Antarctic Sea Ice Proxies from Marine and Ice Core Archives Suitable for Reconstructing Sea Ice over the Past 2000 Years

 , , , ,
, , , ,  ,
,
Abstract
1. Introduction
- In the first section, we collate the published sea ice reconstructions derived from (1) ice cores and (2) marine sediments spanning all, or part, of the past 2 ka. For each archive we review the different proxies and approaches used to reconstruct sea ice.
- We then discuss the potential for combining ice core and marine records and the obstacles which need to be overcome.
2. Results
2.1. Ice Core Based Reconstructions of Sea Ice Spanning the Past 2000 Years
2.2. Proxies for Sea Ice
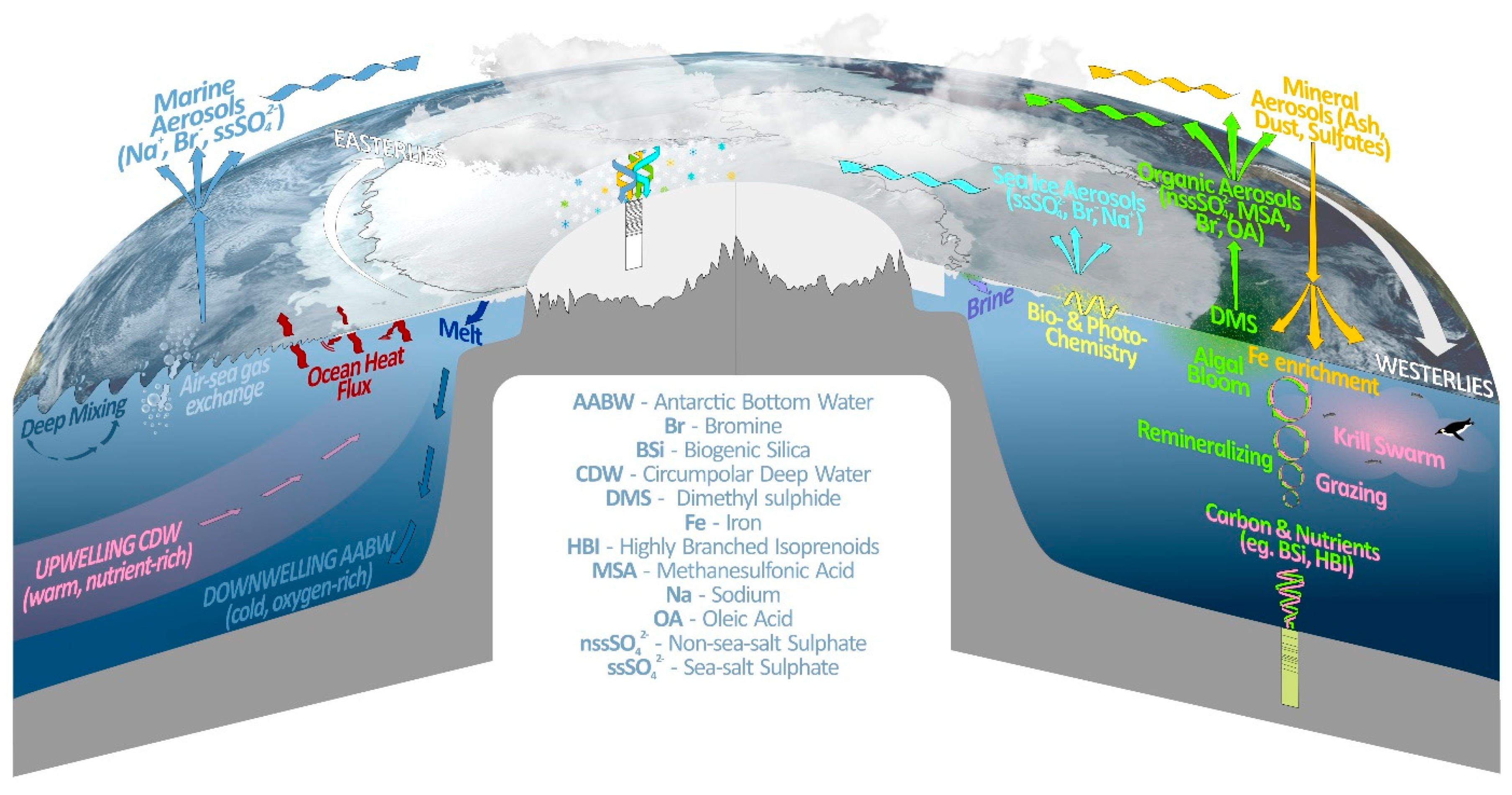
2.2.1. Sea Salts
2.2.2. MSA
2.2.3. Halogens
2.2.4. Novel Organic Compounds
2.2.5. Stable Water Isotopes
2.2.6. Snow Accumulation
2.3. Marine Based Reconstructions of Sea Ice Spanning the Past 2000 Years
2.4. Marine Proxies of Sea Ice
2.4.1. Diatoms
2.4.2. Biomarkers
2.4.3. Foraminifera
2.4.4. Geochemical
3. Advantages and Limitations
3.1. Ice Cores
3.2. Marine Records
4. Feasibility of Combining Marine and Ice Core Records over the Past 2000 Years
4.1. Spatial Coverage
4.2. Temporal Coverage
4.3. Comparable Proxies?
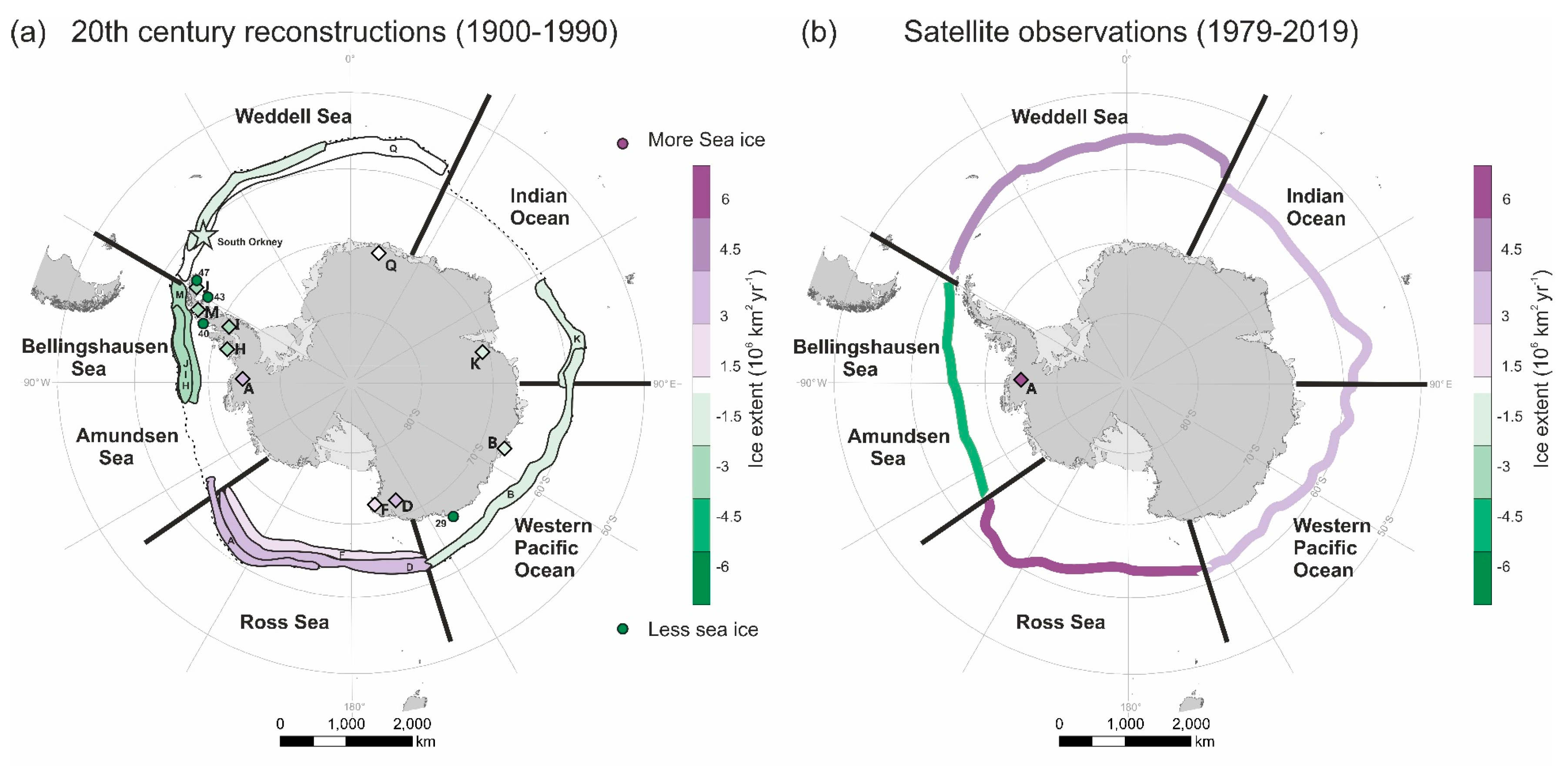
5. Synthesis of Sea Ice Trends during the 20th Century
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vancoppenolle, M.; Bopp, L.; Madec, G.; Dunne, J.; Ilyina, T.; Halloran, P.R.; Steiner, N. Future arctic ocean primary productivity from cmip5 simulations: Uncertain outcome, but consistent mechanisms. Glob. Biogeochem. Cycles 2013, 27, 605–619. [Google Scholar] [CrossRef]
- Turner, J.; Orr, A.; Gudmundsson, G.H.; Jenkins, A.; Bingham, R.G.; Hillenbrand, C.-D.; Bracegirdle, T.J. Atmosphere-ocean-ice interactions in the amundsen sea embayment, west antarctica. Rev. Geophys. 2017, 55, 235–276. [Google Scholar] [CrossRef]
- Trevena, A.J.; Jones, G.B. Dimethylsulphide and dimethylsulphoniopropionate in antarctic sea ice and their release during sea ice melting. Mar. Chem. 2006, 98, 210–222. [Google Scholar] [CrossRef]
- Nomura, D.; Granskog, M.A.; Assmy, P.; Simizu, D.; Hashida, G. Arctic and antarctic sea ice acts as a sink for atmospheric co2 during periods of snowmelt and surface flooding. J. Geophys. Res. Ocean. 2013, 118, 6511–6524. [Google Scholar] [CrossRef]
- Serreze, M.C.; Meier, W.N. The arctic’s sea ice cover: Trends, variability, predictability, and comparisons to the antarctic. Ann. N. Y. Acad. Sci. 2019, 1436, 36–53. [Google Scholar] [CrossRef]
- Stroeve, J.; Holland, M.M.; Meier, W.; Scambos, T.; Serreze, M. Arctic sea ice decline: Faster than forecast. Geophys. Res. Lett. 2007, 34, L09501. [Google Scholar] [CrossRef]
- Zwally, H.J.; Comiso, J.C.; Parkinson, C.L.; Cavalieri, D.J.; Gloersen, P. Variability of antarctic sea ice 1979–1998. J. Geophys. Res. Ocean. 2002, 107, 9–19. [Google Scholar] [CrossRef]
- Turner, J.; Comiso, J.C.; Marshall, G.J.; Lachlan-Cope, T.A.; Bracegirdle, T.; Maksym, T.; Meredith, M.P.; Wang, Z.; Orr, A. Non-annular atmospheric circulation change induced by stratospheric ozone depletion and its role in the recent increase of antarctic sea ice extent. Geophys. Res. Lett. 2009, 36, 37524. [Google Scholar] [CrossRef]
- Parkinson, C.L. A 40-y record reveals gradual antarctic sea ice increases followed by decreases at rates far exceeding the rates seen in the arctic. Proc. Natl. Acad. Sci. USA 2019, 116, 14414–14423. [Google Scholar] [CrossRef]
- Parkinson, C.L.; Cavalieri, D.J. Antarctic sea ice variability and trends, 1979–2010. Cryosphere 2012, 6, 871–880. [Google Scholar] [CrossRef]
- Hobbs, W.R.; Massom, R.; Stammerjohn, S.; Reid, P.; Williams, G.; Meier, W. A review of recent changes in southern ocean sea ice, their drivers and forcings. Glob. Planet. Chang. 2016, 143, 228–250. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to The Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013; p. 1535. [Google Scholar]
- Purich, A.; Cai, W.; England, M.H.; Cowan, T. Evidence for link between modelled trends in antarctic sea ice and underestimated westerly wind changes. Nat. Commun. 2016, 7, 10409. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.; Marshall, J.; Bitz, C.M.; Solomon, S.; Plumb, A. Antarctic ocean and sea ice response to ozone depletion: A two-time-scale problem. J. Clim. 2015, 28, 1206–1226. [Google Scholar] [CrossRef]
- Etourneau, J.; Sgubin, G.; Crosta, X.; Swingedouw, D.; Willmott, V.; Barbara, L.; Houssais, M.N.; Schouten, S.; Damste, J.S.S.; Goosse, H.; et al. Ocean temperature impact on ice shelf extent in the eastern antarctic peninsula. Nat. Commun. 2019, 10, 304. [Google Scholar] [CrossRef]
- Bracegirdle, T.J.; Stephenson, D.B.; Turner, J.; Phillips, T. The importance of sea ice area biases in 21st century multimodel projections of antarctic temperature and precipitation. Geophys. Res. Lett. 2015, 42, 10832–10839. [Google Scholar] [CrossRef]
- Stenni, B.; Curran, M.A.J.; Abram, N.J.; Orsi, A.; Goursaud, S.; Masson-Delmotte, V.; Neukom, R.; Goosse, H.; Divine, D.; van Ommen, T.; et al. Antarctic climate variability on regional and continental scales over the last 2000 years. Clim. Past 2017, 13, 1609–1634. [Google Scholar] [CrossRef]
- Thomas, E.R.; van Wessem, J.M.; Roberts, J.; Isaksson, E.; Schlosser, E.; Fudge, T.J.; Vallelonga, P.; Medley, B.; Lenaerts, J.; Bertler, N.; et al. Regional antarctic snow accumulation over the past 1000 years. Clim. Past 2017, 13, 1491–1513. [Google Scholar] [CrossRef]
- Lenaerts, J.T.M.; Fyke, J.; Medley, B. The signature of ozone depletion in recent antarctic precipitation change: A study with the community earth system model. Geophys. Res. Lett. 2018, 45, 12931–12939. [Google Scholar] [CrossRef]
- Wang, Y.; Thomas, E.R.; Hou, S.; Huai, B.; Wu, S.; Sun, W.; Qi, S.; Ding, M.; Zhang, Y. Snow accumulation variability over the west antarctic ice sheet since 1900: A comparison of ice core records with era-20c reanalysis. Geophys. Res. Lett. 2017, 44, 11482–11490. [Google Scholar] [CrossRef]
- Dalaiden, Q.; Goosse, H.; Klein, F.; Lenaerts, J.T.M.; Holloway, M.; Sime, L.; Thomas, E.R. Surface mass balance of the antarctic ice sheet and its link with surface temperature change in model simulations and reconstructions. Cryosphere Discuss. 2019, 2019, 1–29. [Google Scholar] [CrossRef]
- Medley, B.; Thomas, E.R. Increased snowfall over the antarctic ice sheet mitigated twentieth-century sea-level rise. Nat. Clim. Chang. 2019, 9, 34–39. [Google Scholar] [CrossRef]
- Thomas, E.R.; Abram, N.J. Ice core reconstruction of sea ice change in the amundsen-ross seas since 1702 A.D. Geophys. Res. Lett. 2016, 43, 5309–5317. [Google Scholar] [CrossRef]
- Curran, M.A.J.; van Ommen, T.D.; Morgan, V.I.; Phillips, K.L.; Palmer, A.S. Ice core evidence for antarctic sea ice decline since the 1950s. Science 2003, 302, 1203–1206. [Google Scholar] [CrossRef] [PubMed]
- Vallelonga, P.; Maffezzoli, N.; Moy, A.D.; Curran, M.A.J.; Vance, T.R.; Edwards, R.; Hughes, G.; Barker, E.; Spreen, G.; Saiz-Lopez, A.; et al. Sea-ice-related halogen enrichment at law dome, coastal east antarctica. Clim. Past 2017, 13, 171–184. [Google Scholar] [CrossRef]
- Foster, A.F.M.; Curran, M.A.J.; Smith, B.T.; Van Ommen, T.D.; Morgan, V.I. Covariation of sea ice and methanesulphonic acid in wilhelm ii land, east antarctica. Ann. Glaciol. 2006, 44, 429–432. [Google Scholar] [CrossRef]
- Becagli, S.; Castellano, E.; Cerri, O.; Curran, M.; Frezzotti, M.; Marino, F.; Morganti, A.; Proposito, M.; Severi, M.; Traversi, R.; et al. Methanesulphonic acid (msa) stratigraphy from a talos dome ice core as a tool in depicting sea ice changes and southern atmospheric circulation over the previous 140 years. Atmos. Environ. 2009, 43, 1051–1058. [Google Scholar] [CrossRef]
- Severi, M.; Becagli, S.; Caiazzo, L.; Ciardini, V.; Colizza, E.; Giardi, F.; Mezgec, K.; Scarchilli, C.; Stenni, B.; Thomas, E.R.; et al. Sea salt sodium record from talos dome (east antarctica) as a potential proxy of the antarctic past sea ice extent. Chemosphere 2017, 177, 266–274. [Google Scholar] [CrossRef]
