a first reading.
Section 8 shows how the traditional estimate of transition-zone length can be derived from the proposed new estimate, providing insight into conditions under which the two estimates will be similar; this section, too, can be skipped on a first reading.
Section 9 compares the values of the traditional and proposed estimates of transition-zone length calculated using published data for several streams in eastern New York state (USA).
Section 10 presents a second novel method of estimating the transition-zone length (based on the DO residence-time distribution instead of concentrations of the three types of old and new DO), which yields the same proposed estimate as the first method; it can be skipped on a first reading. Finally,
Section 11 discusses the results, and
Section 12 briefly summarizes the main takeaways from our study.
3. The Traditional Estimate of Transition Zone Length
Grace and Imberger [
26] and Demars et al. [
30] briefly treat an important theoretical question that is closely related to the four problems mentioned in the previous section: within what distance upstream did most of the DO measured by a sonde enter the flowing water? One of the main goals of the present paper is to reconsider this question and the answer these authors suggest. We will begin by addressing the question in the following, closely related form: how far down from the upstream boundary of a reasonably uniform stream reach does the DO concentration first become dominated by DO that entered the flowing water within the focal reach. This is an important question in ecological studies of stream metabolism that employ the FWDO method with 1-station monitoring, because the answer tells us which reach and associated habitat and community types the metabolism estimates apply to.
For example, if 95% of the DO in water flowing past a sonde at a particular location in the focal reach was already in the water when it entered the reach at the upstream boundary, then the measured DO concentrations mainly reflect habitat and community types upstream of the focal reach rather than within it. But if only 5% of the DO in water flowing past the sonde was already present in the water when it entered the focal reach, then the measured DO concentrations mainly reflect the stream habitat and community between the upstream boundary of the reach and the sonde, all of which lies within the focal reach.
Several authors have drawn attention to this problem in the past (e.g., [
21,
22,
26,
29,
30,
31,
48]). These authors argue that 95% of the DO measured by a sonde was contributed by stream processes within an upstream distance
given by
where
[dimensions: Length Time
−1] is mean current velocity and
K [Time
−1] is the atmospheric exchange coefficient (see below), both assumed constant. For example, in the notation of Equation (
1), Grace and Imberger [
26] state that “the usual one-station method integrates stream metabolism for a distance approximated by
” (p. 26) and “the use of only one station means that calculations are based on water integrated from only a vaguely known distance upstream (≈
)” (p. 71). Similarly, Reichert et al. [
21] state that “the downstream dissolved oxygen concentration is primarily determined by processes within a river reach of length
” (p. 4), Roley et al. [
48] state that they “estimated the upstream distance integrated with the 1-station method as
” (p. 1045), and Demars et al. [
30] state that “the 95% footprint (length scale
) of the oxygen sensor is generally calculated as
” (p. 361).
In the conceptual framework where a stream is viewed as a longitudinal sequence of discrete reaches with different dominant habitat and community types and corresponding metabolic rates, the key issue is whether a sonde at a given stream location (and hence the time series of DO concentration, water temperature, and so on it acquires) does or does not lie within the transition zone between the neighboring upstream reach and the focal reach. There are two ways of thinking about this issue: we can center our thinking on the sonde and look upstream toward the boundary of the focal reach, or we can center our thinking on the upstream boundary of the focal reach and look downstream toward the sonde. With the first approach, we are thinking about how far upstream the reach monitored by the sonde extends and wish to know whether this monitored reach lies entirely within the focal reach. With the second approach, we are thinking about how far down from the upstream boundary of the focal reach the DO transition zone extends and wish to know whether the sonde lies downstream of this zone but still within the focal reach. Depending on which view we adopt, the traditional estimate represents the length of the monitored reach or the length of the transition zone.
In most of this paper, we will adopt the downstream-looking view and develop the theory in terms of the length of the transition zone. Our main purpose is to argue that the traditional estimate of the length of the transition zone given by Equation (
1) is not defensible and to propose two closely related alternative approaches for estimating this distance that we believe are more appropriate. To the best of our knowledge, our paper is the first to propose an alternative to the traditional estimate of the transition-zone length, which has been widely accepted by stream ecologists since Grace and Imberger [
26] published their manual on methods for measuring stream metabolism in 2006.
4. An Intuitive Characterization of the Transition Zone
Before developing the theory required for proposing an alternative way to estimate the length of the transition zone, we briefly consider the problem intuitively in order to identify the main biological and physical processes that must be addressed. We will argue in subsequent sections that the traditional estimate
given by Equation (
1) does not adequately account for all of these.
Dating back to the original Streeter-Phelps model in the engineering literature, nearly all published studies of stream metabolism have employed a plug-flow transport model in which transverse slices of water in the stream channel move downstream intact, with no significant longitudinal dispersion or warping. Each slice is assumed to be vertically and tranversely homogeneous with respect to temperature and solute concentrations (implying instantaneous vertical and transverse mixing), so the model only addresses longitudinal patterns in these properties. It is intended to be applied under field conditions where lateral inflow and outflow of water are negligible, so these processes usually are not included (but can be [
49,
50]). The plug-flow assumption greatly simplifies the theory and numerical calculations used to estimate components of stream metabolism, and also makes it easier to think about the problem intuitively. Moreover, real-world applications of the Streeter-Phelps model in many water-quality studies have demonstrated that, despite their rather strong simplifying assumptions, plug-flow models can provide useful characterizations of longitudinal patterns in stream reaches that are reasonably short, relative to the mean current velocity and the actual rate of longitudinal dispersion.
Suppose, then, that we have identified a short stream reach of reasonably uniform habitat and have chosen specific locations as its upstream and downstream boundaries. Let
x denote distance downstream from the upstream boundary. On this scale, the location of the upstream boundary is
, and we may denote the location of the downstream boundary by
(
Figure 4). We place a sonde at location
within the focal reach, with
. As each slice of water passes the sonde, its DO concentration is measured. The question is: where did this DO enter the slice?
When any given slice enters the focal reach at
, it is already carrying DO, all of which was acquired upstream. After the slice enters the focal reach, a portion of this initial DO will be lost to the atmosphere by the time the slice reaches the sonde at
, and another portion will be lost to aerobic respiration. Thus, only a fraction of the DO the slice contained when it entered the focal reach will remain when it reaches the sonde. We will call this residual portion “old DO” (labeled “1” in
Figure 4), since it was produced by stream processes upstream of
, before the slice entered the focal reach.
