de novo Blood Biomarkers in Autism: Autoantibodies against Neuronal and Glial Proteins


 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Diagnostic Criteria
2.3. Exclusion Criteria
2.4. Therapeutic Treatments
3. Demographic Information
3.1. Subjects
3.2. Mothers
3.3. Parental Education
3.4. Signs of ASD in Study Children
4. Materials
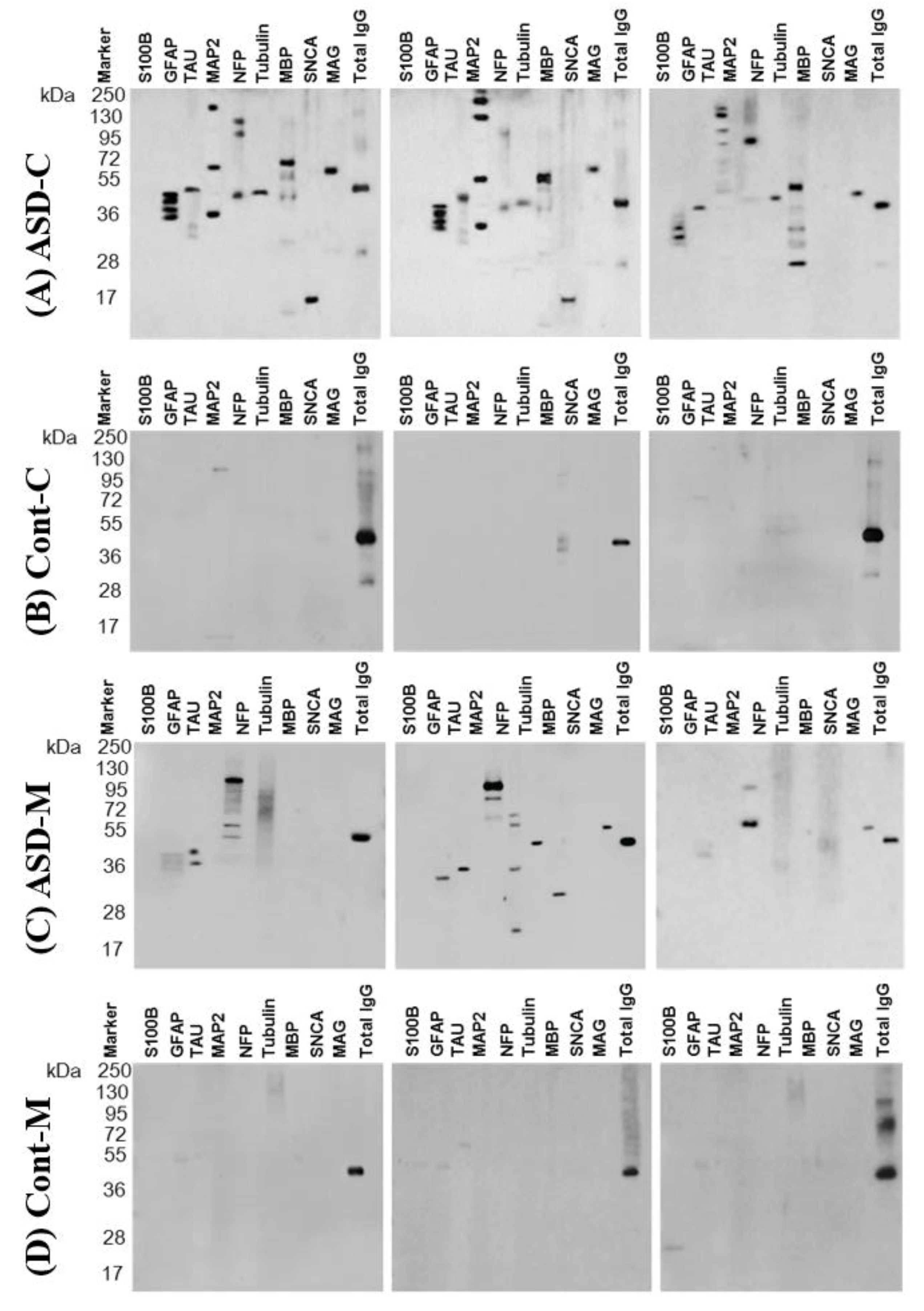
4.1. Western Blot Assay
4.2. Specificity of Serum Autoantibodies
4.3. Calculations
5. Statistics
6. Results
6.1. Autoantibodies against Neuronal and Glial Proteins
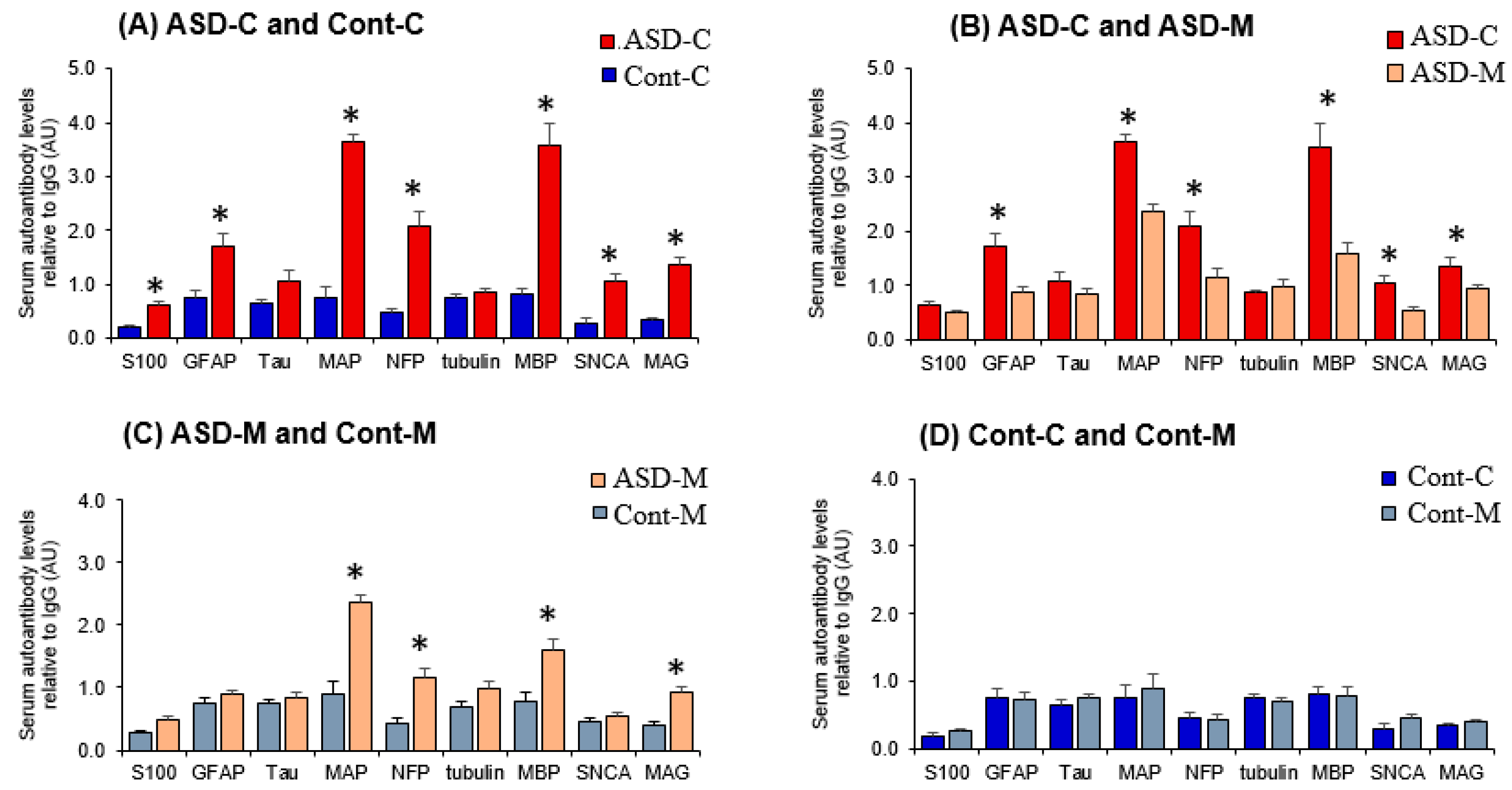
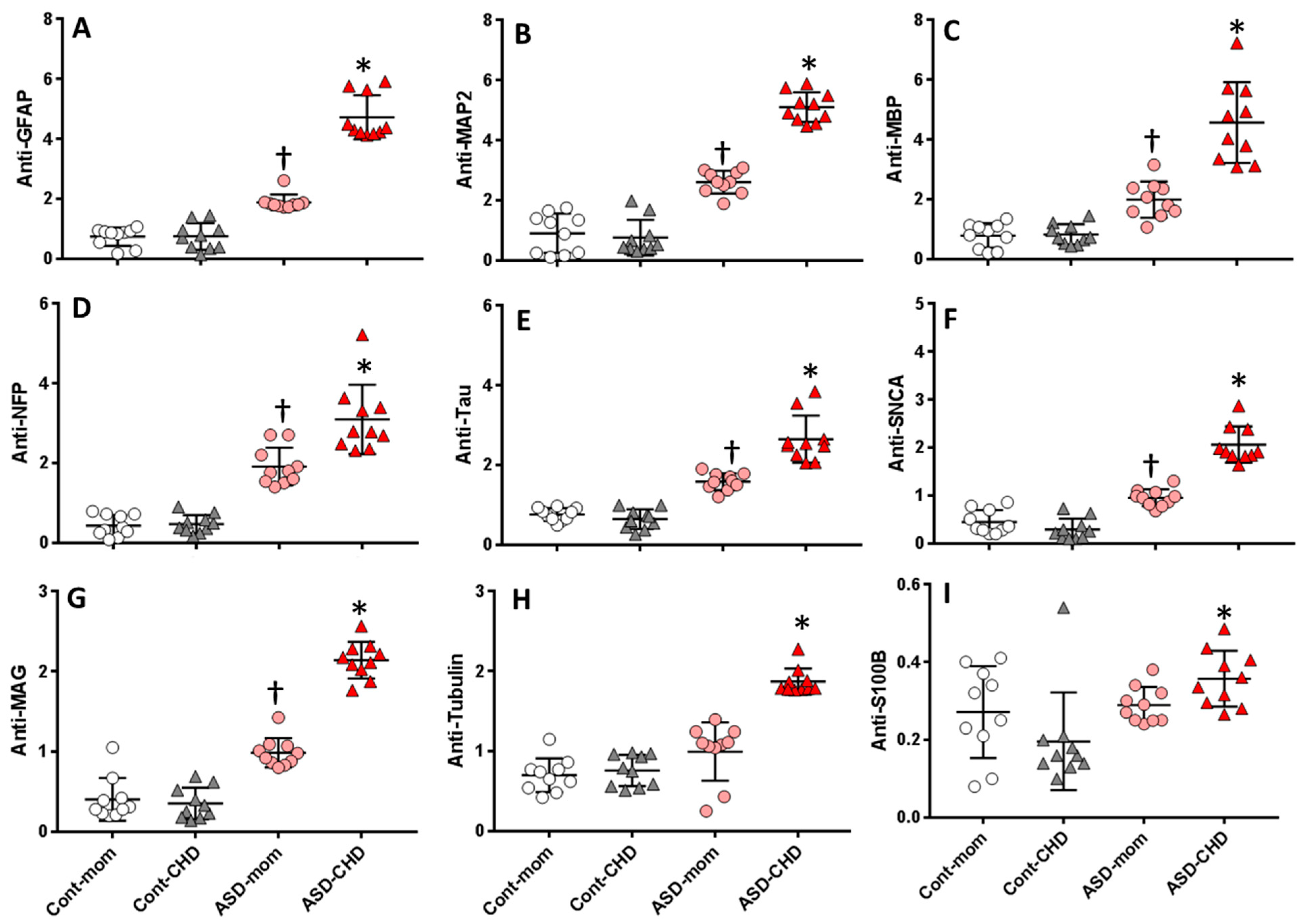
6.2. Levels of Autoantibodies against Neuronal and Glial Proteins in ASD Children and Their Mothers
6.3. Autoantibodies Profile of Neuronal and Glial Proteins in ASD and Control Children and Their Mothers
6.4. Fold Increase in Autoantibodies against Neuronal and Glial Proteins in ASD Children and Their Mothers
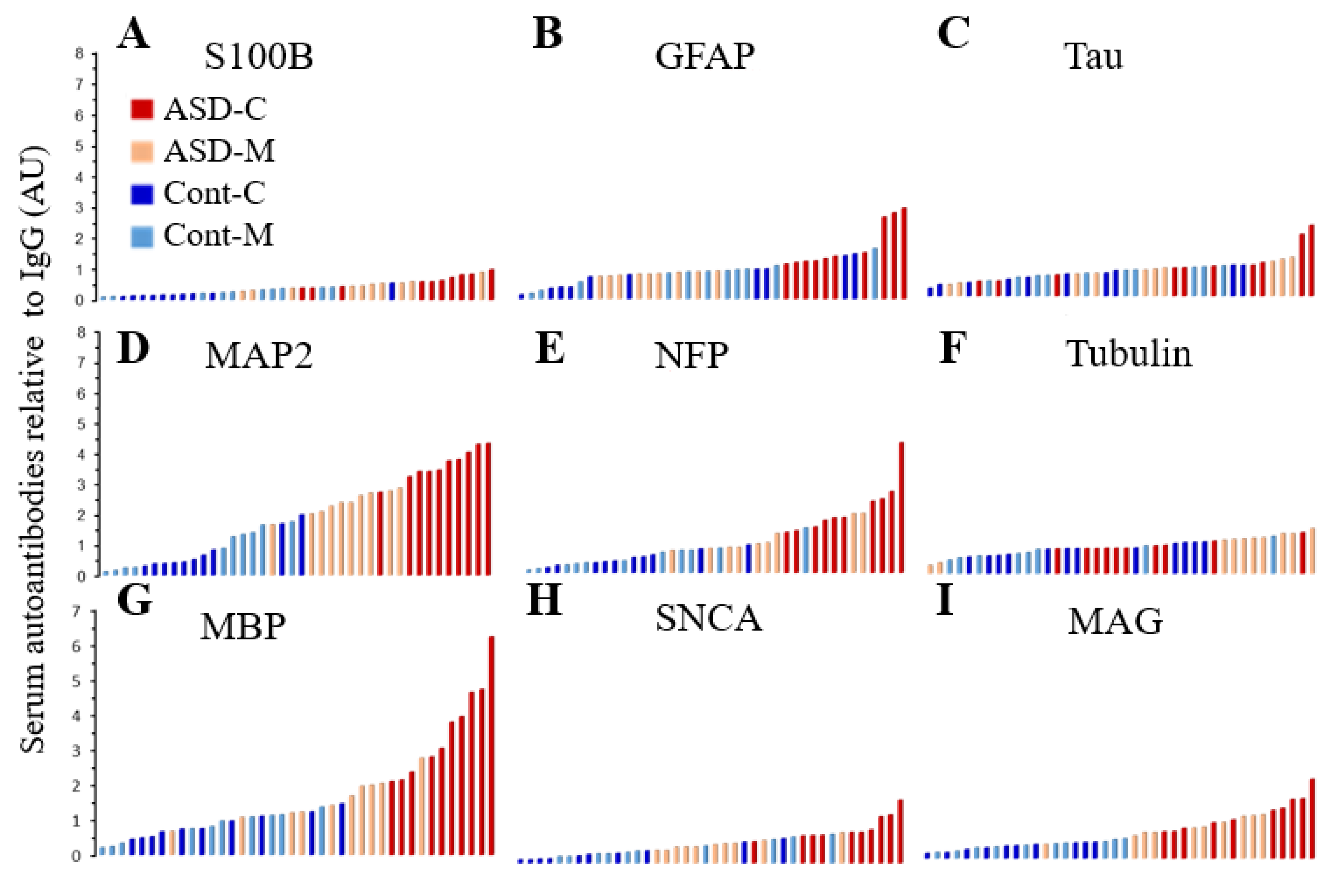
6.5. Distribution of the Biomarkers for ASD Subjects and Controls
7. Discussion
8. Limitations
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| ASD | Autism Spectrum Disorder |
| TD | Typically Developing |
| DD | Developmental Delay |
| ASD-C | Children with Autism Spectrum Disorder |
| ASD-M | Mothers of ASD children |
| Cont-C | Control healthy Children |
| Cont.-M | Mothers of control healthy children |
| AU | Arbitrary Unit |
References
- Gillberg, C.; Wing, L. Autism: Not an extremely rare disorder. Acta Psychiatr. Scand. 1999, 99, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.P.; Myers, S.M. American Academy of Pediatrics Council on Children with D. Identification and evaluation of children with autism spectrum disorders. Pediatrics 2007, 120, 1183–1215. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Fombonne, E. Pervasive developmental disorders in preschool children. JAMA 2001, 285, 3093–3099. [Google Scholar] [CrossRef] [PubMed]
- Spence, S.J.; Schneider, M.T. The role of epilepsy and epileptiform EEGs in autism spectrum disorders. Pediatr. Res. 2009, 65, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Richdale, A.L.; Schreck, K.A. Sleep problems in autism spectrum disorders: Prevalence, nature, & possible biopsychosocial aetiologies. Sleep Med. Rev. 2009, 13, 403–411. [Google Scholar]
- Gorrindo, P.; Williams, K.C.; Lee, E.B.; Walker, L.S.; McGrew, S.G.; Levitt, P. Gastrointestinal dysfunction in autism: Parental report, clinical evaluation, and associated factors. Autism Res. Off. J. Int. Soc. Autism Res. 2012, 5, 101–108. [Google Scholar] [CrossRef]
- Miles, J.H. Autism spectrum disorders—A genetics review. Genet. Med. Off. J. Am. College Med. Genet. 2011, 13, 278–294. [Google Scholar] [CrossRef]
- Miller, M.T.; Stromland, K.; Ventura, L.; Johansson, M.; Bandim, J.M.; Gillberg, C. Autism associated with conditions characterized by developmental errors in early embryogenesis: A mini review. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 2005, 23, 201–219. [Google Scholar] [CrossRef]