- Welch, K.A.; Mayewski, P.A.; Whitlow, S.I. Methanesulfonic acid in coastal antarctic snow related to sea-ice extent. Geophys. Res. Lett. 1993, 20, 443–446. [Google Scholar] [CrossRef]
- Sinclair, K.E.; Bertler, N.A.N.; Bowen, M.M.; Arrigo, K.R. Twentieth century sea-ice trends in the ross sea from a high-resolution, coastal ice-core record. Geophys. Res. Lett. 2014, 41, 3510–3516. [Google Scholar] [CrossRef]
- Meyerson, E.A.; Mayewski, P.A.; Kreutz, K.J.; David Meeker, L.; Whitlow, S.I.; Twickler, M.S. The polar expression of enso and sea-ice variability as recorded in a south pole ice core. Ann. Glaciol. 2002, 35, 430–436. [Google Scholar] [CrossRef]
- Abram, N.J.; Thomas, E.R.; McConnell, J.R.; Mulvaney, R.; Bracegirdle, T.J.; Sime, L.C.; Aristarain, A.J. Ice core evidence for a 20th century decline in sea ice in the bellingshausen sea, antarctica. J. Geophys. Res. 2010, 115, D23101. [Google Scholar] [CrossRef]
- Xiao, C.; Dou, T.; Sneed, S.B.; Li, R.; Allison, I. An ice-core record of antarctic sea-ice extent in the southern indian ocean for the past 300 years. Ann. Glaciol. 2015, 56, 451–455. [Google Scholar] [CrossRef]
- King, A.C.F.; Thomas, E.R.; Pedro, J.B.; Markle, B.; Potocki, M.; Jackson, S.L.; Wolff, E.; Kalberer, M. Organic compounds in a sub-antarctic ice core: A potential suite of sea ice markers. Geophys. Res. Lett. 2019, 46, 9930–9939. [Google Scholar] [CrossRef] [PubMed]
- Porter, S.E.; Parkinson, C.L.; Mosley-Thompson, E. Bellingshausen sea ice extent recorded in an antarctic peninsula ice core. J. Geophys. Res. Atmos. 2016, 121, 13886–13990. [Google Scholar] [CrossRef]
- Criscitiello, A.S.; Das, S.B.; Evans, M.J.; Frey, K.E.; Conway, H.; Joughin, I.; Medley, B.; Steig, E.J. Ice sheet record of recent sea-ice behavior and polynya variability in the amundsen sea, west antarctica. J. Geophys. Res. Ocean. 2013, 118, 118–130. [Google Scholar] [CrossRef]
- Pasteris, D.R.; McConnell, J.R.; Das, S.B.; Criscitiello, A.S.; Evans, M.J.; Maselli, O.J.; Sigl, M.; Layman, L. Seasonally resolved ice core records from west antarctica indicate a sea ice source of sea-salt aerosol and a biomass burning source of ammonium. J. Geophys. Res. Atmos. 2014, 119, 9168–9182. [Google Scholar] [CrossRef]
- Rahaman, W.; Thamban, M.; Laluraj, C. Twentieth-century sea ice variability in the weddell sea and its effect on moisture transport: Evidence from a coastal east antarctic ice core record. Holocene 2016, 26, 338–349. [Google Scholar] [CrossRef]
- Abram, N.J.; Wolff, E.W.; Curran, M.A.J. A review of sea ice proxy information from polar ice cores. Quat. Sci. Rev. 2013, 79, 168–183. [Google Scholar] [CrossRef]
- Wagenbach, D.; Ducroz, F.; Mulvaney, R.; Keck, L.; Minikin, A.; Legrand, M.; Hall, J.S.; Wolff, E.W. Sea-salt aerosol in coastal antarctic regions. J. Geophys. Res. Atmos. 1998, 103, 10961–10974. [Google Scholar] [CrossRef]
- Wolff, E.W.; Rankin, A.M.; Röthlisberger, R. An ice core indicator of antarctic sea ice production? Geophys. Res. Lett. 2003, 30, 2158. [Google Scholar] [CrossRef]
- Rankin, A.M.; Wolff, E.W. A year-long record of size-segregated aerosol composition at halley, antarctica. J. Geophys. Res. Atmos. 2003, 108, 4775. [Google Scholar] [CrossRef]
- Rankin, A.M.; Wolff, E.W.; Martin, S. Frost flowers: Implications for tropospheric chemistry and ice core interpretation. J. Geophys. Res. Atmos. 2002, 107, 4683. [Google Scholar] [CrossRef]
- Roscoe, H.K.; Brooks, B.; Jackson, A.V.; Smith, M.H.; Walker, S.J.; Obbard, R.W.; Wolff, E.W. Frost flowers in the laboratory: Growth, characteristics, aerosol, and the underlying sea ice. J. Geophys. Res. 2011, 116, D12301. [Google Scholar] [CrossRef]
- Xin, Y.; John, A.P.; Richard, A.C. Sea salt aerosol production and bromine release: Role of snow on sea ice. Geophys. Res. Lett. 2008, 35. [Google Scholar] [CrossRef]
- Jiayue, H.; Lyatt, J. Wintertime enhancements of sea salt aerosol in polar regions consistent with a sea ice source from blowing snow. Atmos. Chem. Phys. 2017, 17, 3699–3712. [Google Scholar]
- Vega, C.P.; Isaksson, E.; Schlosser, E.; Divine, D.; Martma, T.; Mulvaney, R.; Eichler, A.; Schwikowski-Gigar, M. Variability of sea salts in ice and firn cores from fimbul ice shelf, dronning maud land, antarctica. Cryosphere 2018, 12, 1681–1697. [Google Scholar] [CrossRef]
- Sneed, S.B.; Mayewski, P.A.; Dixon, D.A. An emerging technique: Multi-ice-core multi-parameter correlations with antarctic sea-ice extent. Ann. Glaciol. 2011, 52, 347–354. [Google Scholar] [CrossRef][Green Version]
- Iizuka, Y.; Hondoh, T.; Fujii, Y. Antarctic sea ice extent during the holocene reconstructed from inland ice core evidence. J. Geophys. Res. Atmos. 2008, 113, D15114. [Google Scholar] [CrossRef]
- Mayewski, P.A.; Carleton, A.M.; Birkel, S.D.; Dixon, D.; Kurbatov, A.V.; Korotkikh, E.; McConnell, J.; Curran, M.; Cole-Dai, J.; Jiang, S.; et al. Ice core and climate reanalysis analogs to predict antarctic and southern hemisphere climate changes. Quat. Sci. Rev. 2017, 155, 50–66. [Google Scholar] [CrossRef]
- Huang, J.; Jaeglé, L.; Shah, V. Using caliop to constrain blowing snow emissions of sea salt aerosols over arctic and antarctic sea ice. Atmos. Chem. Phys. 2018, 18, 16253–16269. [Google Scholar] [CrossRef]
- Mezgec, K.; Stenni, B.; Crosta, X.; Masson-Delmotte, V.; Baroni, C.; Braida, M.; Ciardini, V.; Colizza, E.; Melis, R.; Salvatore, M.C.; et al. Holocene sea ice variability driven by wind and polynya efficiency in the ross sea. Nat. Commun. 2017, 8, 1334. [Google Scholar] [CrossRef] [PubMed]
- Curran, M.A.J.; Jones, G.B. Dimethyl sulfide in the southern ocean: Seasonality and flux. J. Geophys. Res. Atmos. 2000, 105, 20451–20459. [Google Scholar] [CrossRef]
- Dacey, J.W.H.; Wakeham, S.G. Oceanic dimethylsulfide: Production during zooplankton grazing on phytoplankton. Science 1986, 233, 1314–1316. [Google Scholar] [CrossRef] [PubMed]
- Legrand, M.; Ducroz, F.; Wagenbach, D.; Mulvaney, R.; Hall, J. Ammonium in coastal antarctic aerosol and snow: Role of polar ocean and penguin emissions. J. Geophys. Res. Atmos. 1998, 103, 11043–11056. [Google Scholar] [CrossRef]
- Rhodes, R.H.; Bertler, N.A.N.; Baker, J.A.; Sneed, S.B.; Oerter, H.; Arrigo, K.R. Sea ice variability and primary productivity in the ross sea, antarctica, from methylsulphonate snow record. Geophys. Res. Lett. 2009, 36, L10704. [Google Scholar] [CrossRef]
- Spolaor, A.; Vallelonga, P.; Plane, J.M.C.; Kehrwald, N.; Gabrieli, J.; Varin, C.; Turetta, C.; Cozzi, G.; Kumar, R.; Boutron, C.; et al. Halogen species record antarctic sea ice extent over glacial–interglacial periods. Atmos. Chem. Phys. 2013, 13, 6623–6635. [Google Scholar] [CrossRef]
- Impey, G.A.; Shepson, P.B.; Hastie, D.R.; Barrie, L.A.; Anlauf, K.G. Measurements of photolyzable chlorine and bromine during the polar sunrise experiment 1995. J. Geophys. Res. Atmos. 1997, 102, 16005–16010. [Google Scholar] [CrossRef]
- Simpson, W.R.; Carlson, D.; Hönninger, G.; Douglas, T.A.; Sturm, M.; Perovich, D.; Platt, U. First-year sea-ice contact predicts bromine monoxide (bro) levels at barrow, alaska better than potential frost flower contact. Atmos. Chem. Phys. 2007, 7, 621–627. [Google Scholar] [CrossRef]
- Crosta, X.; Sturm, A.; Armand, L.; Pichon, J.J. Late quaternary sea ice history in the indian sector of the southern ocean as recorded by diatom assemblages. Mar. Micropaleontol. 2004, 50, 209–223. [Google Scholar] [CrossRef]
- Spolaor, A.; Vallelonga, P.; Gabrieli, J.; Martma, T.; Björkman, M.P.; Isaksson, E.; Cozzi, G.; Turetta, C.; Kjær, H.A.; Curran, M.A.J.; et al. Seasonality of halogen deposition in polar snow and ice. Atmos. Chem. Phys. 2014, 14, 9613–9622. [Google Scholar] [CrossRef]
- Saiz-Lopez, A.; Baidar, S.; Cuevas, C.A.; Koenig, T.K.; Fernandez, R.P.; Dix, B.; Kinnison, D.E.; Lamarque, J.-F.; Rodriguez-Lloveras, X.; Campos, T.L.; et al. Injection of iodine to the stratosphere. Geophys. Res. Lett. 2015, 42, 6852–6859. [Google Scholar] [CrossRef]