But this old DO is only one portion of the total DO that will be present in the slice when it reaches the sonde. Two additional portions will enter the slice as it travels from
to
: a portion that enters from the atmosphere (labeled “2” in
Figure 4) and, during daytime only, a portion that is produced by oxygenic photosynthesis (labeled “3” in
Figure 4). We will call these portions “new DO”, since they entered the slice after it entered the focal reach. Clearly, the greater the distance between
and
, the smaller the proportion of old DO will be at
and the greater the proportion of new DO (left and middle panels of
Figure 4).
Let the residual concentration of old DO when a slice reaches the sonde at be , let the concentrations of new DO that entered the slice from the atmosphere and from oxygenic photosynthesis during transit between and be and respectively, and let be the total DO concentration at . The proportion of total DO measured in the slice at that was already present when it passed is simply the proportion of old DO at , which is . We argue that this is the appropriate measure of the contribution of stream processes upstream of the focal reach to the DO concentration measured by the sonde.
The complementary proportion of total DO measured in the slice at
that entered the slice during transit between
and
is the proportion of new DO at
, which is
. We argue that this proportion is the appropriate measure of the contribution of stream processes in the reach between locations
and
to the DO concentration measured by the sonde. Based on this argument, the length of the transition zone should be chosen as the travel distance
x from the upstream boundary of the reach at which the proportion
of new DO first increases to 0.95, or equivalently, the proportion
of old DO first decreases to 0.05. But as we show below, this is not the measure that underlies the traditional estimate
in Equation (
1). That the estimate in Equation (
1) is suspicious is already evident from the fact that, based on the above intuitive account and
Figure 4, the rates of photosynthetic production and respiratory consumption of DO clearly are important in determining how rapidly the proportion of old DO declines with distance below upstream reach boundary
and how rapidly the proportion of new DO increases, but neither rate appears in Equation (
1).
5. The Model of Stream DO Dynamics Underlying the FWDO Method
The first step in translating the intuitive argument of
Section 4 into a quantitative estimate of transition-zone length is to specify a model of stream DO dynamics. The model we will use is an extension of the standard model on which most ecological studies of stream metabolism employing the FWDO method have been based since H. T. Odum’s seminal paper [
5] was published in 1956. We outline the standard model in this section, then present an extended version in
Section 6 that we use to deduce a new estimate of transition-zone length. Readers who are already familiar with the standard model may wish to skip or skim this section.
The standard model of stream DO dynamics was designed to avoid the extreme complexity of detailed mechanistic representations of stream hydrodynamics and the dynamics of biological community structure and function by focusing on a short stream reach of reasonably uniform habitat during a short period of time (e.g., one diel cycle), and by making several simplifying assumptions that are rather strong but provide useful approximations in this context. The main assumptions are as follows:
Stream flow, cross-section geometry, and mean current velocity are approximately constant in time and space within the focal reach
The water column is well mixed vertically and transversely
Longitudinal mixing is negligible
There is no significant lateral inflow or outflow of water
There is no significant exchange of water and solutes with the hyporheic zone
The total rates at which oxygen is produced and consumed by organisms suspended in and transported by the flowing water are negligible compared to the total rates for organisms attached to the stream bed
The rates of oxygen production and consumption by benthic organisms are reasonably uniform within the focal stream reach
The only physical variables whose dynamics must be monitored in order to adequately predict the rates at which DO is gained or lost via photosynthesis, respiration, and atmospheric exchange are solar irradiance and water temperature.
As noted above, these simplifying assumptions are intended to facilitate application of the model to short stream reaches of reasonably uniform habitat over short periods of time. Longer reaches and periods are addresed by applying the model separately to multiple short reaches and periods so the different sets of parameter estimates can account for spatial and temporal changes in stream flow, nutrient concentrations, autotrophic and heterotrophic biomass, and other factors that affect stream metabolism but are assumed constant in the model. This pragmatic procedure greatly reduces model complexity, the number of explicit functional forms for process rates that must be verified, and the number of parameters that must be estimated for each application of the model.
Given the above assumptions regarding stream flow and biological community structure and function, the usual generic assumptions of mass balance and mathematical continuity invoked in deriving continuity equations for mass transport in flowing water yield the following semilinear first-order hyperbolic partial differential equation:
subject to initial and boundary conditions
where
t [Time] denotes time,
x [Length] denotes longitudinal position or distance downstream from the upstream boundary of the reach at
,
[Mass Length
−3] denotes DO concentration at time
t and position
x throughout a thin transverse slice of the stream at
x,
[Length Time
−1] denotes mean current velocity,
,
, and
[Mass Length
−3 Time
−1] are functions denoting the instantaneous rates of oxygen production by oxygenic photosynthesis, oxygen consumption by aerobic respiration, and net uptake of oxygen from the atmosphere across the air-water interface, and
and
[Mass Length
−3] are functions specifying the initial DO concentration as a function of downstream distance
x, and the concentration at the upstream boundary
as a function of time
t. For brevity, we will refer to
,
, and
as simply the photosynthesis rate, respiration rate, and atmospheric exchange rate.
The atmospheric exchange rate in Equation (
2) typically is assumed to have the form
where
is the gas transfer velocity for oxygen [Length Time
−1],
H is the mean height (depth) of the water column [Length],
is the gas-exchange equilibrium DO concentration [Mass Length
−3], and
is the atmospheric exchange coefficient [Time
−1]. The functional significance of the gas-exchange equilibrium is better revealed by the alternative representation
where
is the concentration of oxygen in the atmosphere adjacent to the air-water interface and
is Ostwald’s concentration-based gas solubility coefficient for oxygen, which in streams depends mainly on water temperature and salinity [
51,
52].
Equation (
2) is a plug-flow (reaction-advection) model of the spatiotemporal dynamics of DO concentration in a stream reach. The left side of this equation represents the advective derivative of DO concentration with respect to time, with advective flux function
. The right side represents the DO source processes (photosynthesis, DO invasion) and sink processes (respiration, DO evasion).
