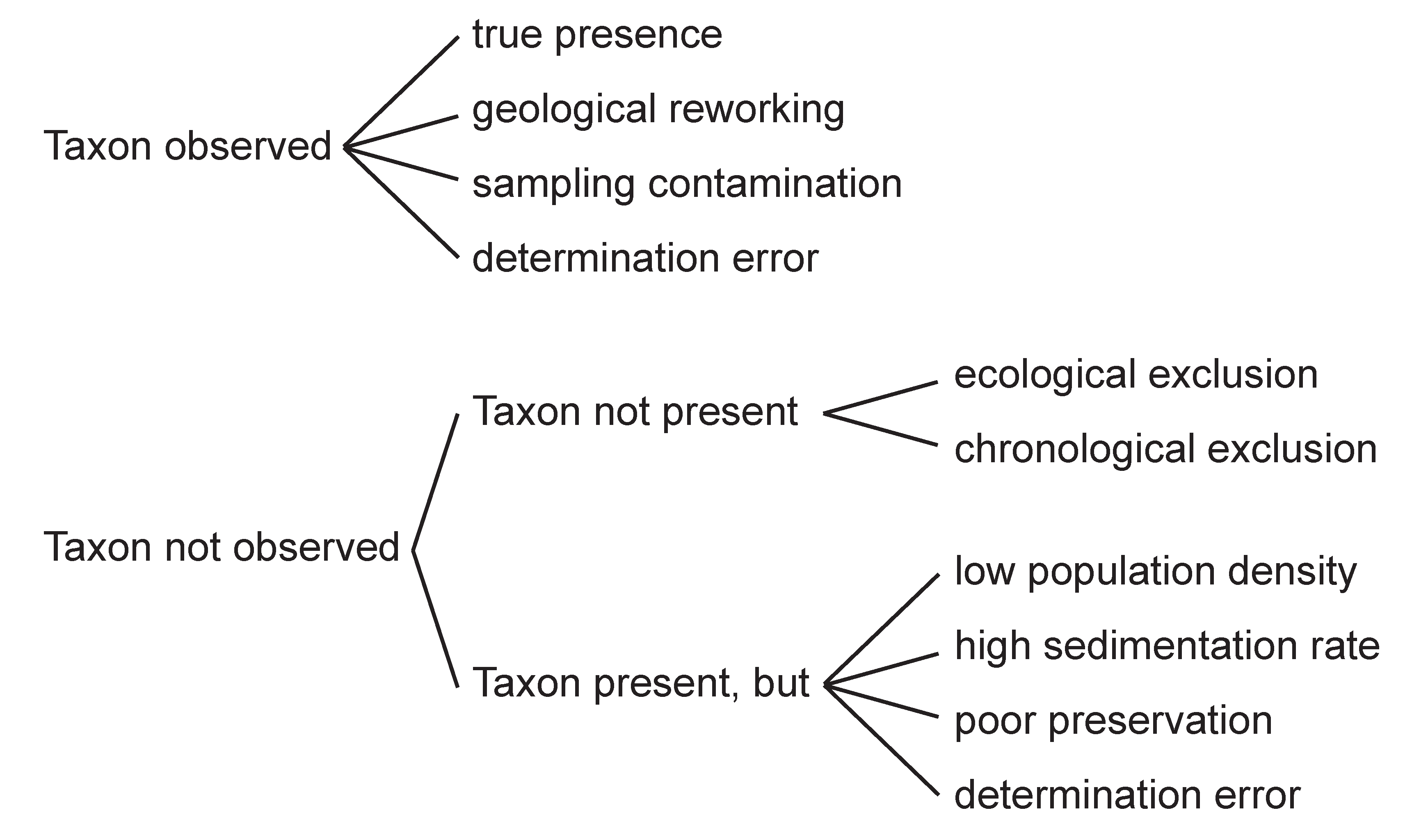
5. Stratigraphic Inventory
Below we describe in stratigraphic order, from Toarcian through Berriasian-early Valanginian our sample material with planktonic foraminifera as free tests from washed sample residues. The listing of these 25 localities worldwide also includes critical literature references with planktonic assemblages from which we do not have specimens or sample material (indicated with a small x).
Figure 3, in an approximate manner shows the majority of localities from which Jurassic planktonic foraminifera are known [
3] including the key ones on which the current analysis is based. The zonal and age assignments, as derived from ammonite or nannofossil zonations follow the chronostratigraphic subdivisions of the Jurassic and earliest Cretaceous stages in [
8,
9,
10]. Note that the early, middle and late subdivision of stages is not formalized and open to change.
Middle Toarcian, SW Turkey (x)
Bifrons—Variabilis Zones, thin-sections,
Globuligerina spp.; tests on average 150 µm [
11].
Early Aalenian, SW Turkey
Opalinum Zone, thin-sections,
Globuligerina spp., (thick walled), tests on average 250 µm [
11].
Early Bajocian, Morocco (RIF) (x)
Sauzi Zone,
Globuligerina spp., tests on average 280–300 µm, maximally 340 µm [
12]
Early Bajocian, NW Australia (x)
Study and locality: Small clay samples from marine dredges acquired along the NW Australian Continental Margin, facing the Argo Abyssal Plain of Tethys contain about 75+ fragile and small, aragonitic planktonic foraminifera.
Age: Early Bajocian, using nannofossils and dinoflagellates.
Microfossil assemblage:Globuligerina oxfordiana, G.bathoniana and a species that maybe endemic to this region. The assemblage needs comparative study.
Reference: Apthorpe (2020) [
13].
Middle Bajocian, Hungary (x)
Study and locality: Hard “Ammonitico Rosso” limestone of Som Hill, Bakony Mountains yielded a diversified and rich planktonic foraminifera assemblage, using weak acidisation to disengage the tests from the limestone matrix. The acidisation does not preserve wall surface textures of the test, necessary to differentiate genera.
