Abstract
Three species belonging to the fern genus Ochotopteris E. Lebedev, including the new species O. lebedevii Herman et Domogatskaya, are revised, described and illustrated in this paper. The fossils come from Albian, Cenomanian, Turonian and Coniacian beds exposed at seven sites in North-Eastern Asia and Northern Alaska. These sites are located in the Cretaceous Arctic, with their palaeolatitudes ranging from 60° N to 80° N. The high endemism characteristic of the mid-Cretaceous Arctic flora is most probably due to adaptations in the globally distinctive Arctic conditions: the unique combination of temperatures, precipitation and the highly seasonal polar light regime. Being an endemic plant of the mid-Cretaceous Arctic, Ochotopteris ferns were adapted to a regional wet temperate to a wet warm temperate palaeoclimate accompanied by marked sunlight seasonality with a prolonged—up to several weeks—winter darkness and continuous summer daylight, also lasting for several weeks. The majority of the mid-Cretaceous Arctic plants were deciduous. Their predominant deciduousness was most probably due to the polar light seasonality: at the end of the growing season, some plants dropped their leaves or leafy shoots, whereas others—including Ochotopteris ferns—died back to rhizome systems.
Keywords:
fern; Arctic; North-Eastern Asia; Northern Alaska; Albian; Cenomanian; Turonian; Coniacian; endemism; palaeoclimate 1. Introduction
Although the study of Albian and Late Cretaceous plants, biostratigraphy and palaeoclimates of the Arctic began in the second half of the 19th century, the most intensive research took place in the second half of the 20th and the beginning of the 21st centuries. Of particular scientific interest is the existence of rich ancient floras that inhabited the high latitude areas of the Arctic, up to palaeolatitude of 85° N [1], during the global “hothouse” climate of the Cretaceous [2,3,4]. The Cretaceous polar vegetation possessed a relatively large number of endemic plants, mainly among ferns such as Arctopteris [5,6], Birisia [7,8], Chaunia [9], Kolymella [9], Ochotopteris [10] and Penzhinopteris [11], and examples of endemic angiosperms include Anadyricarpa [12], Arthollia [13], Barykovia [14], Caricopsis [15], Chachlovia [16], Crataegites [15], Dalembia [17], Disanthophyllum [18], Grebenkia [19], Kenella [20], Palaeotrapa [21], Paraprotophyllum [22], Pseudoprotophyllum [23], Penzhinia [24], Platimelis [25], Ranunculaecarpus [15], Rarytkinia [18], Soninia [16], Sorbites [26], Terechovia [27] and Scheffleraephyllum [28].
In this work, we examine, describe and illustrate three species of the endemic Arctic fern Ochotopteris, including one new species of this genus: O. lebedevii. Two Ochotopteris species, including the type species of the genus, had been published previously in Russian and were rather poorly illustrated with a few leaf drawings and black-and-white photographs. Hence, the information on these ferns was not easily accessible to the general palaeobotanical community. The main point of this paper is to revise this genus, paying particular attention to the geographic and stratigraphic position of the Ochotopteris species. The revision and description of these endemic ferns are followed by our estimation of their palaeolatitudinal positioning and an analysis of the Cretaceous Arctic palaeoclimate that was most likely “responsible” for the high endemism of the high-latitude mid-Cretaceous floras of North-Eastern Asia and Northern Alaska. For the palaeoclimate reconstruction, we use CLAMP, which gives quantitative estimates of past climate parameters.
2. Material and Methods
Our research is based on the study of six plant fossil collections from seven Cretaceous sites, Albian to Coniacian, in North-Eastern Asia and Northern Alaska (Figure 1). These collections are permanently stored in the Geological Institute, Russian Academy of Sciences (GIN RAS), Moscow, Russia: collection no. 3823 (Northwest Kamchatka), no. 3825 (Western Priokhot’ye, Tyl’ River basin) and no. 3381 (Ul’ya Trough, Amka River basin); in the Magadan branch of the Territorial Foundation of Geological Information on the Far Eastern Federal Region (FGSM), Magadan, Russia: collection no. FGSM 1017 (Pekulnei Ridge, western slope) and no. FGSM 233 (Anadyr River basin, Bystraya River basin); and in the National Museum of Natural History, Smithsonian Institution (NMNH), Washington DC, USA: collection USNM (“Smiley’s collection”, Chandler River basin, Northern Alaska; [4]).
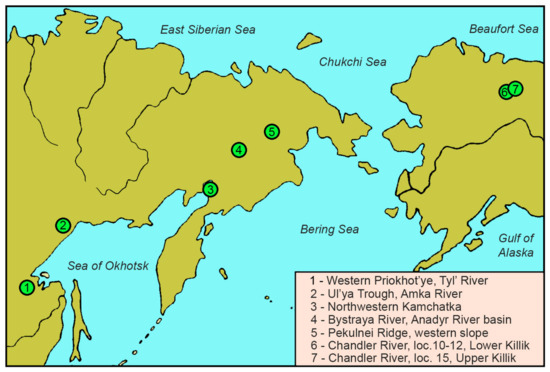
Figure 1.
Ochotopteris plant fossil localities in North-Eastern Asia and Northern Alaska.
The plant fossils studied are represented by impressions of sterile fronds (leaves) and their fragments on siltstones or tuffaceous siltstones. These impressions are often covered by a coaly material or sometimes iron-oxide infilling. Their preservation is moderately good, although the fine venation of the pinnules is poorly preserved and not always observable. Unfortunately, no cuticles were preserved.
Our identifications are based on direct observations made using a binocular eye loupe and a dissecting stereomicroscope Carl Zeiss SM-XX. To highlight the venation of fossil leaves, we used oblique lighting. After cleaning, the specimens were photographed under low-angle incident light from a portable battery-powered LED light source. Images of the plant fossils were taken with a Panasonic Lumix GX9 digital camera equipped with a Leica DG Macro-Elmarit 1:2.8/45 mm macro lens. To enhance contrast, some specimens were examined using two polarizing filters: one in front of the camera or microscope lens and another one in front of the light source. We used Adobe Photoshop CC software to prepare the illustrations.
We calculated the palaeolatitudes of the plant fossil sites (Table 1) using a Palaeolatitude Calculator [29,30]. The palaeoreconstruction of continental configuration was generated using an ODSN Plate Tectonic Reconstruction Service [1,31]. To obtain the quantitative characteristics of the Cretaceous Arctic palaeoclimate, we used the CLAMP method [2,3].

Table 1.
Estimated palaeolatitude of the Ochotopteris localities.
3. Stratigraphic Position and Age of the Plant Fossil Localities
In total, seven localities bear Ochotopteris specimens (Figure 1): (1) Tyl’ River (Western Priokhot’ye Region), Tyl’ Formation, middle–late Albian; (2) Amka River (Ul’ya Trough Region), Yemanra Formation, middle–late Albian (or, possibly, younger Cenomanian–Turonian); (3) Konglomeratovyi Cape (Northwestern Kamchatka), Valizhgen Formation, most probably early Coniacian; (4) Bystraya River (Anadyr River basin, Chukotka), Upper Subformation of the Krivorechenskaya Formation, Cenomanian; (5) western slope of the Pekul’nei Ridge (Chukotka), Tylpegyrgynai Formation, early Coniacian; (6) localities 10–12, Chandler River (Northern Alaska), lower part of the Killik Tongue (Lower Killik), middle–late Albian and (7) locality 15, Chandler River (Northern Alaska), upper part of the Killik Tongue (Upper Killik), Cenomanian.
3.1. Western Priokhot’ye, Tyl’ River
The type species of the genus Ochotopteris, O. ochotensis E. Lebedev, comes from locality 19 (approx. 54°08′ N, 135°02′ E) of the Tyl’ Formation in the Tyl’ River upper stream, Torom Trough, Western Priokhot’ye Region (Figure 1, site 1; [10]). This formation, 400–450 m thick, consists of two parts, with the lower part comprising mostly conglomerates and the upper part consisting of sandstones, siltstones and claystones with numerous plant remains. There are no marine fossils in these deposits, which are of continental origin. Lebedev (1974) who collected O. ochotensis impressions suggested a middle–late Albian age for the plant-bearing deposits of the Tyl’ Formation because the floral assemblage is taxonomically similar to that of the Albian Khatyryk, Buor-Kemuss and Omsukchan floras of North-Eastern Asia [10,32,33], which is dated on the basis of marine mollusc fossils found in deposits regionally underlying the plant-bearing beds. Unfortunately, there are no independent (non-palaeobotanical) dates for the Tyl’ Formation.
3.2. Ul’ya Trough, Amka River
Numerous plant fossils were collected by Lebedev [34] on the right and left banks of the Khetana River, Amka River right tributary, Ul’ya Trough (Figure 1, site 2; Figure 2). According to him, the fossils from localities 101, 107 and 110 (approx. 59°24′ N, 140°23′ E) compose the Yemanra floral assemblage from the volcanogenic and volcanogenic–terrigenous deposits of continental origin belonging to the Yemanra Formation. The majority of plant fossils, including all impressions of Ochotopteris lebedevii sp. nov. described below, were collected from locality 101 (Figure 2). Lebedev [34] pointed out that the Yemanra assemblage is not identical to any known Cretaceous floras of North-Eastern Asia and Northern Alaska but is most similar to those of middle and late Albian age. On this basis, a mid-Albian age is assumed for the Yemanra assemblage in this work. Unfortunately, there are no marine fossils in the Cretaceous deposits of this region, which belong to the Okhotsk–Chukotka volcanogenic belt of North-Eastern Asia.