The general solution of Equation (
2) encompasses two types of particular solutions: those which apply to slices of water that were already present in the focal stream reach at time
, and those which apply to slices that entered the focal reach at some time
. We are interested only in the latter. Consider, therefore, a moving slice of water that crosses upstream boundary
of the focal reach at time
and reaches the sonde at location
at time
(
Figure 5). As this slice travels downstream, its position will follow characteristic base curve
of Equation (
2) in the
plane. Along this curve, we have
. The left side of Equation (
2) then represents the advective derivative of
, and the partial differential equation reduces to an ordinary differential equation in the time domain,
where
,
,
,
, and
. This equation states that the instantaneous rate of change in DO concentration within a moving slice of water is given by the rate at which DO is produced by oxygenic photosynthesis (
), minus the rate at which it is consumed by aerobic respiration (
), plus the difference between the rates at which oxygen is taken up from the atmosphere (
) and lost to the atmosphere (
) across the air-water interface, as illustrated by
Figure 5.
Equation (
3) is subject to initial condition
, which specifies the DO concentration in the slice when it crossed upstream boundary
at time
and entered the focal reach. Our notation calls attention to parameter
, because it alone tells us which slice of water Equation (
3) applies to. Note that the implied value of
can be determined for any slice whose DO concentration is measured at
, because
and the values of
,
, and
will be known.
Solutions of Equation (
3) are functions of time and therefore address temporal patterns. In the present paper, however, we are mainly interested in spatial patterns. For this purpose, it is convenient to express Equation (
2) in the equivalent form,
The characteristic base curves for this equation are the same as for Equation (
2) but are now viewed as solutions of differential equation
, implying
. The left side of Equation (
4) represents the advective derivative of
, and the partial differential equation reduces to an ordinary differential equation in the one-dimensional longitudinal spatial domain,
where
and, for example,
. The initial condition for Equation (
5) is
. The biological interpretation of this equation is similar to that of Equation (
3), except that the rates are now rates with respect to transport distance instead of time. Rates
,
,
, and
on the right side of the equation again represent the time-rates of photosynthetic production, respiration, invasion, and evasion of oxygen (now as functions of downstream transport distance instead of time), but these are converted to distance-rates when divided by mean current velocity
.
7. Numerical Examples with Time-Varying Process Rates
In the previous section, we obtained a simple formula for the length of the transition zone by assuming the photosynthesis rate, respiration rate, atmospheric exchange coefficient, and gas-exchange equilibrium DO concentration are constant over a diel cycle instead of allowing them to vary, as they do in reality. But does the resulting formula for
provide a reasonable estimate of the actual length of the transition zone when these quantities vary over a diel cycle? We now address this question by specifying functional forms for the dependence of instantaneous rates
,
, and
and gas-exchange equilibrium DO concentration
on solar irradiance and water temperature. The resulting model does not appear to admit an explicit closed-form solution, so we use parameter estimates from a study by Zuidema [
2] to compute solutions of Equation (
9) and determine the values of
and
numerically. We then compare these numerical estimates with the corresponding simple estimates of the previous section, as well as with values of traditional estimate
given by Equation (
1). These results yield additional insight into the reasons the various estimates differ and the conditions under which they are likely to be similar. The main conclusion is that the proposed estimate of transition-zone length based on constant process rates often provides a useful approximation to the actual value when process rates are allowed to vary in a realistic way. Readers who are willing to tentatively accept this conclusion may wish to skip this section on a first reading.
We now provide numerical examples, based on the model in Equation (
9), in which the rate of DO production by photosynthesis, rate of DO consumption by community respiration, atmospheric exchange coefficient, and gas-exchange equilibrium DO concentration in a moving slice of water all exhibit diel variation. Variation in photosynthetic DO production is driven mainly by a diel cycle in photon flux from sunlight and is modulated by a diel cycle in water temperature that is driven in turn by solar radiation. Variation in respiration and atmospheric exchange rates and in the gas-exchange equilibrium is driven by the diel cycle in water temperature which, as just noted, is driven by the diel cycle in solar radiation. This model allows us to determine the exact length
of the transition zone for comparison with the constant-rate estimate given by Equation (
16) and the traditional estimate given by Equation (
1).
The functional forms employed for the rate
of DO production by photosynthesis, the rate
of DO consumption by community respiration, and the atmospheric exchange coefficient
in Equations (
8) and (
9) are based on rates at a standard temperature of 20 °C, with Berthelot adjustment to field temperature, and are as follows:
where
is a measure of solar radiation at time
t (assumed uniform throughout the focal stream reach),
,
, and
are the values of
,
, and
at 20 °C,
is water temperature (°C) at time
t (h) and location
x in a thin slice of moving water located distance
x (m) below the upstream reach boundary at
, and
,
, and
are Berthelot parameters used to adjust rates at 20 °C to rates at field temperature. The data set on which parameter values in this example are based employs solar illuminance [klx] as the measure of solar radiation (for sunlight, 1.0 klx illuminance ≈ 7.9 W/m
2 solar irradiance). Parameter values are estimates by Zuidema [
2] for Little Black Creek in July 2017 and are as follows:
mg L
−1 h
−1,
mg L
−1 h
−1,
h
−1,
(
),
(
), and
m h
−1 (5.5 cm s
−1).
The functional form employed for
is the standard Benson-Krause form (e.g., [
54]). Atmospheric pressure was set to 1.0 atm and salinity to 0.0 ‰, so variation in
is driven entirely by water temperature in this example. Solar illuminance at any time
is given by
where
is the diel maximum of solar illuminance,
is the value of
t at which the most recent sunrise occurred,
during daytime is the elapsed time since the most recent sunrise, and
is the duration of daytime at the current geographic location and calendar date.
The dynamics of water temperature and their relationship to solar illuminance were specified by a plug-flow model of thermal energy transport that is commonly used in studies of streams and small rivers (e.g., [
42,
55,
56]). In this modeling framework, the advected property is thermal energy (in joules), and both thermodynamic work and temperature-related changes in water density are ignored. Thus, the longitudinal flux of thermal energy at time
t and stream location
x is
[Energy Length
−2 Time
−1], where
is the density of water [Mass Length
−3] and
is the specific heat capacity of water [Energy Temperature
−1 Mass
−1], both assumed constant. Following Mohseni and Stefan [
56], we ignore thermal exchange with the stream bed and characterize vertical flux across the air-water interface using the temperature equilibrium concept of Edinger et al. [
57], whereby the vertical flux of thermal energy at any time and location is proportional to the difference between a changing equilibrium temperature
and the local water temperature
. The resulting transport equation, after dividing throughout by
, has the form
subject to initial and boundary conditions
where
is a thermal exchange coefficient [Energy Length
−2 Time
−1 Temperature
−1].