- Dufour-Rainfray, D.; Vourc’h, P.; Tourlet, S.; Guilloteau, D.; Chalon, S.; Andres, C.R. Fetal exposure to teratogens: Evidence of genes involved in autism. Neurosci. Biobehav. Rev. 2011, 35, 1254–1265. [Google Scholar] [CrossRef] [PubMed]
- Rodier, P.M. Converging evidence for brain stem injury in autism. Dev. Psychopathol. 2002, 14, 537–557. [Google Scholar] [CrossRef]
- Moore, S.J.; Turnpenny, P.; Quinn, A.; Glover, S.; Lloyd, D.J.; Montgomery, T.; Dean, J.C. A clinical study of 57 children with fetal anticonvulsant syndromes. J. Med. Genet. 2000, 37, 489–497. [Google Scholar] [CrossRef]
- Stromland, K.; Nordin, V.; Miller, M.; Akerstrom, B.; Gillberg, C. Autism in thalidomide embryopathy: A population study. Dev. Med. Child Neurol. 1994, 36, 351–356. [Google Scholar] [CrossRef]
- Malkova, N.V.; Yu, C.Z.; Hsiao, E.Y.; Moore, M.J.; Patterson, P.H. Maternal immune activation yields offspring displaying mouse versions of the three core symptoms of autism. Brain Behav. Immunity 2012, 26, 607–616. [Google Scholar] [CrossRef]
- Mazina, V.; Gerdts, J.; Trinh, S.; Ankenman, K.; Ward, T.; Dennis, M.Y.; Girirajan, S.; Eichler, E.E.; Bernier, R. Epigenetics of autism-related impairment: Copy number variation and maternal infection. J. Dev. Behav. Pediatr. JDBP 2015, 36, 61–67. [Google Scholar] [CrossRef]
- Lee, B.K.; Magnusson, C.; Gardner, R.M.; Blomstrom, A.; Newschaffer, C.J.; Burstyn, I.; Karlsson, H.; Dalman, C. Maternal hospitalization with infection during pregnancy and risk of autism spectrum disorders. Brain Behav. Immunity 2015, 44, 100–105. [Google Scholar] [CrossRef]
- Zerbo, O.; Iosif, A.M.; Walker, C.; Ozonoff, S.; Hansen, R.L.; Hertz-Picciotto, I. Is maternal influenza or fever during pregnancy associated with autism or developmental delays? Results from the CHARGE (CHildhood Autism Risks from Genetics and Environment) study. J. Autism Dev. Disord. 2013, 43, 25–33. [Google Scholar]
- Torrey, E.F.; Hersh, S.P.; McCabe, K.D. Early childhood psychosis and bleeding during pregnancy. A prospective study of gravid women and their offspring. J. Autism Child. Schizophr. 1975, 5, 287–297. [Google Scholar] [CrossRef]
- Yamashita, Y.; Fujimoto, C.; Nakajima, E.; Isagai, T.; Matsuishi, T. Possible association between congenital cytomegalovirus infection and autistic disorder. J. Autism Dev. Disord. 2003, 33, 455–459. [Google Scholar] [CrossRef]
- Shelton, J.F.; Geraghty, E.M.; Tancredi, D.J.; Delwiche, L.D.; Schmidt, R.J.; Ritz, B.; Hansen, R.L.; Hertz-Picciotto, I. Neurodevelopmental disorders and prenatal residential proximity to agricultural pesticides: The CHARGE study. Environ. Health Perspect. 2014, 122, 1103–1109. [Google Scholar] [CrossRef]