Zones and age:Humphriesianum and Niortense Zones, middle Bajocian.
Assemblage: The planktonic assemblage is diverse and dominated by large specimens, often reaching 400 µm in diameter, including Globuligerina aff. dagestanica, Globuligerina oxfordiana (medium and large forms), Globuligerina aff. bathoniana (large form), and Globuligerina avariformis.
Reference: Wernli and Görög (1999) [
14].
Middle Bajocian, France (x)
Study and locality: Find of Jurassic planktonic foraminifera in the type section of the Bajocian Stage, France.
Zone:Humphriesianum Zone, middle Bajocian.
Assemblage: Poorly preserved Globuligerina oxfordiana in a biomicrite with ferriginous oolites, attesting to the shallow marine nature of the sediment.
Reference: Bignot and Janin (1984) [
15].
Late Bajocian, Azerbijan (x)
In 1984 Kasimova and Aliyeva [
16] briefly described
Conoglobigerina avariformis Kasimova from the Bajocian of Azerbaijan. The holotype drawings provide no wall sculpture information, but a metatype specimen illustrated in [
17] has short pseudomurical ridges.
Late Bajocian-Early Bathonian, Dagestan (Russia)
Study and locality: In 1961, Morozova and Moskalenko [
18] described for the first time a rich Jurassic ‘
Globigerina’ biofacies, with abundant free specimens of several taxa. The assemblage occurs in fine grained, bathyal shales near the villages of Gunib and Chokh, Central Dagestan, NE Caucasus. The Gunib section has geographic coordinates 42°23′16’’ N, 46°57′40’’ E, and the Chock section 42°19′18’’ N, 47°1′57’’ E.
Zones and Age:Parkinsonii and Zigzag Zones, late Bajocian-early Bathonian.
Assemblage: Globuligerina dagestanica, Globuligerina balakhmatovae and Globuligerina oxfordiana. The planktonic assemblage is associated with epistominids and discorbids.
Through the courtesy and cooperation of Temirbekova Umuhayabat Temirbekovna (Makhachkala, Dagestan, Russia) we obtained Jurassic planktonic specimens from the type sections near Gunib and Chokh in Central Dagestan. This allowed us to study topotype specimens. Through the courtesy of Ludmila Kopaevich we also obtained micropaleontology slides with planktonic foraminifera of the Gunib and Chokh sections from the Gorbachik collection in Moscow.
References: Morozova and Moskalenko (1961) [
18]; Gradstein et al. (2017) [
1].
Late Bajocian-Early Bathonian, Dagestan (Russia)
Study and locality: Co-author L.G. participated in 2015 in a field trip to a nearby and stratigraphically identical section (42°9′23″ N, 47°7′60″ E) to Gunib and Chokh. This means that the Jurassic planktonic foraminifera in it are essentially topotypes of those described first by Morozova and Moskalenko [
18]. The stratigraphic levels in this new section XУPУKPA (Khouroukra) were studied and sampled together with specialists in ammonite and belemnites. The studied part of the Khouroukra section includes the lower and middle subformation of the Tsudakhar Formation.
The lower subformation shows interbedded light gray siltstones and sandstones. The middle subformation consists of a dark gray claystones, with interbeds of siltstones, rare sandstones, with siderite concretions. There are rare and thin (0.10–0.15 m) interbeds of calcareous rocks.
Zones and age:Parkinsonii and Zigzag Zones, late Bajocian-early Bathonian.
Assemblage: Gastropodes, ostracodes and a relatively rich and diversified benthonic foraminifera assemblage with Hyperammina, Trochammina, Reophax, Ophthalmidium, Ichthyolaria, Dentalina, Lenticulina, Oberhauserella, Epistomina and Discorbis (D. paraspis), together with planktonic foraminifera. The latter are well preserved and common and include Globuligerina dagestanica, Globuligerina oxfordiana, Globuligerina glinskikhae and Globuligerina balakhmatovae.
References: Gradstein et al. (2017) [
1]; Gradstein and Waskowska (2021) [
2].
Bajocian-Bathonian-Callovian, Grand Banks, offshore eastern Canada
Study and stratigraphy: From 1970 until 1995 the senior author F.G. was professionally involved with the biostratigraphy of the sedimentary wedge along the continental margin of eastern Canada.
Zones and age: Garantella-, Globuligerina bathoniana- and Reinholdella crebra var. Zones, Bajocian through Callovian.
Microfossil assemblages: In side-wall-cores (swc) and cuttings (cts) samples between 10600 and 6230 feet in the Eider M-75 well Globuligerina bathoniana and Globuligerina oxfordiana occur in abundance, and Globuligerina balakhmatovae is common. Preservation of the calcite tests is excellent. The same three species also occur in the same stratigraphic interval in the Murre G-67 and Cormorant N-83 wells. In swc’s at 5000 and 5350 feet in the Bittern M-62 well, assigned to the Reinholdella crebra var Zone (Callovian) Globuligerina tojeiraensis occurs with few specimens together with a small and compact globigerinid form tentatively determined as Conoglobigerina grigelisi. Deeper in the same well assigned to the Globuligerina bathoniana Zone (Bathonian) occur Globuligerina balakhmatovae and Globuligerina bathoniana.
References: Gradstein (1976) [
19], Stam (1986) [
20], Gradstein et al. (2017) [
1].
Middle Bathonian, Portugal
Rare specimens of
Globuligerina bathoniana occur in the shallow marine sedimens of middle Bathonian age in the Brenha Road section, Mondego area, Portugal [
20].