For purposes of the present numerical examples, we further simplify the model by assuming that
is constant over
t and
x and that the equilibrium temperature at any time
t and location
x is simply a linear function of solar illuminance
. Equation (
20) therefore becomes
where
[Time
−1] and
Parameters
A,
a, and
b in these equations are positive empirical constants whose values (
h
−1,
°C,
°C klx
−1) were chosen to ensure that predicted dynamics of water temperature resembled the temperature time series shown in
Figure 1.
Now consider a transverse slice of water that crosses the upstream boundary
of the focal stream reach at time
and moves downstream with constant velocity
. Then the position of the slice at time
is
. Along this characteristic base curve, Equation (
21) reduces to the ordinary differential equation
where
. The corresponding concentrations of old DO, new DO that entered the slice from the atmosphere, and new DO that was produced by photosynthesis are given by Equations (
7) and (
9), and the process rates are given by Equations (
17)–(19).
Dividing all terms of Equation (
20) by
yields a partial differential equation with characteristic base curves
, which are solutions of the ordinary differential equation
. Along these curves (which are the same as those for Equation (
20)), the temperature of a slice of water at location
x that entered the focal stream reach at time
is given by the solution of differential equation
where
and
Solutions of Equation (
23) are expressed as functions of transport distance
x instead of time
t, consistent with the focus of this paper on longitudinal spatial patterns.
Numerical solutions of Equations (
7), (
9), (
22) and (
23) were computed and plotted using the R programming language [
58] and the deSolve package [
59].
Figure 8 shows three examples in which the DO concentration and other properties of a slice of water are plotted as a function of travel time as the slice moves downstream from reach boundary
, which it crosses at “real” time (as opposed to travel time)
h. In all examples, real time
h corresponds to sunset for the previous day, and
h corresponds to sunrise for the current day. Panels in column 1 (A, B, C) apply to a slice during its journey over a 36-h period of time after entering the reach at time
(i.e., at sunset for the previous day) with initial temperature 26.275 °C and initial concentrations 7.759, 0.0, 0.0 mg/L of old DO, new DO from the atmosphere, and new DO from photosynthesis. These panels show the solar irradiance and water temperature the slice experiences as it moves downstream (panel A), its total and gas-exchange equilibrium DO concentrations (panel B), and the proportions of its total DO concentration that consist of old DO, new DO derived from atmospheric exchange, new DO derived from photosynthesis, and combined new DO (panel C) as functions of travel distance
x and travel time
. Panels in column 2 (D, E, F) and column 3 (G, H, I) employ the same parameter values, except that the slice’s reach entry time for column 2 is
h (6 h past sunset for the previous day) and that for column 3 is
h (12 h past sunset for the previous day, which coincides with sunrise for the current day); the initial temperatures and DO concentrations in columns 2 and 3 were taken from the solution in column 1 for the times corresponding to
in columns 2 and 3.
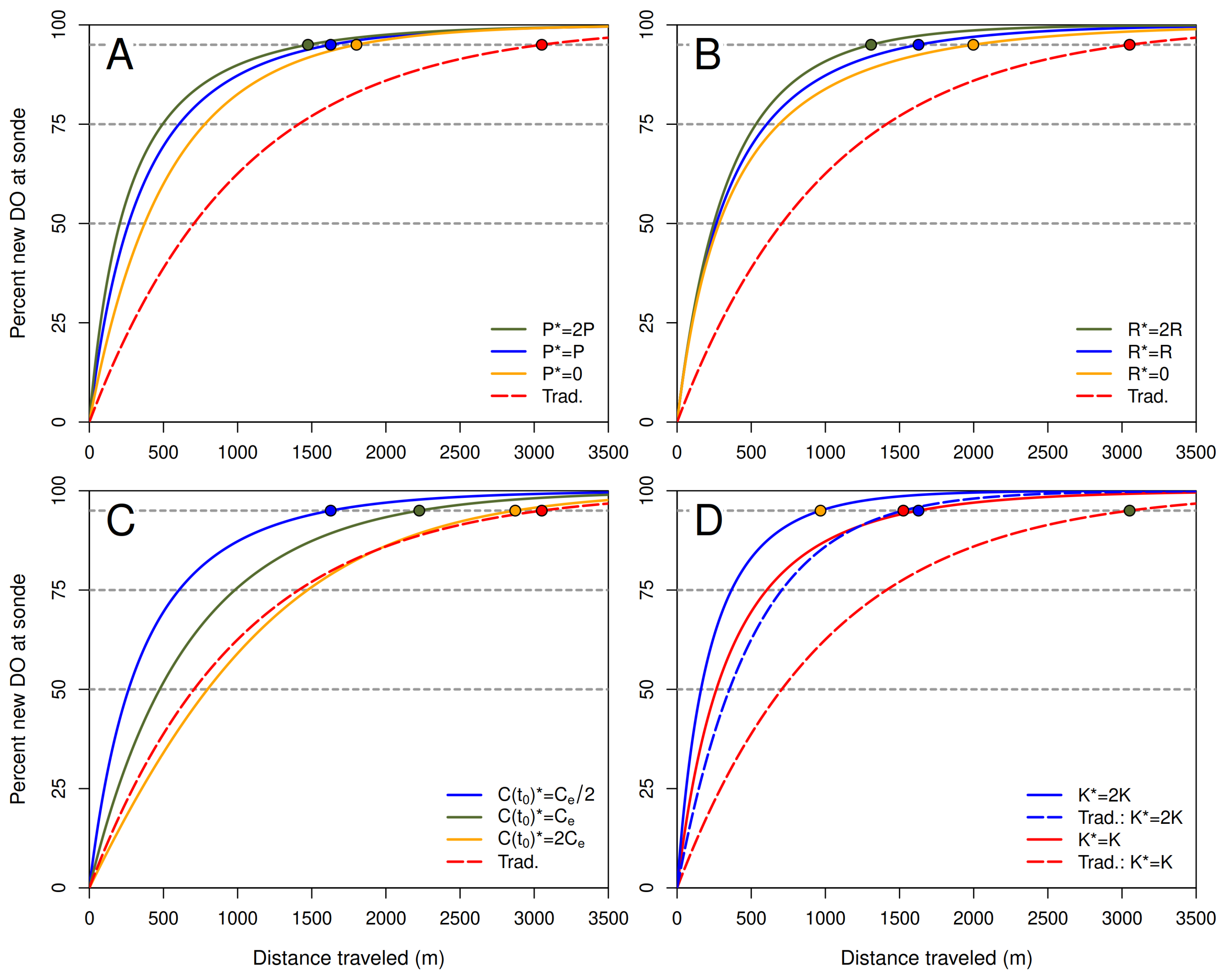
In the first example (column 1), sunrise and the resulting pulse of new DO produced by photosynthesis do not occur until after the proportion of new DO in the slice (red curve) has exceeded 95%. In the third example (column 3), these phenomena occur at the beginning of the journey and therefore affect the dynamics of DO concentration before the proportion of new DO in the slice exceeds 95%. Nevertheless, the distance required for the new-DO threshold to be crossed does not differ greatly for these two extreme examples or for the intermediate (second) example, ranging from about 1.6 to 2.1 km.
