

New Lizard Specimens from the Campanian Wapiti Formation of Alberta, Canada

 ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
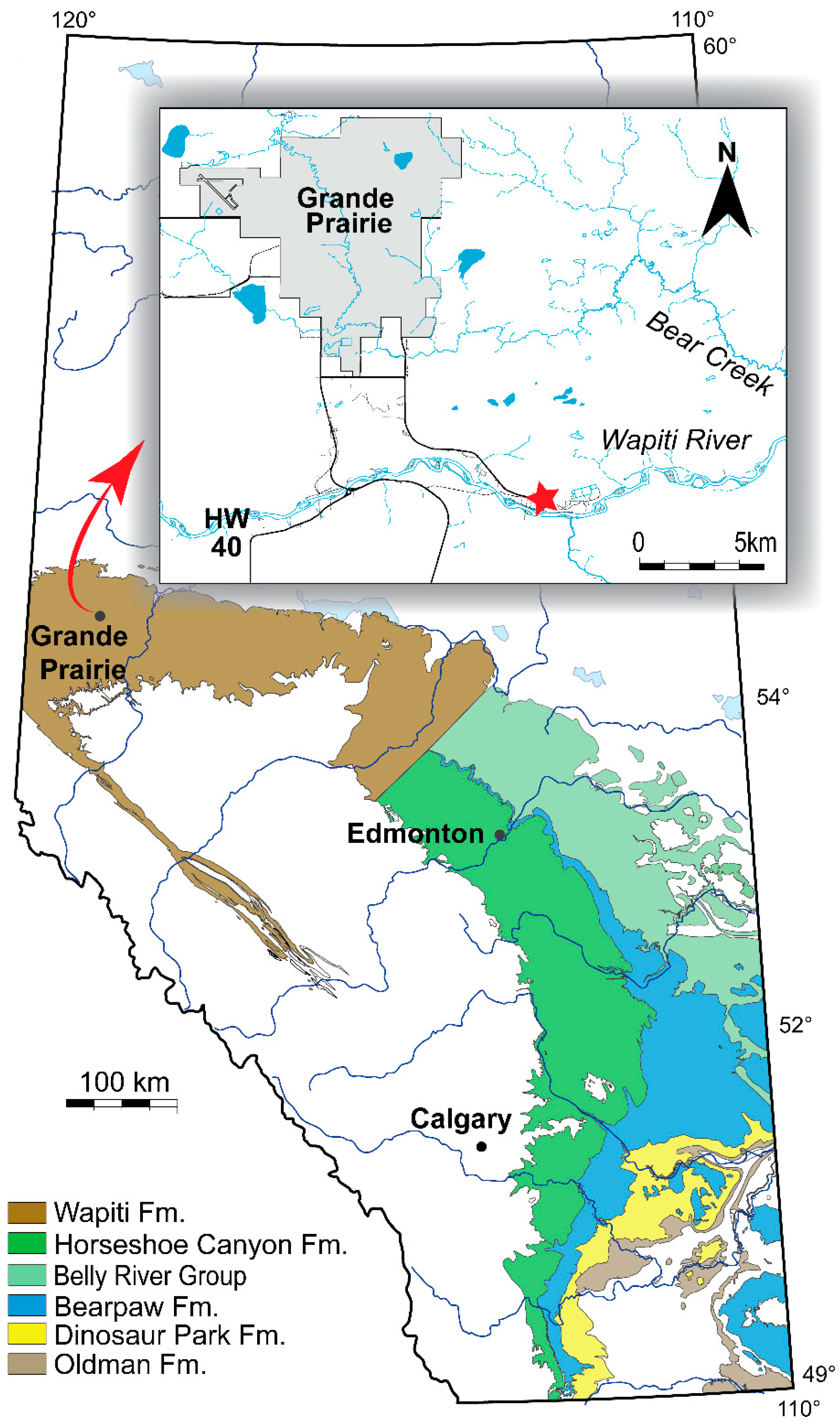
2. Geological Setting
3. Materials and Methods
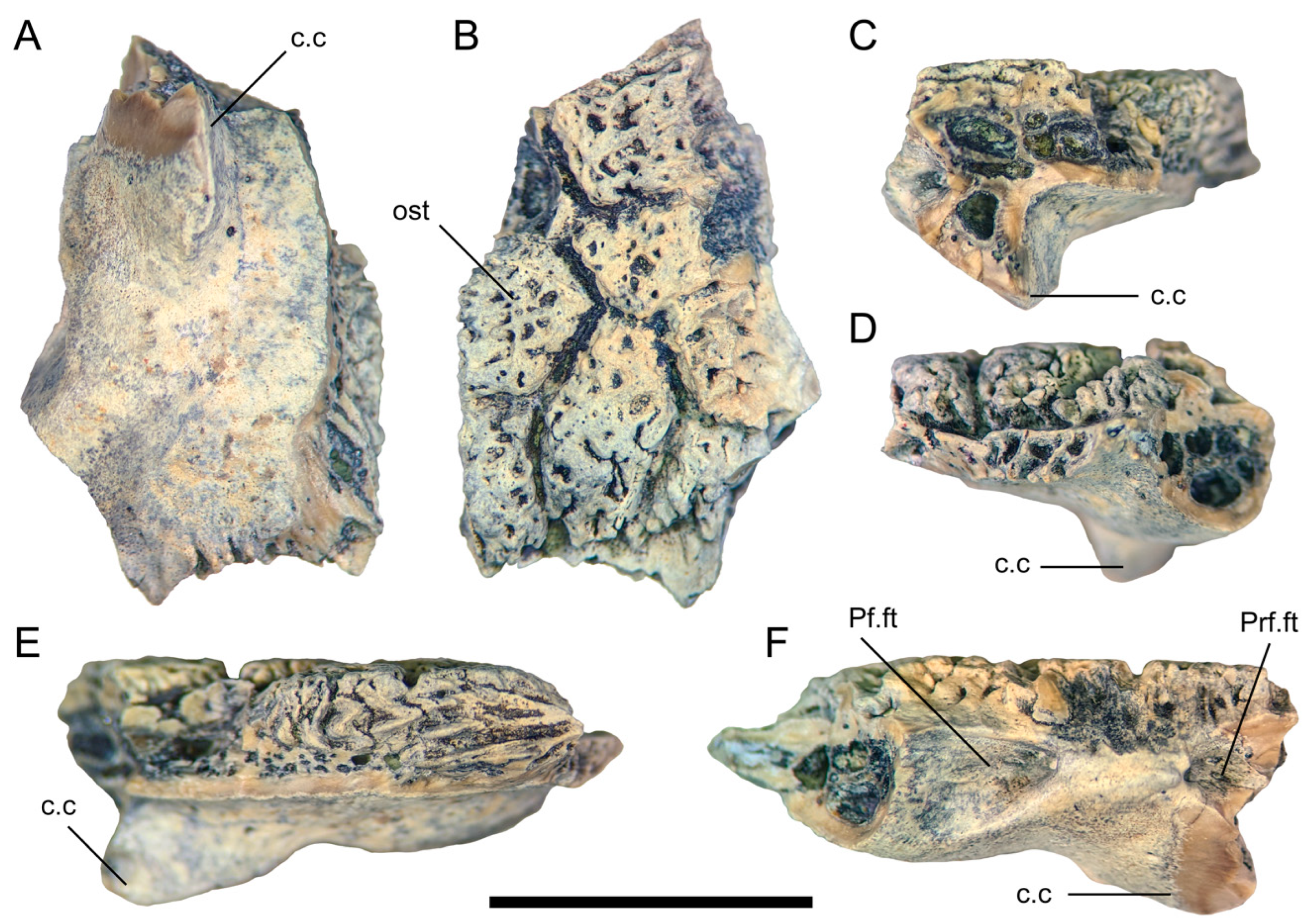
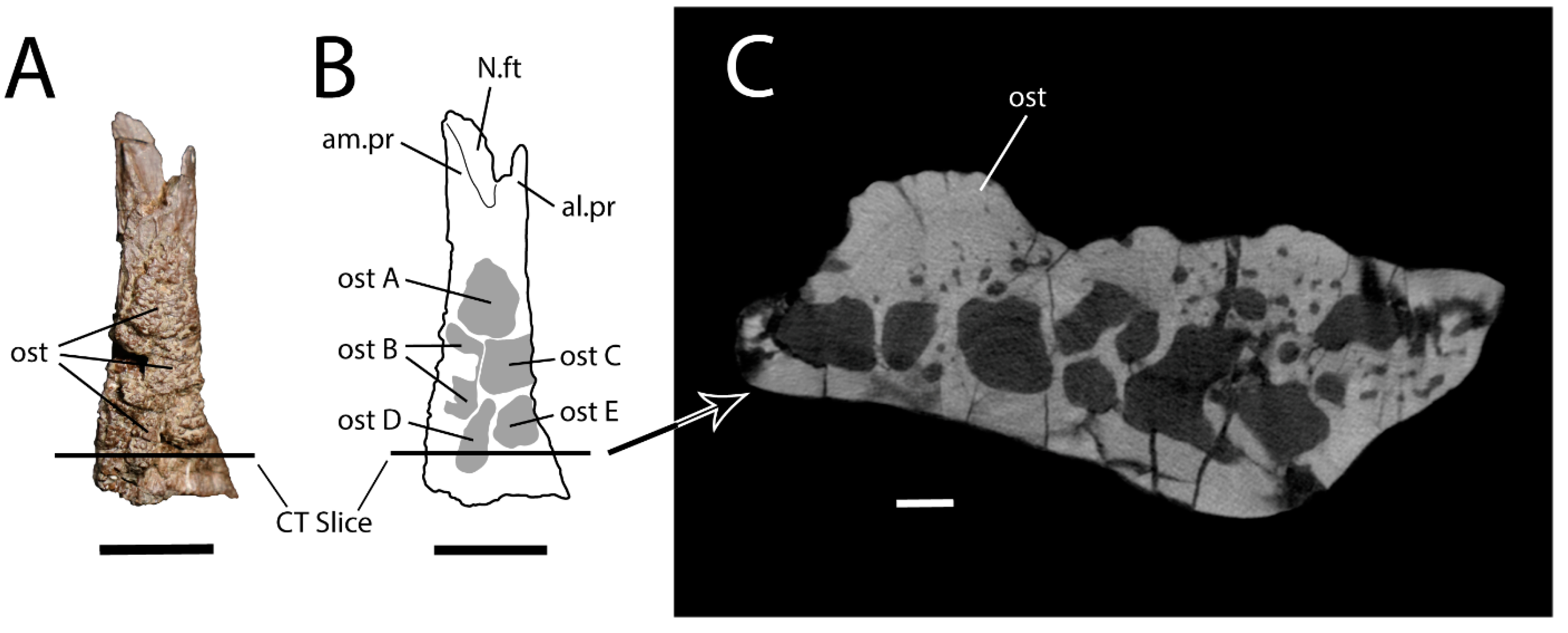
4. Systematic Palaeontology
5. Discussion
5.1. Taxonomic Identification and Ontogenetic Assessment of UALVP 59503
5.2. Identification of UALVP 59947
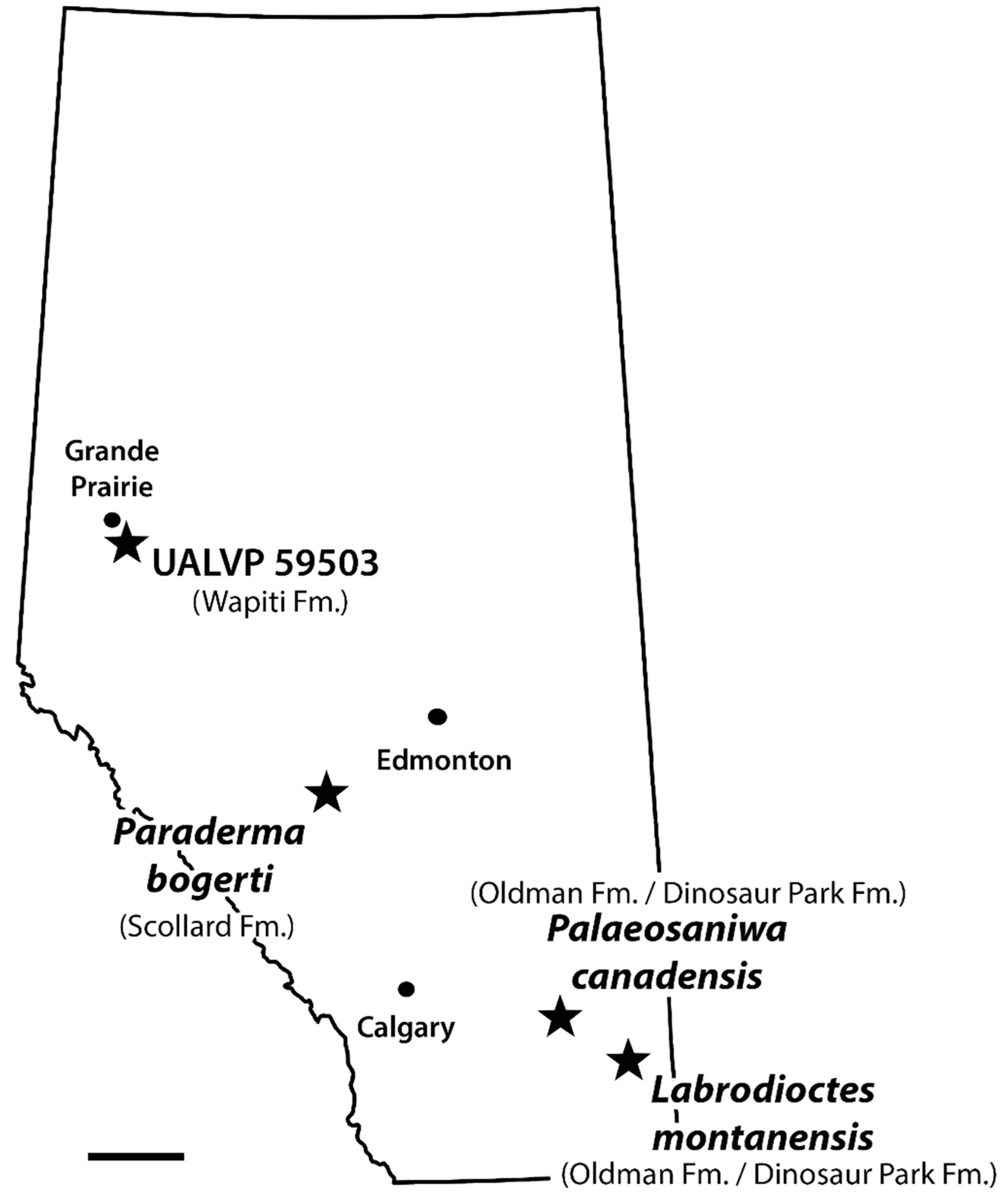
5.3. Taxonomic Status of Labrodioctes montanensis and Palaeosaniwa canadensis
5.4. Implications of the Presence of a Monstersaur cf. Palaeosaniwa canadensis in Unit 3 of the Wapiti Formation
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reiserer, R.S.; Schuett, G.W.; Beck, D.D. Taxonomic reassessment and conservation status of the beaded lizard, Heloderma horridum (Squamata: Helodermatidae). Amphib. Reptile Conserv. 2013, 7, 74–96. [Google Scholar]
- Norell, M.A.; Gao, K. Braincase and phylogenetic relationships of Estesia mongoliensis from the Late Cretaceous of the Gobi Desert and the recognition of a new clade of lizards. Am. Mus. Novit. 1997, 3211, 1–25. [Google Scholar]
- Conrad, J. Phylogeny and systematics of Squamata (Reptilia) based on morphology. Bull. Am. Mus. Nat. Hist. 2008, 310, 1–182. [Google Scholar] [CrossRef]
- Burbrink, F.T.; Grazziotin, F.G.; Pyron, R.A.; Cundall, D.; Donnellan, S.; Irish, F.; Keogh, J.S.; Kraus, F.; Murphy, R.W.; Noonan, B.; et al. Interrogating genomic-scale data for Squamata (lizards, snakes and amphisbaenians) shows no support for key traditional morphological relationships. Syst. Biol. 2020, 69, 502–520. [Google Scholar] [CrossRef]
- Conrad, J.L.; Rieppel, O.; Gautier, J.A.; Norell, M.A. Osteology of Gobiderma pulchrum (Monstersauria, Lepidosauria, Reptilia). Bull. Am. Mus. Nat. Hist. 2011, 362, 1–88. [Google Scholar] [CrossRef]
- Conrad, J.L.; Ast, J.C.; Montanari, S.; Norell, M.A. A combined evidence phylogenetic analysis of Anguimorpha (Reptilia: Squamata). Cladistics 2011, 27, 230–277. [Google Scholar] [CrossRef]
- Reeder, T.W.; Townsend, T.M.; Mulcahy, D.G.; Noonan, B.P.; Wood, P.L., Jr.; Sites, J.W., Jr.; Wiens, J.J. Integrated analyses resolve conflicts over squamate reptile phylogeny and reveal unexpected placements for fossil taxa. PLoS ONE 2015, 10, e0118199. [Google Scholar] [CrossRef]
- Simões, T.R.; Caldwell, M.W.; Tałanda, M.; Bernardi, M.; Palci, A.; Vernygora, O.; Bernardini, F.; Mancini, L.; Nydam, R.L. The origin of squamates revealed by a Middle Triassic lizard from the Italian Alps. Nature 2018, 557, 706–709. [Google Scholar] [CrossRef]
