
Simple Sugars Alter the Odorant Composition of Dairy Cow Manure



Abstract

1. Introduction
2. Materials and Methods
2.1. Manure Incubations
2.2. Biogas Measurement
2.3. Determination of Short-Chain Fatty Acids
2.4. Determination of Odor Compounds
3. Results and Discussion
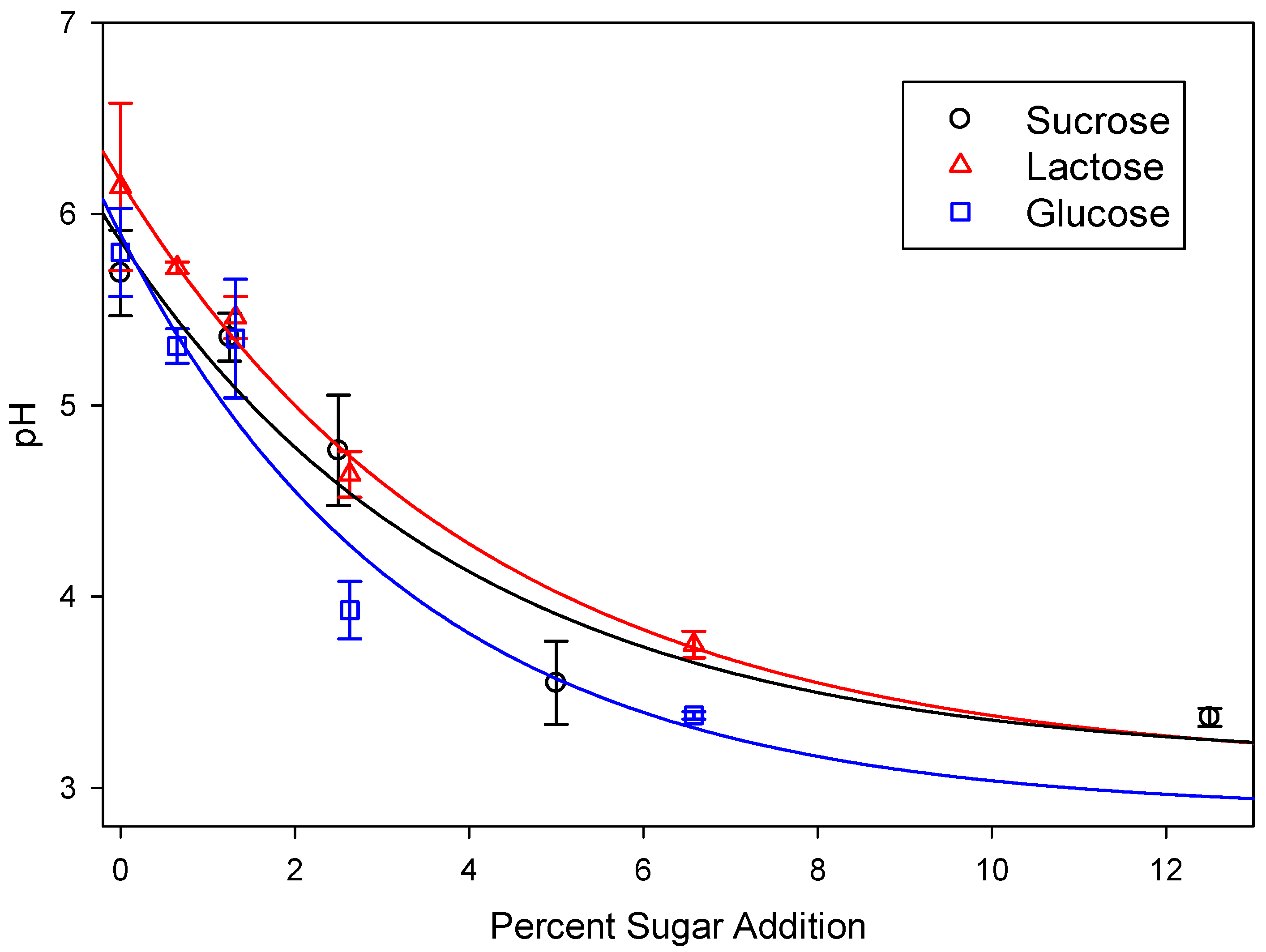
3.1. Biogas Production
3.2. Short-Chain Fatty Acids
3.3. Effect of Sugar Addition on Odorants
3.3.1. Glucose
3.3.2. Lactose
3.3.3. Sucrose
4. Conclusions
Implication of Sugar Addition for Manure Odorants
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Yates, S.R.; McConnell, L.L.; Hapeman, C.J.; Papiernik, S.K.; Gao, S.; Trabue, S.L. Managing agricultural emissions to the atmosphere: State of the science, fate and mitigation, and identifying research gaps. J. Environ. Qual. 2011, 40, 1347–1358. [Google Scholar] [CrossRef]
- Ma, Q.; Meng, N.; Li, Y.; Wang, J. Occurrence, impacts, and microbial transformation of 3-methylindole (skatole): A critical review. J. Hazard. Mat. 2021, 416, 126181. [Google Scholar] [CrossRef]
- Hudson, R. Odor and odorant: A terminological clarification. Chem. Senses. 2000, 25, 693. [Google Scholar] [CrossRef]
- Parker, D.B. Reduction of odor and VOC emissions from a dairy lagoon. Appl. Eng. Agricul. 2008, 24, 647–655. [Google Scholar] [CrossRef]
- Rybarczyk, P.; Szulczyński, B.; Gebicki, J.; Hupka, J. Treatment of malodorous air in biotrickling filters: A review. Biochem. Eng. J. 2019, 141, 146–162. [Google Scholar] [CrossRef]
- Page, L.H.; Ni, J.-Q.; Zhang, H.; Heber, A.J.; Mosier, N.S.; Liu, X.; Joo, H.-S.; Ndegwa, P.M.; Harrison, J.H. Reduction of volatile fatty acids and odor offensiveness by anaerobic digestion and solid separation of dairy manure during manure storage. J. Environ. Manag. 2015, 152, 91–98. [Google Scholar] [CrossRef]
- Szogi, A.A.; Loughrin, J.H.; Vanotti, M.B. Improved water quality and reduction of odorous compounds in anaerobic lagoon columns receiving pre-treated pig wastewater. Environ. Technol. 2018, 39, 2613–2621. [Google Scholar] [CrossRef]
- Spoelstra, S.F. Simple phenols and indoles in anaerobically stored piggery wastes. Sci. Food Agricul. 1977, 28, 415–423. [Google Scholar] [CrossRef]
- Trabue, S.; Kerr, B.; Bearson, B.; Ziemer, C. Swine odor analyzed by odor panels and chemical techniques. J. Environ. Qual. 2011, 40, 1510–1520. [Google Scholar] [CrossRef]