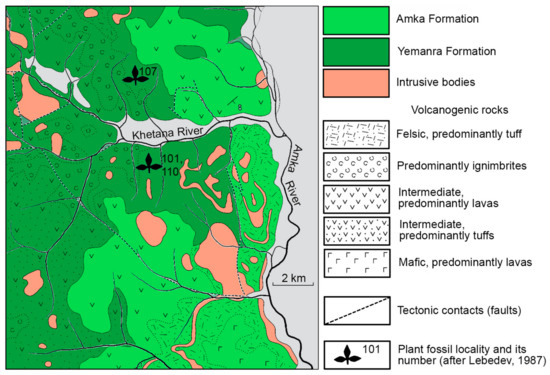
Figure 2.
Geological scheme and plant fossil localities in the Ul’ya Trough, Amka River; modified from [35]; locality numbers after [34].
Recently, Lebedev’s collection of the Yemanra plant fossils was re-examined by Shczepetov and Herman [35]. The following taxa were identified: Thallites sp., Osmunda sp., Ochotopteris sp. nov., Birisia (?) sp., Coniopteris sp., Arctopteris sp., Arctopteris (?) sp., Onychiopsis sp., Cladophlebis sp. 1, Cladophlebis sp. (possibly Birisia sp.), Sphenopteris sp., Polypodiopsida gen. et sp. indet., Heilungia sp., Taeniopteris sp., Ginkgo ex gr. adiantoides (Unger) Heer, G. ex gr. sibirica Heer, Sphenobaiera biloba Prynada, Sphenobaiera sp., Podozamites ex gr. lanceolatus (Lindley et Hutton) F. Braun, Pagiophyllum ex gr. triangulare Prynada, Pagiophyllum (?) sp. (shoot with cones), Elatocladus gyrbykensis Golovneva, E. ex gr. smittiana (Heer) Seward, Cephalotaxopsis ex gr. intermedia Hollick, Florinia (?) sp., Pityophyllum ex gr. staratschinii (Heer) Nathorst, P. ex gr. nordenskioldii (Heer) Nathorst, Cupressinocladus sp., Araucarites sp., Sequoia ochotica Yudova et Golovneva, Sequoia sp. (cone), Sequoia (?) sp., Pityolepis sp. 1–3, Pityospermum sp. 1–4, Pityospermum (?) sp., Pityostrobus sp., cf. Juniperus sp., Desmiophyllum (Podozamites ?) sp. and Dicotylophyllum sp. 1–3. Unfortunately, the age of the plant-bearing deposits is uncertain to some extent: there is no independent (non-palaeobotanical) dating of these deposits, but the taxonomic composition of the Yemanra assemblage does not allow the precise age determination of the plant-bearing beds. We believe that the assemblage is definitely mid-Cretaceous, most probably middle–late Albian and/or Cenomanian–Turonian in age since it resembles some fossil floras dated using marine mollusc finds in deposits regionally underlying the plant-bearing beds.
3.3. Northwestern Kamchatka
The Upper Cretaceous section of Northwestern Kamchatka (Figure 1, site 3) starts with the Mameta Formation overlying the eroded Lower Cretaceous Karmalivayam Formation. The lowermost beds of the Mameta Formation are supposed to be late Albian in age, whereas overlying beds contain the remains of Cenomanian molluscs and correspond to the regional inoceramid zones I. pennatulus and I. nipponicus—I. scalprum at the top of the Cenomanian section of Northwest Kamchatka (Figure 3B). Thus, the age of the Mameta Formation is determined as late Albian—Cenomanian or, possibly, earliest Turonian [36,37].
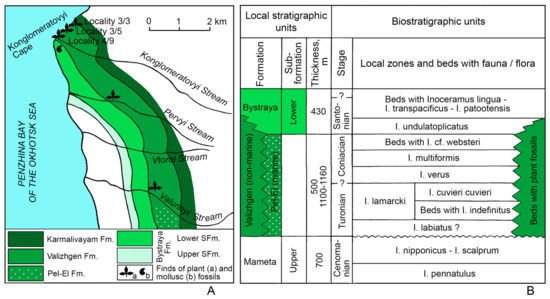
Figure 3.
Geological scheme and plant and marine mollusc fossils localities in the Konglomeratovyi Cape vicinity, Northwest Kamchatka (A) and the Upper Cretaceous local stratigraphy and age of the plant-bearing beds of the Valizhgen Formation (B); modified from [36,37,38]; locality numbers after [36].
To the south of the Karmalivayam River, the Mameta Formation is overlain, possibly unconformably, by the marine deposits of the Pel-El Formation. The latter overlies the eroded Lower Cretaceous Karmalivayam Formation to the north from the Karmalivayam River and in the Valunnyi Stream basin (Figure 3A). The lower part of the Pel-El Formation is not palaeontologically characterised but is placed between the uppermost Cenomanian zone I. nipponicus—I. scalprum and the Turonian zone I. lamarcki. Pergament [37] assumed that these strata corresponded to the lowermost Turonian Inoceramus labiatus zone. The overlying layers of the Pel-El Formation exhibit numerous Turonian and Coniacian inoceramids and ammonites. Therefore, the entire Pel-El Formation is considered to be of Turonian (possibly, excluding lowermost Turonian)—Coniacian age (Figure 3B).
The Valizhgen Formation overlies, disconformably, Aptian—Albian and even older rocks of the Karmalivayam Formation (Figure 3A). The Valizhgen Formation consists of terrigenous and coal-bearing deposits of continental origin featuring plant fossils. According to Pergament’s observations, the continental deposits of the Valizhgen Formation are laterally replaced by the marine rocks of the Pel-El Formation north of latitude 62°N (Figure 3A), with the basal conglomerates of the Pel-El Formation being laterally continuous with the lowermost conglomerates of the Valizhgen Formation [4,11,36].
The Pel-El and Valizhgen formations are conformably overlain by the Bystraya (=Bystrinskaya) Formation (Figure 3). The Lower Bystraya Subformation contains in its lower units the remains of Inoceramus undulatoplicatus Roemer and I. naumanni Yok.; higher up, it contains I. lingua lingua Goldf., I. patootensis sokolovi Perg., I. transpacificus Peg., etc., arguing for a Santonian and, possibly, early Campanian age for these deposits [36,37].
Thus, the plant-bearing deposits of the Valizhgen Formation are considered to be of Turonian (possibly, excluding lowermost Turonian)—Coniacian age (Figure 3). The holotype of Ochotopteris kamtchatica E. Lebedev and the second leaf impression of this species were collected from locality 3/5 on the Konglomeratovyi Cape in the middle part of the Valizhgen Formation (62°06′ N, 164°05′ E), with the age of this site being most probably early Coniacian [11,38].
3.4. Anadyr River Basin, Bystraya River
The latest Albian—earliest Turonian Grebenka flora (Krivorechenskaya Formation, Upper Subformation, Anadyr River middle reaches in Chukotka) has been regarded as “the most important Late Cretaceous palaeofloristic record” [39], p. 100, in North-Eastern Asia. A single leaf impression of Ochotopteris kamtchatica E. Lebedev was reported from the Krivorechenskaya Formation [4]. Here, on the right bank of the Anadyr River, the plant-bearing deposits of this formation extend through the basins of the Orlovka, Grebenka, Gornaya and Bystraya rivers [40,41] with the impression of O. kamtchatica being found at locality 233 (approx. 64°41′ N, 169°17′ E) on the left bank of the Bystraya River (Figure 1, site 4; Figure 4). The Krivorechenskaya Formation overlies Valanginian and Hauterivian strata with an angular discordance and is subdivided into two subformations [42]. The plant-bearing deposits belong to the Upper Krivorechenskaya Subformation and are 700–950 m thick. This subformation is represented by both continental rocks (sandstones and conglomerates) and marine beds (mostly sandstones and siltstones) yielding marine faunal remains. The marine fossils provide the age determination for these deposits [39,40].
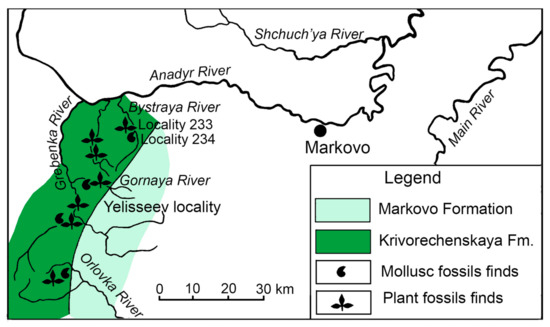
Figure 4.
Distribution of plant-bearing deposits of the Krivorechenskaya Formation and overlying Markovo Formation on the right bank of the Anadyr River middle reaches; modified from [38,41]; numbers of the plant and marine mollusc fossil localities after [39].
On the left bank of the Bystraya River, poorly preserved marine fossils were found in siltstones at locality 234, which overlies the plant-bearing layers of locality 233 (Figure 4). According to Terekhova [39], these marine fossils represent Inoceramus ex gr. nipponicus Nagao et Mat. and Inoceramus sp. indet., most probably of Cenomanian age. Another locality of the Upper Krivorechenskaya Subformation that contains rare plant impressions and numerous marine mollusc remains is situated in the middle reaches of the Gornaya River (Figure 4). The molluscs indicate that the locality belongs to the Mantelliceras zones of the standard scale [43], to the Hypoturrilites gravesianus ammonite zone and to the Inoceramus dunveganensis aiensis inoceramid zone, and it is therefore of early Cenomanian age [40,41]. The plant-bearing deposits of the Yelisseev Locality (64°28′ N, 168°58′ E) (Figure 4), which is very rich in plant fossils but contains no mollusc remains, are situated stratigraphically just below the Gornaya River Locality succession, and thus, are probably not younger than early Cenomanian. Radiometric 40Ar/39Ar dating of tuffaceous sandstones within the Yelisseev Locality confirms the lower Cenomanian age (96.5 ± 0.5 Ma) of the Yelisseev Locality deposits [41]. Hence, the most probable age of the Ochotopteris kamtchatica sample from locality 233 on the left bank of the Bystraya River is Cenomanian.
