The Other-Race and Other-Species Effect during a Sex Categorization Task: An Eye Tracker Study


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Experiment 1
2.1. Materials and Methods
2.1.1. Participants
2.1.2. Stimuli
2.1.3. Apparatus
2.1.4. Eye Tracking
2.1.5. Procedure
2.2. Results
2.2.1. Behavioral Results
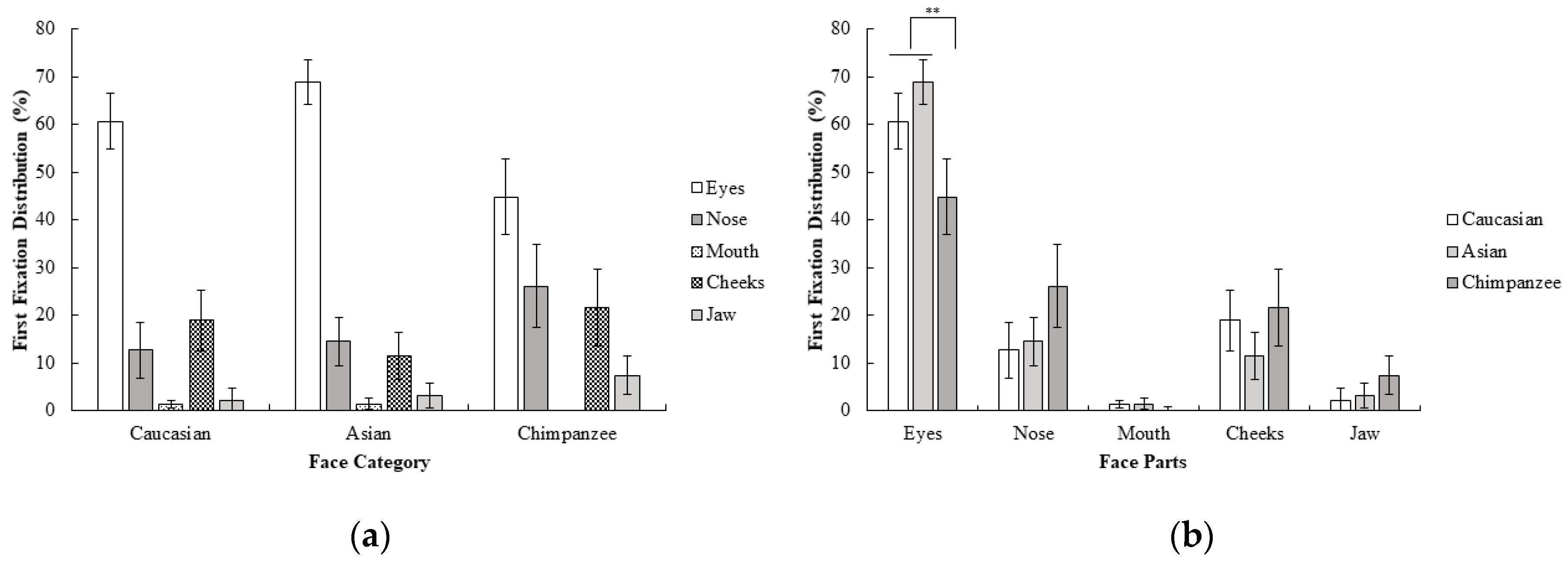
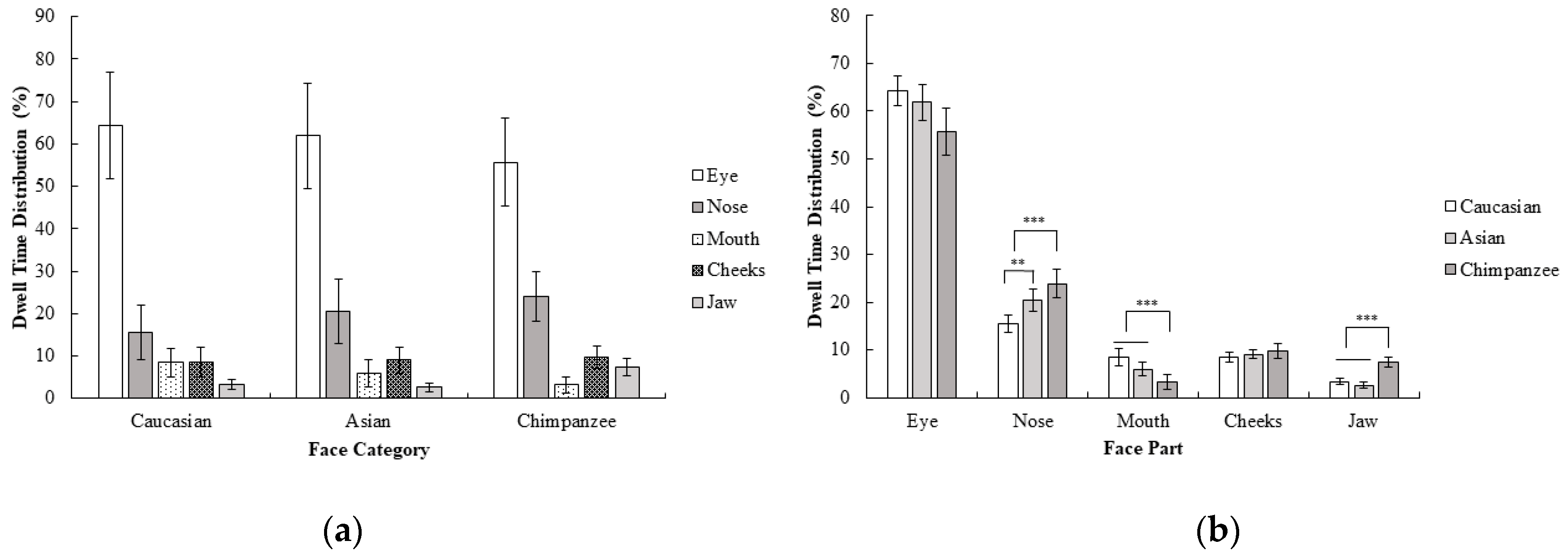
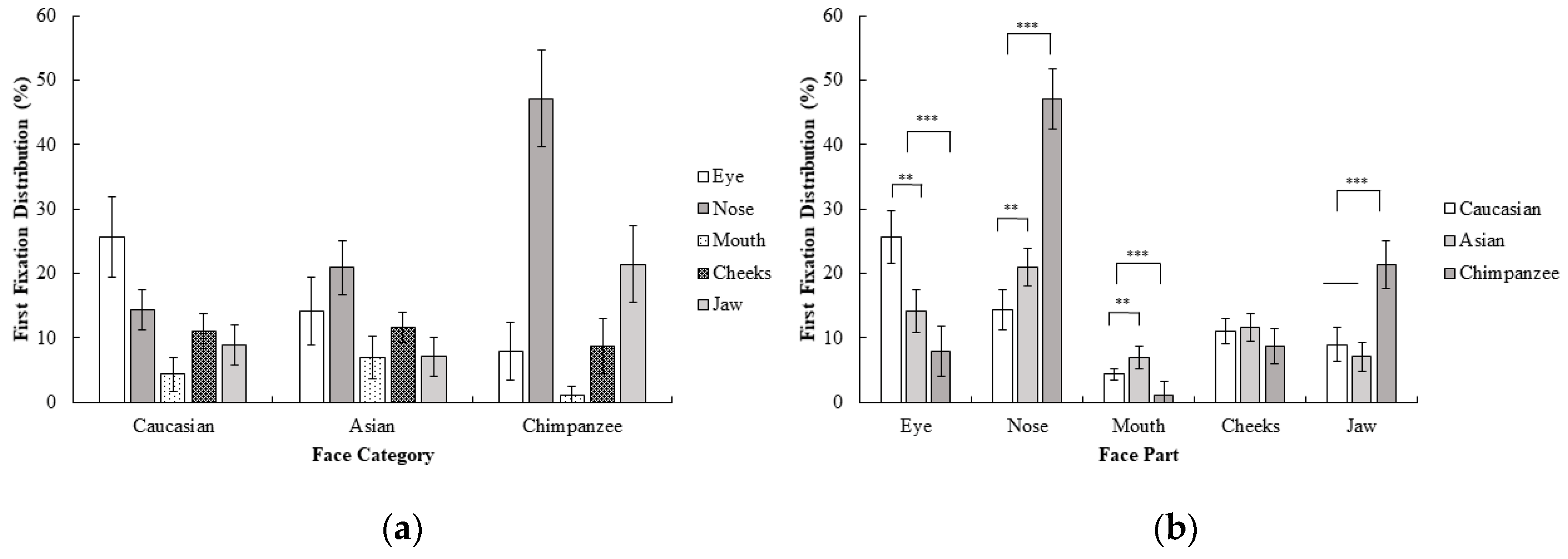
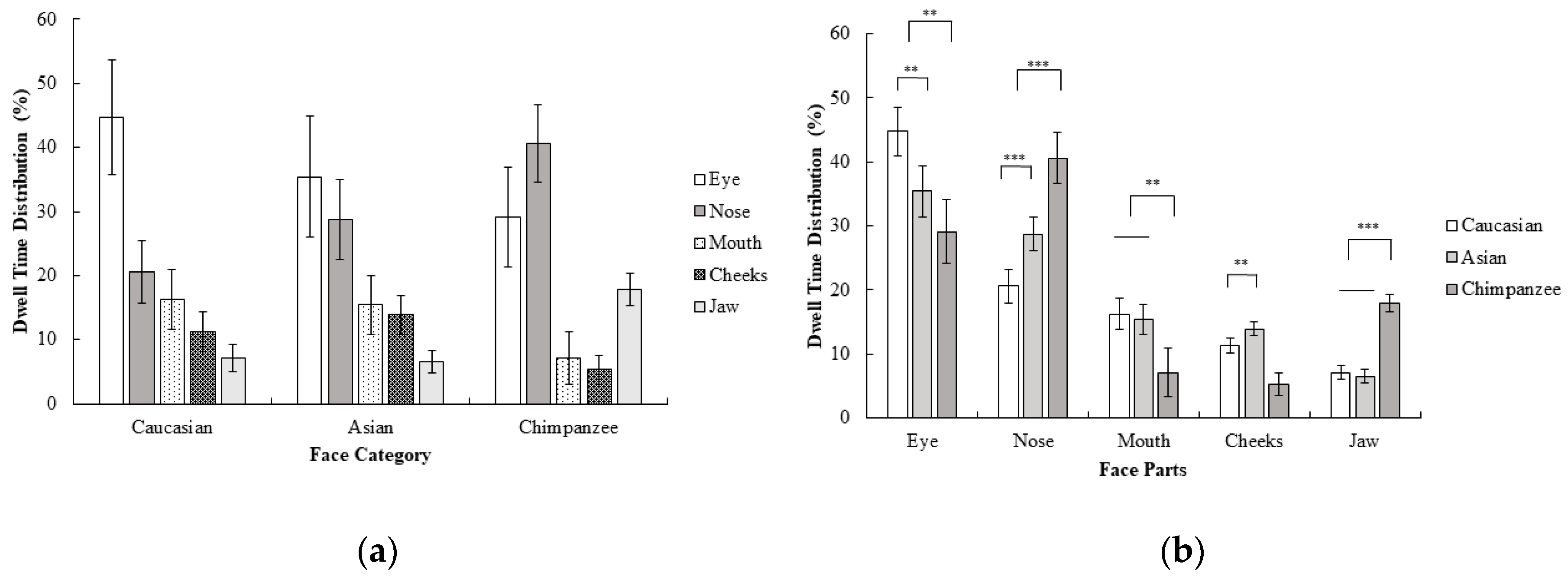
2.2.2. Gaze Behavior
3. Experiment 2
3.1. Materials and Methods
3.1.1. Participants
3.1.2. Procedure
3.2. Results
3.2.1. Behavioral Results
3.2.2. Gaze Behavior
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Parr, L.A. The evolution of face processing in primates. Philos. Trans. R. Soc. B Boil. Sci. 2011, 366, 1764–1777. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.K.; Hirukawa, T.; Kanazawa, S. Judgment of gender through facial parts. Perception 2013, 42, 1253–1265. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, A.J.; Peterson, J.; Deffenbacher, K.A. An “other-race effect” for categorizing faces by sex. Perception 1996, 25, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Reddy, L.; Wilken, P.; Koch, C. Face-gender discrimination is possible in the near-absence of attention. J. Vis. 2004, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Sergent, J.; Ohta, S.; Macdonald, B. Functional Neuroanatomy of Face and Object Processing. Brain 1992, 115, 15–36. [Google Scholar] [CrossRef] [PubMed]