- Whitehead, T.R.; Price, N.; Drake, H.L.; Cotta, M.A. Catabolic pathway for the production of skatole and indoleacetic acid by the acetogen Clostridium drakei, Clostridium scatologenes, and swine manure. Appl. Environ. Microbiol. 2008, 74, 1950–1953. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jensen, R.L.; Højberg, O.; Canibe, N.; Jensen, B.B. Olsenella scatoligenes sp. nov., a 3-methylindole- (skatole) and 4-methylphenol- (p-cresol) producing bacterium isolated from pig faeces. Int. J. Syst. Evol. Microbiol. 2015, 65, 1227–1233. [Google Scholar] [CrossRef]
- Varel, V.H.; Miller, D.N. Effect of carvacrol and thymol on odor emissions from livestock wastes. Water Sci. Technol. 2001, 44, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Varel, V.H.; Miller, D.N.; Lindsay, A.D. Plant oils thymol and eugenol affect cattle and swine waste emissions differently. Water Sci. Technol. 2004, 50, 207–213. [Google Scholar] [CrossRef]
- Kendig, M.D.; Hasebe, K.; McCague, R.; Lee, F.; Leigh, S.-J.; Arnold, R.; Morris, M.J. Adolescent exposure to a solid high-fat, high-sugar ‘cafeteria’ diet leads to more pronounced changes in metabolic measures and gut microbiome composition than liquid sugar in female rats. Appetite 2022, 172, 105973. [Google Scholar] [CrossRef]
- Neves, V.G.O.; de Oliveira, D.T.; Oliveira, D.C.; Perucci, L.O.; dos Santos, T.A.P.; Fernandes, I.C.; de Sousa, G.G.; Barboza, N.R.; Guerr-Sá. High-sugar diet intake, physical activity, and gut microbiota crosstalk: Implications for obesity in rats. Food Sci. Nutr. 2020, 8, 5683–5695. [Google Scholar] [CrossRef] [PubMed]
- Farnsworth, E.R.; Modler, H.W.; Mackie, D.A. Adding Jerusalem artichoke (Helianthus tuberosus L.) to weanling pig diets and the effect on manure composition and characteristics. Anim. Feed Sci. Technol. 1995, 55, 153–160. [Google Scholar] [CrossRef]
- Gerard, J. The Herbal or General History of Plants; Dover Publications: Mineola, New York, USA, 1975; p. 1678. [Google Scholar]
- Chibbar, R.N.; Jaiswal, S.; Gangola, M.; Baga, M. Carbohydrate Metabolism. Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Kolida, S.; Tuohy, K.; Gibson, G.R. Prebiotic effects of inulin and oligofructose. Br. J. Nutr. Suppl. 2002, 87, S193–S197. [Google Scholar] [CrossRef]
- Salazar, N.; Dewulf, E.M.; Neyrinck, A.M.; Bindels, L.B.; Cani, P.D.; Mahillon, J.; de Vos, W.M.; Thissen, J.-P.; Gueimonde, M.; de Los Reyes-Gavilán, C.G.; et al. Inulin-type fructans modulate intestinal Bifidobacterium species populations and decrease fecal short-chain fatty acids in obese women. Clin. Nutr. 2015, 2015, 501–507. [Google Scholar] [CrossRef]
