
Palynostratigraphy of the “Muschelkalk Sedimentary Cycle” in the NW Iberian Range, Central Spain



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
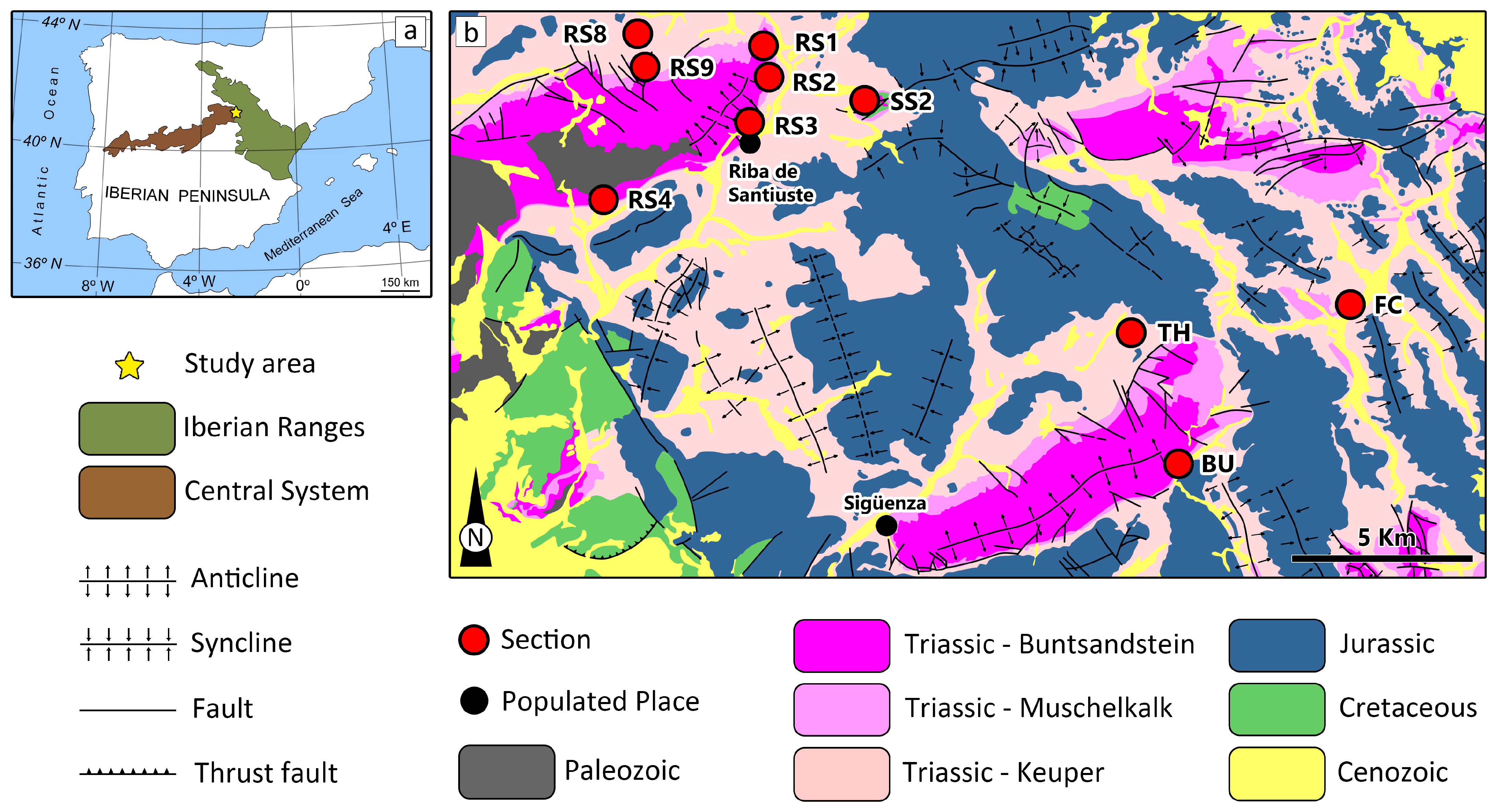
2. Geological Context
3. Materials and Methods
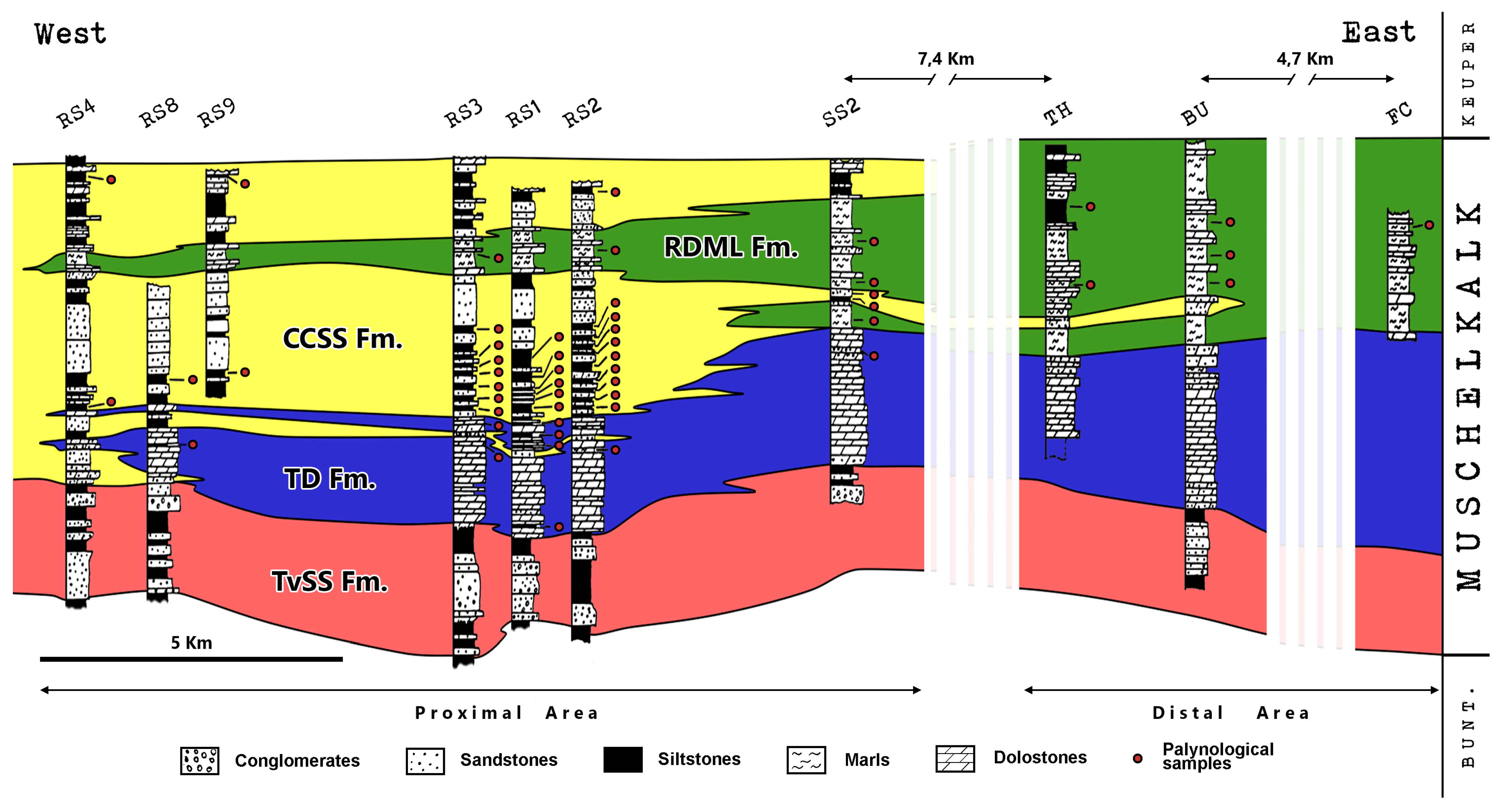
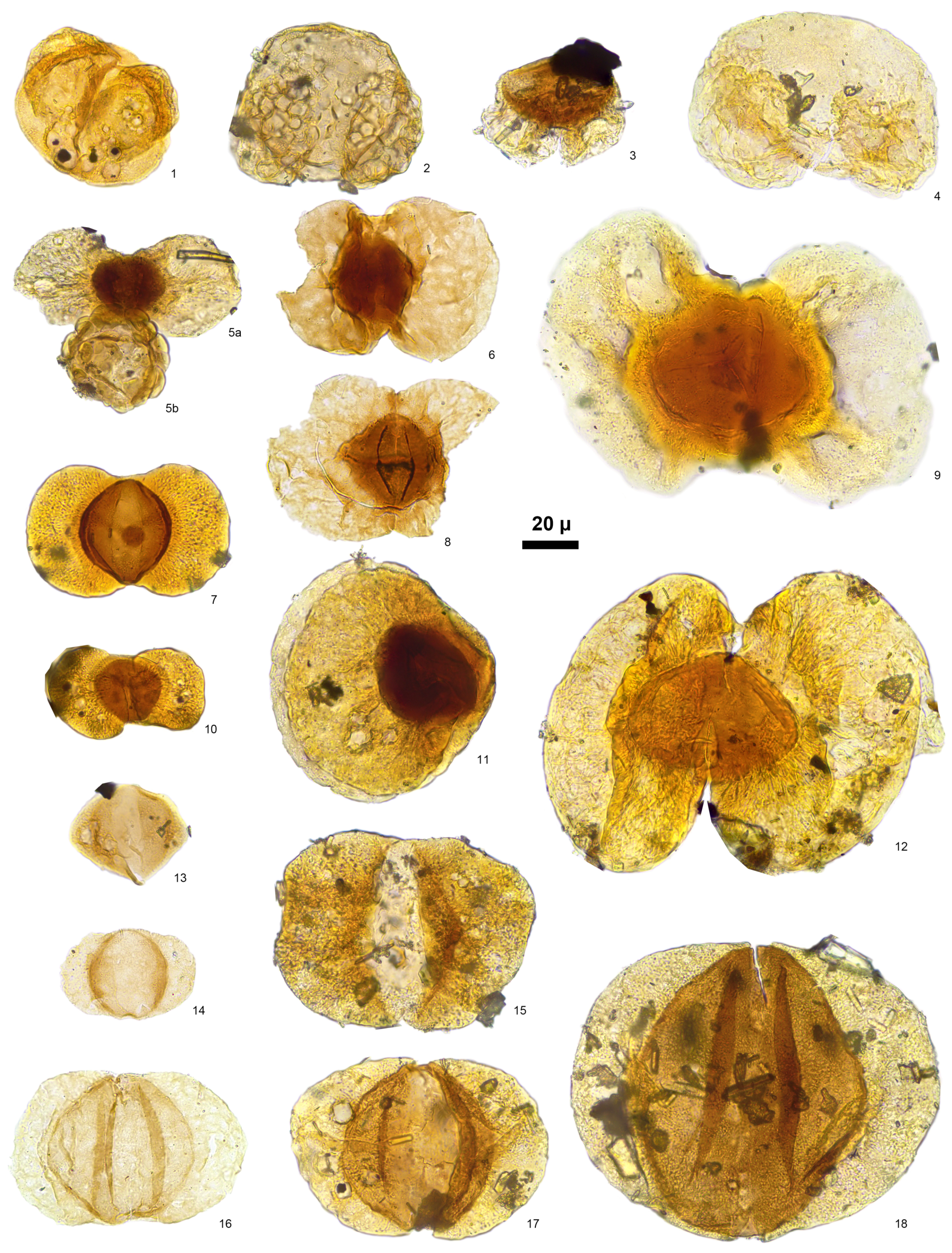
4. Results
4.1. Assemblage 1 (Section RS4, CCSS Fm., Sample RS4-2)
4.2. Assemblage 2 (Section RS8, TD Fm., Sample RS8-1)
4.3. Assemblage 3 (Section RS8, CCSS Fm., Sample RS8-2)
4.4. Assemblage 4 (Section RS9, CCSS Fm., Sample RS9-2)