Figure 9 shows plots of the proportion of new DO as a function of time and distance traveled for the same three examples shown in
Figure 8. The solid dark blue line in each panel is the exact relationship computed using the model with time-varying process rates. The dashed red line was computed using the traditional approach where old DO is treated as an inert tracer, but the atmospheric exchange coefficient
was allowed to vary with temperature and hence time. The required equations were obtained by putting
and
in the first of Equations (
7) and (
9) and solving to obtain
Note the marked difference between the curves resulting from the proposed and traditional approaches and the resulting marked difference in the travel time and distance at which the proportion of new DO crosses the 95% threshold. The length of the transition zone under the proposed approach is roughly a kilometer shorter than under the traditional approach in these examples.
The solid light blue and dashed orange curves in
Figure 9 represent different constant-coefficient approximations. The solid light blue curve was computed using the constant-rate formulas in Equations (
12) and (
13), with the values of
P,
R, and
K being the values at 20 °C adjusted to the 24-h average water temperature and with normalized solar irradiance replaced by its 24-h average, the value of
being the corresponding value for the 24-h average water temperature, and the value of
being the 24-h average DO concentration. The dashed orange curve in each panel is the proportion of new DO according to the traditional approach with the atmospheric exchange coefficient constant and equal to the value calculated using
adjusted to the 24-h average water temperature.
In addition to the 95% threshold for new DO,
Figure 9 also shows the 75% and 50% thresholds. As noted above, the traditional 95% threshold for new DO is a largely arbitrary criterion for determining the length of the transition zone. The 50% threshold is useful as a criterion for estimating where the switch occurs between the upstream reach having the most influence on the observed DO concentration to the focal reach having the most influence. It is interesting to note that the downstream distance at which the 75-th percentile for new DO is first achieved is about half the distance at which the 95-th percentile is first achieved.
The examples shown in
Figure 9 illustrate the fact that the exact length of the
transition zone, based on the proposed approach with time-varying process rates, varies significantly but not greatly with time of day, due to variation in solar irradiance and water temperature and the effects of this variation on oxygen solubility and rates of photosynthetic oxygen production, respiration, and atmospheric exchange. For slices of water entering the focal reach at any time between sunset and sunrise, the length of the transition zone calculated with the time-varying model ranged from 1.61 to 2.07 km, with a mean of 1.84 km. For comparison, the value of the proposed constant-coefficient estimate
was 1.90 km (bias
% relative to the mean exact value), while the length of the the traditional estimate
was 2.75 km (relative bias
%). As can be seen in
Figure 9, the actual percentage of new DO at
is sometimes a bit less than 95% and sometimes a bit more, with a range of about 94–97% in numerical examples. By contrast, the actual percentage of new DO at
is greater than 95% in all numerical examples, with a range of about 98–99%. Thus, the proposed constant-coefficient estimate
was reasonably close to the average true length of the transition zone, and the resulting true percentages of new DO at
were close to 95%. The traditional constant-coefficient estimate
was consistently greater than both the true length of the transition zone and the proposed estimate
, and as a result, the true percentages of new DO at
were consistently much greater than 95%. The traditional estimate
was therefore quite conservative in these examples, often resulting in overestimates of the transition-zone length on the order of a kilometer and implying new DO percentages of roughly 99% instead of 95%.
8. The Traditional Estimate of Transition Zone Length as a Special Case
Most of the published papers that use or mention the traditional estimate
of transition zone length provide no argument to justify it but simply cite a paper by Chapra and Di Toro [
60], which actually addresses a different problem (see below). Two exceptions are the technical manual by Grace and Imberger [
26] (who interpret
as the distance integrated by a sonde) and the supplemental information that accompanies a paper by Demars et al. [
30] (who interpret
as the footprint of a sonde), both of which present clear derivations based exclusively on evasion of old DO from a slice of water as it moves from upstream reach boundary
to the sonde.
To obtain a simple formula, these authors assume the atmospheric exchange coefficient is constant. In contrast to our approach, they focus on the concentration
of old DO in a moving slice of water as a proportion of the concentration
that was initially present in the slice when it entered the focal reach instead of as a proportion of the total DO concentration in the slice at travel time
t. This approach ignores the effects of photosynthesis and respiration within the focal reach. The model that underlies the resulting estimate
of the transition zone length is therefore the first of our Equation (
7), with
and
(both constant).
Solving this equation, we find that the proportion
of DO initially present in a slice of water entering the focal stream reach at time
that is still present at travel time
(as opposed to the proportion of total DO in the slice at travel time
t that is old) is given by
or on a distance scale,
Defining
as the travel distance
x at which this proportion first decreases to
(equivalently, the travel distance at which the proportion of initial DO that has escaped to the atmosphere first increases to
p), we find that
It follows that
for
, which is the traditional estimate.
In explaining the basis of this estimate, Grace and Imberger [
26] point out that “the oxygen concentration at a particular point in a stream, A, is a result of the concentration at some upstream point, B, corrected for the water travel time between B and A, and the processes affecting DO between B and A (i.e., reaeration, primary production, and aerobic respiration)” (p. 76). This is essentially the basis for the alternative estimate
we have proposed. However, the derivations that Grace and Imberger [
26] and Demars et al. [
30] provide only account for evasion of old DO to the atmosphere.