- Gauthier, J.A.; Kearney, M.; Maisano, J.A.; Rieppel, O.; Behlke, A.D.B. Assembling the squamate tree of life: Perspectives from the phenotype and the fossil record. Bull. Peabody Mus. Nat. Hist. 2012, 53, 3–308. [Google Scholar] [CrossRef]
- Longrich, N.R.; Bhullar, B.-A.S.; Gauthier, J.A. Mass extinction of lizards and snakes at the Cretaceous-Paleogene boundary. Proc. Acad. Natl. Sci. USA 2012, 109, 21396–21401. [Google Scholar] [CrossRef]
- Gao, K.; Norell, M.A. Taxonomic revision of Carusia (Reptilia: Squamata) from the Late Cretaceous of the Gobi desert and phylogenetic relationships of anguimorphan lizards. Am. Mus. Novit. 1998, 3230, 1–51. [Google Scholar]
- Nydam, R.L. A new taxon of helodermatid-like lizard from the Albian–Cenomanian of Utah. J. Vertebr. Paleontol. 2000, 20, 285–294. [Google Scholar] [CrossRef]
- Beck, D.D. Biology of Gila Monsters and Beaded Lizards; University of California Press: Berkeley, CA, USA, 2005; pp. 1–211. ISBN 978-0-520-25987-4. [Google Scholar]
- Bogert, C.; del Campo, M. The Gila monster and its allies. Bull. Am. Mus. Nat. Hist. 1956, 109, 1–238. [Google Scholar]
- Fry, B.G.; Vidal, N.; Norman, J.A.; Vonk, F.J.; Scheib, H.; Ramjan, S.F.R.; Kuruppu, S.; Fung, K.; Hedges, S.B.; Richardson, M.K.; et al. Early evolution of the venom system in lizards and snakes. Nature 2006, 439, 584–588. [Google Scholar] [CrossRef]
- Estes, R. Fossil vertebrates from the Late Cretaceous Lance Formation eastern Wyoming. Univ. Calif. Pub. Geol. Sci. 1964, 49, 1–180. [Google Scholar]
- Borsuk-Białynicka, M. Anguimorphans and related lizards from the Late Cretaceous of the Gobi Desert, Mongolia. Palaeontol. Pol. 1984, 46, 5–105. [Google Scholar]
- Pregill, G.K.; Gauthier, J.A.; Greene, H.W. The evolution of helodermatid squamates, with description of a new taxon and an overview of Varanoidea. San. Dieg. Soc. Nat. Hist. 1986, 21, 167–202. [Google Scholar]
- Gao, K.; Fox, R.C. Taxonomy and evolution of Late Cretaceous lizards (Reptilia: Squamata) from western Canada. Bull. Carn. Mus. Nat. Hist. 1996, 3, 1–107. [Google Scholar] [CrossRef]
- Mead, J.I.; Schubert, B.W.; Wallace, S.C.; Swift, S.L. Helodermatid lizard from the Mio-Pliocene oak-hickory forest of Tennessee, eastern USA, and a review of monstersaurian osteoderms. Acta Palaeontol. Pol. 2012, 57, 111–121. [Google Scholar] [CrossRef]
- Estes, R. Sauria Terrestria, Amphisbaenia. Handbuch der Paläoherpetologie, Part 10A; Gustav Fisher Verlag: Stuttgart, Germany, 1983; ISBN 9783437303913. [Google Scholar]