- Atkinson, H.M.; Huang, R.-J.; Chance, R.; Roscoe, H.K.; Hughes, C.; Davison, B.; Schönhardt, A.; Mahajan, A.S.; Saiz-Lopez, A.; Hoffmann, T.; et al. Iodine emissions from the sea ice of the weddell sea. Atmos. Chem. Phys. 2012, 12, 11229–11244. [Google Scholar] [CrossRef]
- Granfors, A.; Ahnoff, M.; Mills, M.M.; Abrahamsson, K. Organic iodine in antarctic sea ice: A comparison between winter in the weddell sea and summer in the amundsen sea. J. Geophys. Res. Biogeosciences 2014, 119, 2276–2291. [Google Scholar] [CrossRef]
- Giorio, C.; Kehrwald, N.; Barbante, C.; Kalberer, M.; King, A.C.F.; Thomas, E.R.; Wolff, E.W.; Zennaro, P. Prospects for reconstructing paleoenvironmental conditions from organic compounds in polar snow and ice. Quat. Sci. Rev. 2018, 183, 1–22. [Google Scholar] [CrossRef]
- O’Dowd, C.D.; Facchini, M.C.; Cavalli, F.; Ceburnis, D.; Mircea, M.; Decesari, S.; Fuzzi, S.; Yoon, Y.J.; Putaud, J.-P. Biogenically driven organic contribution to marine aerosol. Nature 2004, 431, 676–680. [Google Scholar] [CrossRef] [PubMed]
- Pokhrel, A.; Kawamura, K.; Seki, O.; Matoba, S.; Shiraiwa, T. Ice core profiles of saturated fatty acids (c12:0–c30:0) and oleic acid (c18:1) from southern alaska since 1734 ad: A link to climate change in the northern hemisphere. Atmos. Environ. 2015, 100, 202–209. [Google Scholar] [CrossRef]
- Kawamura, K.; Suzuki, I.; Fujii, Y.; Watanabe, O. Ice core record of fatty acids over the past 450 years in greenland. Geophys. Res. Lett. 1996, 23, 2665–2668. [Google Scholar] [CrossRef]
- Baboukas, E.D.; Kanakidou, M.; Mihalopoulos, N. Carboxylic acids in gas and particulate phase above the atlantic ocean. J. Geophys. Res. Atmos. 2000, 105, 14459–14471. [Google Scholar] [CrossRef]
- Rinaldi, M.; Decesari, S.; Carbone, C.; Finessi, E.; Fuzzi, S.; Ceburnis, D.; O’Dowd, C.D.; Sciare, J.; Burrows, J.P.; Vrekoussis, M.; et al. Evidence of a natural marine source of oxalic acid and a possible link to glyoxal. J. Geophys. Res. Atmos. 2011, 116, D16204. [Google Scholar] [CrossRef]
- Dansgaard, W. Stable isotopes in precipitation. Tellus 1964, 16, 436–468. [Google Scholar] [CrossRef]
- Jouzel, J.; Lorius, C.; Petit, J.R.; Genthon, C.; Barkov, N.I.; Kotlyakov, V.M.; Petrov, V.M. Vostok ice core: A continuous isotope temperature record over the last climatic cycle (160,000 years). Nature 1987, 329, 403–408. [Google Scholar] [CrossRef]
- Holloway, M.D.; Sime, L.C.; Singarayer, J.S.; Tindall, J.C.; Bunch, P.; Valdes, P.J. Antarctic last interglacial isotope peak in response to sea ice retreat not ice-sheet collapse. Nat. Commun. 2016, 7, 12293. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.R.; Bracegirdle, T.J.; Turner, J.; Wolff, E.W. A 308 year record of climate variability in west antarctica. Geophys. Res. Lett. 2013, 40, 5492–5496. [Google Scholar] [CrossRef]
- Thomas, E.R.; Bracegirdle, T.J. Precipitation pathways for five new ice core sites in ellsworth land, west antarctica. Clim. Dyn. 2015, 44, 2067–2078. [Google Scholar] [CrossRef]
- Küttel, M.; Steig, E.J.; Ding, Q.; Monaghan, A.J.; Battisti, D.S. Seasonal climate information preserved in west antarctic ice core water isotopes: Relationships to temperature, large-scale circulation, and sea ice. Clim. Dyn. 2012, 39, 1841–1857. [Google Scholar] [CrossRef]
- Holloway, M.D.; Sime, L.C.; Allen, C.S.; Hillenbrand, C.-D.; Bunch, P.; Wolff, E.; Valdes, P.J. The spatial structure of the 128 ka antarctic sea ice minimum. Geophys. Res. Lett. 2017, 44, 11129–11139. [Google Scholar] [CrossRef]
- Tsukernik, M.; Lynch, A.H. Atmospheric meridional moisture flux over the southern ocean: A story of the amundsen sea. J. Clim. 2013, 26, 8055–8064. [Google Scholar] [CrossRef]
- Thomas, E.R.; Hosking, J.S.; Tuckwell, R.R.; Warren, R.A.; Ludlow, E.C. Twentieth century increase in snowfall in coastal west antarctica. Geophys. Res. Lett. 2015, 42, 9387–9393. [Google Scholar] [CrossRef]
- Turner, J.T. Zooplankton fecal pellets, marine snow, phytodetritus and the ocean’s biological pump. Prog. Oceanogr. 2015, 130, 205–248. [Google Scholar] [CrossRef]
- Hillaire–Marcel, C.; Vernal, A.D. Proxies in Late Cenozoic Paleoceanography, 1st, ed.; Elsevier: Amsterdam, The Netherlands, 2007; Volume 1, p. 843. [Google Scholar]
- Leventer, A.; Dunbar, R.B.; DeMaster, D.J. Diatom evidence for late holocene climatic events in granite harbor, antarctica. Paleoceanography 1993, 8, 373–386. [Google Scholar] [CrossRef]
- Leventer, A.; Dunbar, R.B. Recent diatom record of mcmurdo sound, antarctica: Implications for history of sea ice extent. Paleoceanography 1988, 3, 259–274. [Google Scholar] [CrossRef]
- Cunningham, W.L.; Leventer, A.; Andrews, J.T.; Jennings, A.E.; Licht, K. Late pleistocene-holocene marine condtions in the ross sea, antarctica: Evidence from the diatom record. Holocene 1999, 9, 129–139. [Google Scholar] [CrossRef]
- Hemer, M.A.; Harris, P.T. Sediment core from beneath the amery ice shelf, east antarctica, suggests mid-holocene ice-shelf retreat. Geology 2003, 31, 127–130. [Google Scholar] [CrossRef]
- Berg, S.; Wagner, B.; Cremer, H.; Leng, M.; Melles, M. Late quaternary environmental and climate history of rauer group, east antarctica. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 297, 201–213. [Google Scholar] [CrossRef]
- Crosta, X.; Crespin, J.; Swingedouw, D.; Marti, O.; Masson-Delmotte, V.; Etourneau, J.; Goosse, H.; Braconnot, P.; Yam, R.; Brailovski, I.; et al. Ocean as the main driver of antarctic ice sheet retreat during the holocene. Glob. Planet. Chang. 2018, 166, 62–74. [Google Scholar] [CrossRef]
- Denis, D.; Crosta, X.; Barbara, L.; Massé, G.; Renssen, H.; Ther, O.; Giraudeau, J. Sea ice and wind variability during the holocene in east antarctica: Insight on middle–high latitude coupling. Quat. Sci. Rev. 2010, 29, 3709–3719. [Google Scholar] [CrossRef]
- Pike, J.; Crosta, X.; Maddison, E.J.; Stickley, C.E.; Denis, D.; Barbara, L.; Renssen, H. Observations on the relationship between the antarctic coastal diatoms thalassiosira antarctica comber and porosira glacialis (grunow) jørgensen and sea ice concentrations during the late quaternary. Mar. Micropaleontol. 2009, 73, 14–25. [Google Scholar] [CrossRef]
- Taylor, F.; McMinn, A. Late quaternary diatom assemblages from prydz bay, eastern antarctica. Quat. Res. 2002, 57, 151–161. [Google Scholar] [CrossRef]
- McMinn, A. Late holocene increase in sea ice extent in fjords of the vestfold hills, eastern antarctica. Antarct. Sci. 2000, 12, 80–88. [Google Scholar] [CrossRef]
- McMinn, A.; Heijnis, H.; Harle, K.; McOrist, G. Late-holocene climatic change recorded in sediment cores from ellis fjord, eastern antarctica. Holocene 2001, 11, 291–300. [Google Scholar] [CrossRef]
- Allen, C.S.; Oakes-Fretwell, L.M.; Anderson, J.B.; Hodgson, D.A. A record of holocene glacial and oceanographic variability in neny fjord, antarctic peninsula. Holocene 2010, 20, 551–564. [Google Scholar] [CrossRef]
- Peck, V.L.; Allen, C.S.; Kender, S.; McClymont, E.L.; Hodgson, D.A. Oceanographic variability on the west antarctic peninsula during the holocene and the influence of upper circumpolar deep water. Quat. Sci. Rev. 2015, 119, 54–65. [Google Scholar] [CrossRef]
- Taylor, F.; McMinn, A. Evidence from diatoms for holocene climate fluctuation along the east antarctic margin. Holocene 2001, 11, 455–466. [Google Scholar] [CrossRef]
- Shevenell, A.E.; Domack, E.W.; Kernan, G.M. Record of holocene palaeoclimate change alond the antarctic peninsula: Evidence from glacial marine sediments, lallemand fjord. In Climatic Succession and Glacial History of the Southern Hemisphere over the Last Five Million Years; Banks, M.R., Brown, M.J., Eds.; Papers and Proceedings of the Royal Society of Tasmania; Department of Plant Science, University of Tasmania: Tasmania, Australian, 1996; Volume 130, pp. 55–64. [Google Scholar]
- Taylor, F.; Whitehead, J.; Domack, E. Holocene paleoclimate change in the antarctic peninsula: Evidence from the diatom, sedimentary and geochemical record. Mar. Micropaleontol. 2001, 41, 25–43. [Google Scholar] [CrossRef]