3.5. Pekulnei Ridge
Filippova [44] described several Ochotopteris frond impressions from the Tylpegyrgynai Formation of the Pekulnei Ridge western slope (Figure 1, site 5). Only two leaf impressions were illustrated [44,45], but three other specimens belonging to Ochotopteris were also mentioned by Filippova. All of them came from the Tylpegyrgynai Formation (Figure 5), but the exact locations of these samples are sadly not specified in Filippova’s publications.
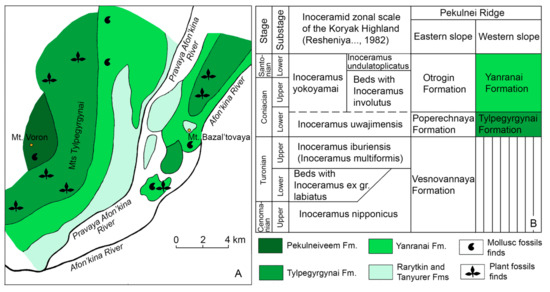
Figure 5.
Geological scheme and plant and marine mollusc fossil localities on the western slope of the Pekulnei Ridge (A) and inoceramid biostratigraphy and age of the plant-bearing beds of the Tylpegyrgynai Formation (western slope) and Poperechnaya Formation (eastern slope) (B); modified from [38,45,46].
The early Coniacian age of the Tylpegyrgynai Formation flora is based on the biostratigraphic correlation of the plant-bearing deposits with underlying and overlying marine units [4,38,44,45,46,47,48]. This formation, 1000–1100 m thick, is exposed in the Tylpegyrgynai Mountains and the Afon’kina River basin (Figure 5). There is an unconformable contact between the Tylpegyrgynai Formation and the underlying marine Pekulneiveem Formation, which is approximately Tithonian to Valanginian in age. The marine Yanranai Formation, 700 to 800 m thick, conformably overlies the plant-bearing unit. Numerous marine molluscs found in this formation have been correlated with the Inoceramus yokoyamai zone, suggesting a late Coniacian to early Santonian age, therefore defining the upper age limit of the Tylpegyrgynai Formation as early Coniacian. The lower age limit of the Tylpegyrgynai Formation cannot be defined precisely due to the unconformable contact with underlying Lower Cretaceous deposits (Figure 5).
The fossil flora from the Tylpegyrgynai Formation is almost identical to that comprising the plant assemblages from the Poperechnaya Formation exposed on the Pekulnei Ridge eastern slope [38,45]. Here, the Vesnovannaya Formation conformably underlies the plant-bearing unit. Fossil molluscs, mainly inoceramids and ammonites, found in this formation allow it to be ascribed to the Inoceramus nipponicus zone (Cenomanian or Cenomanian and lower Turonian) and the Inoceramus iburiensis zone (upper Turonian), thus indicating that the age of the Vesnovannaya Formation is Cenomanian to Turonian (Figure 5B). The Poperechnaya Formation is conformably overlain by marine deposits of the Otrogin Formation which, judging by the fossil molluscs found in it, is late Coniacian to early Santonian in age (Figure 5B). Hence, the age of the non-marine plant-bearing Poperechnaya Formation is early Coniacian [38,45]. A similar taxonomic composition of the floras from the Tylpegyrgynai and Poperechnaya formations suggests that the lower age limit of the Tylpegyrgynai Formation is the same as that of the Poperechnaya Formation, i.e., early Coniacian (Figure 5B). Hence, Ochotopteris fossils found in the Tylpegyrgynai Formation of the western slope of the Pekulnei Ridge (approx. 66°25′ N, 174°49′ E) are most probably of early Coniacian age.
3.6. Chandler River, Northern Alaska
In the Chandler River basin (Figure 1, sites 6 and 7), Ochotopteris kamtchatica E. Lebedev impressions were collected from Smiley’s localities 10, 11, 12 and 15 (approx. 68°47′ N, 152°09′ W) (Figure 6A; [4,49,50]). In this region, the Chandler Formation of the Nanushuk Group consists of predominantly non-marine deposits originally mapped as the Killik Tongue and Niakogon Tongue (Figure 6B). In all outcrops of the Chandler River basin, these two units are separated by the marine strata of the Ninuluk Formation [49]. Numerous finds of inoceramids Inoceramus dunveganensis MacLearn in the Ninuluk Formation define the Cenomanian age of this unit [49].
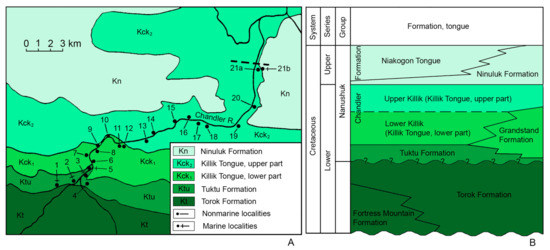
Figure 6.
Geological scheme and plant and marine mollusc fossil localities on the Chandler River middle reaches, Tuktu Bluff, Northern Alaska (A) and local stratigraphy and age of the plant-bearing beds of the Killik Tongue (B); modified from [49,50]; locality numbers after [50].
The predominant fossils of the Killik Tongue are plant remains [4,38,50]. According to [49], in the southern and south-western parts of the Chandler River basin, the Killik Tongue is divisible into two lithologically distinct units named Lower Killik and Upper Killik [50]. The stratigraphically conformable contact of the Killik Tongue with the underlying marine deposits of the middle Albian Tuktu Formation and overlying layers of the Ninuluk Formation yielding the Cenomanian inoceramids, together with a lateral substitution of the plant-bearing deposits by the middle–late Albian marine Grandstand Formation, allow the dating of the Killik Tongue as middle Albian–early Cenomanian. This is corroborated by the findings of the Cenomanian Inoceramus dunveganensis MacLearn in the middle part of the Upper Killik in the Colville River area [50]. Thus, the Lower Killik is middle–late Albian in age, and the Upper Killik is Cenomanian. It follows that the remains of Ochotopteris kamtchatica E. Lebedev are dated from the middle–late Albian (localities 10, 11 and 12; Figure 6) to Cenomanian (locality 15; Figure 6).
4. Systematic Palaeobotany
Class: Polypodiopsida Cronquist, Takht. et W. Zimm.
Order: Polypodiales Link.
Polypodiales incertae sedis.
Genus: Ochotopteris E. Lebedev, 1974.

Figure 7.
Ochotopteris ochotensis E. Lebedev: (a,b)—holotype, collection no. 3825 (GIN RAS), specimen no. 19-5, almost complete leaf; Western Priokhot’ye, Tyl’ River upper reaches, locality 19; Tyl’ Formation; Lower Cretaceous, middle–upper Albian. Scale bar is 1 cm.
Species included Ochotopteris ochotensis E. Lebedev (type species), Ochotopteris kamtchatica E. Lebedev and Ochotopteris lebedevii Herman et Domogatskaya, sp. nov.
Emended diagnosis (modified from [10]): It has a widely ovate sterile bipinnate-pinnatifid frond. The main rachis is straight and stout, bearing nearly oppositely or alternately attached primary pinnae. The basalmost primary pinnae are subopposite. These pinnae and the first basiscopic secondary pinnae, or pinnately parted pinnules, branch out of them asymmetrically: pinnules on the basiscopic side are larger and more divided than the acroscopic ones; the basiscopic side of pinnately parted pinnules is more divided than acroscopic ones, with basiscopic lobes being larger than acroscopic ones. Pinnules vary from ovate to lanceolate, narrowly elliptic or widely to narrowly triangular; unlobed or pinnately lobed to parted or mostly attached by a broad base with a distinctly decurrent proximal margin. Apices are acute or rounded. Margins are entire, undulate or dentate. Pinnules have a midrib and diverging lateral veins. In the majority of pinnules, the midrib enters the pinnule at an acute angle near the basiscopic margin at the base of a pinnule. Lateral veins are thin and forked twice or more.
Comparison and discussion: Although pinnae asymmetry is known in many ferns with sphenopterid foliage, Ochotopteris leaves differ considerably from them in having a predominantly pecopterid shape of the pinnules. The asymmetry of the primary basal pinnae, together with the asymmetry of the first basal basiscopic secondary pinnae, or pinnately parted pinnules, branching out of them (“double asymmetry” of the frond) is a characteristic feature of the genus Ochotopteris, which allows it to be differentiated from other fossil ferns with pecopterid pinnules.
In Ochotopteris, most pinnules are attached to the rachis by their broad bases, thus having the pecopterid shape. However, in O. ochotensis and O. lebedevii sp. nov. basal pinnules on the acroscopic side of the lowest primary pinnae are often attached to the rachis by a constricted (strongly narrowed) base. O. kamtchatica and O. lebedevii sp. nov. basal pinnules on the basiscopic side of the pinnae shift towards, or onto, the main or lowest primary pinna rachis. Isolated fragments of Ochotopteris leaves resemble some ferns possessing pecopterid, or close to pecopterid, pinnules. The majority of the Mesozoic sterile ferns with a predominantly pecopterid shape of the pinnules are usually attributed to the morphogenus Cladophlebis Brongniart, and pecopterid ferns with constricted bases of their pinnules to the morphogenus Raphaelia Debey et Ettingshausen. This means that the correct assignment of fragmentary fern material to Ochotopteris is possible only if the basal part of the fossil frond with basal pinnae is preserved.
Several modern ferns of the family Pteridaceae possess fronds with the “double asymmetry” typical of Ochotopteris. This feature is known in, for example, several Dryopteris and Pteris species. Since no fertile (spore-bearing) structures have been found in Ochotopteris until now, we, following Lebedev [10], consider it as a morphogenus or a genus of a morphological system.