- Norell, M.A.; McKenna, M.C.; Novacek, M.J. Estesia mongoliensis, a new fossil varanoid from the Late Cretaceous Barun Goyot Formation of Mongolia. Am. Mus. Novit. 1992, 3025, 1–24. [Google Scholar]
- Nydam, R.L. Squamates from the Jurassic and Cretaceous of North America. Paleobio. Palaeoenv. 2013, 93, 535–565. [Google Scholar] [CrossRef]
- Yi, H.Y.; Norell, M.A. New materials of Estesia mongoliensis (Squamata: Anguimorpha) and the evolution of venom grooves in lizards. Am. Mus. Novit. 2013, 3767, 1–31. [Google Scholar] [CrossRef]
- Balsai, M.J. The Phylogenetic Position of Palaeosaniwa and the Early Evolution of the Platynotan (Varanoid) Anguimorphs. Ph.D. Thesis, University of Pennsylvania, Philadelphia, PA, USA, 2001; pp. 1–359. [Google Scholar]
- Gilmore, C.W. Fossil lizards of North America. Natl. Acad. Sci. Mem. 1928, 22, 1–201. [Google Scholar]
- Makádi, L.; Caldwell, M.W.; Ősi, A. The first freshwater mosasauroid (Upper Cretaceous, Hungary) and a new clade of basal mosasauroids. PLoS ONE 2012, 7, e51781. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.C.A. Fossil Vertebrates from the Cerro del Pueblo Formation, Coahuila, Mexico, and the Distribution of Late Campanian (Cretaceous) Terrestrial Vertebrate Faunas. Master’s Thesis, Southern Methodist University, Dallas, TX, USA, 2010; pp. 1–135. [Google Scholar]
- Estes, R.; Berberian, P.; Meszoely, C.A.M. Lower vertebrates from the Late Cretaceous Hell Creek Formation, McCone County, Montana. Breviora 1969, 337, 1–33. [Google Scholar]
- Sullivan, R.M. Fossil lizards from Swain Quarry “Fort Union Formation,” middle Paleocene (Torrejonian), Carbon County, Wyoming. J. Paleont. 1982, 56, 996–1010. [Google Scholar]
- McGrew, P.O.; Berman, J.E.; Hecht, M.K.; Hummel, J.M.; Simpson, G.G.; Wood, A.E. The geology and paleontology of the Elk Mountain and Tabernacle Butte area, Wyoming. Bull. Am. Nat. Hist. 1959, 117, 117–176. [Google Scholar]
- McKenna, M.C. Fossil Mammalia from the early Wasatchian Four Mile fauna, Eocene of northwest Colorado. Univ. Calif. Pub. Geol. Sci. 1960, 37, 1–130. [Google Scholar]
- Smith, K.T.; Gauthier, J.A. Early Eocene lizards of the Wasatch Formation near Bitter Creek, Wyoming: Diversity and paleoenvironment during an interval of global warming. Bull. Peabody Mus. Nat. Hist. 2013, 54, 135–230. [Google Scholar] [CrossRef]
- Sternberg, C.M. The lizard Chamops from the Wapiti Formation of northern Alberta: Polyodontosaurus grandis not a lizard. Bull. Nat. Mus. Can. 1951, 123, 256–258. [Google Scholar]