- Alley, K.; Patacca, K.; Pike, J.; Dunbar, R.; Leventer, A. Iceberg alley, east antarctic margin: Continuously laminated diatomaceous sediments from the late holocene. Mar. Micropaleontol. 2018, 140, 56–68. [Google Scholar] [CrossRef]
- Rathburn, A.E.; Pichon, J.J.; Ayress, M.A.; DeDeckker, P. Microfossil and stable-isotope evidence for changes in late holocene palaeoproductivity and palaeoceanographic conditions in the prydz bay region of antarctica. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1997, 131, 485–510. [Google Scholar] [CrossRef]
- Campagne, P.; Crosta, X.; Houssais, M.N.; Swingedouw, D.; Schmidt, S.; Martin, A.; Devred, E.; Capo, S.; Marieu, V.; Closset, I.; et al. Glacial ice and atmospheric forcing on the mertz glacier polynya over the past 250 years. Nat. Commun. 2015, 6, 6642. [Google Scholar] [CrossRef]
- Cremer, H.; Gore, D.; Melles, M.; Roberts, D. Palaeoclimatic significance of late quaternary diatom assemblages from southern windmill islands, east antarctica. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 195, 261–280. [Google Scholar] [CrossRef]
- Kirkup, H.; Melles, M.; Gore, D.B. Late quaternary environment of southern windmill islands, east antarctica. Antarct. Sci. 2002, 14, 385–394. [Google Scholar] [CrossRef]
- Maddison, E.J.; Pike, J.; Dunbar, R. Seasonally laminated diatom-rich sediments from dumont d’urville trough, east antarctic margin: Late-holocene neoglacial sea-ice conditions. Holocene 2012, 22, 857–875. [Google Scholar] [CrossRef]
- Campagne, P.; Crosta, X.; Schmidt, S.; Noëlle Houssais, M.; Ther, O.; Massé, G. Sedimentary response to sea ice and atmospheric variability over the instrumental period off adélie land, east antarctica. Biogeosciences 2016, 13, 4205–4218. [Google Scholar] [CrossRef]
- Kulbe, T.; Melles, M.; Verkulich, S.R.; Pushina, Z.V. East antarctic climate and environmental variability over the last 9400 years inferred from marine sediments of the bunger oasis. Arct. Antarct. Alp. Res. 2001, 33, 223–230. [Google Scholar] [CrossRef]
- Crosta, X.; Debret, M.; Denis, D.; Courty, M.A.; Ther, O. Holocene long- and short-term climate changes off adélie land, east antarctica. Geochem. Geophys. Geosystems 2007, 8, Q11009. [Google Scholar] [CrossRef]
- Crosta, X.; Denis, D.; Ther, O. Sea ice seasonality during the holocene, adelie land, east antarctica. Mar. Micropaleontol. 2008, 66, 222–232. [Google Scholar] [CrossRef]
- Crosta, X.; Romero, O.; Armand, L.K.; Pichon, J.-J. The biogeography of major diatom taxa in southern ocean sediments: 2. Open ocean related species. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 223, 66–92. [Google Scholar] [CrossRef]
- Denis, D.; Crosta, X.; Schmidt, S.; Carson, D.S.; Ganeshram, R.S.; Renssen, H.; Crespin, J.; Ther, O.; Billy, I.; Giraudeau, J. Holocene productivity changes off adelie land (east antarctica). Paleoceanography 2009, 24. [Google Scholar] [CrossRef]
- Kim, S.; Yoo, K.-C.; Lee, J.I.; Khim, B.-K.; Bak, Y.-S.; Lee, M.K.; Lee, J.; Domack, E.W.; Christ, A.J.; Yoon, H.I. Holocene paleoceanography of bigo bay, west antarctic peninsula: Connections between surface water productivity and nutrient utilization and its implication for surface-deep water mass exchange. Quat. Sci. Rev. 2018, 192, 59–70. [Google Scholar] [CrossRef]
- Etourneau, J.; Collins, L.G.; Willmott, V.; Kim, J.H.; Barbara, L.; Leventer, A.; Schouten, S.; Sinninghe Damsté, J.S.; Bianchini, A.; Klein, V.; et al. Holocene climate variations in the western antarctic peninsula: Evidence for sea ice extent predominantly controlled by changes in insolation and enso variability. Clim. Past 2013, 9, 1431–1446. [Google Scholar] [CrossRef]
- Leventer, A. The fate of antarctic “sea-ice diatoms“ and their use as palaeoenvironmental indicators. In Antarctic Sea Ice Biological, Processes, Interactions and Variability; Union, A.G., Ed.; Volume Antarctic Research Series; American Geophysical Union: Washington, DC, USA, 1998; pp. 121–137. [Google Scholar]
- Leventer, A.; Domack, E.W.; Ishman, S.E.; Brachfeld, S.; McClennen, C.E.; Manley, P. Productivity cycles of 200–300 years in the antarctic peninsula region: Understanding linkages among the sun, atmosphere, oceans, sea ice, and biota. Geol. Soc. Am. Bull. 1996, 108, 1626–1644. [Google Scholar] [CrossRef]
- Ishman, S.E.; Sperling, M.R. Benthic foraminiferal record of holocene deep-water evolution in the palmer deep, western antarctic peninsula. Geology 2002, 30, 435–438. [Google Scholar] [CrossRef]
- Shevenell, A.E.; Kennett, J.P. Antarctic holocene climate change: A benthic foraminiferal stable isotope record from palmer deep. Paleoceanography 2002, 17. [Google Scholar] [CrossRef]
- Sjunneskog, C.; Taylor, F. Postglacial marine diatom record of the palmer deep, antarctic peninsula (odp leg 178, site 1098) 1. Total diatom abundance. Paleoceanography 2002, 17. [Google Scholar] [CrossRef]
- Taylor, F.; Sjunneskog, C. Postglacial marine diatom record of the palmer deep, antarctic peninsula (odp leg 178, site 1098) 2. Diatom assemblages. Paleoceanography 2002, 17, PA8001. [Google Scholar] [CrossRef]
- Barbara, L.; Crosta, X.; Schmidt, S.; Masse, G. Diatoms and biomarkers evidence for major changes in sea ice conditions prior the instrumental period in antarctic peninsula. Quat. Sci. Rev. 2013, 79, 99–110. [Google Scholar] [CrossRef]
- Yoon, H.I.; Park, B.-K.; Kim, Y.; Kang, C.Y. Glaciomarine sedimentation and its paleoclimatic implications on the antarctic peninsula shelf over the last 15 000 years. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 185, 235–254. [Google Scholar] [CrossRef]
- Minzoni, R.T.; Anderson, J.B.; Fernandez, R.; Wellner, J.S. Marine record of holocene climate, ocean, and cryosphere interactions: Herbert sound, james ross island, antarctica. Quat. Sci. Rev. 2015, 129, 239–259. [Google Scholar] [CrossRef]
- Barbara, L.; Crosta, X.; Leventer, A.; Schmidt, S.; Etourneau, J.; Domack, E.; Masse, G. Environmental responses of the northeast antarctic peninsula to the holocene climate variability. Paleoceanography 2016, 31, 131–147. [Google Scholar] [CrossRef]
- Heroy, D.C.; Sjunneskog, C.; Anderson, J.B. Holocene climate change in the bransfield basin, antarctic peninsula: Evidence from sediment and diatom analysis. Antarct. Sci. 2008, 20, 69–87. [Google Scholar] [CrossRef]
- Majewski, W.; Anderson, J.B. Holocene foraminiferal assemblages from firth of tay, antarctic peninsula: Paleoclimate implications. Mar. Micropaleontol. 2009, 73, 135–147. [Google Scholar] [CrossRef]
- Michalchuk, B.R.; Anderson, J.B.; Wellner, J.S.; Manley, P.L.; Majewski, W.; Bohaty, S. Holocene climate and glacial history of the northeastern antarctic peninsula: The marine sedimentary record from a long shaldril core. Quat. Sci. Rev. 2009. In Press, Corrected Proof. [Google Scholar] [CrossRef]
- Barcena, M.A.; Fabres, B.; Isla, E.; Flores, J.A.; Sierro, F.J.; Canals, M.; Palanques, A. Holocene neoglacial events in the bransfield strait (antarctica). Palaeocenographic and palaeoclimatic significance. Sci. Mar. 2006, 70, 607–619. [Google Scholar] [CrossRef]
- Barcena, M.A.; Isla, E.; Plaza, A.; Flores, J.A.; Sierro, F.J.; Masque, P.; Sanchez-Cabeza, J.A.; Palanques, A. Bioaccumulation record and paleoclimatic significance in the western bransfield strait. The last 2000 years. Deep-Sea Res. Part. Ii-Top. Stud. Oceanogr. 2002, 49, 935–950. [Google Scholar] [CrossRef]
- Kyrmanidou, A.; Vadman, K.J.; Ishman, S.E.; Leventer, A.; Brachfeld, S.; Domack, E.W.; Wellner, J.S. Late holocene oceanographic and climatic variability recorded by the perseverance drift, northwestern weddell sea, based on benthic foraminifera and diatoms. Mar. Micropaleontol. 2018, 141, 10–22. [Google Scholar] [CrossRef]
- Milliken, K.T.; Anderson, J.B.; Wellner, J.S.; Bohaty, S.M.; Manley, P.L. High-resolution holocene climate record from maxwell bay, south shetland islands, antarctica. Geol. Soc. Am. Bull. 2009, 121, 1711–1725. [Google Scholar] [CrossRef]
- Yoon, H.I.; Yoo, K.C.; Park, B.K.; Kim, Y.; Khim, B.K.; Kang, C.Y. The origin of massive diamicton in marian and potter coves, king george island, west antarctica. Geosci. J. 2004, 8, 1–10. [Google Scholar] [CrossRef]