Middle Bathonian, Poland
Study and locality: In 1969, Pazdrowa published ‘Bathonian Globigerina from Poland’ [
21]. The taxon
Globigerina bathoniana was described with about 150 well-preserved specimens from the ore-bearing clays of Ogrodzieniec, Zawiercie County, Southern Poland at 50°27′ N and 19°31′ E. Author F.G. received from Pazdrowa a foraminiferal assemblage from Ogrodzieniec, including the globigerinid taxon. Later, he undertook self micropaleontological sampling of the strata in the Ogrodzieniec claypit; these samples were used in the Ph.D. study by Stam [
20]. Now, the Ogrodzieniec quarry has been partly filled in and made inaccessible (but see below).
Zone and age:Morrisi Zone, middle Bathonian.
Assemblage: The local foraminiferal assemblage is diverse, and from the list of taxa received from Olga Pazdrowa includes:
Garantella ornata,
Reinholdella crebra,
Epistomina regularis,
E. costifera,
E. nuda,
Paleomiliolina czestochowiensis,
Ophtalmidium agglutinans,
O. carinatum,
Paalzowella pazdroae,
Spirillina radiata and
Conorboides paulus. According to [
22] the Jurassic planktonic foraminifera lived in a shallow marine basin with siliciclastic sedimentation. We interpret this paleo environment as probably deep neritic, using the evidence from epistominids with Jurassic planktonic foraminifera from other basins (Stam, 1986).
References: Pazdrowa (1969) [
21]; Stam (1986) [
20]; Gradstein et al. (2017) [
1].
Middle Bathonian, Poland
Study and locality: After the Ogrodzienie quarry became non-existent (see above), co-author AW undertook fieldwork and sampling of the same Morissi ammonite Zone in the Gnaszyn iron ore quarry in the southern part of Kraków-Silesia. The Gnaszyn (Gnaszyńscy) quarry (coordinates: 50°48’11.4” N, 19°02’31.9” E) is located in the S.W. Częstochowa city area of the Gnaszyn district. The 25 m thick section of dark shales represents the higher part of the Częstochowa ore-bearing clay formation being locally 140 m thick. The monotonous, strongly bioturbated black or dark greyish siltstones and claystones are rich in bioclasts, occasionally with different amount of sandy admixture. It contains seven levels of sphaerosideritic concentrations (N-T horizons).
Zone and age:Morrisi Zone—middle Bathonian.
Fossil assemblage: There is a diverse and rich fossil fauna with ammonites, belemnites, bivalves, scaphopods, gastropods, foraminifers, echinoderms and shark teeths, as well as trace fossils, calcareous nannoplankton, pollen, sphoromorphs, dinoflagellates and driftwoods. Benthic foraminifera include are dominated by Ceratobuliminidae and Vaginulinidae, and smaller numbers of agglutinated taxa.
The Jurassic planktonic foraminifera bearing GN-6 sample was taken in the lower part of the Gnaszyn section. This is a grey and slightly sandy claystone with numerous bioclasts, mainly of mollusks shells. The sample is rich in very well preserved, aragonitic planktonic foraminifera with Globuligerina oxfordiana and fewer Globuligerina bathoniana, Globuligerina glinskhikae and Globuligerina waskowskae.
References: Smoleń (2012) [
22]; Gedl et al. (2012) [
23]; Gradstein et al. (2017) [
1]; Gradstein & Waskowska (2021) [
2].
Middle Callovian, Portugal
Study and locality: Along the Mareta Beach section in southern Portugal, thick and well bedded calcareous shales crop out, locally with thin sand layers with ripple marks, load and flute casts, and upwards in the beach section with more marls and limestones. The shales and marls have common Bositria.
Zone and Age: Bordalo da Rocha (1976) [
24] assigned a late Bajocian through Callovian age to the lithological unit, using a detailed ammonite zonation.
Micropaleontologic assemblage: Middle Jurassic foraminiferal paleoecology and Jurassic planktonic foraminifera in the Mareta Beach section were investigated by Stam (1986). In sample 31 (Coronatum Zone, middle Callovian), the author recognized rare Globuligerina bathoniana. The neritic setting of the Callovian strata, with few epistominids, does not lend itself to find Jurassic planktonic foraminifera in abundance.
References: Bordalo da Rocha (1976) [
24]; Stam (1986) [
20]; Gradstein et al. (2017) [
1].
Early Oxfordian, NW France
Study and locality: In 1966, Bignot and Guyader [
25] discovered several dozen small specimens of
Globuligerina oxfordiana (Grigelis) in the clays of Villers, Le Havre, Normandy, Northwest France.
Zone, stage and microfossil assemblage: The level is lower Oxfordian,
Mariae Zone, about 6 m below the
Cordatum zone with ferruginous oolites. Jurassic planktonic foraminifera are from a low energy, shallow marine embayment, connected to open sea. Using the paleo waterdepth model of Stam (1986) the species lived in a shallow marine environment, not exceeding 150 m in paleo waterdepth (Samson et al., 1992) [
26]. G. Bignot kindly donated
Globuligerina oxfordiana specimens to Gradstein for comparison to Grand Banks specimens.
References: Bignot and Guyader (1966) [
25]; Stam (1986) [
20]; Gradstein et al. (2017) [
1]; Gradstein and Waskowska (2021) [
2].
Middle Oxfordian, Switzerland
Study: In April 2016, author Gradstein visited the Natural History Museum in Basel, Switzerland to study type slides listed to contain Globigerina helvetojurassica Haeusler, and also sample the nearby Eisengraben Section to obtain fresh microfossil material containing this species. The museum type slides did not contain suitably preserved specimens of this taxon, and are unrecognizable, but topotypes of this taxon were successfully sampled, under the expert guidance of Etter and Knappertsbusch.
Locality: The Eisengraben Section, ca 35 km N–W of Zuerich in Canton Aargau, Switzerland is the type section of the Birmenstorfer Schichten,
Tranversarium Zone, middle Oxfordian. The section has coordinates 651,600/264,070 on Schweizerische Landeskarte 1:25,000, Blatt 1069, Frick (reference
Figure 4 in Oesterle, 1968 [
27]).