- Schendan, H.E.; Ganis, G.; Kutas, M. Neurophysiological evidence for visual perceptual categorization of words and faces within 150 ms. Psychophysiology 1998, 35, 240–251. [Google Scholar] [CrossRef]
- Cellerino, A.; Borghetti, D.; Sartucci, F. Sex differences in face gender recognition in humans. Brain Res. Bull. 2004, 63, 443–449. [Google Scholar] [CrossRef]
- O’toole, A.J.; Deffenbacher, K.A.; Valentin, D.; Abdi, H. Structural aspects of face recognition and the other-race effect. Mem. Cogn. 1994, 22, 208–224. [Google Scholar] [CrossRef]
- Scott, L.S.; Fava, E. The own-species face bias: A review of developmental and comparative data. Vis. Cogn. 2013, 21, 1364–1391. [Google Scholar] [CrossRef]
- Zhao, L.; Bentin, S. Own- and other-race categorization of faces by race, gender, and age. Psychon. Bull. Rev. 2008, 15, 1093–1099. [Google Scholar] [CrossRef]
- Franklin, R.G.; Zebrowitz, L.A.; Fellous, J.M.; Lee, A. Generalizing from human facial sexual dimorphism to sex-differentiate macaques: Accuracy and cultural variation. J. Exp. Soc. Psychol. 2013, 49, 344–348. [Google Scholar] [CrossRef]
- Little, A.C.; Jones, B.C.; Waitt, C.; Tiddeman, B.P.; Feinberg, D.R.; Perrett, D.I.; Marlow, F.W. Symmetry is related to sexual dimorphism in faces: Data across culture and species. PLoS ONE 2008, 3. [Google Scholar] [CrossRef] [PubMed]