- Yoo, K.-C.; Yoon, H.I.; Kim, J.-K.; Khim, B.-K. Sedimentological, geochemical and palaeontological evidence for a neoglacial cold event during the late holocene in the continental shelf of the northern south shetland islands, west antarctica. Polar Res. 2009, 28, 177–192. [Google Scholar] [CrossRef]
- Yoon, H.I.; Yoo, K.C.; Bak, Y.S.; Lim, H.S.; Kim, Y.; Lee, J.I. Late holocene cyclic glaciomarine sedimentation in a subpolar fjord of the south shetland islands, antarctica, and its paleoceanographic significance: Sedimentological, geochemical, and paleontological evidence. Geol. Soc. Am. Bull. 2010, 122, 1298–1307. [Google Scholar] [CrossRef]
- Barcena, M.A.; Gersonde, R.; Ledesma, S.; Fabres, J.; Calafat, A.M.; Canals, M.; Sierro, F.J.; Flores, J.A. Record of holocene glacial oscillations in bransfield basin as revealed by siliceous microfossil assemblages. Antarct. Sci. 1998, 10, 269–285. [Google Scholar] [CrossRef]
- Divine, D.V.; Koç, N.; Isaksson, E.; Nielsen, S.; Crosta, X.; Godtliebsen, F. Holocene antarctic climate variability from ice and marine sediment cores: Insights on ocean-atmosphere interaction. Quat. Sci. Rev. 2010, 29, 303–312. [Google Scholar] [CrossRef]
- Ferry, A.J.; Crosta, X.; Quilty, P.G.; Fink, D.; Howard, W.; Armand, L.K. First records of winter sea ice concentration in the southwest pacific sector of the southern ocean. Paleoceanography 2015, 30, 1525–1539. [Google Scholar] [CrossRef]
- Xiao, W.S.; Esper, O.; Gersonde, R. Last glacial—holocene climate variability in the atlantic sector of the southern ocean. Quat. Sci. Rev. 2016, 135, 115–137. [Google Scholar] [CrossRef]
- Bianchi, C.; Gersonde, R. Climate evolution at the last deglaciation: The role of the southern ocean. Earth Planet. Sci. Lett. 2004, 228, 407–424. [Google Scholar] [CrossRef]
- Presti, M.; De Santis, L.; Busetti, M.; Harris, P.T. Late pleistocene and holocene sedimentation on the george v continental shelf, east antarctica. Deep-Sea Res. Part. Ii-Top. Stud. Oceanogr. 2003, 50, 1441–1461. [Google Scholar] [CrossRef]
- Khim, B.K.; Yoon, H.I.; Kang, C.Y.; Bahk, J.J. Unstable climate oscillations during the late holocene in the eastern bransfield basin, antarctic peninsula. Quat. Res. 2002, 58, 234–245. [Google Scholar] [CrossRef]
- Borchers, A.; Dietze, E.; Kuhn, G.; Esper, O.; Voigt, I.; Hartmann, K.; Diekmann, B. Holocene ice dynamics and bottom-water formation associated with cape darnley polynya activity recorded in burton basin, east antarctica. Mar. Geophys. Res. 2016, 37, 49–70. [Google Scholar] [CrossRef]
- Sancetta, C.; Villareal, T.; Falkowski, P. Massive fluxes of rhizosolenid diatoms - a common occurrence. Limnol. Oceanogr. 1991, 36, 1452–1457. [Google Scholar] [CrossRef]
- Villareal, T.A.; Fryxell, G.A. Temperature effects on the valve structure of the bipolar diatoms thalassiosira antarctica and porosira glacialis. Polar Biol. 1983, 2, 163–169. [Google Scholar] [CrossRef]
- Armand, L.K.; Crosta, X.; Romero, O.; Pichon, J.-J. The biogeography of major diatom taxa in southern ocean sediments: 1. Sea ice related species. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 223, 93–126. [Google Scholar] [CrossRef]
- Clarke, D.B.; Ackley, S.F. Sea ice structure and biological activity in the antarctic marginal ice zone. J. Geophys. Res. 1984. [Google Scholar] [CrossRef]
- Zielinski, U.; Gersonde, R.; Sieger, R.; Futterer, D. Quaternary surface water temperature estimations: Calibration of a diatom transfer function for the southern ocean. Paleoceanography 1998, 13, 365–383. [Google Scholar] [CrossRef]
- Gersonde, R.; Zielinski, U. The reconstruction of late quaternary antarctic sea-ice distribution—the use of diatoms as a proxy for sea-ice. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 162, 263–286. [Google Scholar] [CrossRef]
- Schneider-Mor, A.; Yam, R.; Bianchi, C.; Kunz-Pirrung, M.; Gersonde, R.; Shemesh, A. Nutrient regime at the siliceous belt of the atlantic sector of the southern ocean during the past 660 ka. Paleoceanography 2008, 23, PA3217. [Google Scholar] [CrossRef]
- Esper, O.; Gersonde, R. Quaternary surface water temperature estimations: New diatom transfer functions for the southern ocean. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 414, 1–19. [Google Scholar] [CrossRef]
- Denis, D.; Crosta, X.; Zaragosi, S.; Romero, O.; Martin, B.; Mas, V. Seasonal and subseasonal climate changes recorded in laminated diatom ooze sediments, adélie land, east antarctica. Holocene 2006, 16, 1137–1147. [Google Scholar] [CrossRef]
- Minzoni, R.T.; Majewski, W.; Anderson, J.B.; Yokoyama, Y.; Fernandez, R.; Jakobsson, M. Oceanographic influences on the stability of the cosgrove ice shelf, antarctica. Holocene 2017, 27, 1645–1658. [Google Scholar] [CrossRef]
- Esper, O.; Gersonde, R. New tools for the reconstruction of pleistocene antarctic sea ice. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 399, 260–283. [Google Scholar] [CrossRef]
- Maddison, E.J.; Pike, J.; Leventer, A.; Dunbar, R.; Brachfeld, S.; Domack, E.W.; Manley, P.; McClennen, C. Post-glacial seasonal diatom record of the mertz glacier polynya, east antarctica. Mar. Micropaleontol. 2006, 60, 66–88. [Google Scholar] [CrossRef]
- Fryxell, G.A.; Prasad, A.K.S.K. Eucampia antarctica var. Recta (mangin) stat. Nov. (biddulphiaceae, bacillariophyceae): Life stages at the weddell sea ice edge. Phycologia 1990, 29, 27–38. [Google Scholar] [CrossRef]
- Kaczmarska, I.; Barbrick, N.E.; Ehrman, J.M.; Cant, G.P. Eucampia index as an indicator of the late pleistocene oscillations of the winter sea-ice extent at the odp leg 119 site 745b at the kerguelen plateau. Hydrobiologia 1993, 269–270, 103–112. [Google Scholar] [CrossRef]
- Leventer, A.; Domack, E.; Barkoukis, A.; McAndrews, B.; Murray, J. Laminations from the palmer deep: A diatom-based interpretation. Paleoceanography 2002, 17, 1–15. [Google Scholar] [CrossRef]
- Crosta, X.; Pichon, J.J.; Burckle, L.H. Application of modern analog technique to marine antarctic diatoms: Reconstruction of maximum sea-ice extent at the last glacial maximum. Paleoceanography 1998, 13, 284–297. [Google Scholar] [CrossRef]
- Crosta, X.; Pichon, J.J.; Burckle, L.H. Reappraisal of antarctic seasonal sea-ice at the last glacial maximum. Geophys. Res. Lett. 1998, 25, 2703–2706. [Google Scholar] [CrossRef]
- Ferry, A.J.; Prvan, T.; Jersky, B.; Crosta, X.; Armand, L.K. Statistical modeling of southern ocean marine diatom proxy and winter sea ice data: Model comparison and developments. Prog. Oceanogr. 2015, 100–112. [Google Scholar] [CrossRef]
- Xu, L.; Russell, L.M.; Burrows, S.M. Potential sea salt aerosol sources from frost flowers in the pan-arctic region. J. Geophys. Res. Atmos. 2016, 121, 10840–10856. [Google Scholar] [CrossRef]
- Massé, G.; Belt, S.T.; Crosta, X.; Schmidt, S.; Snape, I.; Thomas, D.N.; Rowland, S.J. Highly branched isoprenoids as proxies for variable sea ice conditions in the southern ocean. Antarct. Sci. 2011, 23, 487–498. [Google Scholar] [CrossRef]
- Smik, L.; Belt, S.T.; Lieser, J.L.; Armand, L.K.; Leventer, A. Distributions of highly branched isoprenoid alkenes and other algal lipids in surface waters from east antarctica: Further insights for biomarker-based paleo sea-ice reconstruction. Org. Geochem. 2016, 95, 71–80. [Google Scholar] [CrossRef]
- Smik, L.; Cabedo-Sanz, P.; Belt, S.T. Semi-quantitative estimates of paleo arctic sea ice concentration based on source-specific highly branched isoprenoid alkenes: A further development of the pip25 index. Org. Geochem. 2016, 92, 63–69. [Google Scholar] [CrossRef]
- Belt, S.T. Source-specific biomarkers as proxies for arctic and antarctic sea ice. Org. Geochem. 2018, 125, 277–298. [Google Scholar] [CrossRef]
- Belt, S.T.; Brown, T.A.; Smik, L.; Tatarek, A.; Wiktor, J.; Stowasser, G.; Assmy, P.; Allen, C.S.; Husum, K. Identification of c25 highly branched isoprenoid (hbi) alkenes in diatoms of the genus rhizosolenia in polar and sub-polar marine phytoplankton. Org. Geochem. 2017, 110, 65–72. [Google Scholar] [CrossRef]