4.5. Assemblage 5 (Section RS3, TD Fm., Sample RS3-2)
4.6. Assemblage 6 (Section RS3, CCSS Fm., Samples RS3-3 to RS3-6 and RS3-8)
4.7. Assemblage 7 (Section RS3, RDML Fm., Sample RS3-10)
4.8. Assemblage 8 (Section RS1, TD Fm., Samples RS1-1 to RS1-3)
4.9. Assemblage 9 (Section RS1, CCSS Fm., Samples RS1-5 to RS1-9)
4.10. Assemblage 10 (Section RS2, TD Fm., Sample RS2-1)
4.11. Assemblage 11 (Section RS2, CCSS Fm., Samples RS2-2, 4, 6 to 8, 10, and 12)
4.12. Assemblage 12 (Section RS2, RDML Fm., Sample RS2-11)
4.13. Assemblage 13 (Section SS2, TD Fm., Sample SS2-1)
4.14. Assemblage 14 (Section SS2, CCSS Fm., Sample SS2-3)
4.15. Assemblage 15 (Section SS2, RDML Fm., Samples SS2-2, SS2-5 and SS2-6)
4.16. Assemblage 16 (Section TH, RDML Fm., Samples TH-1 and TH-2)
4.17. Assemblage 17 (Section BU, RDML Fm., Samples BU-1 to BU-3)
4.18. Assemblage 18 (Section FC, RDML Fm., Sample FC-1)
5. Discussion
5.1. Reinterpretation of Previous Data
5.2. Interpretation of the New Data
5.3. Paleoecological and Paleoclimatic Interpretation and Taphonomic Bias
6. Conclusions
- –
- From the upper levels of the TD Fm. (inclusive) to the highest studied levels, the age of those levels can be placed within the Longobardian sensu lato, and possibly extending to the base of the Julian (Lower Carnian).
- –
- The contemporaneous age of CCSS, TD, and RDML formations supports the hypothesis that the termination of the two carbonate formations that constitute the upper Muschelkalk unit in the region (i.e., TD Fm. and RDML Fm.) is resolved by a lateral facies change to the terrigenous CCSS Fm.