Zone-stage: Transversarium Zone, middle Oxfordian.
Microfossil assemblage: Directly below level 4 in the Eisengraben Section, samples with common and well-preserved specimens were obtained that we consider to be topotypes of Conoglobigerina helvetojurassica (Haeusler). Also, rare specimens were obtained of small sized Globuligerina bathoniana and G.oxfordiana. Level 4 top has many agglutinated taxa, but hardly any planktonic specimens. Sample 4 mid, at the level of Stam’s sample with ‘helvetojurassica’ has best preserved specimens, and also includes well-preserved benthics belonging in Bigenerina, Textularia, Ophthalmidium, Ammodiscus, Glomospira, Trochammina, Haplophragmoides, Dentalina, Nodosaria, Lenticulina plus some indeterminate taxa. Micro gastropods also occur. Small limonitic concretions are common; small glauconite lumps also occur, testifying to a likely neritic mode of deposition. There are no epistominid benthic taxa.
Micropaleontology slides of Oesterle (1968) [
27] in the Basel Museum from the Birmenstorfer type section contain 50+ of many more (often broken) specimens which we studied that show the same test moulds of crystallized carbonate fragments. The peculiar preservation is the result of a rigorous processing technique (likely a form of acetolysis). At least five morphotypes may be distinguished, which, despite poor preservation, can easily be assigned to the species
Globuligerina aff.
oxfordiana,
Globuligerina aff.
bathoniana,
Globuligerina aff.
jurassica (with an elongated, high spired and irregularly trochospiral test),
Globuligerina aff. balakhmatovae,
Globuligerina aff
. tojeiraensis (with slightly elongated last chamber) and
Conoglobigerina aff
. grigelisi. However, the latter might be
Globuligerina aff
. avariformis, since we cannot determine wall texture. For sampling details and plates with SEM illustrations of all planktonic taxa, including the topotypes of
Conoglobigerina helvetojurassica see [
28].
References Oesterle (1968) [
27]; Gradstein et al. (2017) [
1].
Oxfordian, Lithuania and Russia
Study and localities: A brief description is provided of the type localities of Globuligerina oxfordiana based on the author’s (A. Grigelis) notes made on boreholes during his field work in Lithuania in 1956, and in the Nikitino region of central Russia in 1977. Microfaunal remnants were picked by the author after standard sediment disintegration and hand-sieving procedure. The shallow burial at the Oxfordian localities accounts for excellent preservation of the Globuligerina oxfordiana taxon.
Geologically, the Lithuanian area belongs to the Precambrian East European Craton. Relatively thin Mesozoic sedimentary strata underly Quaternary surface deposits. Marine Jurassic is distributed in the western-south-western part of Lithuania, with thickness changing from less than 30 m in the northwest to 250 m in the southwest, against the border with Poland.
Microfossil assemblage: Holo- and paratypes of
Globuligerina oxfordiana come from dark claystone and dark siltstone in borehole Jotija (Jotija village), Sakiai District, Lithuania at 143 m below surface in sample 10, lower Oxfordian (type specimens are provided by A. Grigelis on Plate 12 in [
1]). In the original description it is not indicated on which evidence geological age is assigned, but the foraminiferal assemblage contains
Spirophthalmidium birmenstorfense,
Lenticulina brueckmanni,
L. comptala, Vaginulina flabellata,
Trocholina transversarii,
Epistomina volgensis,
E. brueckmanni and
Epistominoides sp. GPS coordinates of the borehole site are 55°02′17’’ N, 23°11′42’’ E.
Well-preserved specimens of Globuligerina oxfordiana also occur in dark, micaceous silty clay in the interval of 55–66 m in borehole Lyduvenai, Raseiniai District, Lithuania. GPS coordinates of this site are 55°30‘29’’ N, 23°04‘59’’ E. The type level contains the ammonite Cardioceras tenuicostatum Nikitin (early Oxfordian) and the foraminifera Epistomina volgensis, Epistomina intermedia and Lenticulina brueckmanni.
In Central Russia, well preserved specimens of this taxon (as assigned by Grigelis) also occur in sample 801 in the Shatriche-2 outcrop at the Nikitino settlement on the river Oka, Ryazan District. The actual outcrop is on high right bank of River Oka, ca 2.5 km below the confluence with River Pronya, at outskirt of village Shatriche, in a river washout called “Durnyanki”. The sampled sediments are assigned to the middle to late Oxfordian, but no specific fossil information is provided. Type specimens of this species provided by A.Grigelis on Plate 12 in [
1].
References: Grigelis (1958) [
29]; Gradstein et al. (2017) [
1].
Oxfordian, Poland
Study and locality: A prominent Globuligerina limestone in Poland occurs in the Pieniny Klippen Belt (PKB). Co-author A.W. obtained hard limestone samples of Oxfordian age from the isolated Obłazowa Klippe block in Nowa Biała village (coordinates: 49°25’42,4” N, 20°07’35,5” E).
Micropaleontologic assemblage: Thin-sections show that Globuligerina oxfordiana is abundant and rock-forming, and Globuligerina bathoniana is rare.
References: Birkenmajer (1977, 1986) [
30,
31]; Wierzbowski et al. (1999) [
32]; Gradstein et al. (2017) [
1].
Oxfordian, Blake Bahama Basin, North Atlantic Ocean
Study: Deep Sea Drilling Site (DSDP) 534, on the landward side of marine Jurassic Magnetic Quiet Zone (JQZ), in the Blake-Bahama Basin, established a Middle Jurassic (Bathonian-Callovian) age for the opening of the N. Atlantic Ocean.