- Belt, S.T.; Smik, L.; Brown, T.A.; Kim, J.H.; Rowland, S.J.; Allen, C.S.; Gal, J.K.; Shin, K.H.; Lee, J.I.; Taylor, K.W.R. Source identification and distribution reveals the potential of the geochemical antarctic sea ice proxy ipso25. Nat. Commun. 2016, 7, 12655. [Google Scholar] [CrossRef]
- Belt, S.T.; Masse, G.; Rowland, S.J.; Poulin, M.; Michel, C.; LeBlanc, B. A novel chemical fossil of palaeo sea ice: Ip25. Org. Geochem. 2007, 38, 16–27. [Google Scholar] [CrossRef]
- Riaux-Gobin, C.; Dieckmann, G.S.; Poulin, M.; Neveux, J.; Labrune, C.; Vetion, G. Environmental conditions, particle flux and sympagic microalgal succession in spring before the sea-ice break-up in adelie land, east antarctica. Polar Res. 2013, 32, 19675. [Google Scholar] [CrossRef]
- Schmidt, K.; Brown, T.A.; Belt, S.T.; Ireland, L.C.; Taylor, K.W.R.; Thorpe, S.E.; Ward, P.; Atkinson, A. Do pelagic grazers benefit from sea ice? Insights from the antarctic sea ice proxy ipso25. Biogeosciences 2018, 15, 1987–2006. [Google Scholar] [CrossRef]
- Belt, S.T.; Müller, J. The arctic sea ice biomarker ip25: A review of current understanding, recommendations for future research and applications in palaeo sea ice reconstructions. Quat. Sci. Rev. 2013, 79, 9–25. [Google Scholar] [CrossRef]
- Brown, T.A.; Belt, S.T. Biomarker-based h-print quantifies the composition of mixed sympagic and pelagic algae consumed by artemia sp. J. Exp. Mar. Biol. Ecol. 2017, 488, 32–37. [Google Scholar] [CrossRef]
- Rontani, J.-F.; Belt, S.T.; Vaultier, F.; Brown, T.A. Visible light induced photo-oxidation of highly branched isoprenoid (hbi) alkenes: Significant dependence on the number and nature of double bonds. Org. Geochem. 2011, 42, 812–822. [Google Scholar] [CrossRef]
- Collins, L.G.; Allen, C.S.; Pike, J.; Hodgson, D.A.; Weckström, K.; Massé, G. Evaluating highly branched isoprenoid (hbi) biomarkers as a novel antarctic sea-ice proxy in deep ocean glacial age sediments. Quat. Sci. Rev. 2013, 79, 87–98. [Google Scholar] [CrossRef]
- Sinninghe Damsté, J.S.; Rijpstra, W.I.C.; Coolen, M.J.L.; Schouten, S.; Volkman, J.K. Rapid sulfurisation of highly branched isoprenoid (hbi) alkenes in sulfidic holocene sediments from ellis fjord, antarctica. Org. Geochem. 2007, 38, 128–139. [Google Scholar] [CrossRef]
- Cabedo Sanz, P.; Smik, L.; Belt, S.T. On the stability of various highly branched isoprenoid (hbi) lipids in stored sediments and sediment extracts. Org. Geochem. 2016, 97, 74–77. [Google Scholar] [CrossRef]
- Vorrath, M.E.; Muller, J.; Esper, O.; Mollenhauer, G.; Haas, C.; Schefuss, E.; Fahl, K. Highly branched isoprenoids for southern ocean sea ice reconstructions: A pilot study from the western antarctic peninsula. Biogeosciences 2019, 16, 2961–2981. [Google Scholar] [CrossRef]
- Müller, J.; Massé, G.; Stein, R.; Belt, S.T. Variability of sea-ice conditions in the fram strait over the past 30,000 years. Nat. Geosci. 2009, 2, 772. [Google Scholar] [CrossRef]
- Müller, J.; Wagner, A.; Fahl, K.; Stein, R.; Prange, M.; Lohmann, G. Towards quantitative sea ice reconstructions in the northern north atlantic: A combined biomarker and numerical modelling approach. Earth Planet. Sci. Lett. 2011, 306, 137–148. [Google Scholar] [CrossRef]
- Mortyn, P.G.; Charles, C.D. Planktonic foraminiferal depth habitat and δ18o calibrations: Plankton tow results from the atlantic sector of the southern ocean. Paleoceanography 2003, 18, 1037. [Google Scholar] [CrossRef]
- Dieckmann, G.S.; Spindler, M.; Lange, M.A.; Ackley, S.F.; Eicken, H. Antarctic sea ice - a habitat for the foraminifer neogloboquadrina-pachyderma. J. Foraminifer. Res. 1991, 21, 182–189. [Google Scholar] [CrossRef]
- Kramer, M.; Swadling, K.M.; Meiners, K.M.; Kiko, R.; Scheltz, A.; Nicolaus, M.; Werner, I. Antarctic sympagic meiofauna in winter: Comparing diversity, abundance and biomass between perennially and seasonally ice-covered regions. Deep-Sea Res. Ii 2011, 9–10, 62–89. [Google Scholar] [CrossRef]
- Lipps, J.; Krebs, W. Planktonic foraminifera associated with antarctic sea ice. J. Foraminifer. Res. 1974, 4, 80–85. [Google Scholar] [CrossRef]
- Spindler, M.; Dieckmann, G.S. Distribution and abundance of the planktic foraminifer neogloboquadrina pachyderma in sea ice of the weddell sea (antarctica). Polar Biol. 1986, 5, 185–191. [Google Scholar] [CrossRef]
- Vautravers, M.J.; Hodell, D.A.; Channell, J.E.T.; Hillenbrand, C.-D.; Hall, M.; Smith, J.; Larter, R.D. Palaeoenvironmental records from the west antarctic peninsula drift sediments over the last 75 ka. Geol. Soci. Lond. Spec. Publ. 2013, 381, 263–276. [Google Scholar] [CrossRef]
- Mikis, A.; Hendry, K.; Pike, J.; Schmidt, D.N.; Edgar, K.M.; Peck, V.; Peeters, F.J.C.; Leng, M.; Meredith, M.J.; Todd, C.L.; et al. Temporal variability in foraminiferal morphology and geochemistry at the west antarctic peninsula: A sediment trap study. Biogeosciences Discuss. 2019, 16, 3267–3282. [Google Scholar] [CrossRef]
- Anderson, J.B. Ecology and distribution of foraminifera in the weddell sea of antarctica. Micropalaeontology 1975, 21, 69–96. [Google Scholar] [CrossRef]
- Ishman, S.E.; Domack, E.W. Oceanographic controls on benthic foraminifera from 12 the bellingshausen margin of the antarctic peninsula. Mar. Micropalaeontol. 1994, 24, 119–155. [Google Scholar] [CrossRef]
- Milam, R.W.; Anderson, J.B. Distribution and ecology of recent benthonic foraminifera of the adelie-george v continental shelf and slope, antarctica. Mar. Micropaleontol. 1981, 6, 297–325. [Google Scholar]
- Schmidt, M.; Botz, R.; Stoffers, P.; Anders, T.; Bohrmann, G. Oxygen isotopes in marine diatoms: A comparative study of analytical techniques and new results on the isotope composition of recent marine diatoms. Geochim. Et Cosmochim. Acta 1997, 61, 2275–2280. [Google Scholar] [CrossRef]
- Shemesh, A.; Charles, C.D.; Fairbanks, R.G. Oxygen isotopes in biogenic silica: Global changes in ocean temperature and isotopic composition. Science 1992, 256, 1434–1436. [Google Scholar] [CrossRef] [PubMed]
- Swann, G.E.A.; Leng, M.J. A review of diatom δ18o in palaeoceanography. Quat. Sci. Rev. 2009, 28, 384–398. [Google Scholar] [CrossRef]
- Meredith, M.P.; Venables, H.J.; Clarke, A.; Ducklow, H.W.; Erickson, M.; Leng, M.J.; Lenaerts, J.T.M.; Broeke, M.R.V.D. The freshwater system west of the antarctic peninsula: Spatial and temporal changes. J. Clim. 2013, 26, 1669–1684. [Google Scholar] [CrossRef]
- Meredith, M.P.; Wallace, M.I.; Stammerjohn, S.E.; Renfrew, I.A.; Clarke, A.; Venables, H.J.; Shoosmith, D.R.; Souster, T.; Leng, M.J. Changes in the freshwater composition of the upper ocean west of the antarctic peninsula during the first decade of the 21st century. Prog. Oceanogr. 2010, 87, 127–143. [Google Scholar] [CrossRef]
- Crespin, J.; Yam, R.; Crosta, X.; Masse, G.; Schmidt, S.; Campagne, P.; Shemesh, A. Holocene glacial discharge fluctuations and recent instability in east antarctica. Earth Planet. Sci. Lett. 2014, 394, 38–47. [Google Scholar] [CrossRef]
- Dickens, W.A.; Kuhn, G.; Leng, M.J.; Graham, A.G.C.; Dowdeswell, J.A.; Meredith, M.P.; Hillenbrand, C.D.; Hodgson, D.A.; Roberts, S.J.; Sloane, H.; et al. Enhanced glacial discharge from the eastern antarctic peninsula since the 1700s associated with a positive southern annular mode. Nature Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Pike, J.; Swann, G.E.A.; Leng, M.J.; Snelling, A.M. Glacial discharge along the west antarctic peninsula during the holocene. Nat. Geosci. 2013, 6, 199–202. [Google Scholar] [CrossRef]
- Swann, G.E.A.; Pike, J.; Snelling, A.M.; Leng, M.J.; Williams, M.C. Seasonally resolved diatom delta o-18 records from the west antarctic peninsula over the last deglaciation. Earth Planet. Sci. Lett. 2013, 364, 12–23. [Google Scholar] [CrossRef]
- Gibson, J.A.E.; Trull, T.; Nichols, P.D.; Summons, R.E.; McMinn, A. Sedimentation of c-13-rich organic matter from antarctic sea-ice algae: A potential indicator of past sea-ice extent. Geology 1999, 27, 331–334. [Google Scholar] [CrossRef]