- Yang, Y.-X.; He, M.; Hu, G.; Wei, J.; Pages, P.; Yang, X.-H.; Bourdu-Maturel, S. Effect of a fermented milk containing Bifidobacterium lactis DN-173010 on Chinese constipated women. World J. Gastroenterol. 2008, 14, 6237–6243. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.; Stasse-Wolthuis, M. The bifidogenic effect of inulin and oligofructose and its consequences for gut health. Eur. J. Clin. Nutr. 2009, 63, 1277–1289. [Google Scholar] [CrossRef]
- Ghouri, Y.A.; Richards, D.M.; Rahimi, E.F.; Krill, J.T.; Jelinek, K.A.; DuPont, A.W. Systematic review of randomized controlled trials of probiotics, prebiotics, and synbiotics in inflammatory bowel disease. Clin. Exp. Gastroenterol. 2014, 7, 473–487. [Google Scholar] [CrossRef]
- De Graff, A.A.; Venema, K. Gaining insight into microbial physiology in the large intestine: A special role for stable isotopes. Adv. Microb. Physiol. 2008, 53, 73–168. [Google Scholar] [CrossRef]
- Lincoln, L.; More, S.S. Bacterial invertases: Occurrence, production, biochemical characterization, and significance of transfructosylation. J. Basic Microbiol. 2017, 57, 803–813. [Google Scholar] [CrossRef]
- Long, C.-X.; He, L.; Guo, Y.-F.; Xiao, N.-X.; Tan, Z.-J. Diversity of bacterial lactase genes in intestinal contents of mice with antibiotics-induced diarrhea. World J. Gastroenterol. 2017, 23, 7584–7593. [Google Scholar] [CrossRef]
- Müller, V. Bacterial Fermentation. In Encyclopedia of Life Sciences; John Wiley & Sons: Hoboken, NJ, USA, 2001. [Google Scholar] [CrossRef]
- Lay, J.-J.; Li, Y.-Y.; Noike, T. The influence of pH and ammonia concentration on the methane production in high-solids digestion processes. Water Environ. Res. 1998, 70, 1075–1079. [Google Scholar] [CrossRef]
- Bintis, T. Lactic acid bacteria as starter cultures: An update in their metabolism and genetics. AIMS Microbiol. 2018, 4, 665–684. [Google Scholar] [CrossRef]
- Pietzke, M.; Meiser, J.; Vazquez, A. Formate metabolism in health and disease. Mol. Metab. 2020, 33, 23–37. [Google Scholar] [CrossRef]
- Peck, S.M.; Rosenfeld, H.; Leifer, W.; Bierman, W. Role of sweat as a fungicide: With special reference to the use of constituents of sweat in the therapy of fungous infections. Arch. Derm. Syphilol. 1939, 39, 126–148. [Google Scholar] [CrossRef]