Microfossil assemblage: The site cored abyssal (>2.7 km paleo waterdepth) Jurassic sediments with well preserved and common benthic foraminifera, nannofossils, radiolarians and dinoflagellates. It failed to find in situ Jurassic planktonic foraminifera. Rare, small and poorly preserved Globuligerina aff. oxfordiana specimens in Core 110 of Oxfordian age, are interpreted as transported with gravity flows from shallower strata. DSDP Leg 11, Site 105, drilled in this region in 1972, in this region yielded similar observation.
References: Gradstein (1983) [
33]; Luterbacher (1972) [
34].
Early Kimmeridgian, Portugal
Study: Between 1978 and 2016, author FMG with colleagues and students undertook several stratigraphic surveys and sampling in road outcrops, mountain sections and beach cliffs with Jurassic marine strata in Central and South Portugal. The research was part of a stratigraphic study of the Grand Banks of Newfoundland and its conjugate basin. The Grand Banks and Portugese Jurassic basins were conjugate, prior to mid-Cretaceous oceanisation of this segment of the North Atlantic.
Over the years, several stratigraphers and paleontologists generously provided guidance in the field, and assisted with analysis of ammonites and microfossils collected. These specialists included: Mouterde and Ruget Perrot (University of Lyon, Lyon, France), Ramalho (Geological Survey, Lisbon, Portugal), da Rocha (New University of Lisbon, Lisbon, Portugal), Wilson (The Open University, UK), Poulton (GSC, Calgary, AB, Canada), Gale and Turner (University of Portsmouth, Portsmouth, UK) and Watkins (Department of Earth and Atmospheric Sciences, Nebraska University, Lincoln, NE, USA).
Locality: Several sections in the marly Tojeira Formation, over 70 m thick, in hills near the hamlet of Ramada and the villages of Pereiro, Tojeira an Vila Verde dos Francos, Montejunto area, Estramadura, west central Portugal. Details are in [
1].
Zone-stage:Platynota—Hypselocyclum Zones, early Kimmeridgian
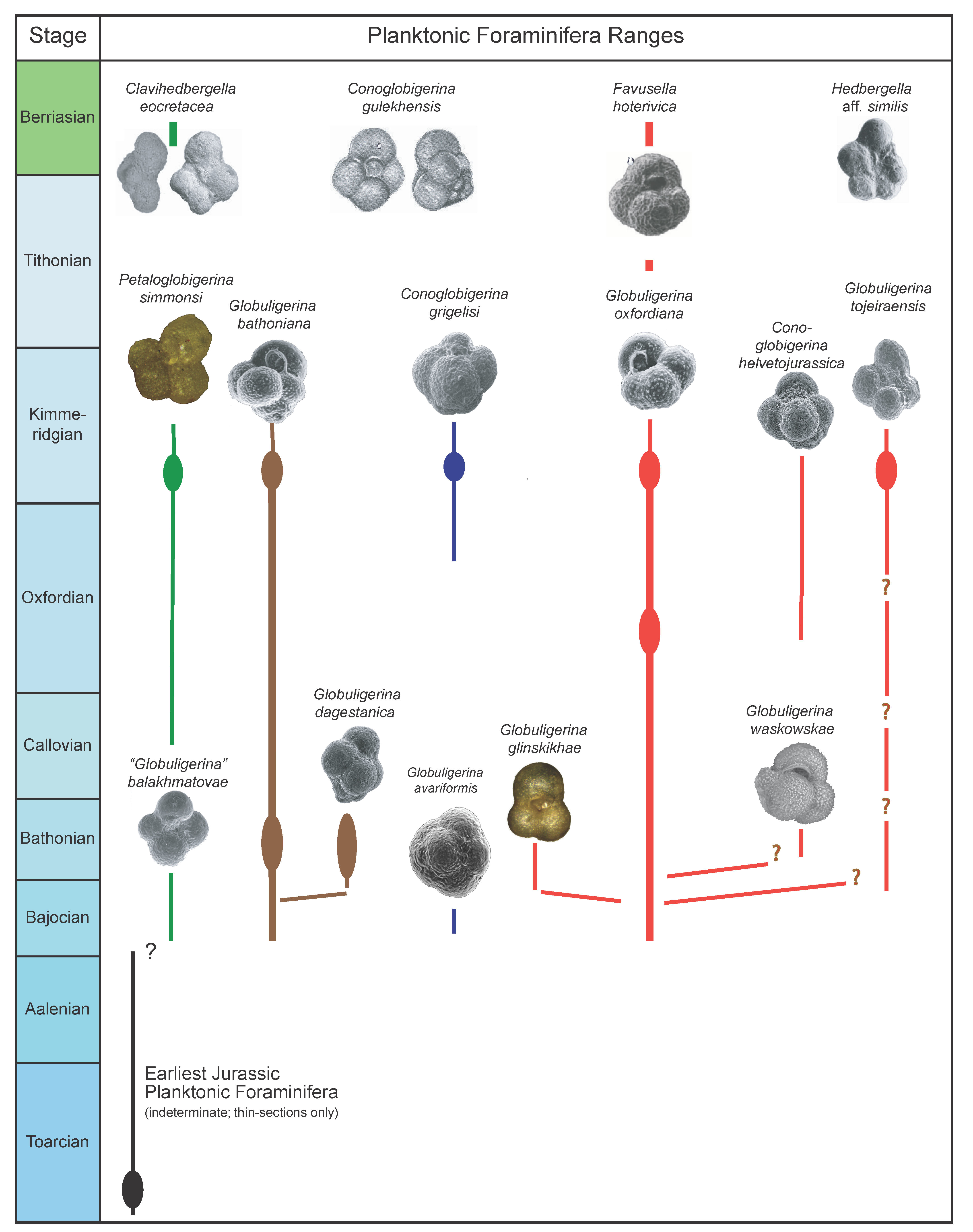