- Nydam, R.L.; Caldwell, M.W.; Fanti, F. Borioteiioidean lizard skulls from Kleskun Hill (Wapiti Formation; Upper Campanian), west-central Alberta, Canada. J. Vertebr. Paleontol. 2010, 30, 1090–1099. [Google Scholar] [CrossRef]
- Nydam, R.L. Lizards and snakes from the Cenomanian through Campanian of southern Utah: Filling the gap in the fossil record of Squamata from the Late Cretaceous of the Western Interior of North America. In At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah; Titus, A.L., Loewen, M.A., Eds.; Indiana University Press: Bloomington, IN, USA, 2013; pp. 370–423. ISBN 978-0253008831. [Google Scholar]
- Currie, P.J.; Langston, W., Jr.; Tanke, D.H. A New Horned Dinosaur from an Upper Cretaceous Bone Bed in Alberta; NRC Research Press: Ottawa, ON, Canada, 2008; pp. 1–144. ISBN 978-0-660-19820-0. [Google Scholar]
- Fanti, F.; Bell, P.R.; Vavrek, M.; Larson, D.; Koppelhus, E.; Sissons, R.L.; Langone, A.; Campione, N.E.; Sullivan, C. Filling the Bearpaw gap: Evidence for palaeoenvironment-driven taxon distribution in a diverse, non-marine ecosystem from the late Campanian of west-Central Alberta, Canada. Palaeogeogr. Palaeocl. Palaeoecol. 2022, 592, 1–18. [Google Scholar] [CrossRef]
- Fanti, F.; Catuneanu, O. Fluvial sequence stratigraphy: The Wapiti Formation, west-central Alberta, Canada. J. Sediment. Res. 2010, 80, 320–338. [Google Scholar] [CrossRef]
- Oppel, M. Die Ordnungen, Familien und Gattungen der Reptilien als Prodrom Einer Naturgeschichte Derselben; Joseph Lindauer Verlag: Munich, Germany, 1811; pp. 1–86. [Google Scholar]
- Fürbringer, M. Zur vergleichenden Anatomie des Brustschulterapparates und der Schultermuskeln. Jan. Zeitschr. 1900, 34, 215–718. [Google Scholar]
- Lee, M.S.Y. The phylogeny of varanoid lizards and the affinities of snakes. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1997, 352, 53–91. [Google Scholar] [CrossRef]
- Evans, S.E. The skull of lizards and tuatara. In Biology of the Reptilia; Gans, C., Gaunt, A.S., Adler, K., Eds.; Society for the Study of Amphibians and Reptiles: Ithaca, NY, USA, 2008; Volume 20, pp. 1–347. ISBN 978-0-91698-476-2. [Google Scholar]
- McDowell, S.B.; Bogert, C.M. The systematic position of Lanthanotus and the affinities of the anguinomorphan lizards. B. Am. Mus. Nat. Hist. 1954, 105, 1–142. [Google Scholar]
- Mo, J.-y.; Xu, X.; Evans, S.E. A large predatory lizard (Platynota, Squamata) from the Late Cretaceous of South China. J. Syst. Palaeont. 2012, 10, 333–339. [Google Scholar] [CrossRef]
- Yatkola, D.A. Fossil Heloderma (Reptilia, Helodermatidae). Occas. Pap. Mus. Nat. Hist. Univ. Kans. 1976, 51, 1–14. [Google Scholar]