- –
- The palynological assemblage indicates a warm and predominantly dry climate, dominated by xerophytic conifers adapted to arid conditions. However, the presence of hygrophytic spores and algae suggests the existence of localized wet areas, possibly associated with coastal or palustrine environments.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alberti, F. Ueberblick über Die Trias, mit Berücksichtigung Ihres Vorkommens in der Alpen; Verlag der J. G. Gottaschen Bachhandlung: Stuttgart, Germany, 1864; pp. 1–353. [Google Scholar] [CrossRef]
- Virgili, C.; Sopeña, A.; Ramos, A.; Hernando, S. Problemas de la cronoestratigrafía del Trías en España. Cuad. Geol. Ibérica 1977, 4, 57–88. [Google Scholar]
- Sopeña, A.; Virgili, C.; Arche, A.; Ramos, A.; Hernando, S. El Triásico. In Libro jubilar J.M. Rios, Geología de España; Instituto Geológico y Minero de España: Madrid, Spain, 1983; Volume 2, pp. 47–63. [Google Scholar]
- García-Gil, S. Estudio Sedimentológico y Paleogeográfico del Triásico en el Tercio Noroccidental de la Cordillera Ibérica (provincias de Gualajara y Soria). Ph.D. Thesis, Universidad Complutense Madrid, Madrid, Spain, 1989. [Google Scholar]
- García-Gil, S. Las unidades litoestratigráficas del Muschelkalk en el NW de la Cordillera Ibérica (España). Bol. R. Soc. Esp. Hist. Nat. 1991, 86, 21–51. [Google Scholar]
- García-Gil, S. The sedimentological significance of a clastic wedge in the western basin margin of the Triassic Tethys (Iberian Range, Spain). Cuad. Geol. Ibérica 1991, 15, 209–239. [Google Scholar]
- Virgili, C. El Triásico de las Catalánides. Bol. Inst. Geol. Mm. 1958, 69, 1–186. [Google Scholar]
- Dinarès-Turell, J.; Diez, J.B.; Rey, D.; Arnal, I. “Buntsandstein” magnetostratigraphy and biostratigraphic reappraisal from Eastern Iberia: Early and Middle Triassic stage boundary definitions through correlation to Tethyan sections. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 229, 158–177. [Google Scholar] [CrossRef]
- Ardell Argilés, F.; González Lodeiro, F.; Tena-Dávila Ruiz, M. Mapa geológico de la Hoja nº 343 (Barahona). Mapa Geológico de España E. 1:50.000. In Segunda Serie (MAGNA), Primera Edición; IGME: Madrid, Spain, 1982. [Google Scholar]
- Lendínez González, A. Mapa geológico de la Hoja nº 435 (Arcos de Jalón). Mapa Geológico de España E. 1:50.000. In Segunda Serie (MAGNA), Primera Edición; IGME: Madrid, Spain, 1991. [Google Scholar]
- Ardell Argilés, F.; González Lodeiro, F.; Tena-Dávila Ruiz, M. Mapa geológico de la Hoja nº 461 (Sigüenza). Mapa Geológico de España E. 1:50.000. In Segunda Serie (MAGNA), Primera Edición; IGME: Madrid, Spain, 1981. [Google Scholar]
- Bascones Alvira, L.; Martínez Álvarez, F. Mapa Geológico de la Hoja nº 462 (Maranchón); Mapa Geológico de España E. 1:50.000. Segunda Serie (MAGNA), Primera Edición; IGME: Madrid, Spain, 1981. [Google Scholar]
- Sopeña, A. Estratigrafía del Pérmico y Triásico del noroeste de la provincia de Guadalajara. In Seminarios de Estratigrafía, Serie Monografías 5; Universidad Complutense: Madrid, Spain, 1979; p. 329. [Google Scholar]
- Geiger, M.E.; Hopping, C.A. Triassic stratigraphy of the Southern North Sea Basin. In Philosophical Transactions of the Royal Society of London Series B; The Royal Society: London, UK, 1968; Volume 254, pp. 1–36. [Google Scholar]