- Arndt, T.L.; Stodgell, C.J.; Rodier, P.M. The teratology of autism. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 2005, 23, 189–199. [Google Scholar] [CrossRef]
- Geschwind, D.H. Advances in autism. Annu. Rev. Med. 2009, 60, 367–380. [Google Scholar] [CrossRef]
- Stoner, R.; Chow, M.L.; Boyle, M.P.; Sunkin, S.M.; Mouton, P.R.; Roy, S.; Wynshaw-Boris, A.; Colamarino, S.A.; Lein, E.S.; Courchesne, E. Patches of disorganization in the neocortex of children with autism. N. Engl. J. Med. 2014, 370, 1209–1219. [Google Scholar] [CrossRef] [PubMed]
- Kern, J.K.; Geier, D.A.; Sykes, L.K.; Geier, M.R. Evidence of neurodegeneration in autism spectrum disorder. Transl. Neurodegener. 2013, 2, 17. [Google Scholar] [CrossRef]
- Braunschweig, D.; Van de Water, J. Maternal autoantibodies in autism. Arch. Neurol. 2012, 69, 693–699. [Google Scholar] [CrossRef]
- Siniscalco, D.; Sapone, A.; Giordano, C.; Cirillo, A.; de Magistris, L.; Rossi, F.; Fasano, A.; Bradstreet, J.J.; Maione, S.; Antonucci, N. Cannabinoid receptor type 2, but not type 1, is up-regulated in peripheral blood mononuclear cells of children affected by autistic disorders. J. Autism Dev. Disord. 2013, 43, 2686–2695. [Google Scholar] [CrossRef] [PubMed]
- Abou-Donia, M.B.; Abou-Donia, M.M.; ElMasry, E.M.; Monro, J.A.; Mulder, M.F. Autoantibodies to nervous system-specific proteins are elevated in sera of flight crew members: Biomarkers for nervous system injury. J. Toxicol. Environ. Health Part A 2013, 76, 363–380. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, G.A.; El-Sherif, D.F.; Al-Ayadhi, L.Y. Systemic auto-antibodies in children with autism. J. Neuroimmunol. 2014, 272, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, A.W.; Connors, S.L.; Matteson, K.J.; Lee, L.C.; Singer, H.S.; Castaneda, J.A.; Pearce, D.A. Maternal antibrain antibodies in autism. Brain Behav. Immunity 2007, 21, 351–357. [Google Scholar] [CrossRef]
- Wills, S.; Rossi, C.C.; Bennett, J.; Martinez-Cerdeno, V.; Ashwood, P.; Amaral, D.G.; Van de Water, J. Further characterization of autoantibodies to GABAergic neurons in the central nervous system produced by a subset of children with autism. Mol. Autism 2011, 2, 5. [Google Scholar] [CrossRef]
- Abdel-Rahman, A.; Shetty, A.K.; Abou-Donia, M.B. Subchronic dermal application of N,N-diethyl m-toluamide (DEET) and permethrin to adult rats, alone or in combination, causes diffuse neuronal cell death and cytoskeletal abnormalities in the cerebral cortex and the hippocampus, and Purkinje neuron loss in the cerebellum. Exp. Neurol. 2001, 172, 153–171. [Google Scholar]
- Abou-Donia, M.B.; Conboy, L.A.; Kokkotou, E.; Jacobson, E.; Elmasry, E.M.; Elkafrawy, P.; Neely, M.; Bass, C.R.; Sullivan, K. Screening for novel central nervous system biomarkers in veterans with Gulf War Illness. Neurotoxicol. Teratol. 2017, 61, 36–46. [Google Scholar] [CrossRef]