- Maisano, J.A. Terminal fusions of skeletal elements as indicators of maturity in squamates. J. Vertebr. Paleontol. 2002, 22, 268–275. [Google Scholar] [CrossRef]
- Estes, R.; de Queiroz, K.; Gauthier, J.A. Phylogenetic relationships within Squamata. In Phylogenetic Relationships of the Lizard Families; Estes, R., Pregill, G., Eds.; Stanford University Press: Stanford, CA, USA, 1988; pp. 119–291. ISBN 9780804714358, 0804714355. [Google Scholar]
- Richter, A. Lacertilier aus der Unteren Kreide von Uña und Galve (Spanien) und Anoual (Marokko). Berl. Geowiss. Abh. 1994, 14, 1–147. [Google Scholar]
- Vickaryous, M.K.; Sire, J.Y. The integumentary skeleton of tetrapods: Origin, evolution, and development. J. Anat. 2009, 214, 441–464. [Google Scholar] [CrossRef] [PubMed]
- Bever, G.S.; Bell, C.J.; Maisano, J.A. The ossified braincase and cephalic osteoderms of Shinisaurus crocodilurus (Squamata, Shinisauridae). Palaeont. Electr. 2005, 8, 4A. [Google Scholar]
- Bhullar, B.-A.S. Cranial osteology of Exostinus serratus (Squamata: Anguimorpha), fossil sister taxon to the enigmatic clade Xenosaurus. Zool. J. Linn. Soc. 2010, 159, 921–953. [Google Scholar] [CrossRef]
- Bhullar, B.-A.S. The power and utility of morphological characters in systematics: A fully resolved phylogeny of Xenosaurus and its fossil relatives (Squamata: Anguimorpha). Bull. Mus. Comp. Zool. 2011, 160, 65–181. [Google Scholar] [CrossRef]
- Dong, L.; Xu, X.; Wang, Y.; Evans, S.E. The lizard genera Bainguis and Parmeosaurus from the Upper Cretaceous of China and Mongolia. Cret. Res. 2018, 85, 95–108. [Google Scholar] [CrossRef]
- Camp, C.L. Classification of the lizards. Bull. Am. Mus. Nat. Hist. 1923, 48, 289–481. [Google Scholar]
- Mead, J.I.; Arroyo-Cabrales, J.; Johnson, E. Pleistocene lizards (Reptilia: Squamata) from San Josecito Cave, Nuevo León, Mexico. Copeia 1999, 1999, 163–173. [Google Scholar] [CrossRef]
- Thorn, K.M.; Fusco, D.A.; Hutchinson, M.N.; Gardner, M.G.; Clayton, J.L.; Prideaux, G.J.; Lee, M.S.Y. A giant armoured skink from Australia expands lizard morphospace and the scope of the Pleistocene extinctions. Proc. R. Soc. B 2023, 290, 20230704. [Google Scholar] [CrossRef]
- Bauer, A.M.; Russell, A.P. Supraorbital ossifications in geckos. Can. J. Zool. 1989, 67, 678–684. [Google Scholar] [CrossRef]
- de Buffrénil, V.; Sire, J.Y.; Rage, J.C. The histological structure of glyptosaurine osteoderms (Squamata: Anguidae), and the problem of osteoderm development in squamates. J. Morphol. 2010, 271, 729–737. [Google Scholar] [CrossRef]