- Schyns, P.G.; Bonnar, L.; Gosselin, F. Show me the features! Understanding recognition from the use of visual information. Psychol. Sci. 2002, 13, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Pearson, A.M.; Henderson, J.M.; Schyns, P.G.; Gosselin, F. Task Dependent Eye Movements during Face Perception. Abstr. Psychon. Soc. 2003, 8, 84. [Google Scholar]
- Sæther, L.; Belle, W.V.; Laeng, B.; Brennen, T.; Øvervoll, M. Anchoring gaze when categorizing faces’ sex: Evidence from eye-tracking data. Vis. Res. 2009, 49, 2870–2880. [Google Scholar] [CrossRef]
- Peterson, M.F.; Eckstein, M.P. Looking just below the eyes is optimal across face recognition tasks. Proc. Natl. Acad. Sci. USA 2012, 109, E3314–E3323. [Google Scholar] [CrossRef]
- Blais, C.; Jack, R.E.; Scheepers, C.; Fiset, D.; Caldara, R. Culture shapes how we look at faces. PLoS ONE 2008, 3. [Google Scholar] [CrossRef]
- Watson, T.; Robins, R. The nature of holistic processing in face and object recognition: Current opinions. Front. Psychol. 2014, 5. [Google Scholar] [CrossRef]
- Rossion, B.; Michel, C. An Experience-Based Holistic Account of the Other-Race Face Effect. In Oxford Handbook of Face Perception; Calder, A., Rhodes, G., Johnson, M., Haxby, J., Eds.; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Langner, O.; Dotsch, R.; Bijlstra, G.; Wigboldus, D.H.J.; Hawk, S.T.; van Knippenberg, A. Presentation and validation of the radboud faces database. Cogn. Emot. 2010, 24, 1377–1388. [Google Scholar] [CrossRef]