Morphological taxonomy is independent of the “natural”, or “real”, taxonomy in that it deals not with complete plants but with their dispersed parts, like leaves, wood, pollen, spores, etc. It gives us a system of morphological taxa, or parataxa (like Ochotopteris), which are not the same as natural taxa (like Dryopteris). Theoretically, we can claim that some Dryopteris ferns possess sterile leaves belonging to Ochotopteris. However, if we only have a fossil sterile foliage similar to, let us say, Dryopteris, it is risky to attribute this foliage to that taxon within the natural system of ferns. In fact, an assignment of a fossil leaf to a modern genus or family implies (as a rule, unjustifiably) that all other, usually unknown, organs of the fossil plant are the same as in the living representatives of this taxon. However, normally we lack any proof that this is the case. According to an evocative metaphor of Sergei Meyen (pers. comm.), in this case, we act like someone who found a metal nut on the railway and claimed to have found a steam locomotive. Thus, the major difference between generic taxa Dryopteris and Ochotopteris is that for the first one, we know the complete plant, whereas for the second one, we only know the sterile foliage.
Ochotopteris ochotensis E. Lebedev
(Figure 7a,b)
Holotype: specimen no. 19-5, almost complete leaf (Figure 7a,b).
Repository: GIN RAS, Moscow, Russia, collection no. 3825.
Type locality: Western Priokhot’ye Region, Torom Trough, Tyl’ River upper reaches, Ilinurek-Makit Stream, locality 19 (approx. 54°08′ N, 135°02′ E).
Type stratum: Tyl’ Formation; Lower Cretaceous, middle–upper Albian.
Material: holotype only.
Emended diagnosis (modified from [10]): It has a widely ovate sterile frond, once pinnate-pinnatifid in an apical part and bipinnate-pinnatifid in a basal part. The main rachis is robust. The basalmost primary pinnae are asymmetrical: the basal pinnule of the acroscopic side is entirely unlobed while the basal pinnule of the basiscopic side is pinnately parted. This pinnule is also asymmetrical, where the basiscopic side is divided into more lobes than the acroscopic one, with basiscopic lobes being larger than acroscopic ones. The basal basiscopic pinnule of deeply pinnatifid pinnae arising near or from the base of pinna rachis. The first basal acroscopic pinnule of basalmost primary pinnae is attached with a distinctly constricted base; the second one is attached with strongly contracted basiscopic and acroscopic base margins and the third one is attached with a slightly contracted basiscopic margin. Basal basiscopic lobes arise near, or from, the base of the midrib. Lateral veins arise nearly opposite and become catadromous.
Description: The holotype (Figure 7a,b) represents an impression of a sterile, widely ovate, almost completely preserved leaf, once pinnate-pinnatifid in the apical part and bipinnate-pinnatifid in the basal part. The total frond size is about 35 mm long and 45 mm wide, with a robust main rachis. The petiole is not preserved. The second from the base pinna is lance-ovate, slightly curved in shape, up to 18 mm long and 6 mm wide. The basalmost primary pinnae are asymmetrical, about 23 mm long and 15 mm wide, subopposite, emerging at an angle of 40–60°, while the second acroscopic pinnule is inserted at an angle of 100–120°. Towards the apex of the frond, the insertion angle of the primary pinnae gradually diminishes.
The pinnules are ovate, lance-ovate, widely triangular, triangular or narrowly triangular and unlobed or pinnately lobed to parted; most of them are attached by a broad base with a decurrent basal margin. Triangular unlobed pinnules are up to 3 × 2 mm in size, while the size of narrowly triangular unlobed pinnules is up to 8 × 2.5 mm. Apices are acute or rounded. Margins are entire or dentate. The basal acroscopic pinnule, or lobes of primary branches, tend to be parallel with the main rachis. Basal basiscopic pinnules and lobes emerge near or from the base of the pinna rachis or midrib and have a slightly contracted basiscopic margin.
The first basal acroscopic pinnule of the basal primary pinnae has an undulate margin. It is attached to the axis by a distinctly constricted base; the second one is attached with strongly contracted basiscopic and acroscopic base margins and the third one is attached with a slightly contracted basiscopic margin. The basal basiscopic pinnately parted pinnule of the basal primary pinnae is asymmetrically compound: the basiscopic side is more divided than the acroscopic one, with the basiscopic lobes being larger than the acroscopic ones. These pinnules are up to 11 mm long and 5 mm wide, and the basal lobe size is 4.5 × 2 mm.
Venation is pinnate. In the majority of pinnules, the midrib enters the pinnule at an acute angle near the basiscopic margin at the base of the pinnule; towards the frond apex, the midrib becomes narrower and sinuous, branching and disappearing near the apex. In the basal acroscopic pinnule of the lowest primary pinnae, the midrib enters the pinnule in the middle of its base at a right angle. Lateral veins are subtle and therefore often difficult to observe. Narrowly triangular unlobed pinnules have up to eight lateral veins on each side of the midrib. These veins fork up to three times in the middle part of the pinnule. The first basiscopic lateral vein emerges near, or from, the midrib base. Lateral veins emerge nearly opposite and become catadromous.
Comparison: The differences between O. ochotensis and the other two species of the genus are given in the descriptions of them.
Ochotopteris kamtchatica E. Lebedev

Figure 8.
Ochotopteris kamtchatica E. Lebedev: (a,b)—holotype, collection no. 3823 (GIN RAS), specimen no. 3/5–39, almost complete leaf; (c,d)—specimen no. 3/5–23, collection no. 3823 (GIN RAS); Northwestern Kamchatka, Konglomeratovyi Cape, locality 3/5; Valizhgen Formation, lower Coniacian. Scale bar 1 cm.

Figure 9.
Ochotopteris kamtchatica E. Lebedev: (a)—unnumbered specimen, collection FGSM (FGSM, Magadan); Anadyr River basin, Bystraya River, locality 233; Krivorechenskaya Formation, Cenomanian; (b)—specimen no. 19; (c)—specimen no. 26; (d)—specimen no. 23; (e)—specimen no. 21; (b–e)—“Smiley’s collection” (National Museum of Natural History, Washington DC); Northern Alaska, Chandler River, locality 10KcK1; Lower Killik, middle–upper Albian. Scale bar is 1 cm.
Holotype: specimen no. 3/5–39, almost complete leaf (Figure 8a,b).
Repository: GIN RAS, Moscow, Russia, collection no. 3823.
Type locality: Northwestern Kamchatka, Konglomeratovyi Cape, locality 3/5 (62°06′ N, 164°05′ E).
Type stratum: Valizhgen Formation; Upper Cretaceous, lower Coniacian.
Material: samples 3/5–26 and 3/5–39 (collection no. 3823, Northwestern Kamchatka; GIN RAS, Moscow); single unnumbered specimen from locality 233 (collection FGSM, Anadyr River basin, Bystraya River. This specimen comes from the collection stored in the Territorial Foundation of Geological Information in Magadan, Russia, where the majority of geological collections are not properly curated and numbered); specimen 1017/216 (collection FGSM, Pekulnei Ridge, western slope; FGSM, Magadan); 16 samples [4]: “Smiley’s collection”, Chandler River; NMNH, Washington DC.
Occurrence: Northern Alaska: Lower Killik, middle–upper Albian; Upper Killik, Cenomanian; Eastern Chukotka, Anadyr River middle reaches: Krivorechenskaya Formation, Cenomanian; Northwest Kamchatka: Valizhgen Formation, lower Coniacian; Pekul’nei Ridge: Tylpegyrgynai Formation, lower Coniacian.
Emended diagnosis (modified from [11]): It has a widely ovate sterile frond, once pinnate-pinnatifid in the apical part and bipinnate-pinnatifid in basal part. The main rachis is robust. The basalmost primary pinnae are asymmetrical: the basal pinnule of the acroscopic side is unlobed, while the first basiscopic branch is a pinnately parted pinnule, or typically a deeply pinnatifid pinna. These first basiscopic branches of the basalmost primary pinnae are also asymmetrical: the basiscopic side is more divided than the acroscopic one, and the basiscopic pinnules and lobes are larger than the acroscopic ones. Pinnules are attached by a broad base. Basal basiscopic pinnule arises from the base of the primary pinna rachis or is decurrent to the main rachis or the rachis of the basalmost primary pinnae. The basal basiscopic lobe arises from the base of the midrib or is decurrent to the rachis it emerges from. Lateral veins are nearly opposite and become catadromic or alternate in catadromic order.
Description: It has widely ovate sterile leaves, once pinnate-pinnatifid in apical part with pinnately lobed-cleft pinnules (Figure 9e) or typically with deeply pinnatifid pinnae (near bipinnate) and bipinnate-pinnatifid in the basal part with pinnately parted pinnules (Figure 9d,e) or typically with deeply pinnatifid pinnae (near tripinnate); from 41 to 80 mm long and from 37 to 97 mm wide; the main rachis is robust, and the petiole is unknown. The shapes of the pinnae (except for the lowest primary pinnae) are narrowly elliptic, slightly curved. Sizes of the second from the base deeply pinnatifid pinnae are 20 to 32 mm long and 7 to 10 mm wide. The basalmost primary pinnae are from 26 to 53 mm long and from 17 to 40 mm wide, sub-opposite, emerging at an acute angle, near the second acroscopic pinnule located at an angle of almost 90° (Figure 9e), about 90° (Figure 8a–d; Figure 9b–d) or slightly more than 90° (Figure 9a). The angle of the primary branch departure gradually diminishes towards the leaf apex, and the last ones start to arise alternately near the leaf base (Figure 9c), gradually towards the leaf apex (Figure 9d,e) or just near the apex (O. kamtchatica from Kamchatka (Figure 8a–d) and Chukotka (Figure 9a)). The lowest primary pinnae, and deeply pinnatifid first basal basiscopic secondary pinnae branching out of them, are asymmetrical. These secondary pinnae are 15 to 32 mm long and 5 to 13 mm wide.