- Paluh, D.J.; Griffing, A.H.; Bauer, A.M. Sheddable armour: Identification of osteoderms in the integument of Geckolepis maculata (Gekkota). Afr. J. Herpet. 2017, 66, 12–24. [Google Scholar] [CrossRef]
- Scarpetta, S.G. Peltosaurus granulosus (Squamata, Anguidae) from the middle Oligocene of Sharps Corner, South Dakota, and the youngest known chronostratigraphic occurrence of Glyptosaurinae. J. Vertebr. Paleont. 2019, 39, e1622129. [Google Scholar] [CrossRef]
- Maisano, J.A.; Bell, C.J.; Gauthier, J.A.; Rowe, T. The osteoderms and palpebral in Lanthanotus borneensis (Squamata: Anguimorpha). J. Herpetol. 2002, 36, 678–682. [Google Scholar] [CrossRef]
- Erickson, G.M.; de Ricqles, A.; de Buffrénil, V.; Molnar, R.E.; Bayless, M.K. Vermiform bones and the evolution of gigantism in Megalania–how a reptilian fox became a lion. J. Vertebr. Paleont. 2003, 23, 966–970. [Google Scholar] [CrossRef]
- Maisano, J.A.; Laduc, T.J.; Bell, C.J.; Barber, D. The cephalic osteoderms of Varanus komodoensis as revealed by high-resolution X-ray computed tomography. Anat. Rec. 2019, 147, 1–6. [Google Scholar] [CrossRef]
- Rieppel, O. The Phylogeny of Anguimorph Lizards; Birkhäuser Verlag: Basel, Switzerland, 1980; pp. 1–86. ISBN 978-3-7643-1224-4. [Google Scholar]
- Boulenger, G.A. Notes on the osteology of Heloderma horridum and H. suspectum, with remarks on the systematic position of the Helodermatidae and on the vertebrae of the Lacertilia. J. Zool. 1891, 59, 109–118. [Google Scholar] [CrossRef]
- Hoffstetter, R. Un saurien hélodematidé (Eurheloderma gallicum nov. gen. et sp.) dans la faune fossile des phosphorites du Quercy. Bull. Soc. Géo. Fr. 1957, 7, 775–786. [Google Scholar] [CrossRef]
- Stevens, M.S. Further study of Castolon local fauna (early Miocene) Big Bend National Park, Texas. Pearche-Sellards Ser. Tex. Mem. Mus. 1977, 28, 1–69. [Google Scholar]
- Moreno, K.; Wroe, S.; Clausen, P.; McHenry, C.; D’Amore, D.C.; Rayfield, E.J.; Cunningham, E. Cranial performance in the Komodo dragon (Varanus komodoensis) as revealed by high-resolution 3-D finite element analysis. J. Anat. 2008, 212, 736–746. [Google Scholar] [CrossRef]
- Collar, D.C.; Schulte, J.A., II; Losos, J.B. Evolution of extreme body size disparity in monitor lizards (Varanus). Evolution 2011, 65, 2664–2680. [Google Scholar] [CrossRef]
- Brinkman, D. Structural correlates of tarsal and metatarsal functioning in Iguana (Lacertilia; Iguanidae) and other lizards. Can. J. Zool. 1980, 58, 277–289. [Google Scholar] [CrossRef]
- Evans, S.E. At the feet of the dinosaurs: The early history and radiation of lizards. Biol. Rev. 2003, 78, 513–551. [Google Scholar] [CrossRef]