- Abou-Donia, M.B.; Lieberman, A.; Curtis, L. Neural autoantibodies in patients with neurological symptoms and histories of chemical/mold exposures. Toxicol. Ind. Health 2018, 34, 44–53. [Google Scholar] [CrossRef]
- Reneman, L.; Schagen, S.B.; Mulder, M.; Mutsaerts, H.J.; Hageman, G.; de Ruiter, M.B. Cognitive impairment and associated loss in brain white microstructure in aircrew members exposed to engine oil fumes. Brain Imaging Behav. 2016, 10, 437–444. [Google Scholar] [CrossRef]
- Nordahl, C.W.; Braunschweig, D.; Iosif, A.M.; Lee, A.; Rogers, S.; Ashwood, P.; Amaral, D.G.; Van de Water, J. Maternal autoantibodies are associated with abnormal brain enlargement in a subgroup of children with autism spectrum disorder. Brain Behav. Immunity 2013, 30, 61–65. [Google Scholar] [CrossRef]
- Hashemi, E.; Ariza, J.; Lechpammer, M.; Noctor, S.C.; Martinez-Cerdeno, V. Abnormal white matter tracts resembling pencil fibers involving prefrontal cortex (Brodmann area 47) in autism: A case report. J. Med. Case Rep. 2016, 10, 237. [Google Scholar] [CrossRef]
- Dajani, D.R.; Uddin, L.Q. Local brain connectivity across development in autism spectrum disorder: A cross-sectional investigation. Autism Res. Off. J. Int. Soc. Autism Res. 2016, 9, 43–54. [Google Scholar] [CrossRef]
- Tagliaferro, P.; Javier Ramos, A.; Onaivi, E.S.; Evrard, S.G.; Lujilde, J.; Brusco, A. Neuronal cytoskeleton and synaptic densities are altered after a chronic treatment with the cannabinoid receptor agonist WIN 55,212-2. Brain Res. 2006, 1085, 163–176. [Google Scholar] [CrossRef]
- Liliang, P.C.; Liang, C.L.; Weng, H.C.; Lu, K.; Wang, K.W.; Chen, H.J.; Chuang, J.H. Tau proteins in serum predict outcome after severe traumatic brain injury. J. Surg. Res. 2010, 160, 302–307. [Google Scholar] [CrossRef]
- Salama, M.; Shalash, A.; Magdy, A.; Makar, M.; Roushdy, T.; Elbalkimy, M.; Elrassas, H.; Elkafrawy, P.; Mohamed, W.; Abou Donia, M.B. Tubulin and Tau: Possible targets for diagnosis of Parkinson’s and Alzheimer’s diseases. PLoS ONE 2018, 13, e0196436. [Google Scholar] [CrossRef]
- Jauch, E.C.; Lindsell, C.; Broderick, J.; Fagan, S.C.; Tilley, B.C.; Levine, S.R.; Group Nr-PSS. Association of serial biochemical markers with acute ischemic stroke: The National Institute of Neurological Disorders and Stroke recombinant tissue plasminogen activator Stroke Study. Stroke 2006, 37, 2508–2513. [Google Scholar] [CrossRef]
- Bido, S.; Soria, F.N.; Fan, R.Z.; Bezard, E.; Tieu, K. Mitochondrial division inhibitor-1 is neuroprotective in the A53T-alpha-synuclein rat model of Parkinson’s disease. Sci. Rep. 2017, 7, 7495. [Google Scholar] [CrossRef]
- Middeldorp, J.; Hol, E.M. GFAP in health and disease. Prog. Neurobiol. 2011, 93, 421–443. [Google Scholar] [CrossRef]
- Donato, R. S100: A multigenic family of calcium-modulated proteins of the EF-hand type with intracellular and extracellular functional roles. Int. J. Biochem. Cell Biol. 2001, 33, 637–668. [Google Scholar] [CrossRef]