Pinnules widely triangular, triangular or narrowly triangular, from unlobed to pinnately parted, are attached by a broad base mostly possessing a distinctly decurrent basal margin. Apices are acute, rounded or cuspidate. Margins are entire or dentate. The basal acroscopic pinnule of the primary pinnae tends to be parallel to the main rachis. Basal basiscopic pinnule arises from the base of the pinna rachis or decurrent to the main rachis or rachis of the basal primary pinna. The basal basiscopic lobe arises from the midrib base or is decurrent to the rachis. Some basal basiscopic pinnules or lobes of primary pinnatifid branches and lowest basal secondary branches are attached with a slightly contracted basal margin but without a decurrent basal margin.
In the middle part of the leaf possessing deeply pinnatifid pinnae, the pinnules are purely unlobed. The largest pinnules are situated on the second from the base pinnae, they are up to 7 mm long and 2.5 mm wide. Pinnules of deeply pinnatifid pinnae in the upper third are either larger than ones in the lower third (O. kamtchatica from Kamchatka (Figure 8a–d) and Chukotka (Pl. 9, a)) or smaller than ones in the lower third (O. kamtchatica from Alaska (Figure 9b–d)).
In the basalmost pinnae, the basal pinnule of the acroscopic side is always unlobed. Each subsequent pinnule of the specimens, preserving the largest leaves (Figure 8a,b; Figure 9a), is larger and more divided, up to being pinnately lobed to being parted in the middle part of the pinnae. Then, pinnules start to diminish gradually and thus become less divided up to unlobed near the pinna apex. In smaller leaves (Figure 8c,d; Figure 9b–d), all pinnules of the acroscopic side of the first basal primary pinna are unlobed, with the largest of them being located in the middle part of the pinna. The first branch of the basiscopic side of the basal primary pinna is either a pinnately parted pinnule (Figure 9b,d,e) or a deeply pinnatifid secondary pinna (Figure 8a–d; Figure 9a,c), with subsequent branches gradually diminishing and becoming less divided up to unlobed pinnules near the pinna apex. The basal basiscopic pinnately parted pinnule (secondary branch) is asymmetrically compound: the basiscopic side is more divided than the acroscopic, and basiscopic lobes are larger than acroscopic ones; each pinnule is up to 16 mm long and 5 mm wide, and the basiscopic lobe measures up to 4.5 × 2 mm.
Pinnules of the deeply pinnatifid lowest secondary pinna are mostly unlobed. Basiscopic ones are larger than those that are acroscopic. In the holotype, the largest pinnule (third branch) is about 10 mm × 4 mm (Figure 8a,b).
The venation is pinnate. The midrib enters the pinnule at an acute angle near the basiscopic margin at the base of the pinnule. Towards the apex, the midrib branches become gradually narrower and indistinct. The midrib and secondary (lateral) veins are distinct, and higher order vein branches are indistinct. Unlobed narrowly triangular pinnules have up to seven, or perhaps more, forked and diverging lateral veins on each side of the midrib. The first basiscopic lateral vein arises from the midrib base, or from the main rachis or the rachis of the basal primary or first basiscopic secondary pinnae. Lateral veins arise nearly opposite and become catadromic or alternate in catadromic order. They fork up to three, perhaps more, times.
Comparison and discussion: Most of O. kamtchatica leaves are distinctly larger and more divided than the only known leaf of O. ochotensis. However, the least divided O. kamtchatica leaf (Figure 9e) resembles O. ochotensis both in the frond dissection and size. O. kamtchatica differs from O. ochotensis mainly in that all its pinnules are attached by a broad base without obviously narrowed base margins. In O. ochotensis, the first basal acroscopic pinnule of the lowest primary pinnae is attached by a distinctly narrowed base, and the second one is attached with strongly narrowed basiscopic and acroscopic base margins. The basiscopic basal pinnule of O. kamtchatica arises from the base of the primary pinna rachis or is quite often decurrent to the main rachis or the rachis of the basalmost primary pinna; the basal basiscopic lobe arises from the base of the midrib or is decurrent to the rachis or rachis it emerges from; the first basiscopic lateral vein arises from the midrib base or the main rachis or the rachis of the lowest primary or secondary pinna. The similar O. ochotensis pinnule arises near or from the base of the pinna rachis; lobes of the same kind and vein arise near or from the midrib base.
A very similar specimen to the least divided O. kamtchatica in size, form, degree of division, attachment of pinnules and other features from Alaska (Figure 9e) was described as Ochotopteris pekulneensis Philippova [44,45] from the lower Coniacian Tylpegyrgynai Formation of the Pekulnei Ridge western slope. According to Filippova, O. pekulneensis differs from O. kamtchatica from the type locality in its smaller size and absence of lobed pinnules, and the pinnules have cuspidate apices. We, however, believe that these criteria are not sufficient enough to consider Pekulnei leaves of Ochotopteris, and the similar frond from Alaska (Figure 9e) is a species different from O. kamtchatica.
Ochotopteris lebedevii Herman and Domogatskaya, sp. nov.

Figure 10.
Ochotopteris lebedevii Herman and Domogatskaya, sp. nov., holotype: (a,b)—specimen no. 101-352b-1, (c,d)—specimen no. 101-352a-1 (counterpart); collection no. 3381 (GIN RAS); Ul’ya Trough, locality 101; Yemanra Formation, middle Albian to Turonian. Scale bar is 1 cm.

Figure 11.
Ochotopteris lebedevii Herman and Domogatskaya, sp. nov.: (a)—specimen no. 101-374-1, (b)—specimen no. 101-101-374-2, (c)—specimen no. 101-101-374-3, (d)—specimen no. 101-101-352a-3; collection no. 3381 (GIN RAS); Ul’ya Trough, locality 101; Yemanra Formation, middle Albian to Turonian. Scale bar is 1 cm.

Figure 12.
Ochotopteris lebedevii Herman and Domogatskaya, sp. nov.: (a)—specimen no. 101–8; (b)—specimen no. 101–351; collection no. 3381 (GIN RAS); Ul’ya Trough, locality 101; Yemanra Formation, middle Albian to Turonian. Scale bar is 1 cm.
2023 Ochotopteris sp.: Shczepetov and Herman, pl. XI, Figure 2, Figure 3, Figure 4, Figure 5 and Figure 6 and 9, pl. XII, Figure 2, pl. XIII, Figure 1; Figure 2 [35].
Holotype: designated here, specimen no. 3381/101-352a-1 and 3381/101-352b-1 (counterpart) (Figure 10a–d).
Repository: GIN RAS, Moscow, Russia, collection no. 3381.
Type locality: Ul’ya Trough, Amka River, locality 101 (approx. 59°24′ N, 140°23′ E).
Type stratum: Yemanra Formation, middle Cretaceous, and middle Albian to Turonian.
Etymology: in honour of palaeobotanist E.L. Lebedev.
Material: samples 101-8, 101-351, 101-352a, b-1, 101-352b-3, and 101-374 (collection no. 3381, Ul’ya Trough, Amka River; GIN RAS, Moscow).
Occurrence: Ul’ya Trough, Amka River, Yemanra Formation, middle Cretaceous and most probably middle–upper Albian and/or Cenomanian–Turonian.
Diagnosis: It has a sterile widely ovate frond, once pinnate-pinnatifid in apical part and bipinnate-pinnatifid in basal part. The main rachis is robust, with a long and almost straight petiole. The basalmost primary pinnae are asymmetrical: basal pinnule of the acroscopic side is unlobed while the basal pinnule of the basiscopic side is pinnately parted, or the pinnule of the acroscopic side is unlobed to be pinnately parted while the first acroscopic branch is a deeply pinnatifid secondary pinna. In bipinnate–deeply pinnatifid (nearly tripinnate) leaves, a basal basiscopic pinnule arises from the base of pinna rachis or is decurrent to the rachis it emerges from; in the basalmost primary pinnae, a basal acroscopic pinnule is attached by constricted base; the other secondary acroscopic branches, excluding the basal one and the upper third ones, always have the form of pinnately lobed–parted pinnules or secondary pinnae. The basal basiscopic lobe arises from the base of the midrib or is decurrent to the main rachis or basalmost primary pinna rachis. Lateral veins arise nearly opposite becoming catadromous or alternate in catadromous order.
Description: It has a sterile widely ovate frond, once pinnate-pinnatifid in the apical part with pinnately parted pinnules, or typically with deeply pinnatifid pinnae (nearly bipinnate) and bipinnate-pinnatifid in the basal part with pinnately parted pinnules, or typically with deeply pinnatifid pinnae (near tripinnate). The lamina is 18 to 57 mm long and 19 to 90 mm wide, with a robust main rachis. The petiole is up to 22 mm long, almost straight. Pinnae, except for the basalmost primary ones, are narrowly elliptic and slightly curved in shape. The largest deeply pinnatifid (second from the base) pinnae are 15 to 33 mm long and 5 to 10 mm wide. The basalmost primary pinnae are 11 to 50 mm long and 7 to 35 mm wide, sub-opposite; emerging at an angle of 20–45°, near the second acroscopic branch located at an angle of 60–110°. Towards the frond apex, the angle of the primary pinnae departure gradually diminishes; the last primary pinnae are alternate. The basalmost primary pinnae emerging from them are deeply pinnatifid basiscopic, and the basalmost secondary pinnae are asymmetrical. These secondary pinnae are up to 25 × 7 mm in size.