- Wang, X.; Tang, X. Face photo-sketch synthesis and recognition. IEEE Trans. Pattern Anal. Mach. Intell. 2009, 31, 1955–1967. [Google Scholar] [CrossRef] [PubMed]
- Mollison, J. James Other Apes; Chris Boot: London, UK, 2004; ISBN 10 0954689402. [Google Scholar]
- Cobb, S.N.; O’Higgins, P. The ontogeny of sexual dimorphism in the facial skeleton of the African apes. J. Hum. Evol. 2007, 53, 176–190. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.M.; Machanda, Z.; Bernard, A.B.; Donovan, R.M.; Papakyrikos, A.M.; Muller, M.N.; Wrangham, R. First molar eruption, weaning, and life history in living wild chimpanzees. Proc. Natl. Acad. Sci. USA 2013, 110, 2787–2791. [Google Scholar] [CrossRef] [PubMed]
- Baxter, J.C. Interpersonal spacing in natural settings. Sociometry 1970, 33, 444–456. [Google Scholar] [CrossRef] [PubMed]
- Bindemann, M.; Scheepers, C.; Burton, A.M. Viewpoint and center of gravity affect eye movements to human faces. J. Vis. 2009, 9, 7. [Google Scholar] [CrossRef]
- Hsiao, J.H.W.; Cottrell, G. Two fixations suffice in face recognition. Psychol. Sci. 2008, 19, 998–1006. [Google Scholar] [CrossRef]
- Reichle, E.D.; Rayner, K.; Pollatsek, A. The E-Z Reader model of eye-movement control in reading: Comparisons to other models. Behav. Brain Sci. 2003, 26. [Google Scholar] [CrossRef]
- Brielmann, A.A.; Bülthoff, I.; Armann, R. Looking at faces from different angles: Europeans fixate different features in Asian and Caucasian faces. Vis. Res. 2014, 100, 105–112. [Google Scholar] [CrossRef]