- Machado, M.G.; Sencio, V.; Trottein, F. Short-chain fatty acids as a potential treatment for infections: A closer look at the lungs. Infect. Immun. 2021, 89, e00188-21. [Google Scholar] [CrossRef]
- Kadry, A.A.; El-Antrawy, M.A.; El-Ganiny, A.M. Impact of short chain fatty acids (SCFAs) on antimicrobial activity of new β-lactam/β-lactamase inhibitor combinations and on virulence of Escherichia coli isolates. J. Antibiot. 2023, 76, 225–235. [Google Scholar] [CrossRef]
- Aldunate, M.; Srbinovski, D.; Hearps, A.C.; Latham, C.F.; Ramsland, P.A.; Gugasyan, R.; Cone, R.A.; Tachedjian, G. Antimicrobial and immune modulatory effects of lactic acid and short chain fatty acids produced by vaginal microbiota associated with eubiosis and bacterial vaginosis. Front. Physiol. 2015, 6, 164. [Google Scholar] [CrossRef] [PubMed]
- López, M.L.; Lavilla, M.T.; Riba, M.; Vendrell, M. Comparison of volatile compounds in two seasons in apples: Golden delicious and Granny Smith. J. Food Qual. 1998, 21, 155–166. [Google Scholar] [CrossRef]
- Yokoyama, M.T.; Carlson, J.R. Production of Skatole and para-Cresol by a Rumen Lactobacillus sp. Appl. Environ. Microbiol. 1981, 41, 71–76. [Google Scholar] [CrossRef]
- Nowak, A.; Libudzisz, Z. Ability of intestinal lactic bacteria to bind or/and metabolise phenol and p-cresol. Ann. Microbio. 2007, 57, 329–3325. [Google Scholar] [CrossRef]
- Lim, J. Hedonic scaling: A review of methods and scaling. Food Qual. Pref. 2011, 22, 733–747. [Google Scholar] [CrossRef]
- Zarso, M. Hedonic judgements of chemical compounds are correlated with molecular size. Sensors 2011, 11, 3667–3686. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Koziel, J.A.; Cai, L.; Hoff, S.J.; Heathcote, K.Y.; Chen, L.; Jacobson, L.D.; Akdeniz, N.; Hetchler, B.P.; Parker, D.B.; et al. Odor and odorous chemical emissions from animal buildings: Part 5. Simultaneous chemical and sensory analysis with gas chromatography-mass spectrometry-olfactometry. Trans. ASABE 2015, 58, 1349–1359. [Google Scholar] [CrossRef]
- Nagata, Y.; Takeuchi, N. Measurement of odor threshold by triangle odor bag method. Odor Meas. Rev. 2003, 118, 118–127. [Google Scholar]
- Øverland, M.; Kjos, N.K.; Fauske, A.K.; Teige, J.; Sørum, H. Easily fermentable carbohydrates reduce skatole formation in the distal intestine of entire male pigs. Livestock Sci. 2011, 140, 206–217. [Google Scholar] [CrossRef]
- Liu, S.-Q.; Holland, R.; Crow, V.L. Esters and their biosynthesis in fermented dairy products: A review. Int. Dairy J. 2004, 14, 923–945. [Google Scholar] [CrossRef]