Pinnules are ovate, lance-ovate or lanceolate, narrowly elliptic, triangular or narrowly triangular in shape; unlobed or pinnately lobed to parted; attached typically by a broad base with a distinctly decurrent basal margin. Apices are acute or rounded. Margins are entire, sometimes undulate, or dentate. The basal acroscopic pinnule of the primary pinnae tends to be parallel to the main rachis. The basal basiscopic pinnule arises from the base of the pinna rachis or is decurrent to the rachis it emerges from. A basiscopic basal lobe arises from the midrib base or is often decurrent to the main rachis or rachis of the basal primary pinna.
In the middle part of the frond with deeply pinnatifid primary pinnae, the pinnules are unlobed. The largest pinnules are situated in the middle part of the pinna, and they are up to 6 mm long and 2.5 mm wide. Pinnules in the upper third of a pinna are slightly smaller than ones in the lower third, or they are of the same size. Some basal pinnules possess rounded teeth or undulate margins and are attached with slightly contracted lower and upper base margins: two or less teeth of basiscopic side, one or less of acroscopic side. Least divided leaves (Figure 11b,d) do not have pinnules of this kind. The holotype (Figure 10a–d) has one basiscopic and one acroscopic pinnule of this kind (located on the second and third from the base pinnae). The largest leaves (Figure 11a; Figure 12b) have one acroscopic and two basiscopic pinnules of this kind on the second pinnae and one acroscopic pinnule on the third pinnae.
In the basalmost primary pinnae of the least divided leaf (Figure 11d), the apical part of the pinna, approximately one-third, is not pinnately divided; the acroscopic pinnule has an undulate margin and is attached with a strongly contracted basal margin of its base; the basiscopic basal branch has the form of a pinnately cleft pinnule, and the other pinnules are unlobed. The first basal acroscopic pinnule of basalmost pinnae of other leaves is unlobed with rounded teeth (Figure 11c), or usually pinnately lobed-cleft with up to five lobes with rounded margins on each side of the midrib, is attached by a constricted base and is up to 9 mm long and 3 mm wide. The largest and the most divided pinnules of the acroscopic side, almost of the same degree of division as the secondary pinnae, are situated in the middle part of the basalmost pinna, while pinnules in the upper third of the pinna are usually unlobed and smaller than the ones in its lower third. The basal basiscopic lobe of the second acroscopic pinnately parted pinnule (Figure 12a) also has rounded teeth and is attached with a slightly contracted basiscopic base margin. The first branch of the basiscopic side has the form of a deeply pinnatifid secondary pinna, while subsequent branches gradually diminish and become less divided, up to unlobed, near the apex.
The pinnules of deeply pinnatifid first basiscopic secondary pinnae are unlobed. Basiscopic ones are larger than acroscopic ones, up to 5 mm long and 2 mm wide. The base of the basal basiscopic pinnule of the basal secondary pinnae is slightly narrowed on both sides.
Venation is pinnate. The midrib enters the pinnule near the basiscopic margin at its base and at an acute angle. The midrib becomes narrower and sinuous towards the frond apex, then branches and disappears near it. The midrib and lateral veins are distinct, while higher-order vein branches are less distinct. The first basiscopic lateral vein arises from the midrib base, the main rachis or rachis of the lowest primary pinnae. The lateral veins are nearly opposite and become catadromous or, in the most divided pinnules, alternate in catadromous order. Unlobed pinnules have up to eight forking and diverging lateral veins on each side of the midrib. Lateral veins are forked two to three, and perhaps more, times.
Comparison and discussion. Both in O. lebedevii and O. ochotensis, the first basal acroscopic pinnule of the basalmost primary pinnae is attached by the noticeably constricted base. O. lebedevii differs from O. ochotensis in its larger size and more divided pinnae and pinnules. Apart from that, the basiscopic basal pinnule of O. lebedevii arises from the rachis base or quite often is decurrent to the rachis it emerges from; the basal basiscopic lobe arises from the pinnule base, or quite often is decurrent to the main rachis or rachis of basalmost primary pinna. The first basiscopic lateral vein arises from the midrib base, or main or pinna rachis. Pinnules of the same kind in O. ochotensis arise near to or from the base of the pinna rachis, while the lobes and veins of the same kind arise near or from the midrib base. O. lebedevii, with deeply pinnatifid pinnae and an unlobed acroscopic basal pinnule of the basalmost primary pinnae, resembles O. ochotensis in size. The main difference of such Ochotopteris lebedevii sp. nov. fronds is that the successive acroscopic branches (excepting the upper third ones) of the basal primary pinnae have the form of pinnately lobed-parted pinnules or a secondary pinna, whereas in O. ochotensis, these pinnules are unlobed to barely lobed.
All O. lebedevii sp. nov. leaves have long, almost straight, petioles, while both O. ochotensis and O. kamtchatica have no petioles, or have a small part of them, usually curved near the bases of sub-opposite basal pinnae. The most apparent difference between the new species and O. kamtchatica is that in the former the basal acroscopic pinnule of the basalmost primary pinnae is attached by the notably constricted base, which is not observed in O. kamtchatica. The other difference is that in O. lebedevii with deeply pinnatifid pinnae, almost all acroscopic pinnules (except the upper ones) of the lowest primary pinnae resemble pinnately lobed-parted pinnules or a secondary pinna, while O. kamtchatica has unlobed or pinnately lobed-cleft (only in the middle pinna part) acroscopic pinnules. Another difference is that O. lebedevii with deeply pinnatifid pinnae has basal acroscopic pinnules of the second, and sometimes the third from the base, primary pinnae attached with contracted basiscopic and acroscopic margins, while in O. kamtchatica acroscopic pinnules are attached by a broad base without the contracted base margins.
5. Discussion and Conclusions: The “Extinct” Palaeoclimate of the mid-Cretaceous Arctic
The known habitats of Ochotopteris ferns were all situated in high latitudes of the Northern Hemisphere (Figure 13) with the estimated palaeolatitudes of the sites being between 60.4° N (Western Priokhot’ye, Tyl’ River, Albian) and 80.4°N (Chandler River, Albian) (Table 1). Among the Ochotopteris species, O. kamtchatica is the most “Arctic”, with this species being situated north of palaeolatitude of 72° N. The mid-Cretaceous Arctic palaeoclimate of this area was frost-free temperate to warm temperate and wet [2]. It possessed the typical insolation for near-polar latitudes with continuous summer daylight for several weeks (“polar day”) and continuous winter darkness for several weeks (“polar night”) [51]. The combination of these characteristics makes the mid-Cretaceous Arctic palaeoclimate unique in that near-polar southern latitudes lack the thermal moderation of a polar ocean. In our modern “icehouse” Earth, there are no analogues of this kind of climate—it simply does not exist.
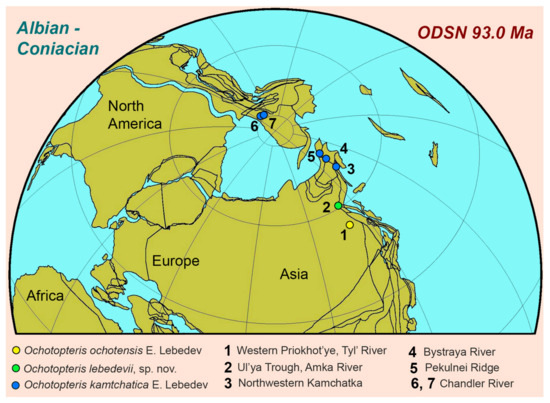
Figure 13.
Palaeogeographic distribution of Ochotopteris species.
As was pointed out above, the mid-Cretaceous Arctic flora possessed a large number of endemic plants, mainly ferns (including Ochotopteris) and angiosperms. The high endemism of the high-latitude mid-Cretaceous floras of North-Eastern Asia and Northern Alaska most probably reflect the adaptation of these floras to the very unusual environmental conditions of the Cretaceous Arctic: the unique combination of warm and wet summers with continuous daylight and predominantly warm winters with the pronounced scarcity of the winter sunlight.
In summer, daylight duration at high latitudes far exceeds that at low latitudes. In summer, in near-polar regions, the amount of photosynthetically useful solar radiation is greater than that at low latitudes due to a considerably longer day. Moreover, high-latitude solar energy is different to that at lower latitudes in that it consists of more diffuse light. The prevalence of diffuse light at high latitudes is caused by the relatively low sun position above the horizon (low angle of light incidence), high vapour content in the atmosphere of polar areas, and frequent mists and cloudiness [52]. Diffuse light is utilized by plants more efficiently than direct light, and it is absorbed more effectively and does not cause the overheating of leaves. Hence, during the long summer growing season, the near-polar plants did not suffer from a scarcity of warmth, moisture and solar energy. Abundant and diverse mid-Cretaceous near-polar floras were highly productive.
This high-latitude mid-Cretaceous palaeoclimate can be exemplified by the climatic conditions experienced by the latest Albian—Cenomanian Grebenka flora from the Upper Subformation of the Krivorechenskaya Formation on the Anadyr River right bank, North-eastern Russia [40,41,52,53]. The Grebenka flora seems to be one of the most diverse middle Cretaceous floras of Eurasia despite its position within the mid-Cretaceous Arctic Circle. The calculated palaeolatitude of the early Cenomanian Yelisseev locality of the Grebenka Flora is 73.5° N (Table 1). This means that each year the ancient plants experienced a continuous summer daylight for 14–15 weeks in May–August and a continuous winter darkness for about 13 weeks in November–January bounded before and after that by 3–4 weeks of twilight periods [51].