- Hernando, S.; Doubinger, J.; Adloff, M.C. Datos cronoestratigráficos del Triásico superior de la región Ayllón-Atienza. Cuad. Geol. Ibérica 1977, 4, 399–410. [Google Scholar]
- Dunay, R.E.; Fisher, M.J. The Karnian palynofloral succession in the Northern Calcareous Alps, Lunz-am-See, Austria. Pollen Spores 1978, 20, 177–187. [Google Scholar]
- Visscher, H.; Krystyn, L. Aspects of late Triassic palynology. 4. A palynological assemblage from ammonoid-controlled Late Karnian (Tuvalian) sediments of Sicily. Rev. Palaeobot. Palynol. 1978, 26, 93–112. [Google Scholar] [CrossRef]
- Ramos, A. Estratigrafía y paleogeografía del Pérmico y Triásico al Oeste de Molina de Aragón (provincia de Guadalajara). In Seminarios de Estratigrafía, Serie Monografías 6; Universidad Complutense: Madrid, Spain, 1979; p. 313. [Google Scholar]
- García-Gil, S.; Sopeña, A. Estratigrafía y Sedimentología del Triásico en el sector medinaceli-Somaén (Provincia de Soria). Cuad. Geol. Ibérica 1987, 11, 707–735. [Google Scholar]
- Hinkelbein, K. El Triásico y el Jurásico de los alrededores de Albarracín. Teruel 1969, 4, 35–75. [Google Scholar]
- Pérez-Arlucea, M.; Sopeña, A. Estratigrafía del Pérmico y Triásico en el sector central de la Rama Castellana de la Cordillera Ibérica. Estud. Geol. 1985, 41, 207–222. [Google Scholar] [CrossRef]
- Diez, J.B. Geología y Palaeobotánica de la Facies Buntsandstein en la Rama Aragonesa de la Cordillera Ibérica. Implicaciones Bioestratigráficas en el Peritethys Occidental. Ph.D. Thesis, Universidad de Zaragoza (Spain), Zaragoza, Spain, Université Paris VI (France), Paris, France, 2000. [Google Scholar]
- García-Ávila, M.; De la Horra, R.; de Miguel Chaves, C.; Juncal, M.A.; Pérez-García, A.; Ortega, F.; Diez, J.B. Palynological and sedimentological implications of the sauropterygian Upper Triassic site of El Atance (Central Iberian Peninsula). Rev. Palaeobot. Palynol. 2021, 295, 104541. [Google Scholar] [CrossRef]
- Wood, D.G.; Gabriel, A.M.; Lawson, J.C. Palynological techniques—Processing and microscopy. In Palynology: Principles and Applications; Jansonius, J., McGregor, D.C., Eds.; American Association of Stratigraphic Palynologists Foundation: Houston, TX, USA, 1996; Volume 1, pp. 29–50. [Google Scholar]
- Visscher, H.; Brugman, W.A. Ranges of selected palynomorphs in the Alpine Triassic of Europe. Rev. Palaeobot. Palynol. 1981, 34, 115–128. [Google Scholar] [CrossRef]
- Besems, R.E. Aspects of Middle and Late Triassic Palynology. 1. Palynostratigraphical data from the Chiclana de Segura Formation of the Linares-Alcaraz region (southeastern Spain) and correlation with palynological assemblages from the Iberian Peninsula. Rev. Palaeobot. Palynol. 1981, 32, 257–273. [Google Scholar] [CrossRef]
- Van Der Eem, J.G.L.A. Aspects of Middle and Late Triassic Palynology: 6. Palynological investigations in the Ladinian and Lower Karnian of the Western Dolomites, Italy. Rev. Palaeobot. Palynol. 1983, 39, 189–300. [Google Scholar] [CrossRef]
- Roghi, G. Analisi Palinologica del Trias Medio del Sudalpino. Ph.D. Thesis, University of Padova, Padova, Italy, 1995. [Google Scholar]
- Stockar, R.; Baumgartner, P.O.; Condon, D. Integrated Ladinian bio-chronostratigraphy and geochronology of Monte San Giorgio (Southern Alps, Switzerland). Swiss J. Geosci. 2012, 105, 85–108. [Google Scholar] [CrossRef]