- Tortell, P.D.; Mills, M.M.; Payne, C.D.; Maldonado, M.T.; Chierici, M.; Fransson, A.; Alderkamp, A.C.; Arrigo, K.R. Inorganic c utilization and c isotope fractionation by pelagic and sea ice algal assemblages along the antarctic continental shelf. Mar. Ecol. Prog. Ser. 2013, 483, 47–66. [Google Scholar] [CrossRef]
- Schoepfer, S.; Shen, J.; Wei, H.; Tyson, R.; Ingall, E. Toc, organic p, and biogenic ba accumulation rates as proxies for marine primary productivity and export flux. Earth Sci. Rev. 2014, 149, 23–52. [Google Scholar] [CrossRef]
- Paytan, A. Ocean paleoproductivity. In Encyclopedia of Paleoclimatology and Ancient Environments; Gornitz, V., Ed.; Springer: Dordrecht, The Netherlands, 2009; pp. 644–651. [Google Scholar]
- Ragueneau, O.; Tréguer, P.; Leynaert, A.; Anderson, R.F.; Brzezinski, M.A.; DeMaster, D.J.; Dugdale, R.C.; Dymond, J.; Fischer, G.; François, R.; et al. A review of the si cycle in the modern ocean: Recent progress and missing gaps in the application of biogenic opal as a paleoproductivity proxy. Glob. Planet. Chang. 2000, 26, 317–365. [Google Scholar] [CrossRef]
- Chase, Z.; Kohfeld, K.E.; Matsumoto, K. Controls on biogenic silica burial in the southern ocean. Glob. Biogeochem. Cycles 2015, 29, 1599–1616. [Google Scholar] [CrossRef]
- Chase, Z.; Anderson, R.F.; Fleisher, M.Q.; Kubik, P.W. Accumulation of biogenic and lithogenic material in the pacific sector of the southern ocean during the past 40,000 years. Deep Sea Res. Part. Ii Top. Stud. Oceanogr. 2003, 50, 799–832. [Google Scholar] [CrossRef]
- Bradtmiller, L.I.; Anderson, R.F.; Fleisher, M.Q.; Burckle, L.H. Opal burial in the equatorial atlantic ocean over the last 30 ka: Implications for glacial0interglacial changes in the ocean silicon cycle. Paleoceanography 2007, 22, 1–15. [Google Scholar] [CrossRef]
- Hodell, D.A.; Kanfoush, S.L.; Shemesh, A.; Crosta, X.; Charles, C.D.; Guilderson, T.P. Abrupt cooling of antarctic surface waters and sea ice expansion in the south atlantic sector of the southern ocean at 5000 cal yr b.P. Quat. Res. 2001, 56, 191–198. [Google Scholar] [CrossRef]
- Crosta, X.; Shemesh, A. Reconciling down core anticorrelation of diatom carbon and nitrogen isotopic ratios from the southern ocean. Paleoceanography 2002, 17. [Google Scholar] [CrossRef]
- Thomas, E.R.; Bracegirdle, T.J. Improving ice core interpretation using in situ and reanalysis data. J. Geophys. Res. Atmos. 2009, 114, D20116. [Google Scholar] [CrossRef]
- Andrews, J.T.; Domack, E.W.; Cunningham, W.L.; Leventer, A.; Licht, K.J.; Jull, A.J.T.; DeMaster, D.J.; Jennings, A.E. Problems and possible solutions concerning radiocarbon dating of surface marine sediments, ross sea, antarctica. Quat. Res. 1999, 52, 206–216. [Google Scholar] [CrossRef]
- Hall, B.L.; Koffman, T.; Denton, G.H. Reduced ice extent on the western antarctic peninsula at 700–970 cal. Yr B.P. Geology 2010, 38, 635–638. [Google Scholar] [CrossRef][Green Version]
- Jull, A.J.T.; Burr, G.S.; Hodgins, G.W.L. Radiocarbon dating, reservoir effects, and calibration. Quat. Int. 2013, 299, 64–71. [Google Scholar] [CrossRef]
- Negrete, J.; Soibelzon, E.; Tonni, E.P.; Carlini, A.; Soibelzon, L.H.; Poljak, S.; Huarte, R.A.; Carbonari, J.E. Antarctic radiocarbon reservoir: The case of the mummified crabeater seals (lobodon carcinophaga)in bodman cape, seymour island, antarctica. Radiocarbon 2011, 53, 161–166. [Google Scholar] [CrossRef]
- Murphy, E.J.; Clarke, A.; Abram, N.J.; Turner, J. Variability of sea-ice in the northern weddell sea during the 20th century. J. Geophys. Res. Ocean. 2014, 119, 4549–4572. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
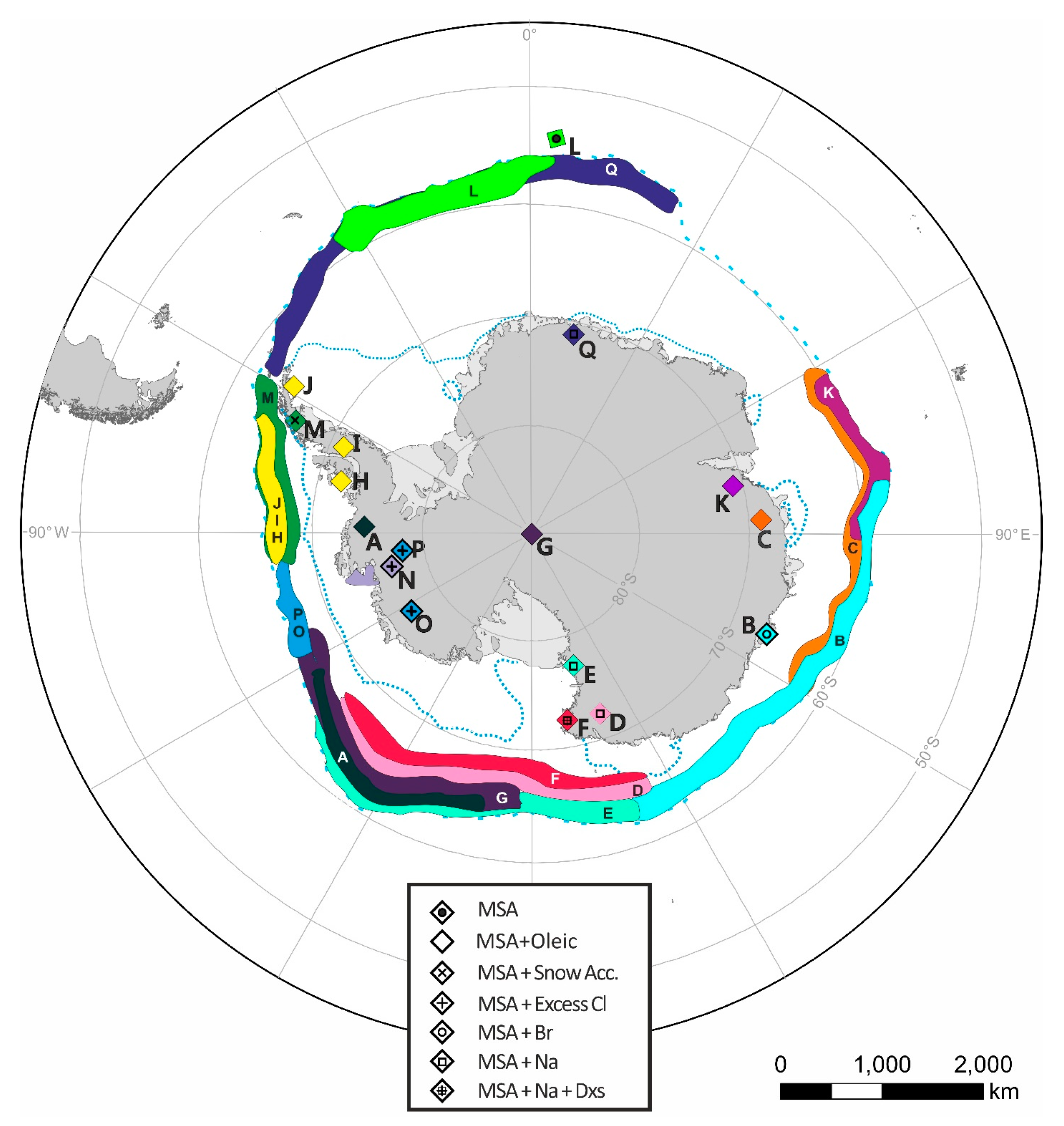
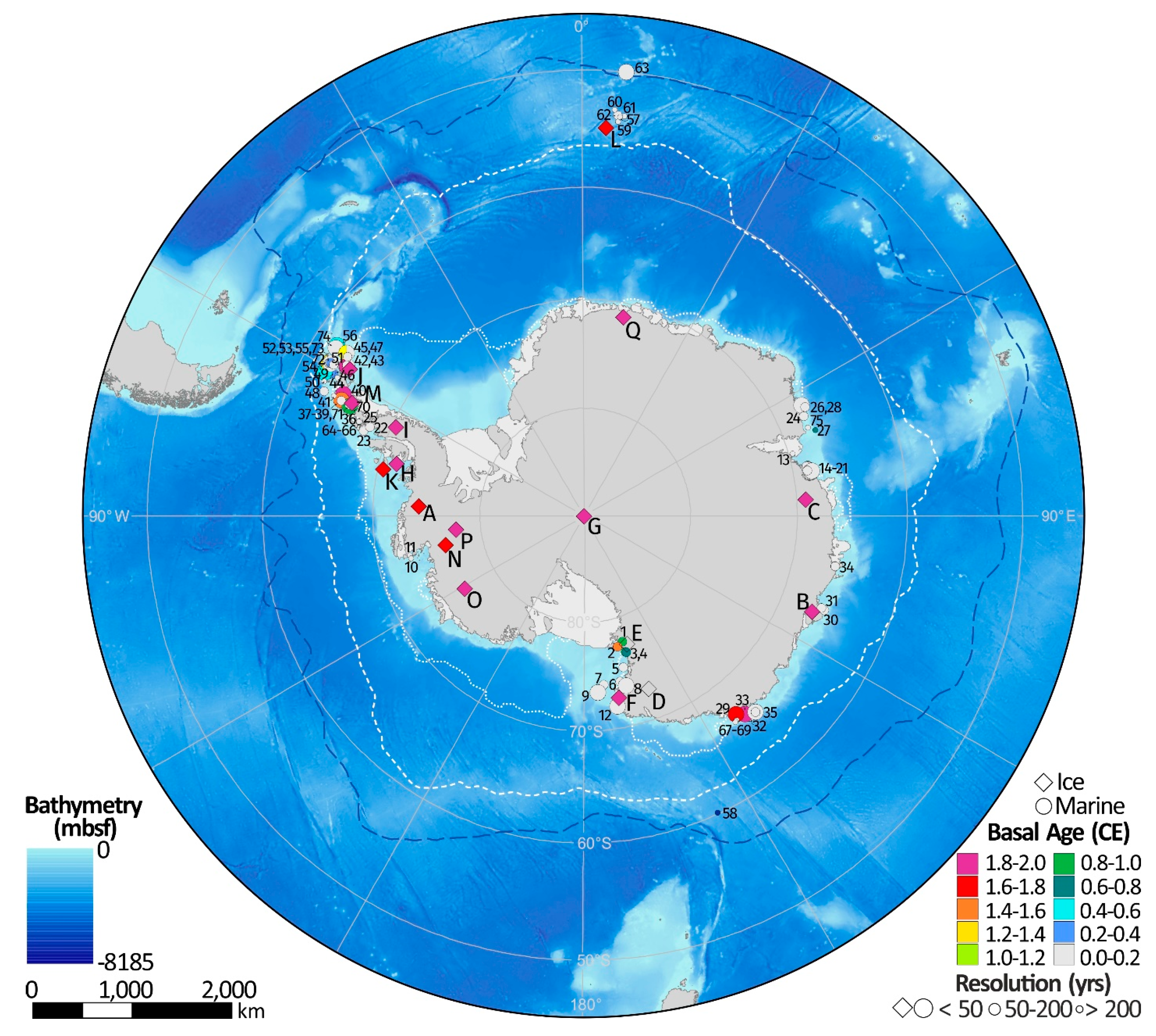
| Map Reference | Core ID | Latitude (°) | Longitude (°) | Location | Dates | Sea Ice Proxy | Reference |
|---|---|---|---|---|---|---|---|
| A | F10 | −74.57 | −86.90 | Ferrigno | 1703–2010 | MSA | [23] |
| B | DSS0506 | −66.77 | 112.81 | Law Dome (Summit) | 1841–2012 | MSA, Br | [24,25] |
| C | Mount Brown | −69.13 | 86.00 | Mount Brown, Wilkes Land | 1984–1999 | MSA | [26] |
| D | TALDICE | −72.80 | 159.06 | Talos Dome, East Antarctica | MSA, Na | [27,28] | |
| E | NG | −77.58 | 162.50 | Newell Glacier, Victoria Land | MSA | [29] | |
| F | WHG | −72.90 | 169.83 | Whitehall Glacier, Victoria Land | 1883–2004 | MSA, Na, Dxs | [30] |
| G | SP95 | −90.00 | 0.00 | South Pole | 1801–1991 | MSA | [31] |
| H | B92 | −71.90 | 74.60 | Beethoven Peninsula | 1949–1991 | MSA | [32] |
| I | DY90 | −70.68 | −64.87 | Dyer Plateau, Antarctic Peninsula | 1900–1988 | MSA | [32] |
| J | JRI97 | −64.22 | −57.68 | James Ross Island, Antarctic Peninsula | 1832–1997 | MSA | [32] |
| K | LGB69 | −70.83 | −77.07 | Princess Elizabeth Land, East Antarctica | 1745–1996 | MSA | [33] |
| L | B17 | −54.42 | 3.39 | Bouvet Island, South Atlantic | 2001–2017 | MSA, Oleic acid | [34] |
| M | BP | −66.04 | −64.08 | Bruce Plateau, Antarctic Peninsula | 1900–2009 | MSA, snow accumulation | [35] |
| N | DIV2010 | −76.77 | −101.74 | Thwaites Glacier, West Antarctica | 1786–2010 | MSA, ExCl- | [36,37] |
| O | THW2010 | −76.95 | −121.22 | Thwaites Glacier, West Antarctica | 1867–2010 | MSA, ExCl- | [36,37] |
| P | PIG2010 | −77.96 | −95.96 | Pine Island Glacier, West Antarctica | 1918–2010 | MSA, ExCl- | [36,37] |
| Q | IND-25/B5 | −71.33 | 11.58 | Central Dronning Maud Land | 1905–2005 | Na | [38] |
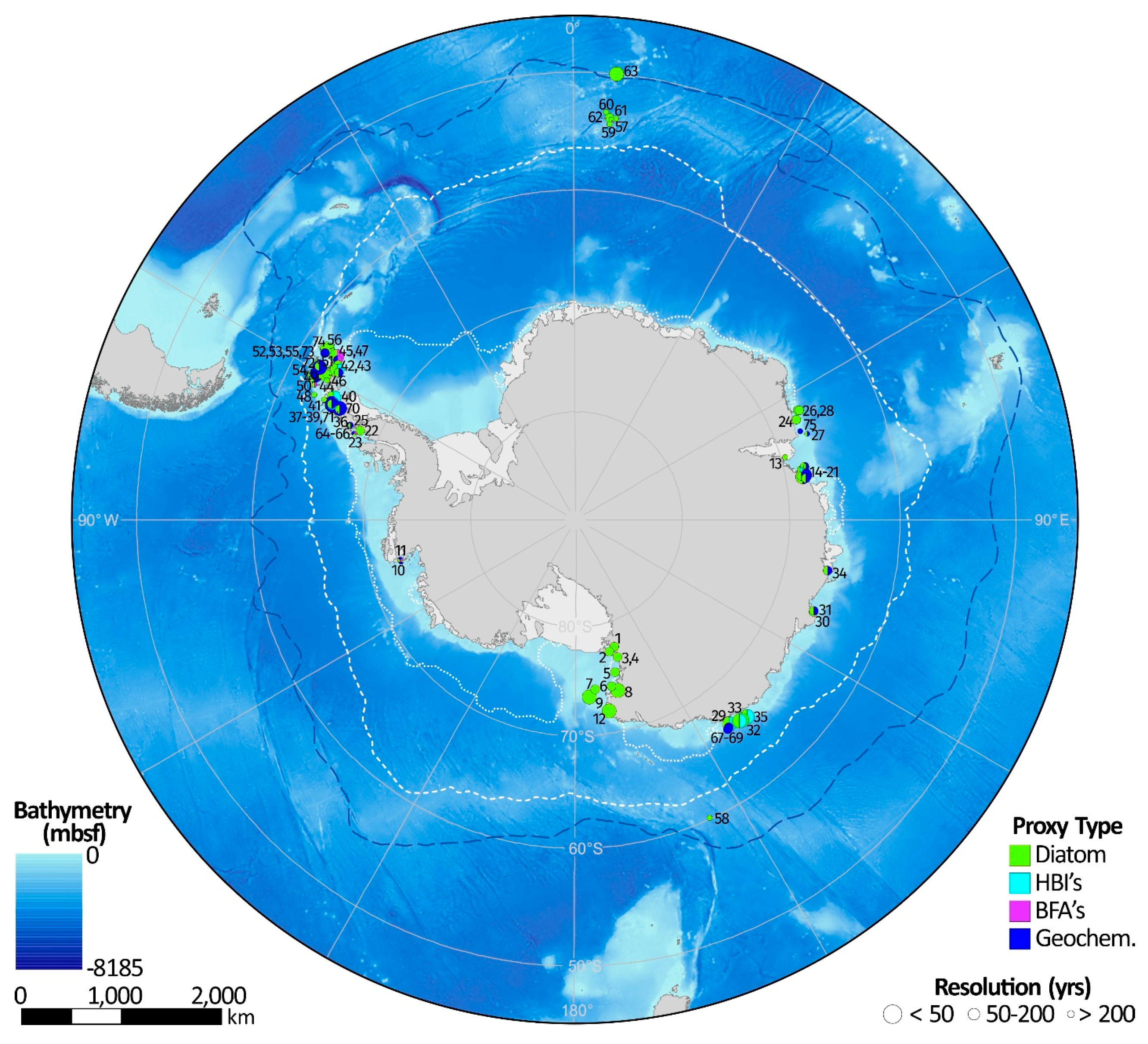
| Map Reference | Core/Site ID | Latitude (°) | Longitude (°) | Location Name | Dates ** | Resolution (Lowest) $ | Resolution (Highest) $ | Sea Ice Proxy | Reference(s) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | WG35 | −77.989 | 162.853 | Granite Harbor, Ross Sea | 1.2 to 0.6 ka BP | 60 | 32 | Diatom | [82] |
| 2 | Multiple | −77.668 | 165.500 | McMurdo Sound; Ross Sea | 0.5 to 0 ka BP | 100 | 56 | Diatom | [83] |
| 3 | WG17 | −77.000 | 162.850 | Granite Harbor, Ross Sea | 1.2 to 0.6 ka BP | 60 | 32 | Diatom | [82] |
| 4 | KC208.09 | −76.972 | 162.876 | Granite Harbor, Ross Sea | 1.3 to 0 ka BP | 65 | 33 | Diatom | [82] |
| 5 | KC31 | −75.700 | 165.418 | Western Ross Sea | 2.0 to 0 14C ka BP | 400 | 222 | Diatom | [84] |
| 6 | KC37 | −74.499 | 167.744 | Western Ross Sea | 2.0 to 0 14C ka BP | 400 | 222 | Diatom | [84] |
| 7 | KC39 | −74.474 | 173.474 | Western Ross Sea | 2.0 to 0 14C ka BP | 400 | 222 | Diatom | [84] |
| 8 | BAY05-43c | −74.000 | 166.050 | Wood Bay, Western Ross Sea | 2.0 to ~0.5 ka BP | 38 | 15 | Diatom | [52] |
| 9 | ANTA99-cJ5 | −73.817 | 175.650 | Joides Basin, Western Ross Sea | 2.0 to 0 ka BP | 23 | 9 | Diatom | [52] |
| 10 | KC17 | −73.420 | −102.827 | Ferrero Bay, Amundsen Sea Embayment | 2.0 to 0 ka BP | 2000 | 500 | Diatom; BFA; Geochemistry | [52] |
| 11 | KC15 | −73.360 | −101.836 | Ferrero Bay, Amundsen Sea Embayment | 2.0 to 0 ka BP | 2000 | 500 | Diatom; BFA; Geochemistry | [52] |
| 12 | BAY05-20c | −72.300 | 170.050 | Cape Hallet, Western Ross Sea | 2.0 to ~0.1 ka BP | 48 | 19 | Diatom | [52] |
| 13 | AM02 | −69.713 | 72.640 | Amery Ice Shelf, East Antarctica | 2.0 to 0.0 14C ka BP | 400 | 222 | Diatom | [85] |
| 14 | CO1011 | −68.827 | 77.760 | Flag Island Inlet, Rauer Group, Prydz Bay, EA | 2.0 to 0 ka BP | 2000 | 500 | Diatom | [86] |
| 15 | CO1010 | −68.817 | 77.833 | Filla Island Inlet, Rauer Group, Prydz Bay, EA | 2.0 to 0 ka BP | 400 | 222 | Diatom | [86] |
| 16 | JPC24 | −68.694 | 76.709 | Svenner Channel, Prydz Bay, EA | 2.0 to 0.6 ka BP | 70 | 36 | Diatom; HBIs | [87,88,89] |
| 17 | KROCK-15-GC29 | −68.664 | 76.696 | Prydz Bay, EA | 2.0 to 0 14C ka BP | 400 | 222 | Diatom | [90] |
| 18 | Abel Bay | −68.650 | 78.400 | Abel Bay, Vestfold Hills, EA | 2.0 to 0 ka BP | 200 | 51 | Diatom; Geochemistry | [91] |
| 19 | Watts Basin | −68.603 | 78.213 | Ellis Fjord, Vestfold Hills, EA | 2.0 to 0.2 14C ka BP | 45 | 18 | Diatom; Geochemistry | [92] |
| 20 | Deep Basin | −68.560 | 78.199 | Ellis Fjord, Vestfold Hills, EA | 2.0 to 0.8 14C ka BP | 30 | 12 | Diatom; Geochemistry | [92] |
| 21 | Platcha Bay | −68.515 | 78.478 | Platcha Bay, Vestfold Hills, EA | 2.0 to ~0.8 ka BP | 120 | 63 | Diatom; Geochemistry | [91] |
| 22 | JPC43 | −68.257 | −66.962 | Neny Fjord, Marguerite Bay, AP | 2.0 to 0 ka BP | 100 | 51 | Diatom | [93] |