Based on its systematic composition and physiognomy, the Grebenka flora could be characterised as mixed coniferous and broad-leaved deciduous, or predominantly deciduous, forest with horsetails, ferns, caytonialeans, leptostrobaleans, cycadaleans and ginkgoaleans in understorey [40,41]. The high floristic diversity of the Grebenka flora provides a reliable quantitative climatic signal of conditions near the mid-Cretaceous North Pole. Using the leaf physiognomic multivariate analysis CLAMP [54] (http://clamp.ibcas.ac.cn/, accessed on 28 June 2023) on the whole composite flora, we determined the palaeoclimate variables experienced by its plants [2]. A preliminary estimation of the palaeoclimate variables was made using the most complete PHYSG3ARC set of morphological data on modern floras and the corresponding GRIDMET3ARC modern climate calibration data set. This estimation showed that the Grebenka flora plants did not experience sub-zero temperatures during the year. This made it possible to obtain more accurate results with the reduced error of the method using the set of morphological data PHYSG3BRC, since this data set does not include the floras growing in cold climates, and the corresponding set of high-resolution grid climate calibration GRIDMET3BR. The following paleoclimatic variables were obtained (for each of them, the standard deviation ± σ is indicated):
- Mean annual temperature (MAT) 12.9 ± 1.1 °C;
- Warm month mean temperature (WMMT) 20.8 ± 1.4 °C;
- Cold month mean temperature (CMMT) 5.9 ± 1.9 °C;
- Length of the growing season (LGS) 7.4 ± 0.7 months;
- Mean growing season precipitation (GSP) 823 ± 196 mm;
- Mean monthly growing season precipitation (MMGSP) 93 ± 26 mm;
- Precipitation during the three wettest months (3WET) 577 ± 138 mm;
- Precipitation during the three driest months (3DRY) 145 ± 32 mm;
- Relative humidity (RH) 72.5 ± 5.2%;
- Specific humidity (SH) 8.0 ± 1.0 g/kg;
- Enthalpy (ENTHAL) 317 ± 5 kJ/kg.
The estimates indicate that according to the Köppen classification of global climates [55,56], the plants of the Grebenka flora, as well as the majority of other mid-Cretaceous Arctic floras, grew in a warm humid climate with a warm summer and a mild frost-free winter (Cfb climate). There is little reason to assume cold winters with substantial (weeks or months) periods of subfreezing conditions existed in North-Eastern Asia and Northern Alaska at this time. The above results show that the growing of these floras in high latitudes is unlikely to have been restricted by temperature, moisture or scarcity of daylight during the growing season.
A large number of cycadophyte plants in the Grebenka flora (genera Cycadites, Nilssonia, Nilssoniocladus and Taeniopteris), and the abundance of their fossils in the burials, also indicate a warm and frost-free palaeoclimate. Although there was a small number of evergreen plants in the Grebenka Flora [41,52], in general, the evergreenness (overwintering without leaf shedding) was highly unfavourable for the plants: a warm temperate and frost-free climate, combined with the near-polar light regime and evergreenness, would inevitably have led to a metabolical drain on the plant’s resources during the dark and above freezing winter. Most of the Grebenka flora plants were deciduous, i.e., able to drop the individual leaves, or shoots with leaves attached, during winter. Examples include caytonealeans, ginkgoaleans, czekanowskialeans, some conifers (Cephalotaxopsis, Taxites, Elatocladus, some Sequoia and Parataxodium) and all the angiosperms. At least some cycads were deciduous as well: plants with Nilssoniocladus shoots were capable not only of shedding the individual leaves but also of discarding their “unwanted” leaf load extremely rapidly in the short autumn by abscising short shoots and attached leaves as single units [57]. The absence of a long winter period cold enough to suppress plant metabolism enough to make leaf retention energetically worthwhile implies that the deciduousness of most of the Grebenka plants reflects their strong adaptation to high latitude light seasonality and the prolonged winter darkness. Horsetails and ferns including Ochotopteris probably died back to rhizome systems during the polar winters.
Author Contributions
A.B.H.: fossil plant taxonomy and palaeoecology, regional stratigraphy and palaeoclimate; K.V.D.: taxonomic description and photographic illustrations of the Ochotopteris species. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding. The research was performed within the framework of the State program No. FMMG-2023-0003 of the Geological Institute, Russian Academy of Sciences, Moscow, Russia.
Data Availability Statement
http://arcticfossils.nsii.org.cn/, accessed on 28 June 2023.
Acknowledgments
We are sincerely grateful to Sergei Shczepetov (Komarov Botanical Institute, Russian Academy of Sciences, St. Petersburg, Russia) for his helpful comments on the local stratigraphy of North-Eastern Asia and Jiří Kvaček (National Museum, Praha, the Czech Republic) for his advice on plant taxonomy. For many years, we discussed our ideas and results on the Cretaceous Arctic palaeobotany, stratigraphy and palaeoclimate with Bob Spicer (The Open University, Milton Keynes, UK). He also kindly agreed to correct the English of our paper. We highly appreciate his friendship and help. We are sincerely grateful to the Editor and two anonymous reviewers for their helpful amendments and comments on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hay, W.W.; DeConto, R.; Wold, C.N.; Wilson, K.M.; Voigt, S.; Schulz, M.; Wold, A.R.; Dullo, W.-C.; Ronov, A.B.; Balukhovsky, A.N.; et al. Alternative Global Cretaceous Palaeogeography. In The Evolution of Cretaceous Ocean/Climate Systems; Barrera, E., Johnson, C., Eds.; Special Paper 332; Geological Society of America: Boulder, CO, USA, 1999; pp. 1–47. [Google Scholar]
- Spicer, R.A.; Herman, A.B. The Late Cretaceous environment of the Arctic: A quantitative reassessment based on plant fossils. Palaeogeogr. Palaeoclimat. Palaeoecol. 2010, 295, 423–442. [Google Scholar] [CrossRef]
- Spicer, R.; Valdes, P.; Hughes, A.; Yang, J.; Spicer, T.; Herman, A.; Farnsworth, A. New insights into the thermal regime and hydrodynamics of the early Late Cretaceous Arctic. Geol. Mag. 2020, 157, 1729–1746. [Google Scholar] [CrossRef]
- Spicer, R.; Herman, A.; Spicer, T.; Yang, J. Arctic Plant Fossils. Polar Vegetation in the Cretaceous and Paleogene. Available online: http://arcticfossils.nsii.org.cn/ (accessed on 25 June 2023).
- Samylina, V.A. Mesozoic flora of the Kolyma River left bank (Zyrianka coal basin), Part 1: Horsetails, ferns, cycads and bennettites. Proc. Botan. Inst. USSR Acad. Sci. Ser. 8 Palaeobot. 1964, 5, 40–79. (In Russian) [Google Scholar]
- Grabovskiy, A.A. Genus Arctopteris (Pteridaceae) in the Cretaceous floras of the Northeast of Russia. Palaeobotany 2021, 12, 34–55. [Google Scholar] [CrossRef]
- Samylina, V.A. Birisia—New genus of the Cretaceous ferns of Siberia. Bot. Zhurnal 1972, 57, 94–101. (In Russian) [Google Scholar]
- Grabovskiy, A.A.; Golovneva, L.B. Genus Birisia (Dicksoniaceae) in the Cretaceous floras of Siberia. Palaeobotany 2018, 9, 32–53. (In Russian) [Google Scholar] [CrossRef]
- Samylina, V.A.; Filippova, G.G. New Cretaceous ferns of North-eastern USSR. Paleontol. Zhurnal 1970, 2, 90–97. (In Russian) [Google Scholar]
- Lebedev, E.L. Al’bskaya Flora i Stratigrafiya Nizhnego mela Zapadnogo Priokhot’ya (Albian Flora and Lower Cretaceous Stratigraphy of Western Priokhot’ye); Nauka Publishers: Moscow, Russia, 1974; Volume 254, 148p. (In Russian) [Google Scholar]
- Herman, A.B.; Lebedev, E.L. Stratigrafiya i Flora Melovykh Otlozhenii Severo-Zapadnoi Kamchatki (Cretaceous Stratigraphy and Flora of Northwestern Kamchatka); Nauka Publishers: Moscow, Russia, 1991; Volume 468, 189p. (In Russian) [Google Scholar]
- Maslova, N.P.; Herman, A.B. New finds of fossil Hamamelids and data on the phylogenetic relationships between the Platanaceae and Hamamelidaceae. Paleontol. J. 2004, 38, 563–575. [Google Scholar]
- Herman, A.B.; Golovneva, L.B. New genus of the Late Cretaceous platanoids of the North-Eastern USSR. Bot. Zhurnal 1988, 73, 1456–1467. (In Russian) [Google Scholar]
- Moiseeva, M.G. Barykovia, a new genus of angiosperms from the Campanian of northeastern Russia. Rev. Palaeobot. Palynol. 2012, 178, 1–12. [Google Scholar] [CrossRef]
- Samylina, V.A. Angiosperms from the Lower Cretaceous of the Kolyma River. Bot. Zhurnal 1960, 45, 335–352. (In Russian) [Google Scholar]
- Alekseev, P.I.; Herman, A.B.; Shchepetov, S.V. New angiosperm genera from Cretaceous sections of Northern Asia. Stratigr. Geol. Correl. 2014, 22, 606–617. [Google Scholar] [CrossRef]
- Lebedev, E.L.; Herman, A.B. A new genus of Cretaceous angiosperms—Dalembia. Rev. Palaeobot. Palynol. 1989, 59, 77–91. [Google Scholar] [CrossRef]
- Golovneva, L.B.; Abramova, L.N. New plants of the Rarytkin flora (Late Cretaceous, Koryak Highland). Bot. Zhurnal 1990, 75, 92–97. (In Russian) [Google Scholar]
- Lebedev, E.L. New genus of the Cretaceous angiosperms of the North-eastern Asia—Grebenkia. Paleontol. Zhurnal 1986, 1, 134–139. (In Russian) [Google Scholar]
- Samylina, V.A. Early Cretaceous angiosperms of the Soviet Union based on leaf and fruit remains. Bot. J. Linn. Soc. 1968, 61, 207–218. [Google Scholar] [CrossRef][Green Version]
- Golovneva, L.B. New genus Palaeotrapa (Trapaceae?) and new species of Quereuxia from the Rarytkin Formation (Koryak Highland, Maastrichtian-Danian). Bot. Zhurnal 1991, 76, 601–606. (In Russian) [Google Scholar]
- Herman, A.B. New genus of platanoid angiosperms from the Upper Cretaceous deposits of Kamchatka. Paleontol. Zhurnal 1984, 1, 71–79. (In Russian) [Google Scholar]
- Hollick, A. The Upper Cretaceous floras of Alaska. U.S. Geol. Surv. Prof. Pap. 1930, 159, 1–123. [Google Scholar]
- Herman, A.B. New angiosperms from the Turonian of Northwestern Kamchatka. Paleontol. Zhurnal 1987, 4, 96–105. (In Russian) [Google Scholar]
- Golovneva, L.B. New genus Platimelis in the Late Cretaceous—Early Paleogene floras of Arctic. Bot. Zhurnal 1994, 79, 98–107. (In Russian) [Google Scholar]
- Filippova, G.G. New Cretaceous angiosperms from the Anadyr River basin. Paleontol. Zhurnal 1978, 1, 138–144. (In Russian) [Google Scholar]
- Filippova, G.G. New angiosperms from the Coniacian deposits of the northern part of the Pekulney Ridge. Paleontol. Zhurnal 1991, 3, 85–93. (In Russian) [Google Scholar]
- Filippova, G.G. Fossil plants from the Cretaceous deposits of the Arman River basin (Northern Priokhot’ye). Paleontol. Zhurnal 1988, 4, 88–95. (In Russian) [Google Scholar]
- A Paleolatitude Calculator for Paleoclimate Studies. Available online: http://paleolatitude.org/ (accessed on 23 May 2023).