- Evans, S.E.; Jones, M.E.H. The origin, early history and diversification of lepidosauromorph reptiles. In New Aspects of Mesozoic Biodiversity; Bandyopadhyay, Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 27–44. ISBN 978-3-642-10310-0. [Google Scholar]
- International Commission on Zoological Nomenclature (ICZN). International Code of Zoological Nomenclature, 4th ed.; International Trust for Zoological Nomenclature: London, UK, 1999; pp. 1–306. [Google Scholar]
- van Hinsbergen, D.J.J.; de Groot, L.V.; van Schaik, S.J.; Spakman, W.; Bijl, P.K.; Sluijs, A.; Langereis, C.G.; Brinkhuis, H. A paleolatitude calculator for paleoclimate studies. PLoS ONE 2015, 10, e0126946. [Google Scholar] [CrossRef]
- Beck, D.D.; Lowe, C.H. Ecology of the beaded lizard, Heloderma horridum, in a tropical dry forest in Jalisco, Mexico. J. Herpet. 1991, 25, 395–406. [Google Scholar] [CrossRef]
- Jerzykiewicz, T.; Currie, P.J.; Fanti, F.; Lefeld, J. Lithobiotopes of the Nemegt Gobi Basin. Can. J. Earth Sci. 2021, 58, 829–851. [Google Scholar] [CrossRef]
- Eagle, R.A.; Enriquez, M.; Grellet-Tinner, G.; Pérez-Huerta, A.; Hu, D.; Tütken, T.; Montanari, S.; Loyd, S.J.; Ramirez, P.; Tripati, A.K.; et al. Isotopic ordering in eggshells reflects body temperatures and suggests differing thermophysiology in two Cretaceous dinosaurs. Nat. Commun. 2015, 6, 8296. [Google Scholar] [CrossRef]
- Naugolnykh, S.V.; Kuleshov, V.N. New palaeoclimatic insights on Late Cretaceous environments of Mongolia based on the isotope data (δ13C, δ18O) of dinosaur eggshells and pedogenic carbonates from Bayn Dzak section. Glob. Geol. 2020, 23, 199–213. [Google Scholar] [CrossRef]
- Cockx, P.; McKellar, R.; Tappert, R.; Vavrek, M.; Muehlenbachs, K. Bonebed amber as a new source of paleontological data: The case of the Pipestone Creek deposit (Upper Cretaceous), Alberta, Canada. Gond. Res. 2020, 81, 378–389. [Google Scholar] [CrossRef]
- Quinney, A.; Therrien, F.; Zelenitsky, D.; Eberth, D.A. Palaeoenvironmental and palaeoclimatic reconstruction of the Upper Cretaceous (late Campanian-early Maastrichtian) Horseshoe Canyon Formation, Alberta, Canada. Palaeogeogr. Palaeocl. Palaeoecol. 2013, 371, 26–44. [Google Scholar] [CrossRef]
- Eberth, D.A.; Kamo, S.L. High-precision U-Pb CA-ID-TIMS dating and chronostratigraphy of the dinosaur-rich Horseshoe Canyon Formation (Upper Cretaceous, Campanian-Maastrichtian), Red Deer River valley, Alberta, Canada. Can. J. Earth Sci. 2020, 57, 1220–1237. [Google Scholar] [CrossRef]
