Dynamic Variations in Fecal Bacterial Community and Fermentation Profile of Holstein Steers in Response to Three Stepwise Density Diets

 , , ,
, , , 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals, Experimental Design, and Sampling
2.3. VFA Detection and DNA Isolation
2.4. 16S rRNA Gene Sequencing and Analysis
2.5. Statistical Analysis
3. Results
3.1. Fecal pH Value, VFA, and NH3-N Concentration
3.2. Sequencing Depth and Coverage
3.3. Alpha Diversity
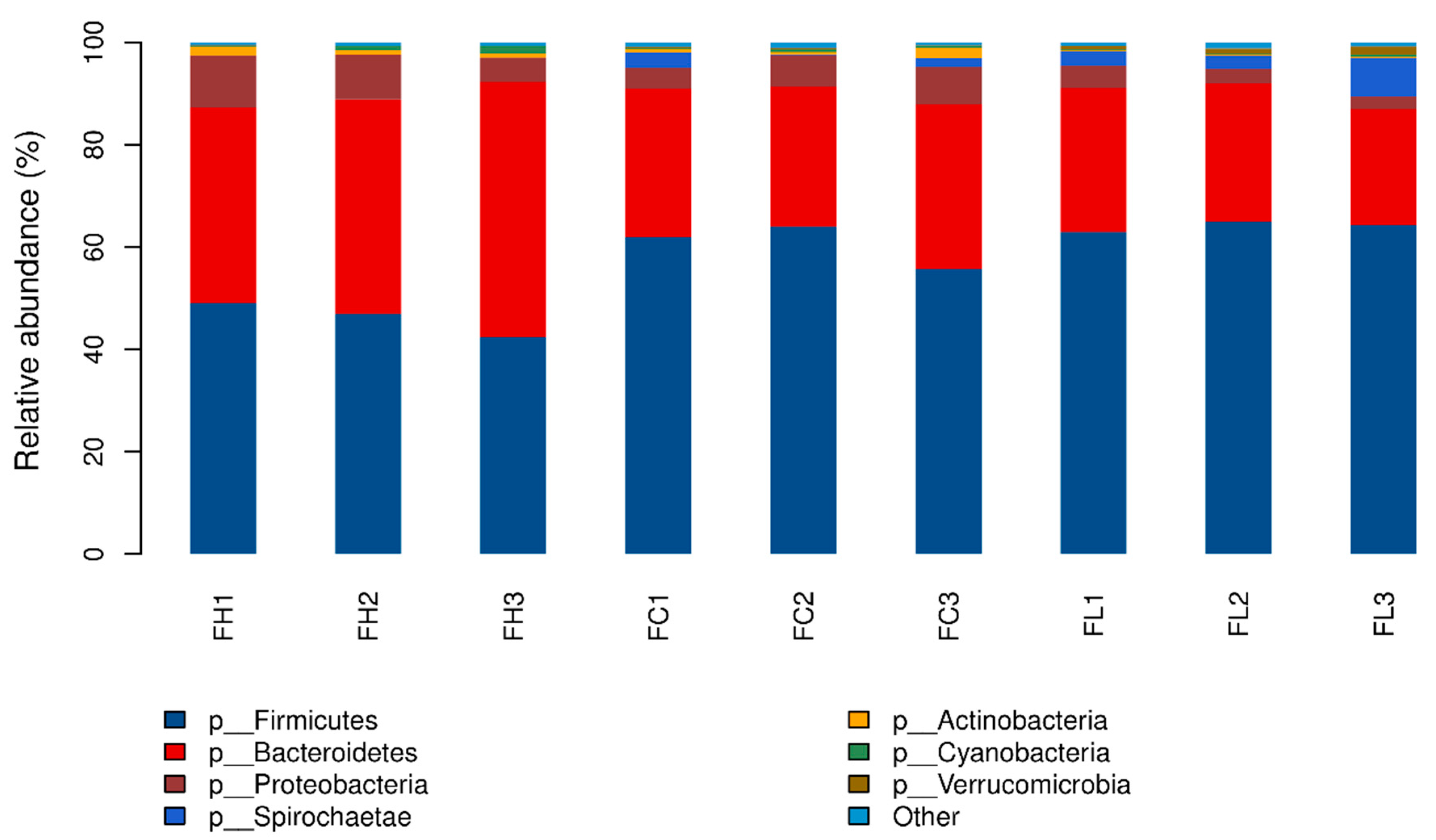
3.4. Taxonomic Profiles
4. Discussion
4.1. Fecal Fermentation Profile
4.2. Fecal Bacterial Diversity and Profile
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gressley, T.F.; Hall, M.B.; Armentano, L.E. Ruminant nutrition symposium: Productivity, digestion, and health responses to hindgut acidosis in ruminants. J. Anim. Sci. 2011, 89, 1120–1130. [Google Scholar] [CrossRef] [PubMed]
- Varadyova, Z.; Zelenak, I.; Siroka, P. In vitro study of the rumen and hindgut fermentation of fibrous materials (meadow hay, beech sawdust, wheat straw) in sheep. Anim. Feed Sci. Technol. 2000, 83, 127–138. [Google Scholar] [CrossRef]
- Lopes, D.R.G.; La Reau, A.J.; Duarte, M.D.; Detmann, E.; Bento, C.B.P.; Mercadante, M.E.Z.; Bonilha, S.F.M.; Suen, G.; Mantovani, H.C. The bacterial and fungal microbiota of Nelore steers is dynamic across the gastrointestinal tract and its fecal-associated microbiota is correlated to feed efficiency. Front. Microbiol. 2019, 10, 1263. [Google Scholar] [CrossRef] [PubMed]
- Doelman, J.; McKnight, L.L.; Carson, M.; Nichols, K.; Waterman, D.F.; Metcalf, J.A. Postruminal infusion of calcium gluconate increases milk fat production and alters fecal volatile fatty acid profile in lactating dairy cows. J. Dairy Sci. 2019, 102, 1274–1280. [Google Scholar] [CrossRef] [PubMed]