- Lewis, M.B. The lady’s not for turning: Rotation of the Thatcher illusion. Perception 2001, 30, 769–774. [Google Scholar] [CrossRef]
- Hancock, K.J.; Rhodes, G. Contact, configural coding and the other-race effect in face recognition. Br. J. Psychol. 2008, 99, 45–56. [Google Scholar] [CrossRef]
- Tanaka, J.W.; Kiefer, M.; Bukach, C.M. A holistic account of the own-race effect in face recognition: Evidence from a cross-cultural study. Cognition 2004, 93, B1–B9. [Google Scholar] [CrossRef] [PubMed]
- Michel, C.; Rossion, B.; Han, J.; Chung, C.S.; Caldara, R. Holistic processing is finely tuned for faces of one’s own race. Psychol. Sci. 2006, 17, 608–615. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bulut, M.; Erdeniz, B. The Other-Race and Other-Species Effect during a Sex Categorization Task: An Eye Tracker Study. Behav. Sci. 2020, 10, 24. https://doi.org/10.3390/bs10010024
Bulut M, Erdeniz B. The Other-Race and Other-Species Effect during a Sex Categorization Task: An Eye Tracker Study. Behavioral Sciences. 2020; 10(1):24. https://doi.org/10.3390/bs10010024
Chicago/Turabian StyleBulut, Merve, and Burak Erdeniz. 2020. "The Other-Race and Other-Species Effect during a Sex Categorization Task: An Eye Tracker Study" Behavioral Sciences 10, no. 1: 24. https://doi.org/10.3390/bs10010024
APA StyleBulut, M., & Erdeniz, B. (2020). The Other-Race and Other-Species Effect during a Sex Categorization Task: An Eye Tracker Study. Behavioral Sciences, 10(1), 24. https://doi.org/10.3390/bs10010024