- Geier, D.A.; Audhya, T.; Kern, J.K.; Geier, M.R. Blood mercury levels in autism spectrum disorder: Is there a threshold level? Acta Neurobiol. Exp. 2010, 70, 177–186. [Google Scholar]
- Yochum, C.L.; Bhattacharya, P.; Patti, L.; Mirochnitchenko, O.; Wagner, G.C. Animal model of autism using GSTM1 knockout mice and early post-natal sodium valproate treatment. Behav. Brain Res. 2010, 210, 202–210. [Google Scholar] [CrossRef]
- Ballough, G.P.; Martin, L.J.; Cann, F.J.; Graham, J.S.; Smith, C.D.; Kling, C.E.; Forster, J.S.; Phann, S.; Filbert, M.G. Microtubule-associated protein 2 (MAP-2): A sensitive marker of seizure-related brain damage. J. Neurosci. Methods 1995, 61, 23–32. [Google Scholar] [CrossRef]
- Jensen, K.F.; Lapadula, D.M.; Anderson, J.K.; Haykal-Coates, N.; Abou-Donia, M.B. Anomalous phosphorylated neurofilament aggregations in central and peripheral axons of hens treated with tri-ortho-cresyl phosphate (TOCP). J. Neurosci. Res. 1992, 33, 455–460. [Google Scholar] [CrossRef]
- Zanier, E.R.; Refai, D.; Zipfel, G.J.; Zoerle, T.; Longhi, L.; Esparza, T.J.; Spinner, M.L.; Bateman, R.J.; Brody, D.L.; Stocchetti, N. Neurofilament light chain levels in ventricular cerebrospinal fluid after acute aneurysmal subarachnoid haemorrhage. J. Neurol. Neurosurg. Psychiatry 2011, 82, 157–159. [Google Scholar] [CrossRef]
- Arolt, V.; Peters, M.; Erfurth, A.; Wiesmann, M.; Missler, U.; Rudolf, S.; Kirchner, H.; Rothermundt, M. S100B and response to treatment in major depression: A pilot study. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2003, 13, 235–239. [Google Scholar] [CrossRef]
- Grabe, H.J.; Ahrens, N.; Rose, H.J.; Kessler, C.; Freyberger, H.J. Neurotrophic factor S100 beta in major depression. Neuropsychobiology 2001, 44, 88–90. [Google Scholar] [CrossRef]
- Machado-Vieira, R.; Lara, D.R.; Portela, L.V.; Goncalves, C.A.; Soares, J.C.; Kapczinski, F.; Souza, D.O. Elevated serum S100B protein in drug-free bipolar patients during first manic episode: A pilot study. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2002, 12, 269–272. [Google Scholar] [CrossRef]
- Zurek, J.; Fedora, M. The usefulness of S100B, NSE, GFAP, NF-H, secretagogin and Hsp70 as a predictive biomarker of outcome in children with traumatic brain injury. Acta Neurochir. 2012, 154, 93–103, discussion 103. [Google Scholar] [CrossRef]
- Goines, P.; Van de Water, J. The immune system’s role in the biology of autism. Curr. Opin. Neurol. 2010, 23, 111–117. [Google Scholar] [CrossRef]
- Vargas, D.L.; Nascimbene, C.; Krishnan, C.; Zimmerman, A.W.; Pardo, C.A. Neuroglial activation and neuroinflammation in the brain of patients with autism. Ann. Neurol. 2005, 57, 67–81. [Google Scholar] [CrossRef]
- Braunschweig, D.; Ashwood, P.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.; Croen, L.A.; Pessah, I.N.; Van de Water, J. Autism: Maternally derived antibodies specific for fetal brain proteins. Neurotoxicology 2008, 29, 226–231. [Google Scholar] [CrossRef]