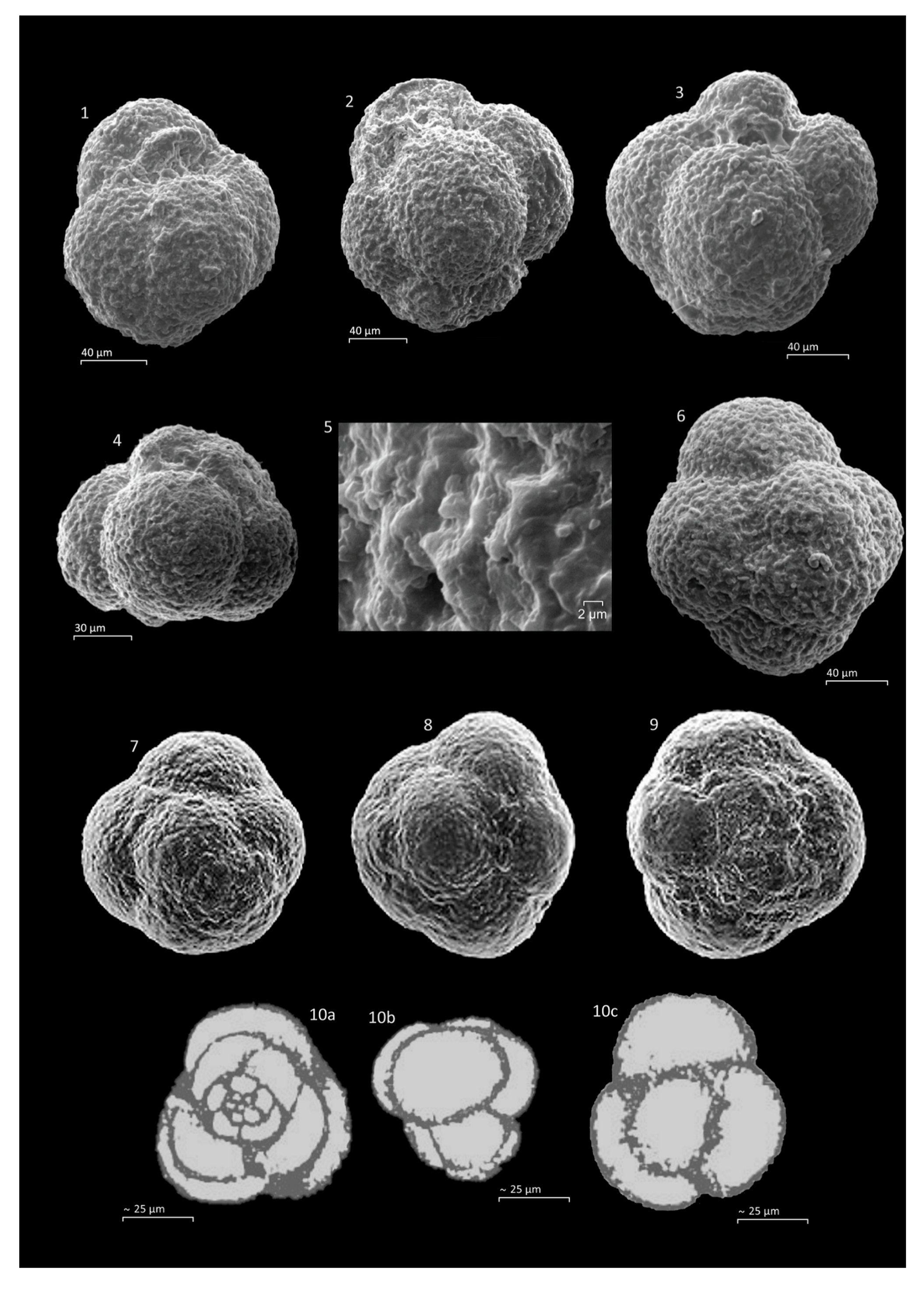
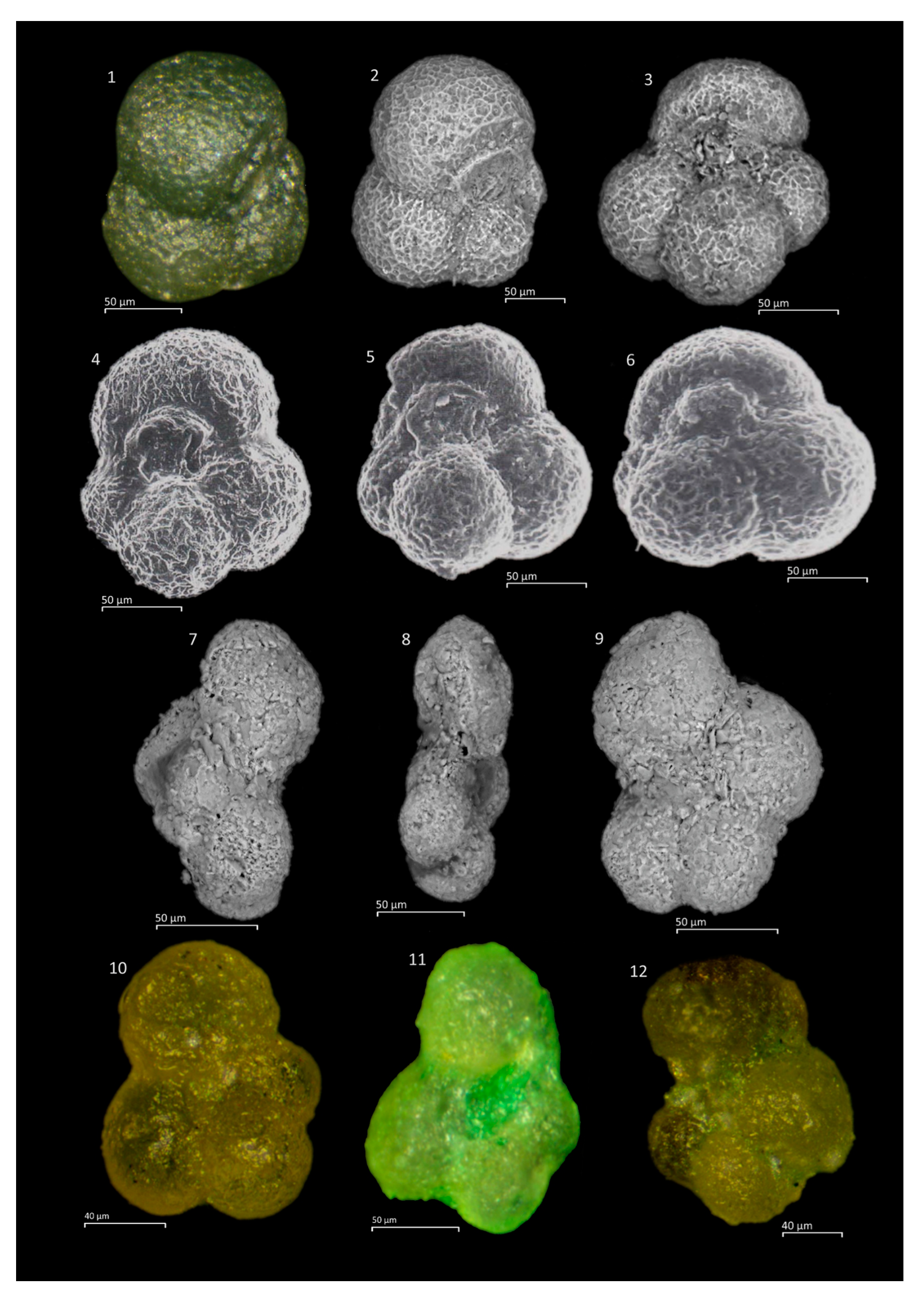
Microfossil assemblage: The Tojeira Formation is rich in fairly well preserved planktonic foraminifera and a very large collection of specimens was made, with seven taxa described. The taxa include
Globuligerina oxfordiana,
Globuligerina bathoniana,
Globuligerina tojeiraensis,
Globuligerina balakhmatovae,
Petaloglobigerina simmonsi,
Conoglobigerina grigelisi and
Conoglobigerina helvetojurassica. Planktonic foraminiferal tests are calcitic, nor aragonitic. The Portugese Jurassic planktonic foraminifera occur together with a rich epistominid assemblage, also found in Grand Banks sections with
Pseudolamarckina rjasanensis,
Epistomina mosquensis and
Epistomina uhligi. Agglutinated foraminiferal taxa and micro-gastropods are common; the calcareous benthic foraminifera
Discorbis paraspis and
Discorbis scutiliformis are rare. The benthic foraminiferal assemblage is only known from deeper neritic and bathyal settings [
20].
References: Stam (1986) [
20]; Agterberg et al. (1989) [
35]; Gradstein et al. (2017) [
1]; Turner et al. (2017) [
36]; Gradstein and Waskowska (2021) [
2]; Görög and Wernli (2013) [
37].
Middle Kimmeridgian, Portugal
Study and locality: In 1978, as part of the extensive Tojeira Formation sampling (see above), Rogerio Bordalo da Rocha (Lisbon) kindly guided author F.G. and students to two dark shales outcrops, one along the road just outside Villa Verde dos Francos and another one outside Pereiro, both in the Montejunto area. The fossiliferous shales with ammonites and Bositria are of the Abadia Formation, that overly the Tojeira Formation and underly thick-bedded conglomeratic sands. In the Montejunto area, the Abadia Formation, with shales, sands, conglomerates and olistolithic limestone blocks reach a thickness of over 1 km.
Zone and age: Middle Kimmeridgian, according ammonite studies of the Portugese Geological Survey.