- Pang, X.-N.; Chen, C.; Huang, X.-N.; Yan, Y.-Z.; Chen, J.-Y.; Han, B.-Z. Influence of indigenous lactic acid bacteria on the volatile flavor profile of light-flavor Baijiu. LWT 2021, 147, 111540. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Treatment | Milliliters of Biogas 2 | µg L−1 CO2 3 | µg L−1 CH4 3 |
|---|---|---|---|
| Glucose | |||
| Control | 175 ± 12.3 | 591,000 ± 31,400 | 163,300 ± 39,600 |
| 6.5 g L−1 | 190 ± 8.4 | 779,000 ± 66,000 | 84,500 ± 5640 |
| 13.2 g L−1 | 329 ± 16.0 | 773,000 ± 52,900 | 28,700 ± 3690 |
| 26.3 g L−1 | 230 ± 77.5 | 751,000 ± 75,600 | 34,200 ± 23,400 |
| 65.8 g L−1 | 213 ± 21.8 | 579,000 ± 393,000 | 21,200 ± 14,900 |
| Lactose | |||
| Control | 330 ± 34.3 | 561,000 ± 91,000 | 69,000 ± 24,300 |
| 6.5 g L−1 | 311 ± 37.4 | 542,000 ± 76,500 | 8470 ± 1460 |
| 13.2 g L−1 | 301 ± 95.0 | 533,000 ± 61,200 | 7760 ± 3550 |
| 26.3 g L−1 | 366 ± 48.5 | 569,000 ± 59,800 | 7320 ± 3080 |
| 65.8 g L−1 | 497 ± 171 | 638,000 ± 89,600 | 5530 ± 4090 |
| Sucrose | |||
| Control | 411 ± 162 | 649,000 ± 43,200 | 164,000 ± 17,300 |
| 12.5 g L−1 | 335 ± 38.0 | 846,000 ± 139,000 | 56,800 ± 21,100 |
| 25 g L−1 | 350 ± 194 | 869,000 ± 80,700 | 47,400 ± 15,600 |
| 50 g L−1 | 204 ± 55.2 | 832,000 ± 157,000 | 33,700 ± 10,900 |
| 125 g L−1 | 205 ± 14.8 | 745,000 ± 131,000 | 34,400 ± 10,200 |
| Treatment | |||||
|---|---|---|---|---|---|
| Glucose Addition (g L−1) | |||||
| 0 | 6.5 | 13.2 | 26.3 | 65.8 | |
| Acid Concentration (mg L−1) | |||||
| Lactic | 44.5 | 289 | 287 | 19,800 | 35,700 |
| Formic | nd 2 | nd | nd | nd | nd |
| Acetic | 1010 | 4650 | 2720 | 3900 | 3870 |
| Propionic | 201 | 2300 | 970 | 1940 | 1680 |
| Butyric | 306 | 2470 | 1000 | 1640 | 1370 |
| iso-Butyric | 408 | 346 | 4760 | 1 | 159 |
| Total identified | 1970 | 10,100 | 9740 | 27,300 | 42,800 |
| Lactose Addition (g L−1) | |||||
| 0 | 6.5 | 13.2 | 26.3 | 65.8 | |
| Lactic | 960 | 513 | 495 | 7260 | 29,300 |
| Formic | nd | nd | nd | 213 | nd |
| Acetic | 3830 | 7480 | 7110 | 12,500 | 15,100 |
| Propionic | 3030 | 4010 | 2670 | 1570 | 2010 |
| Butyric | 2730 | 2040 | 4059 | 1390 | 283 |
| iso-Butyric | 831 | 1230 | 1460 | nd | 2500 |
| Total identified | 11,400 | 15,300 | 15,800 | 22,900 | 49,200 |
| Sucrose Addition (g L−1) | |||||
| 0 | 12.5 | 25 | 50 | 125 | |
| Lactic | nd | 463 | 7330 | 29,500 | 44,900 |
| Formic | nd | nd | 120 | nd | nd |
| Acetic | 7620 | 5590 | 10,900 | 7950 | 6340 |