- Braunschweig, D.; Krakowiak, P.; Duncanson, P.; Boyce, R.; Hansen, R.L.; Ashwood, P.; Hertz-Picciotto, I.; Pessah, I.N.; Van de Water, J. Autism-specific maternal autoantibodies recognize critical proteins in developing brain. Transl. Psychiatry 2013, 3, e277. [Google Scholar] [CrossRef]
- Piras, I.S.; Haapanen, L.; Napolioni, V.; Sacco, R.; Van de Water, J.; Persico, A.M. Anti-brain antibodies are associated with more severe cognitive and behavioral profiles in Italian children with Autism Spectrum Disorder. Brain Behav. Immunity 2014, 38, 91–99. [Google Scholar] [CrossRef]
- Abou-Donia, M.B. Organophosphorus ester-induced chronic neurotoxicity. Arch. Environ. Health 2003, 58, 484–497. [Google Scholar] [CrossRef]
- Rosas, L.G.; Eskenazi, B. Pesticides and child neurodevelopment. Curr. Opin. Pediatr. 2008, 20, 191–197. [Google Scholar] [CrossRef]
- De Felice, A.; Greco, A.; Calamandrei, G.; Minghetti, L. Prenatal exposure to the organophosphate insecticide chlorpyrifos enhances brain oxidative stress and prostaglandin E2 synthesis in a mouse model of idiopathic autism. J. Neuroinflamm. 2016, 13, 149. [Google Scholar] [CrossRef]
- Karr, C.J.; Solomon, G.M.; Brock-Utne, A.C. Health effects of common home, lawn, and garden pesticides. Pediatr. Clin. N. Am. 2007, 54, 63–80. [Google Scholar] [CrossRef]
- Shaw, W. Elevated Urinary Glyphosate and Clostridia Metabolites with Altered Dopamine Metabolism in Triplets with Autistic Spectrum Disorder or Suspected Seizure Disorder: A Case Study. Integr. Med. 2017, 16, 50–57. [Google Scholar]
- Roberts, E.M.; English, P.B.; Grether, J.K.; Windham, G.C.; Somberg, L.; Wolff, C. Maternal residence near agricultural pesticide applications and autism spectrum disorders among children in the California Central Valley. Environ. Health Perspect. 2007, 115, 1482–1489. [Google Scholar] [CrossRef]
- Singer, H.S.; Morris, C.M.; Gause, C.D.; Gillin, P.K.; Crawford, S.; Zimmerman, A.W. Antibodies against fetal brain in sera of mothers with autistic children. J. Neuroimmunol. 2008, 194, 165–172. [Google Scholar] [CrossRef]
- Goines, P.; Haapanen, L.; Boyce, R.; Duncanson, P.; Braunschweig, D.; Delwiche, L.; Hansen, R.; Hertz-Picciotto, I.; Ashwood, P.; Van de Water, J. Autoantibodies to cerebellum in children with autism associate with behavior. Brain Behav. Immun. 2011, 25, 514–523. [Google Scholar] [CrossRef]
- Kalcheva, N.; Albala, J.; O’Guin, K.; Rubino, H.; Garner, C.; Shafit-Zagardo, B. Genomic structure of human microtubule-associated protein 2 (MAP-2) and characterization of additional MAP-2 isoforms. Proc. Natl. Acad. Sci. USA 1995, 92, 10894–10898. [Google Scholar] [CrossRef]
- Burre, J.; Vivona, S.; Diao, J.; Sharma, M.; Brunger, A.T.; Sudhof, T.C. Properties of native brain alpha-synuclein. Nature 2013, 498, E4–E6, discussion E6–E7. [Google Scholar] [CrossRef]