- Amiot, R.; Lécuyer, C.; Buffetaut, E.; Fluteau, F.; Legendre, S.; Martineau, F. Latitudinal temperature gradient during the Cretaceous Upper Campanian-Middle Maastrichtian: δ18O record of continental vertebrates. Earth Plan. Sci. Let. 2004, 226, 255–272. [Google Scholar] [CrossRef]
- Druckenmiller, P.S.; Erickson, G.M.; Brinkman, D.; Brown, C.M.; Eberle, J.J. Nesting at extreme polar latitudes by non-avian dinosaurs. Curr. Biol. 2021, 31, 1–10. [Google Scholar] [CrossRef]
- Salazar-Jaramillo, S.; McCarthy, P.J.; Ochoa, A.; Fowell, S.J.; Longstaffe, F.J. Paleoclimate reconstruction of the Prince Creek Formation, Arctic Alaska, during Maastrichtian global warming. Palaeogeogr. Palaeocl. Palaeoecol. 2019, 532, 109265. [Google Scholar] [CrossRef]
- Burgener, L.; Hyland, E.; Huntington, K.W.; Kelson, J.R.; Sewall, J.O. Revisiting the equable climate problem during the Late Cretaceous greenhouse using paleosol carbonate clumped isotope temperatures from the Campanian of the Western Interior Basin, USA. Palaeogeogr. Palaeocl. Palaeoecol. 2019, 516, 244–267. [Google Scholar] [CrossRef]
- Falcon-Lang, H.J. Growth interruptions in silicified conifer woods from the Upper Cretaceous Two Medicine Formation, Montana, USA: Implications for palaeoclimate and dinosaur palaeoecology. Palaeogeogr. Palaeocl. Palaeoecol. 2003, 199, 299–314. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamilton, S.M.; Paparella, I.; Bell, P.R.; Campione, N.E.; Fanti, F.; Larson, D.W.; Sissons, R.L.; Vavrek, M.J.; Balsai, M.J.; Sullivan, C. New Lizard Specimens from the Campanian Wapiti Formation of Alberta, Canada. Geosciences 2023, 13, 337. https://doi.org/10.3390/geosciences13110337
Hamilton SM, Paparella I, Bell PR, Campione NE, Fanti F, Larson DW, Sissons RL, Vavrek MJ, Balsai MJ, Sullivan C. New Lizard Specimens from the Campanian Wapiti Formation of Alberta, Canada. Geosciences. 2023; 13(11):337. https://doi.org/10.3390/geosciences13110337
Chicago/Turabian StyleHamilton, Samantha M., Ilaria Paparella, Phil R. Bell, Nicolás E. Campione, Federico Fanti, Derek W. Larson, Robin L. Sissons, Matthew J. Vavrek, Michael J. Balsai, and Corwin Sullivan. 2023. "New Lizard Specimens from the Campanian Wapiti Formation of Alberta, Canada" Geosciences 13, no. 11: 337. https://doi.org/10.3390/geosciences13110337
APA StyleHamilton, S. M., Paparella, I., Bell, P. R., Campione, N. E., Fanti, F., Larson, D. W., Sissons, R. L., Vavrek, M. J., Balsai, M. J., & Sullivan, C. (2023). New Lizard Specimens from the Campanian Wapiti Formation of Alberta, Canada. Geosciences, 13(11), 337. https://doi.org/10.3390/geosciences13110337