Micropaleontologic assemblage: Our samples 4/1 and 25/1 have large size benthic foraminifera with Epistomina stellicostata, E.volgensis and E.uhligi. The first two taxa do not occur in the underlying Tojeira Formation, and have their first, regional stratigraphic occurrence in these two samples. In correlation with eastern Canada, this benthic foraminiferal assemblage is Kimmeridgian-Tithonian in age.
Planktonic foraminifera include Globuligerina bathoniana (abundant), common Globuligerina oxfordiana, rare Globuligerina tojeiraensis, rare Petaloglobigerina simmonsi and rare Conoglobigerina grigelisi. No Conoglobigerina helvetojurassica was observed, a species common in the upper part of the underlying Tojeira Formation. Some Globuligerina oxfordiana specimens have a reticulate wall texture, a feature taken up in the section on Evolution.
References: Ascoli et al. (1984) [
38]; Stam (1986) [
20]; Williamson and Stam (1988) [
39].
Tithonian, Hungary (x)
Study: Co-author A.W. and colleagues in 2019 visited the Paprét-árok profile in the Gerecse mountains of Hungary. This locality is described with outcrop details and a Tithonian planktonic foraminiferal assemblage in [
40]. It turns out the locality does not exist, and its publication and description are based solely on archival data. The locality originally was a railway cut.
Locality: Paprét-árok, Gerecse Mts, Hungary (x).
Lithology and Zone-Age: In the Paprét-árok profile the succession begins with red radiolarite (Lókút Radiolarite Formation) of Middle Jurassic-Kimmeridgian age. It is followed upwards with a massive and hard bank about 75 cm in thickness, grey, pink and purple in colour. This massive bank is subdivided in five beds numbered 6 to 2, from the base to the top: Beds 6–7 belong to the Beckeri and the Hybonotum Zones, Bed 5 questionably to the Darwini Zone and Bed 4 questionably to the Fallauxi Zone, all considered early Tithonian in age. Limestone Beds 3 and 2 are not zoned, but considered as probably late Tithonian in age. Above this massive limestone bank occur grey marl and breccia layers, belonging to the Lower Cretaceous Bersek Marl Formation.