- Gilquin, B.; Cannon, B.R.; Hubstenberger, A.; Moulouel, B.; Falk, E.; Merle, N.; Assard, N.; Kieffer, S.; Rousseau, D.; Wilder, P.T.; et al. The calcium-dependent interaction between S100B and the mitochondrial AAA ATPase ATAD3A and the role of this complex in the cytoplasmic processing of ATAD3A. Mol. Cell. Biol. 2010, 30, 2724–2736. [Google Scholar] [CrossRef]
- Wills, S.; Cabanlit, M.; Bennett, J.; Ashwood, P.; Amaral, D.G.; Van de Water, J. Detection of autoantibodies to neural cells of the cerebellum in the plasma of subjects with autism spectrum disorders. Brain Behav. Immun. 2009, 23, 64–74. [Google Scholar] [CrossRef]
- LeBien, T.W.; Tedder, T.F. B lymphocytes: How they develop and function. Blood 2008, 112, 1570–1580. [Google Scholar] [CrossRef]
- Isenbergm, D.A. B-cell targeted therapies in autoimmune diseases. J. Ruuatol. 2008, 33 (Suppl. 77), 24–28. [Google Scholar]
- Edwards, J.C.W.; Szczepanski, L.; Szechinski, J.; Filipowicz-Sosnowska, A.; Emery, P.; Close, D.R.; Stevens, R.M.; Shaw, T. Efficacy of B-cell-targeted therapy with rituximab in patients with rheumatoid arthritis. N. Engl. J. Med. 2004, 350, 2572. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Early Indicators | YES | NO | |
| 1 | No babbling or pointing by age 2 | 2 | 8 |
| 2 | No single words by 16 months or two-word phrases by age 2 | 6 | 4 |
| 3 | No response to name | 7 | 3 |
| 4 | loss of language or social skills | 9 | 1 |
| 5 | Poor eye contact | 5 | 5 |
| 6 | Excessive lining up of toys or object | 3 | 7 |
| 7 | No smiling or social responsiveness. | 7 | 3 |
| Later indicators | YES | NO | |
| 1 | Impaired ability to make friends with peers | 10 | 0 |
| 2 | Impaired ability to initiate or sustain a conversation with others | 8 | 2 |
| 3 | Absence or impairment of imaginative and social play | 8 | 2 |
| 4 | Serotyped, repetitive, or unusual use of language | 10 | 0 |
| 5 | Restricted patterns of interest that are abnormal in intensity or focus | 4 | 6 |
| 6 | Preoccupation with certain objects or subjects | 4 | 6 |
| Neural Protein | Mass kDa | Location in the Brain |
|---|---|---|
| NFL, NFM, NFH | 70,160, 200 | Axon is mainly found in large myelinated axons |
| Tubulin | 55 | Axon consists of 10–20% of soluble protein in the brain, axonal transport |
| Tau | 45–62 | Axon, More abundant in the white matter |
| SNCA | 14 | Axon, a neuroprotective protein, with respect to oxidative stress |
| MAP2 | 200 | Dendrites, Purkinje cells, Cerebellum |
| MBP | 38 | Myelin |
| MAG | 100 | Myelin, near the axon |
| GFAP | 52 | Astrocytes, contribute to white matter, myelination and BBB. forms Glial Scar, |
| S100B | 10–11 | Astrocytes, neurotropic at nanomolar concentration and apoptotic at micromolar |
| Protein Name | ASD-C | Cont.-C | ASD-M | Cont.-M | P Value |
|---|---|---|---|---|---|