The other papers that mention or use the traditional estimate
cite the paper by Chapra and Di Toro [
60] as justification. But when we look at this paper, we find no claim about the stream reach integrated by a sonde, the footprint of a sonde, or the length of the transition zone. Instead, the authors provide an estimate of how far down from the upstream boundary of a uniform stream reach the longitudinal distribution of DO concentration first becomes approximately constant, so that
. Specifically, Chapra and Di Toro state: “For situations where the plants [or more generally, the dominant oxygenic photoautotrophs] are uniformly distributed for a sufficiently long distance (
), deficit does not vary spatially (
)” (p. 641). The term “deficit” here refers to the DO deficit
. The form of the continuity equation for DO dynamics used by these authors assumes that gas-exchange equilibrium
is constant in time and space, in which case
if and only if
. Thus, the claim is that the longitudinal profile of DO concentration in a uniform stream reach will be approximately flat (
) for distances greater than
down from the upstream boundary of the reach.
This distance clearly is not equivalent to the distance at which the proportion of new DO first increases to 95%, because it addresses only the total DO concentration, not where or how various components of that total entered the stream. To demonstrate this nonequivalence, it suffices to consider the hypothetical case where process rates are constant and the DO concentration of water entering the focal reach is constant and equal to equilibrium concentration
. Since the DO concentration in each slice of water is already at the equilibrium for the focal reach when the slice enters at the upstream boundary, it will remain constant at
as the slice moves through the reach. Since this is true for all slices moving through the focal reach, the DO concentration in the reach will be constant longitudinally, implying that the distance to achieve longitudinal constancy is zero. But the proportion of new DO will nevertheless be zero at the upstream boundary of the reach and will steadily increase downstream as new DO enters and old DO exits until it eventually reaches 95%, with the required travel distance being given by Equation (
16) with
.
Finally, it is instructive to consider what additional specializing assumptions are required in order to obtain the traditional estimate
in Equation (
1) as a special case of our proposed estimate
. To be consistent with the derivations by Grace and Imberger [
26] and Demars et al. [
30], we assume the photosynthesis rate, respiration rate, and atmospheric exchange coefficient are constant in time and space, that the photosynthesis and respiration rates are in fact zero, and that water from upstream enters the focal reach with its DO concentration already at the steady-state concentration for the reach. (The last assumption is required because the derivation of
is based on the proportion of old DO in a moving slice of water at the sonde relative to the initial total DO concentration in the slice when it crossed the upstream boundary of the reach, whereas
is based on the proportion of old DO in the slice at the sonde relative to the present total DO concentration at the sonde; to make this difference irrelevant, we must assume the DO concentration of a slice remains constant as it moves through the focal reach and therefore was already at the steady state when it entered the reach.) In terms of Equation (
7), these assumptions mean that
and
for all
,
(constant), and
. It follows at once that
and
in Equations (
15) and (
16), whence
Thus,
can be obtained from
by imposing additional artificial specializing assumptions. However, the fundamental difference between these two estimates of the transition-zone length is that our proposed estimate (
) focuses on old DO at travel time
t as a proportion of total DO in the slice at
t (thus requiring us to account for photosynthesis, respiration, and invasion of DO from the atmosphere in addition to evasion of old DO), whereas the traditional estimate (
) focuses on old DO at travel time
t as a proportion of the total DO in the slice when it entered the focal reach at travel time 0 (thereby excluding the effects of photosynthesis, respiration, and invasion of DO from the atmosphere).
9. Example: Application to Streams in Eastern New York, USA
Over the course of each diel cycle, there is a continually shifting imbalance between processes that tend to increase DO concentrations (oxygenic photosynthesis, invasion from the atmosphere) and opposing processes that tend to decrease DO concentrations (aerobic respiration, evasion to the atmosphere). The instantaneous rates of these processes in the models of
Section 5 and
Section 6 therefore change significantly over a diel cycle. However, the numerical examples of
Section 7 suggest that the lengths of the transition zone under the traditional and newly proposed approaches with time-varying process rates can be usefully approximated by replacing time-varying instantaneous rates in the model of DO dynamics with (constant) diel averages. This procedure makes it possible to calculate and compare estimates of the transition-zone length under the traditional and newly proposed approaches using data sometimes reported in published studies of metabolism in real streams.
A study that is particularly useful for this purpose is detailed by Bott et al. [
20] and Newbold et al. [
61]. It provides estimates of 24-h photosynthetic production and respiratory consumption of DO, atmospheric exchange rates, water temperature, stream depth, and current velocity (and several other properties) for 10 streams in eastern New York state, USA, in three successive years. The reported values were obtained using consistent methods across streams and years. They include sufficient information for calculating
and
using Equations (
15) and (
16), as well as the traditional estimate
given by Equation (
1), thus making comparison of the two types of estimate possible for multiple streams and years.
The data employed in these calculations and the resulting estimates are summarized in
Appendix A. Data for daily mean water temperature, minimum and maximum DO as percent gas-exchange equilibrium (“saturation”), and mean
K were taken from Table 1 of Bott et al. [
20], estimates of 24-h cumulative DO production by photosynthesis were digitized from
Figure 3 (same paper), estimates of 24-h cumulative DO consumption by respiration were digitized from
Figure 5 (same paper), and data for mean current velocity and depth were taken from Appendix 1 of Newbold et al. [
61]. No values corresponding directly to
were provided, so the reported values of the minimum and maximum DO as percent saturation, converted to concentrations, were used as reasonable estimates of the range of potential values of
.