- Doubinger, J.; Bühmann, D. Röt bei Borken und bei Schlüchtern (Hessen, Deutschland). Palynologie und Tonmineralogie. Z. Dtsch. Geol. Ges. 1981, 132, 421–449. [Google Scholar] [CrossRef]
- Doubinger, J.; Adloff, M.C. Triassic Palynomorphs of the Mediterranean Area; Centre de Sédimentologie et Géochimie de la Surface: Strasbourg, France, 1983. [Google Scholar]
- Orłowska-Zwolińska, T. Palynostratigraphy of the upper part of Triassic Epicontinental sediments in Poland. Pr. Inst. Geol. 1983, 104, 1–89. [Google Scholar]
- Orłowska-Zwolińska, T. Palynological zones of the Polish epicontinental Triassic. Bull. Pol. Acad. Sci. Earth Sci. 1985, 33, 107–117. [Google Scholar]
- Eshet, Y. The palynostratigraphy of the Permian Triassic boundary in Israel: Two approaches to biostratigraphy. Isr. J. Earth Sci. 1990, 39, 1–15. [Google Scholar]
- Kürschner, W.M.; Waldemaar Herngreen, G.F.W. Triassic palynology of central and northwestern Europe: A review of palynofloral diversity patterns and biostratigraphic subdivisions. In Triassic Timescale; Lucas, S.G., Ed.; Geological Society of London: London, UK, 2010; Special Publications 334; pp. 263–283. [Google Scholar] [CrossRef]
- Scheuring, B.W. Palynologische und palynostratigraphische Utersuschungen des Keupers im Bölchentunnel (Solothurner Jura). Schweiz. Paläont. Abh. 1970, 88, 1–187. [Google Scholar]
- Scheuring, B.W. Mikrofloren aus den Meridelkalken des Monte San Giorgio (Kanton Tessin). Schweiz. Paläont. Abh. 1978, 100, 1–205. [Google Scholar]
- Visscher, H.; Van Route, M.; Brugman, W.A.; Poort, R.J. Rejection of a Carnian (Late Triassic) “pluvial event” in Europe. Rev. Palaeobot. Palynol. 1994, 83, 217–226. [Google Scholar] [CrossRef]
- Roghi, G. Palynological investigations in the Carnian of the Cave del Predil area (Julian Alps, NE Italy). Rev. Palaeobot. Palynol. 2004, 132, 1–35. [Google Scholar] [CrossRef]
- Zavialova, N.E.; Roghi, G. Exine morphology and ultrastructure of Duplicisporites from the Triassic of Italy. Grana 2005, 44, 337–342. [Google Scholar] [CrossRef]
- Buratti, N.; Cirilli, S. Microfloristic provincialism in the Upper Triassic Circum-Mediterranean area and palaeogeographic implication. Geobios 2007, 40, 133–142. [Google Scholar] [CrossRef]
- Roghi, G.; Gianolla, P.; Minarelli, L.; Pilati, C.; Preto, N. Palynological correlation of Carnian humid pulses throughout western Tethys. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 290, 89–106. [Google Scholar] [CrossRef]
- Kustatscher, E.; Ash, S.; Karasev, E.; Pott, C.; Vajda, V.; Yu, J.; McLoughlin, S. Flora of the Late Triassic. In The Late Triassic World; Tanner, L.H., Ed.; Springer: Berlin/Heidelberg, Germany, 2017; Topics in Geobiology; 2018; Volume 46, pp. 545–622. [Google Scholar] [CrossRef]
- Balme, B.E. Fossil in situ spores and pollen grains: An annotated catalogue. Rev. Palaeobot. Palynol. 1995, 87, 1–323. [Google Scholar] [CrossRef]
- Looy, C.V.; Brugman, W.A.; Dilcher, D.L.; Visscher, H. The delayed resurgence of equatorial forests after the Permian–Triassic ecologic crisis. Proc. Natl. Acad. Sci. USA 1999, 96, 13857–13862. [Google Scholar] [CrossRef]
- Kustatscher, E.; Heunisch, C.; Van Konijnenburg-Van Cittert, J.H.A. Taphonomical implications of the Ladinian megaflora and palynoflora of Thale (Germany). Palaios 2012, 27, 753–764. [Google Scholar] [CrossRef]