| S100B | 0.36 ± 0.06 * | 0.20 ± 0.04 | 0.29 ± 0.06 | 0.27 ± 0.04 | 0.022 |
| GFAP | 4.72 ± 0.23 * | 0.75 ± 0.14 | 1.88 ± 0.08 † | 0.74 ± 0.10 | 0.001 |
| Tau | 2.63 ± 0.19 * | 0.65 ± 0.08 | 1.58 ± 0.09 | 0.76 ± 0.05 | 0.005 |
| MAP2 | 5.09 ± 0.16 * | 0.76 ± 0.19 | 2.84 ± 0.12 † | 0.91 ± 0.21 | 0.001 |
| NFP | 3.10 ± 0.28 * | 0.47 ± 0.07 | 1.92 ± 0.15 † | 0.44 ± 0.08 | 0.001 |
| Tubulin | 1.87 ± 0.05 * | 0.76 ± 0.06 | 0.99 ± 0.12 | 0.70 ± 0.07 | 0.043 |
| MBP | 4.56 ± 0.43 * | 0.83 ± 0.11 | 1.89 ± 0.18 † | 0.79 ± 0.13 | 0.001 |
| SNCA | 2.06 ± 0.12 * | 0.30 ± 0.08 | 0.95 ± 0.05 | 0.45 ± 0.08 | 0.001 |
| MAG | 2.14 ± 0.16 * | 0.35 ± 0.04 | 0.99 ± 0.08 † | 0.40 ± 0.04 | 0.001 |
| Protein | ASD-C Over Cont-C | ASD-M Over Cont-M | P-Value |
|---|---|---|---|
| S100B | 1.83 ± 0.31 * | 1.07 ± 0.21 | 0.022 |
| GFAP | 6.29 ± 0.31 * | 2.54 ± 0.11 * | 0.005 |
| Tau | 4.08 ± 0.29 * | 2.08 ± 0.12 * | 0.005 |
| MAP2 | 6.70 ± 0.21 * | 3.13 ± 0.13 * | 0.001 |
| NFP | 6.57 ± 0.55 * | 4.39 ± 0.33 * | 0.001 |
| tubulin | 2.46 ± 0.07 * | 1.41 ± 0.17 | 0.025 |
| MBP | 5.51 ± 0.71 * | 2.38 ± 0.24 * | 0.026 |
| SNCA | 6.99 ± 0.40 * | 2.13 ± 0.11 * | 0.036 |
| MAG | 6.05 ± 0.54 * | 2.47 ± 0.20 * | 0.001 |
| Protein Mass (kDa) | Corresponding Neural Protein | Location in the Brain | References |
|---|---|---|---|
| 10–11 | S100B | Brain glia and astrocytes | [68] |
| 14 | SNCA | neocortex, hippocampus, substantia nigra, thalamus, cerebellum | [14] |
| 36 | MBP | Cerebellum | [64] |
| 37 | MBP | Cerebellum | [24,55,56,57] |
| 39 | MBP | Cerebellum | [24,55,56,57] |
| 45–62 | Tau | Cerebellum | [24,59,65] |
| 200 | MAP2 | Brain | [66] |
| 52–55 | Tubulin | Cerebellum | [69] |
| 67–71 | MAG | Cerebellum | [24,56] |
| 70–150 | NFP | caudate, putamen and prefrontal cortex | [56,57,64] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abou-Donia, M.B.; Suliman, H.B.; Siniscalco, D.; Antonucci, N.; ElKafrawy, P.; Brahmajothi, M.V. de novo Blood Biomarkers in Autism: Autoantibodies against Neuronal and Glial Proteins. Behav. Sci. 2019, 9, 47. https://doi.org/10.3390/bs9050047
Abou-Donia MB, Suliman HB, Siniscalco D, Antonucci N, ElKafrawy P, Brahmajothi MV. de novo Blood Biomarkers in Autism: Autoantibodies against Neuronal and Glial Proteins. Behavioral Sciences. 2019; 9(5):47. https://doi.org/10.3390/bs9050047
Chicago/Turabian StyleAbou-Donia, Mohamed B., Hagir B. Suliman, Dario Siniscalco, Nicola Antonucci, Passent ElKafrawy, and Mulugu V. Brahmajothi. 2019. "de novo Blood Biomarkers in Autism: Autoantibodies against Neuronal and Glial Proteins" Behavioral Sciences 9, no. 5: 47. https://doi.org/10.3390/bs9050047
APA StyleAbou-Donia, M. B., Suliman, H. B., Siniscalco, D., Antonucci, N., ElKafrawy, P., & Brahmajothi, M. V. (2019). de novo Blood Biomarkers in Autism: Autoantibodies against Neuronal and Glial Proteins. Behavioral Sciences, 9(5), 47. https://doi.org/10.3390/bs9050047