Microfossil assemblage: ‘Protoglobigerinids’ are described from bed 5? Darwini Zone using thin sections and free specimens obtained with weak acetic acid. The authors consider the planktonic foraminiferal assemblage to be monospecific, with a species transitional between Globuligerina oxfordiana and Favusella hoterivica, but no pore features and wall texture can be determined, making the interpretation tentative. Stratigraphically higher-up, it states that the same section might yield Berriasian planktonic foraminifera, but none are listed.
Reference: Görög and Wernli (2004) [
40].
Tithonian-Valanginian, North Africa
Other localities: There is a meagre record of unspecified planktonic foraminifera in thin sections from Tethyan limestones of Tithonian through Valanginian age in N. Africa and Central Europe [
40].
Early Berriasian, Crimea
Study: For our study of Mesozoic planktonic foraminifera, Ludmila Kopaevich (Moscow) obtained samples from the Tonas River Basin in E. Crimea, Russia. The hard marly sediment interval is assigned to the
Jacobi ammonite Zone, early Berriasian, not far above the Jurassic-Cretaceous boundary. The Tonas River Basin is a locality from which Gorbachik and Poroshina [
41] described
Globuligerina caucasica and
Globuligerina gulekhensis.
Locality, with coordinates: Outcrop 2 km south of the village Krasnoselivka in the Tonas Basin, Crimea, Russia. Geographic coordinates are 44.927712 N, 34.634905 E.
Zones, stage: Calcareous nannofossil Zone CC2, late early to middle Berriasian;
Ammonites of the Jacobi and Grandis subzones of the Jacobi Zone of early Berriasian age. Benthic foraminifera of the Protopeneroplis ultragranulatus—Siphoninella antiqua Zone and a younger one—Quadratina tonassica—Siphoninella antiqua Zone.
Microfossil assemblage: Fairly well-preserved benthic foraminifera, with Epistomina stellicostata, E. volgensis and E. uhligi, Lamarckina sp., common Neobulimina sp., Patellina sp., Lenticulina spp., several nodosariid taxa, Bigenerina sp., Spiroplectammina spp., Textularia sp., Trochammina sp.
Sample Cr1 contains about 100 specimens of planktonic foraminifera taxa, all microperforate, including with frequent specimens Favusella hoterivica, with common specimens Clavihedbergella eocretacea and Conoglobigerina gulekhensis and with less than 10 specimens each of the taxa Hedbergella aff. similis, ?Favusella sp., Hedbergella aff. handousi and ? Globuligerina sp.
Only Conoglobigerina gulekhensis was previously known from Berriasian strata, and this only from two regions, Crimea and Azerbaijan. Favusella hoterivica (Subbotina) with certainty only is known from Valanginian through Aptian marine strata, and the other taxa only from Valanginian, Hauterivian or even younger stratigraphic intervals. All taxa are of Tethyan or sub-Tethyan origin.
Note: As outlined in some detail in the geological history study by Nikishin et al. (2017) [
42], the Crimea region underwent late Berriasian vertical movements, leading to a regional unconformity with erosion and karstification of underlying strata. The Krasnoselivka section preserves non-karstified Berriasian limestones, subject of our study.
References: Arkadiev et al. (2005) [
43]; Gorbachik and Poroshina (1979) [
41]; Gradstein et al. (2018) [
44].
Berriasian-early Valanginian, Scotian Shelf, offshore eastern Canada
Study: In 2019, author F.G. studied foraminifera in micropaleontology slides and washed residues (65–250 µm) from exploration well samples stored with the Geological Survey of Canada, Bedford Institute of Canada, Dartmouth, NS, Canada.
Locality: Exploration wells with interval studied: Oneida O-25, 8630–9460′below casing point at 6876′; Onondaga E-84, 11730–11810′, below casing point at 8121′; Glenelg J-48, 4165–4155 m, below casing point at 4134 m.
Zone, stage: Using dinoflagellates and benthic foraminifera, the studied interval is assigned a Berrisian-early Valanginian age.
Microfossil assemblage: In total about 50 specimens of Favusella hoterivica were obtained, many pyritized and with good wall texture details. Some questionable Conoglobuligerina aff. gulekhensis were found, but preservation is insufficient to provide reliable identification. Benthic foraminifera include Lenticulina busnardoi, Epistomina minireticulata, E.stellicostata, Neobulimina sp, Discorbis aff. valdendisensis, Discorbis sp., agglutinated indet. Micro gastropods are common to abundant, as they are in the Kimmeridgian Tojeira Formation in Portugal.
References: Ascoli (1976) [
45]; Wernli et al. (1995) [
46]; Barss et al. (1979) [
47].
Berriasian-early Valanginian, North Mexico (x)
Study: North Mexico calpionellid publication with thin sections.
Locality: Taraises Formation, north Mexico
Zone, stage: Calpionellid assemblages assigned to Berriasian-early Valanginian.
Microfossil assemblage: Poor thin-section illustrations of
Conoglobigerina gulekhensis,
Caucasella hoterivica,
Compactogerina sp. and
Gorbachikella sp. by Omaña et al. (2017) [
48] from Berriasian-early Valanginian limestones of the Taraises Formation in north Mexico. The images are non-determinate, but the illustration on
Figure 5 of Omaña et al. (2017) likely is
Favusella hoterivica. The type of the monotypic genus
Compactogerina is coarse perforate and belongs to Neogene
Neogloboquadrina pachyderma [
1,
3]. Lourdes Omaña kindly communicated to Gradstein (May 2018) that it is currently not advised to visit the Mexican site due to drug-related violence.
Reference: Omaña et al. (2017) [
48].
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}