- Lindström, S.; Erlström, M.; Piasecki, S.; Nielsen, L.H.; Mathiesen, A. Palynology and terrestrial ecosystem change of the Middle Triassic to lowermost Jurassic succession of the eastern Danish Basin. Rev. Palaeobot. Palynol. 2017, 244, 65–95. [Google Scholar] [CrossRef]
- Brugman, W.A.; Van Bergen, P.R.; Kerp, J.H.E. A quantitative approach to Triassic palynology: The Lettenkeuper of the Germanic Basin as an example. In Sedimentation of Organic Particles; Traverse, A., Ed.; Cambridge University Press: Cambridge, UK, 1994; pp. 409–430. [Google Scholar] [CrossRef]
- Césari, S.N.; Colombi, C. Palynology of the Late Triassic Ischigualasto Formation, Argentina: Paleoecological and paleogeographic implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 449, 365–384. [Google Scholar] [CrossRef]
- Mueller, S.; Hounslow, M.W.; Kürschner, W.M. Integrated stratigraphy and palaeoclimate history of the Carnian Pluvial Event in the Boreal realm; new data from the Upper Triassic Kapp Toscana Group in central Spitsbergen (Norway). J. Geol. Soc. 2015, 173, 186–202. [Google Scholar] [CrossRef]
- Hochuli, P.A.; Hermann, E.; Vigran, J.O.; Bucher, H.; Weissert, H. Rapid demise and recovery of plant ecosystems across the end-Permian extinction event. Glob. Planet. Change 2010, 74, 144–155. [Google Scholar] [CrossRef]
- Lindström, S.; Vosgerau, H.; Piasecki, S.; Nielsen, L.H.; Dybkjær, K.; Erlström, M. Ladinian palynofloras in the Norwegian–Danish Basin: A regional marker reflecting a climate change. GEUS Bull. 2009, 17, 21–24. [Google Scholar] [CrossRef]
- Hauschke, N.; Heunisch, C. Lithologie und Palynologie der Bohrung USB 3 (Horn-Bad Meinberg, Ostwestfalen): Ein Beitrag zur Faziesentwicklung im Keuper. Neues Jahrb. Geol. Paläontol. Abh. 1990, 181, 79–105. [Google Scholar] [CrossRef]
- Vigran, J.O.; Mangerud, G.; Mørk, A.; Worsley, D.; Hochuli, P.A. Palynology and Geology of the Triassic Succession of Svalbard and the Barents Sea; Special Publication, 14; Geological Survey of Norway: Trondheim, Norway, 2014; p. 247. [Google Scholar] [CrossRef]
- Duringer, P.; Doubinger, J. La palynologie: Un outil de caractérisation des faciès marins et continentaux à la limite Muschelkalk supérieur—Lettenkohle. Sci. Géol. Bull. 1985, 38, 19–34. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Ávila, M.; García-Gil, S.; Diez, J.B. Palynostratigraphy of the “Muschelkalk Sedimentary Cycle” in the NW Iberian Range, Central Spain. Geosciences 2025, 15, 299. https://doi.org/10.3390/geosciences15080299
García-Ávila M, García-Gil S, Diez JB. Palynostratigraphy of the “Muschelkalk Sedimentary Cycle” in the NW Iberian Range, Central Spain. Geosciences. 2025; 15(8):299. https://doi.org/10.3390/geosciences15080299
Chicago/Turabian StyleGarcía-Ávila, Manuel, Soledad García-Gil, and José B. Diez. 2025. "Palynostratigraphy of the “Muschelkalk Sedimentary Cycle” in the NW Iberian Range, Central Spain" Geosciences 15, no. 8: 299. https://doi.org/10.3390/geosciences15080299
APA StyleGarcía-Ávila, M., García-Gil, S., & Diez, J. B. (2025). Palynostratigraphy of the “Muschelkalk Sedimentary Cycle” in the NW Iberian Range, Central Spain. Geosciences, 15(8), 299. https://doi.org/10.3390/geosciences15080299