| Propionic | 3320 | 3060 | 1360 | 1230 | 816 |
| Butyric | 340 | 4870 | 2411 | 920 | 667 |
| iso-Butyric | 2150 | 555 | nd | 1050 | 1490 |
| Total identified | 13,400 | 14,500 | 22,100 | 40,700 | 54,200 |
| Treatment 1 | |||||
|---|---|---|---|---|---|
| Compound | Glucose concentration (g L−1) | ||||
| 0 | 6.5 | 13.2 | 26.3 | 65.8 | |
| Manure malodorants, ng mL−1 | |||||
| Dimethyl disulfide | 2.25 ± 1.50 b | 288 ± 179 a | 362 ± 306 a | 173 ± 332 ab | 3.0 ± 2.0 b |
| Phenol | 37.0 ± 17.2 a | 36.3 ± 14.3 a | 37.0 ± 13.8 a | 22.5 ± 16.7 a | 26.8 ± 14.9 a |
| p-Cresol | 472 ± 127 a | 289 ± 85.8 ab | 323 ± 121 ab | 185 ± 126 bc | 130 ± 94.4 c |
| p-Ethylphenol | 36.0 ± 14.0 a | 25.5 ± 15.8 a | 32.7 ± 12.3 a | 39.5 ± 37.6 a | 20.8 ± 5.1 a |
| p-Propylphenol | 131 ± 38.6 a | 69.5 ± 45.2 b | 19.3 ± 4.73 c | 14.8 ± 10.4 c | 20.8 ± 5.1 c |
| Indole | 3.5 ± 0.6 b | 2.0 ± 0 bc | 5.0 ± 1.7 a | 0.8 ± 1.0 c | nd 2 c |
| Skatole | 6.5 ± 0.6 b | 14.0 ± 4.2 a | 6.0 ± 3.6 b | 3.3 ± 2.4 b | 2.0 ± 2.3 b |
| Total identified | 688 ± 195 a | 724 ± 78.1 a | 785 ± 171 a | 439 ± 419 ab | 203 ± 120 b |
| Aliphatic esters, ng mL−1 | |||||
| Ethyl butyrate | nd b | nd b | nd b | 529 ± 140 a | 556 ± 197 a |
| Propyl propanoate | nd b | nd b | nd b | 158 ±141 a | 220 ± 129 a |
| Propyl butyrate | nd b | nd b | nd b | 139 ± 88.3 a | 88.3 ± 19.8 a |
| Butyl butyrate | nd b | nd b | nd b | 20.8 ± 14.0 a | 16.8 ± 15.3 a |
| Ethyl hexanoate | nd b | nd b | nd b | 111 ± 118 ab | 29.3 ± 24.1 a |
| Total identified | 0 b | 0 b | 0 b | 958 ± 771 a | 910 ± 99.1 a |
| Treatment 1 | |||||
|---|---|---|---|---|---|
| Compound | Lactose concentration (g L−1) | ||||
| 0 | 6.5 | 13.2 | 26.3 | 65.8 | |
| Manure malodorants, ng mL−1 | |||||
| Dimethyl disulfide | nd b | 56.8 ± 110 a | 222 ± 152 a | 6.5 ± 1.9 b | 4.5 ± 0.6 b |
| Phenol | 100 ± 24.9 a | 88.0 ± 11.6 a | 81.3 ± 8.4 a | 44.0 ± 18.6 b | 41.5 ± 20.6 b |
| p-Cresol | 364 ± 166 a | 309 ± 168 a | 192 ± 90.8 ab | 108 ± 53.6 b | 95.8 ± 43.3 b |
| p-Ethylphenol | 38.5 ± 11.6 a | 36.0 ± 29.4 a | 36.0 ± 19.1 a | 32.8 ± 13.1 a | 30.3 ± 11.4 a |
| p-Propylphenol | 82.5 ± 7.1 a | 60.0 ± 42.8 ab | 20.3 ± 3.2 b | 38.8 ± 27.2 b | 36.5 ± 24.9 b |
| Indole | 3.3 ± 2.1 a | 3.8 ± 2.2 a | 2.8 ± 2.5 ab | nd 2 b | nd b |
| Skatole | 9.8 ± 7.9 a | 10.3 ± 7.4 a | 1.3 ± 2.3 b | nd b | nd b |
| Total identified | 608 ± 170 a | 564 ± 339 a | 556 ± 72.5 a | 230 ± 74.2 b | 209 ± 60.5 b |
| Aliphatic esters, ng mL−1 | |||||