- Van Hinsbergen, D.J.J.; de Groot, L.V.; van Schaik, S.J.; Spakman, W.; Bijl, P.K.; Sluijs, A.; Langereis, C.G.; Brinkhuis, H. A Paleolatitude Calculator for Paleoclimate Studies. PLoS ONE 2015, 10, e0126946. [Google Scholar] [CrossRef]
- ODSN Plate Tectonic Reconstruction Service. Available online: http://www.odsn.de/odsn/services/paleomap/paleomap.html (accessed on 12 May 2023).
- Samylina, V.A. Rannemelovye Flory Severo-Vostoka SSSR (k Probleme Stanovleniya Flor Kainofita) (Early Cretaceous Floras of Northeastern USSR (On the Problem of Cenophytic Floras Establishing)); Nauka Publishers: Leningrad, Russia, 1974; Volume XXVII, 56p. (In Russian) [Google Scholar]
- Herman, A.B. Albian—Paleocene flora of the North Pacific: Systematic composition, palaeofloristics and phytostratigraphy. Stratigr. Geol. Correl. 2013, 21, 689–747. [Google Scholar] [CrossRef]
- Lebedev, E.L. Stratigrafiya i Vozrast Okhotsko-Chukotskogo Vulkanogennogo Poyasa (Stratigraphy and Age of the Okhotsk-Chukotka Volcanigenic Belt); Nauka Publishers: Moscow, Russia, 1987; Volume 421, 175p. (In Russian) [Google Scholar]
- Shczepetov, S.V.; Herman, A.B. Volcanogenic Cretaceous of North-eastern Asia: Taphofloras of the Ul’ya and Urak interfluve (on the knowledge of E.L. Lebedev’s legacy). Stratigr. Geol. Correl. 2023, 31. in press. [Google Scholar]
- Pergament, M.A. Stratigrafiya Verkhnemelovykh Otlozhenii Severo-Zapadnoi Kamchatki (Penzhinskii Raion) (Stratigraphy of the Upper Cretaceous Deposits of North-Western Kamchatka, Penzhina Region); The Academy of Sciences of the U.S.S.R.: Moscow, Russia, 1969; Volume 39, 147p. (In Russian) [Google Scholar]
- Pergament, M.A. Stratigrafiya i Inotseramy Verkhnego Mela Severnogo Polushariya (Upper Cretaceous Stratigraphy and Inoceramids of the Northern Hemisphere); Nauka Publishers: Moscow, Russia, 1978; Volume 322, 214p. (In Russian) [Google Scholar]
- Herman, A.B. Al’bskaya–Paleotsenovaya Flora Severnoi Patsifiki (Albian–Paleocene Flora of the North Pacific Region); GEOS Publications: Moscow, Russia, 2011; Volume 592, 280p. (In Russian) [Google Scholar]
- Terekhova, G.P. On the age of the Krivorechenskaya Formation and the Grebenka floral assemblage. In Stratigrafiya i Paleontologiya Fanerozoya Severo-Vostoka SSSR (Phanerozoic Stratigraphy and Palaeontology of the Northeastern USSR); North-Eastern Integrated Scientific Research Institution: Magadan, Russia, 1988; pp. 100–117. (In Russian) [Google Scholar]
- Shchepetov, S.V.; Herman, A.B.; Belaya, B.V. Srednemelovaya Flora Pravoberezh’ya r. Anadyr’: Stratigraficheskoye Polozheniye, Sistematicheskii Sostav, Atlas Iskopayemykh Rasteniy (Mid-Cretaceous Flora of the Anadyr River Right Bank: Stratigraphic Setting, Systematic Composition, Plant Fossils Atlas); North-Eastern Integrated Scientific Research Institution: Magadan, Russia, 1992; 166p. (In Russian) [Google Scholar]
- Spicer, R.A.; Ahlberg, A.; Herman, A.B.; Kelley, S.P.; Raikevich, M.I.; Rees, P.M. Palaeoenvironment and ecology of the middle Cretaceous Grebenka Flora of Northeastern Asia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 184, 65–105. [Google Scholar] [CrossRef]
- Devyatilova, A.D.; Nevretdinov, E.B.; Filippova, G.G. Upper Cretaceous stratigraphy of the Anadyr River middle reaches basin. Geol. I Geofiz. 1980, 12, 62–70. (In Russian) [Google Scholar]
- Pokhialainen, V.P. Mel Severo-Vostoka Rossii (Cretaceous of the North-Eastern Russia); North-Eastern Integrated Scientific Research Institute, Far Eastern Branch of the Russian Academy of Sciences (NEISRI RAS): Magadan, Russia, 1994; 37p. (In Russian) [Google Scholar]
- Filippova, G.G. Cretaceous stratigraphy and flora of the northern part of the Pekulnei Ridge (Chukotka). Stratigr. Geol. Correl. 2005, 13, 79–99. (In Russian) [Google Scholar]
- Filippova, G.G. Stratigrafiya i Flora Melovykh Otlozhenii Severnoi Chasti Khrebta Pekulnei (Cretaceous Stratigraphy and Flora of the Northern Part of the Pekulnei Ridge); North-Eastern Integrated Scientific Research Institute, Far Eastern Branch of the Russian Academy of Sciences (NEISRI RAS): Magadan, Russia, 2010; 203p. (In Russian) [Google Scholar]
- Terekhova, G.P.; Filippova, G.G. Tylpegyrgynai Formation and its floral assemblage (Pekulney Ridge, north-eastern USSR). Dokl. Akad. Nauk. SSSR (USSR Acad. Sci. Rep.) 1984, 278, 1443–1446. (In Russian) [Google Scholar]
- Filippova, G.G. Coniacian flora of the northern part of the Pekulney Ridge. Kolyma 1994, 3, 13–21. (In Russian) [Google Scholar]
- Craggs, H.J. Late Cretaceous climate signal of the northern Pekulney Range flora of Northeastern Russia. Palaeogeogr. Palaeoclimat. Palaeoecol. 2005, 217, 25–46. [Google Scholar] [CrossRef]
- Detterman, R.L.; Bickel, R.S.; Gryc, G. Geology of the Chandler River region, Alaska. U.S. Geol. Surv. Prof. Pap. 1963, 303-E, 223–324. [Google Scholar]
- Smiley, C.J. Cretaceous Floras of Chandler-Colville Region, Alaska: Stratigraphy and Preliminary Floristics. Amer. Assoc. Petrol. Geol. Bull. 1969, 53, 482–502. [Google Scholar]
- Central Intelligence Agency. Handbook (1978) Polar Regions Atlas; National Foreign Assessment Center; Central Intelligence Agency: Washington, DC, USA, 1978; pp. 1–66.
- Herman, A.B.; Spicer, R.A. Mid-Cretaceous Grebenka Flora of the North-eastern Russia: Two strategies of overwintering. In Proceedings of the Fifth European Palaeobotanical-Palynological Conference, Krakow, Poland, 26–30 June 1998; Polska Akademia Nauk, Instytut Botaniki im W. Szafera: Krakow, Poland, 1999; (Suppl. S2), pp. 107–110. [Google Scholar]
- Filippova, G.G.; Abramova, L.N. Pozdnemelovaya Flora Severo-Vostoka Rossii (Late Cretaceous Flora of the North-Eastern Russia); Nedra Publications: Moscow, Russia, 1993; pp. 1–348. (In Russian) [Google Scholar]
- CLAMP Online. Climate Leaf Analysis Multivariate Program. Available online: http://clamp.ibcas.ac.cn/ (accessed on 17 April 2023).
- Köppen, W. Das geographische System der Klimate. In Handbuch der Klimatologie; Köppen, W., Geiger, R., Eds.; Gebrüder Bornträger: Berlin, Germany, 1936; pp. 1–44. [Google Scholar]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Spicer, R.A.; Herman, A.B. Nilssoniocladus in the Cretaceous Arctic: New species and biological insights. Rev. Palaeobot. Palynol. 1996, 92, 229–243. [Google Scholar] [CrossRef]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).