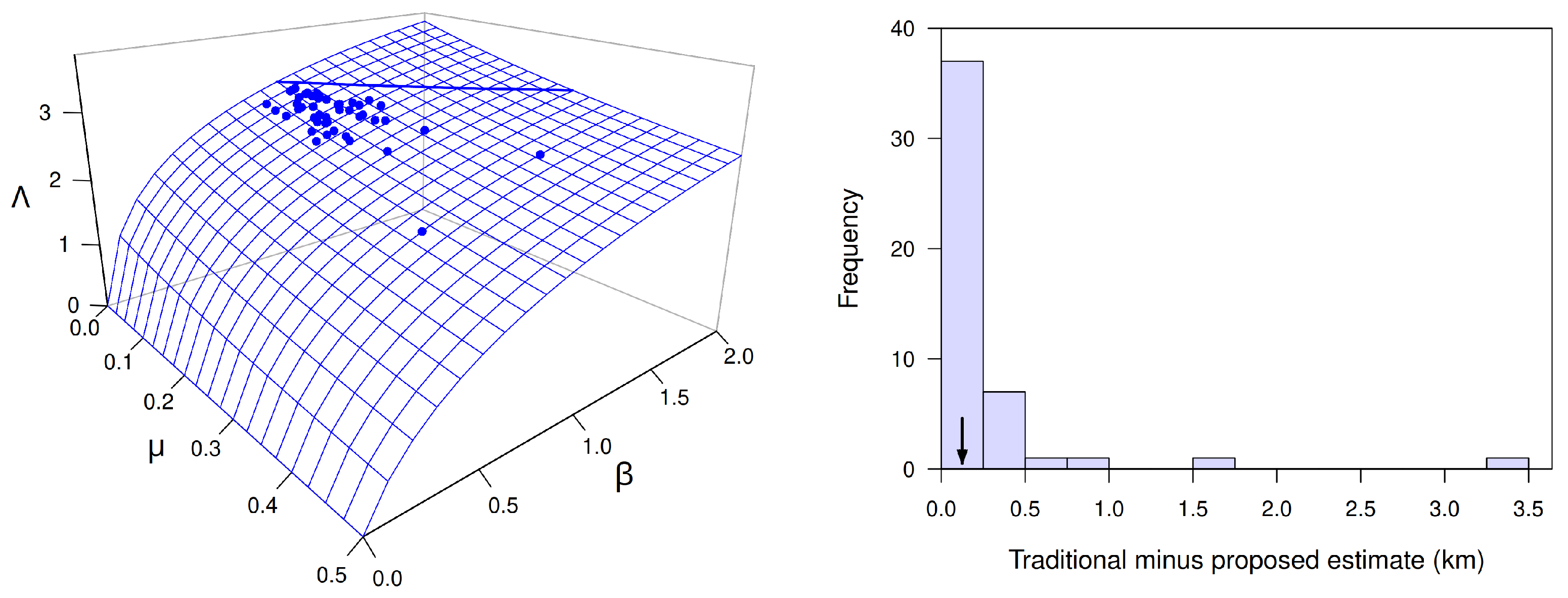
Figure 10 displays the key results. The left panel shows the values of
, calculated from estimates of
and
, plotted on the portion of the
surface shown in
Figure 6 for values of
between 0 and 2 and values of
between 0 and
. Note that all values of
are less than the traditional value of 3, implying that
is shorter than
. The right panel in
Figure 10 shows a histogram of the difference
between the traditional and newly proposed estimates of the transition-zone length, all of which are positive. Nearly all of the 48 differences are less than
km, and the median difference is
km. One of the 48 estimates, however, is greater than
km and another is greater than 3 km. These results suggest that the traditional estimate
often will exceed the proposed estimate
by less than 200 m but occasionally will exceed it by more than a kilometer.
11. Discussion
As noted in the Introduction, streams often consist of a longitudinal sequence of more-or-less distinct reaches dominated by different habitat and community types, each with its own characteristic community metabolism. As the flowing water exits one reach and enters the next, there is a transition zone in which physical and chemical properties of the water, such as temperature and DO concentration, change from being representative of the upstream reach to being representative of the downstream reach. The theoretical approaches we have developed provide practical ways to estimate the length of this transition zone for DO. Rutherford et al. [
42,
43] address the corresponding problem for water temperature.
The extent of the DO transition zone between one stream reach and the next is important in metabolism studies employing the FWDO method with 1-station monitoring, since this method derives its metabolism estimates from time series acquired at a single location per reach. If a sonde is placed so close to the upstream boundary of the focal reach that it lies well within the transition zone, the DO time series it acquires may largely reflect DO that entered the stream upstream of the focal reach rather than within it. Another way to say this is that the reach actually being monitored may extend well upstream of the focal reach. Viewed either way, it clearly is unlikely in this case that metabolism estimates based on time series acquired by the sonde will mainly reflect the habitat and community type in the focal reach.
Obtaining metabolism estimates that are representative of specific types of stream habitat is important in characterizing and comparing streams. The well-known longitudinal patterns of change in stream and river habitat over long distances are a central part of the river continuum concept of Vannote et al. [
62]. These patterns require that useful comparisons between streams be stratified in order to control for position within the river continuum as indicated by stream order or magnitude. Otherwise, effects of the large-scale trend of changing habitat conditions along lengthy streams and rivers (e.g., water depth, light attenuation, current velocity, suspended particulate matter, shading by riparian vegetation) are confounded with effects of other properties (e.g., local climate, anthropogenic stressors) that can affect stream ecosystem function. It is also important to be aware that marked changes in stream habitat can occur even over short distances (e.g., shifts between shaded and unshaded reaches, or between riffles and pools), and these can have pronounced effects on local stream metabolism at a given position within the river continuum (e.g., [
2]).
In addition to its importance for studies of stream metabolism, the length of the transition zone is also important in studies concerned specifically with predicting stream DO concentrations and how they will respond to potential management options. This, of course, is the problem addressed by the original Streeter-Phelps model for cases where metabolism is dominated by catabolism of allochthonous organic matter discharged to a river by upstream point sources. Another example is the design of stream restoration projects to improve fish habitat, where both DO concentration and water temperature may be important. The key point is that the temperature and DO concentration at a given stream location are strongly influenced by habitat upstream of the location. This dependence must be accounted for in estimating and interpreting components of stream metabolism via the FWDO method, or in designing stream restoration projects to improve DO or temperature conditions in a particular reach.