| Ethyl butyrate | nd | nd | 94.3 ± 164 | 247 ± 79.4 | 188 ± 82.4 |
| Propyl propanoate | nd b | nd b | 22.3 ± 38.7 b | 674 ± 207 a | 514 ± 349 a |
| Propyl butyrate | nd b | nd b | 9.0 ± 15.6 b | 179 ± 79.4 a | 118 ± 82.4 a |
| Butyl butyrate | nd a | nd a | 19.7 ± 34.1 a | 1.8 ± 1.0 a | 1.0 ± 1.2 a |
| Ethyl hexanoate | nd a | nd a | nd a | nd a | 0.8 ± 1.5 a |
| Total identified | 0 b | 0 b | 145 ± 255 a | 1102 ± 246 a | 822 ± 555 a |
| Treatment 1 | |||||
|---|---|---|---|---|---|
| Compound | Sucrose concentration (g L−1) | ||||
| 0 | 12.5 | 25 | 50 | 125 | |
| Manure malodorants, ng mL−1 | |||||
| Dimethyl disulfide | 2.3 ± 1.5 b | 317 ± 280 a | 5.5 ± 4.4 b | 3.5 ±0.6 b | 2.8 ± 1.9 b |
| Phenol | 32.0 ± 14.2 a | 27.3 ± 12.9 a | 17.8 ± 8.5 a | 18.0 ± 8.8 a | 17.0 ± 8.4 a |
| p-Cresol | 442 ± 211 a | 349 ± 187 a | 117 ± 72.5 b | 117 ± 69.5 b | 113 ± 61.9 b |
| p-Ethylphenol | 30.0 ± 11.1 a | 29.3 ± 9.7 a | 23.8 ± 9.7 a | 24.8 ± 9.7 a | 24.5 ± 8.9 a |
| p-Propylphenol | 137 ± 27.1 a | 45.8 ± 18.9 b | 31.5 ± 14.2 b | 31.8 ± 11.9 b | 34.8 ± 11.0 b |
| Indole | 7.3 ± 5.1 a | 3.3 ± 0.5 ab | nd 2 b | nd b | nd b |
| Skatole | 10.3 ± 5.9 a | 12.5 ± 8.7 a | nd b | nd b | nd b |
| Total identified | 661 ± 274 a | 784 ± 401 a | 196 ± 103 b | 195 ± 99.3 b | 202 ± 91.7 b |
| Aliphatic esters, ng mL−1 | |||||
| Ethyl butyrate | nd b | nd b | 512 ± 159 a | 427 ± 49.5 a | 420 ± 13.8 a |
| Propyl propanoate | nd b | nd b | 547 ± 323 a | 133 ± 102 ab | 47.5 ± 36.9 ab |
| Propyl butyrate | nd b | nd b | 454 ± 159 a | 87.3 ± 49.5 b | 43.3 ± 13.8 b |
| Total identified | 0 b | 0 b | 1513 ± 457 a | 647 ± 220 a | 511 ± 113 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loughrin, J.H.; Agga, G.E.; Lovanh, N. Simple Sugars Alter the Odorant Composition of Dairy Cow Manure. Environments 2024, 11, 145. https://doi.org/10.3390/environments11070145
Loughrin JH, Agga GE, Lovanh N. Simple Sugars Alter the Odorant Composition of Dairy Cow Manure. Environments. 2024; 11(7):145. https://doi.org/10.3390/environments11070145
Chicago/Turabian StyleLoughrin, John H., Getahun E. Agga, and Nanh Lovanh. 2024. "Simple Sugars Alter the Odorant Composition of Dairy Cow Manure" Environments 11, no. 7: 145. https://doi.org/10.3390/environments11070145
APA StyleLoughrin, J. H., Agga, G. E., & Lovanh, N. (2024). Simple Sugars Alter the Odorant Composition of Dairy Cow Manure. Environments, 11(7), 145. https://doi.org/10.3390/environments11070145