| 23 | TPC522 | −67.856 | −68.205 | Marguerite Bay, AP | 2.0 to ~0.8 ka BP | 240 | 133 | Diatom; BFA; Geochemistry | [94] |
| 24 | KROCK-125-GC2 | −67.474 | 64.973 | Nielsen Bay, MacRobertson Shelf, EA | 2.0 to 0 14C ka BP | 100 | 133 | Diatom | [95] |
| 25 | GC1 | −67.180 | −66.797 | Lallemand Fjord, AP | 2.0 to 0 14C ka BP | 200 | 133 | Diatom; Geochemistry | [96,97] |
| 26 | JPC41 | −67.131 | 62.990 | Iceberg Alley, MacRobertson Shelf, EA | 2.0 to 0 ka BP | <1 * | Diatom | [98] | |
| 27 | GC 5 | −67.059 | 69.016 | MacRobertson Shelf, Prydz Bay, EA | 1.3 to 0 14C ka BP | 700 | 175 | Diatom; Geochemistry | [99] |
| 28 | KROCK-128-GC1 | −66.983 | 63.154 | Iceberg Alley, MacRobertson Shelf, EA | 2.0 to 0.2 14C ka BP | 100 | 51 | Diatom | [95] |
| 29 | CB2010 | −66.906 | 142.436 | Commonwealth Bay, Prydz Bay | 0.25 to 0 ka BP | 6 | 3 | Diatom; HBIs | [100] |
| 30 | PG1433 | −66.465 | 110.572 | Browning Bay, Windmill Islands, EA | 2.0 to 0.3 ka BP | 85 | 44 | Diatom | [101] |
| 31 | PG1430 | −66.453 | 110.498 | Peterson Inlet, Windmill Islands, EA | 2.0 to 0 ka BP | 100; 50 | 51; 20 | Diatom; Geochemistry | [101,102]; |
| 32 | MD03-2597 | −66.412 | 140.421 | Dumont d’Urville Trough, EA | 2.0 to 0.7 ka BP | <1 * | Diatom | [103] | |
| 33 | DTCI2010 | −66.411 | 140.445 | Dumont d’Urville Trough, EA | 0.04 to 0 ka BP | 0.4 | <0.4 | Diatom; HBIs | [104] |
| 34 | PG1173 | −66.267 | 100.750 | Rybiy Khvost Bay, Bunger Oasis, EA | 2.0 to 0 ka BP | 50; 100 | 20; 51 | Diatom; Geochemistry | [105] |
| 35 | MD03-2601 | −66.052 | 138.557 | Dumont d’Urville Trough, EA | 2.0 to 1.0 ka BP | 25; 50 | <10; 26 | Diatom; HBIs | [88,89,106,107,108,109] |
| 36 | WAP13-GC47 | −65.613 | −64.759 | Bigo Bay, AP | 2.0 to 0.1 ka BP | 190 | 100 | Diatom; Geochemistry | [110] |
| 37 | JPC10 | −64.883 | −64.200 | Palmer Deep, AP | 2.0 to 0.1 ka BP | 50 | 20 | Diatom; HBIs | [111] |
| 38 | PD92-30/178-1098A | −64.862 | −64.208 | Palmer Deep, AP | 2.0 to 0.1 ka BP | 200; 33 | 44; 13 | Diatom; Geochemistry | [96,111,112,113] |
| 39 | 178-1098B/C | −64.862 | −64.208 | Palmer Deep, AP | 2.0 to 0.2 ka BP | 100; 50; 18; | 51; 20; <18 | Diatom; BFA; Geochemistry | [114,115,116,117] |
| 40 | MTC18A | −64.772 | −62.829 | Andvord Drift, Gerlache Strait, AP | 0.13 to 0 ka BP | 3 | 1 | Diatom; HBIs | [118] |
| 41 | GC 02 | −64.000 | −64.000 | Anvers Shelf, AP | 2.0 to 0.9 14C ka BP | 220 | 122 | Diatom | [119] |
| 42 | KC2B | −63.971 | −57.759 | Herbert Sound, James Ross Island, AP | 2.0 to 0 ka BP | 200; 100 | 105; 51 | Diatom; Geochemistry | [120] |
| 43 | MTC38C; JPC38 | −63.717 | −57.411 | Vega Drift, Prince Gustav Channel, AP | 0.08 to 0 ka BP; 2.0 to 0 ka BP *** | 2; 50; 20 | 1; 20; <20 | Diatom; HBIs | [118,121] |
| 44 | PC61 | −63.389 | −60.319 | Bransfield Strait, AP | 2.0 to 0 ka BP | 200 | 105 | Diatom | [122] |
| 45 | JPC02 | −63.343 | −55.887 | Firth of Tay, AP | 2.0 to 0 ka BP | 200; 50; 50 | 105; 20; 20 | Diatom; BFA; Geochemistry | [123] [124] |
| 46 | A-3 | −63.168 | −59.302 | Bransfield Strait, AP | 1.7 to 0 ka BP | 95 | 44 | Diatom | [125,126] |
| 47 | JPC36 | −63.089 | −55.411 | Perseverence Drift, Joinville Island, AP | 0.8 to 0 ka BP; 2.0 to 0 ka BP *** | 40; 20 | 21; <20 | Diatom; BFA | [127] |
| 48 | GC 03 | −63.000 | −64.000 | Anvers Shelf, AP | 2.0 to 0.12 14C ka BP | 188 | 99 | Diatom | [119] |
| 49 | A-6 | −62.912 | −59.970 | Bransfield Strait, AP | 1.8 to 0.1 ka BP | 80 | 41 | Diatom | [125,126] |
| 50 | NCS 09 | −62.594 | −62.254 | Outer Shelf, S. Shetland Islands | 2.0 to 0 14C ka BP | 400 | 222 | Diatom; BFA | [119] |
| 51 | Gebra-2 | −62.589 | −58.542 | Bransfield Strait, AP | 2.0 to 0.2 ka BP | 90 | 46 | Diatom | [125,126] |
| 52 | 1B | −62.282 | −58.754 | Maxwell Bay, S. Shetland Islands | 2.0 to 0.1 ka BP | 1900; 200 | 475;105 | Diatom; Geochemistry | [128] |
| 53 | MC-01 | −62.202 | −58.727 | Marian Cove, S. Shetland Islands | 1.7 to 0 14C ka BP | 17 | <17 | Diatom; Geochemistry | [129] |
| 54 | WB2 | −62.200 | −60.700 | Outer Shelf, S. Shetland Islands | 1.5 to 0 14C ka BP | 38 | 15 | Diatom; Geochemistry | [130] |
| 55 | CB2 | −62.191 | −58.833 | Collins Harbour, Maxwell Bay, S. Shetland Islands | 2.0 to 0 14C ka BP | 20 | <20 | Diatom; Geochemistry | [131] |
| 56 | Gebra-1 | −61.943 | −55.170 | Bransfield Strait, AP | 1.6 to 0.1 ka BP | 90 | 15 | Diatom | [125,132] |
| 57 | 13PC (TN057-13PC4) | −53.200 | 5.100 | Cape Basin, South Atlantic | 2.0 to 0 ka BP | 20 | <20 | Diatom | [133] |
| 58 | E27-23 | −59.618 | 155.238 | Emerald Basin, South Indian-Pacific | 2.0 to 1.5 ka BP | 500 | 100 | Diatom | [134] |
| 59 | PS1652-2 | −53.664 | 5.100 | Cape Basin, South Atlantic | 2.0 to 0.6 ka BP | 280 | 156 | Diatom | [135] |
| 60 | PS1768-8 | −52.593 | 4.476 | Cape Basin, South Atlantic | 2.0 to 0.8 ka BP | 600 | 300 | Diatom | [135] |
| 61 | 177-1094/PS2090-1 | −53.179 | 5.132 | Cape Basin, South Atlantic | 2.0 to 0.8 ka BP | 240 | 133 | Diatom | [135,136] |
| 62 | PS2102-2 | −53.073 | 4.986 | Cape Basin, South Atlantic | 2.0 to 0.2 ka BP | 180 | 95 | Diatom | [135] |
| 63 | 17PC (TN057-17PC1) | −50.000 | 6.000 | Cape Basin, South Atlantic | 2.0 to 4.0 ka BP | 40 | 16 | Diatom | [133] |
| 64 | KC72 | −67.210 | −66.888 | Lallemand Fjord, AP | 2.0 to 0 14C ka BP | 200 | 105 | Geochemistry | [96] |
| 65 | KC5 | −67.166 | −66.950 | Lallemand Fjord, AP | 2.0 to 0 14C ka BP | 200 | 105 | Geochemistry | [96] |
| 66 | KC75 | −67.140 | −66.797 | Lallemand Fjord, AP | 2.0 to 0 14C ka BP | 200 | 105 | Geochemistry | [96] |
| 67 | 26PC12 | −66.560 | 142.950 | Mertz Ninnis Trough, EA | 2.0 to 0 14C ka BP | 2000 | 500 | Geochemistry | [137] |
| 68 | 17PC02 | −66.540 | 143.200 | Mertz Ninnis Trough, EA | 2.0 to 0 14C ka BP | 400 | 222 | Geochemistry | [137] |
| 69 | 11GC03 | −66.490 | 143.420 | Mertz Ninnis Trough, EA | 2.0 to 0 14C ka BP | 320 | 178 | Geochemistry | [137] |
| 70 | WAP13-GC45 | −65.752 | −64.525 | Bigo Bay, AP | 1.2 to 0.2 ka BP | 10 | <10 | Geochemistry | [110] |
| 71 | LMG_KC1 | −64.862 | −64.217 | Palmer Deep, AP | 0.5 to 0 ka BP | 25 | 13 | Geochemistry | [115] |
| 72 | WB1 | −62.300 | −60.100 | Outer Shelf, S. Shetland Islands | 0.8 to 0 14C ka BP | 20 | 8 | Geochemistry | [130] |
| 73 | CB1 | −62.173 | −58.826 | Collins Harbour, Maxwell Bay, S. Shetland Islands | 2.0 to 0 14C ka BP | 20 | <20 | Geochemistry | [131] |
| 74 | A9-EB2 | −61.982 | −55.957 | Bransfield Strait, AP | 2.0 to 0 ka BP | 50 | 20 | Geochemistry | [138] |
| 75 | PS69/849-2 | −67.583 | 68.125 | Burton Basin, MacRobertson Shelf, EA | 2.0 to 0.3 ka BP | 850 | 320 | Geochemistry | [139] |
| Archive | Advantage | Disadvantage |
|---|---|---|
| Ice cores |
|
|
| Marine sediments |
|
|
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomas, E.R.; Allen, C.S.; Etourneau, J.; King, A.C.F.; Severi, M.; Winton, V.H.L.; Mueller, J.; Crosta, X.; Peck, V.L. Antarctic Sea Ice Proxies from Marine and Ice Core Archives Suitable for Reconstructing Sea Ice over the Past 2000 Years. Geosciences 2019, 9, 506. https://doi.org/10.3390/geosciences9120506
Thomas ER, Allen CS, Etourneau J, King ACF, Severi M, Winton VHL, Mueller J, Crosta X, Peck VL. Antarctic Sea Ice Proxies from Marine and Ice Core Archives Suitable for Reconstructing Sea Ice over the Past 2000 Years. Geosciences. 2019; 9(12):506. https://doi.org/10.3390/geosciences9120506
Chicago/Turabian StyleThomas, Elizabeth R., Claire S. Allen, Johan Etourneau, Amy C. F. King, Mirko Severi, V. Holly L. Winton, Juliane Mueller, Xavier Crosta, and Victoria L. Peck. 2019. "Antarctic Sea Ice Proxies from Marine and Ice Core Archives Suitable for Reconstructing Sea Ice over the Past 2000 Years" Geosciences 9, no. 12: 506. https://doi.org/10.3390/geosciences9120506
APA StyleThomas, E. R., Allen, C. S., Etourneau, J., King, A. C. F., Severi, M., Winton, V. H. L., Mueller, J., Crosta, X., & Peck, V. L. (2019). Antarctic Sea Ice Proxies from Marine and Ice Core Archives Suitable for Reconstructing Sea Ice over the Past 2000 Years. Geosciences, 9(12), 506. https://doi.org/10.3390/geosciences9120506