In studies of stream metabolism, the length of the transition zone is an important constraint on where the sonde should be placed within a focal reach when 1-station monitoring is used. Another important consideration is whether the length of a focal reach and the longitudinal pattern of change in DO concentration within it is suitable for estimating components of metabolism. This problem is discussed by Grace and Imberger [
26] and Reichert et al. [
21], among others. For example, 2-station monitoring often fails to produce plausible estimates of metabolism if the distance between the two sondes is so short, relative to the current velocity and the rates of metabolism and atmospheric exchange, that little change in DO concentration occurs between them [
26]. In the case of 1-station monitoring, it is commonly noted that obtaining reliable estimates of metabolism components requires the longitudinal profile of DO concentration to be approximately uniform (flat) in the vicinity of the sonde (e.g., [
26]). More precisely, this form of the FWDO method assumes that
in the fundamental continuity equation stated in Equation (
2), and that the gas-exchange equilibrium and the photosynthesis, respiration, and atmospheric exchange rates all depend on time
t but not on location
x within the reach. The DO continuity equation then reduces to the simpler ordinary differential equation,
which is the basis for estimating parameter values from DO and temperature time series when 1-station monitoring is used.
The requirement that
(approximately) in the vicinity of each sonde in metabolism studies employing 1-station monitoring imposes a second constraint on admissible sonde locations, in addition to that imposed by the length of the transition zone. To illustrate the relationship between these two constraints, we briefly consider the conditions under which the longitudinal profile of DO concentration will be approximately uniform in the vicinity of a sonde. We have already mentioned this problem in discussing the basis of the traditional estimate
of the transition-zone length in
Section 8, since this is actually the problem that Chapra and Di Toro [
60] claim their commonly-cited estimate addresses. We now consider it more carefully to clarify the meaning of Chapra and Di Toro’s result, which turns out to be somewhat different than these authors suggest.
Given the usual model of stream DO transport and dynamics outlined in
Section 5, and replacing the various process rates with averages over a suitable time period (e.g., a diel cycle), the longitudinal profile of DO concentration
will converge to a stationary pattern
as
t becomes large. In reality, such a stationary pattern will never be attained, mainly because of the pronounced diel cycle in solar irradiance which, directly and indirectly, forces diel changes in rates of the various processes that tend to increase and decrease stream DO concentrations. Nevertheless, the constant-rate approximation to the model of DO transport and dynamics provides useful approximations to results of the variable-rate model, as we saw in
Section 7 for estimates of the transition-zone length.
One way to find the stationary longitudinal DO profile
is by setting
in Equation (
2). To prevent dependence of the results on physical dimensions, we introduce dimensionless variables
and
in the resulting differential equation. Solving this equation (see
Appendix B), we find that the absolute value of the slope of the dimensionless longitudinal DO profile
is given by
where dimensionless parameter
(which is consistent with the definition of
in Equation (
11)). The absolute value of the slope is therefore greatest at the upstream boundary of the focal reach and decreases exponentially with increasing dimensionless distance
downstream.
Let
denote the minimum value of
such that
, where
is a positive dimensionless number much smaller than unity. Clearly
if
. Otherwise,
. The corresponding dimensional distance
is therefore given by
Thus, if the DO concentration
of water entering the focal reach is already close enough to
so that
, then the slope of the DO profile will be sufficiently close to zero for all
and there is no constraint on the location of the sonde. But if the initial DO concentration is substantially different from
, then
and its value will be greater, the greater the initial discrepancy measure
is.
As an example, if we choose
, and if
, then
This estimate may be compared with that of Chapra and Di Toro [
60], which is the same as the traditional estimate of the transition-zone length in Equation (
1). Their estimate does not depend on the magnitude of the initial discrepancy between
and
, which is physically incorrect (unless one argues that
, which is physically unjustified). The estimate of Chapra and Di Toro actually tells us the distance at which the discrepancy
at downstream distance
x first becomes sufficiently small
relative to initial discrepancy . This is an appropriate distance-based measure of the relative responsiveness of the stream ecosystem to displacements from steady state but is not appropriate as a measure of the absolute distance at which
becomes approximately constant, which clearly depends on the magnitude of the initial discrepancy.
When employing the FWDO method with 1-station monitoring to estimate components of stream metabolism in a particular reach, length estimates
and
impose two constraints that jointly determine the allowable locations for the sonde. One of these constraints—the main focus of the present paper—assures that the time series of DO concentrations acquired by the sonde mainly reflects the stream community in the focal reach. The other assures that the longitudinal profile of DO concentration is approximately flat in the vicinity of the sonde, which is a condition for obtaining valid metabolism estimates with 1-station monitoring. Both constraints must be satisfied if metabolism estimates based on the acquired time series are to be valid and attributable to the focal reach. To satisfy both constraints, the distance
between the upstream boundary of the focal reach and the sonde must be at least as large as both
and
for suitable choices of
p and
. That is,
If the sonde is placed closer than
to the upstream boundary of the focal reach, then the FWDO method with 1-station monitoring will yield unreliable estimates of the components of community metabolism in the focal reach. A corollary is that it will not be possible to obtain reliable estimates if the total length of the focal reach is less than
.
Two extreme cases are instructive. First, note that
approaches zero as
does (with
fixed). It follows from Equation (
15) that
, and therefore
. This case addresses the artificial but conceptually important case where the DO concentration of water entering the focal reach is approximately zero, and all DO at every location within the reach is therefore new. It follows that the first constraint will be satisfied regardless of where the sonde is placed within the focal reach, so only the second constraint restricts placement of the sonde. The other extreme case is where
. Here,
and
for all
in Equation (
34), so
. In this case, then, the slope of the longitudinal DO profile is zero at all locations within the focal reach, so only the first constraint restricts placement of the sonde. Aside from these extreme cases, both constraints impose significant restrictions on sonde placement, and the condition in Equation (
35) becomes nontrivial.
But how should the values of thresholds
p and
in
and
be chosen? As we have indicated in previous sections of this paper, the traditional choice of
p is
. Our own subjective view is that this value is probably higher than necessary and should be regarded as conservative. On subjective grounds, we also suggest that
is a reasonable choice for
, but we have no evidence that this value is small enough to be conservative. Objective evidence for appropriate values of
p and
must come from numerical studies in which a specific statistical method is employed for estimating components of metabolism from artificial time series generated by models like the one presented in
Section 7, with random noise added. In this way, the true values of photosynthesis, respiration, and atmospheric exchange parameters will be known and the error resulting from estimating these parameters from time series acquired at different distances from the upstream boundary of the focal reach can therefore be rigorously quantified. Numerical values of thresholds
p and
that are protective but not overly so can then be determined. This task, however, is beyond the scope of the present paper.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}