- Taschuk, R.; Griebel, P.J. Commensal microbiome effects on mucosal immune system development in the ruminant gastrointestinal tract. Anim. Health Res. Rev. 2012, 13, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Oikonomou, G.; Teixeira, A.G.V.; Foditsch, C.; Bicalho, M.L.; Machado, V.S.; Bicalho, R.C. Fecal microbial diversity in pre-weaned dairy calves as described by pyrosequencing of metagenomic 16S rDNA. associations of faecalibacterium species with health and growth. PLoS ONE 2013, 8, e63157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shi, H.; Wang, Y.; Cao, Z.; Yang, H.; Li, S. Effect of limit-fed diets with different forage to concentrate rations on fecal bacterial and archaeal community composition in Holstein heifers. Front. Microbiol. 2018, 9, 976. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, J.; Kuehn, L.A.; Bono, J.L.; Berry, E.D.; Kalchayanand, N.; Freetly, H.C.; Benson, A.K.; Wells, J.E. Investigation of bacterial diversity in the feces of cattle fed different diets. J. Anim. Sci. 2014, 92, 683–694. [Google Scholar] [CrossRef]
- Mao, S.; Zhang, R.; Wang, D.; Zhu, W. The diversity of the fecal bacterial community and its relationship with the concentration of volatile fatty acids in the feces during subacute rumen acidosis in dairy cows. BMC Vet. Res. 2012, 8, 237. [Google Scholar] [CrossRef]
- Shanks, O.C.; Kelty, C.A.; Archibeque, S.; Jenkins, M.; Newton, R.J.; McLellan, S.L.; Huse, S.M.; Sogin, M.L. Community structures of fecal bacteria in cattle from different animal feeding operations. Appl. Environ. Microbiol. 2011, 77, 2992–3001. [Google Scholar] [CrossRef]
- Klein-Jobstl, D.; Schornsteiner, E.; Mann, E.; Wagner, M.; Drillich, M.; Schmitz-Esser, S. Pyrosequencing reveals diverse fecal microbiota in Simmental calves during early development. Front. Microbiol. 2014, 5, 622. [Google Scholar] [CrossRef] [PubMed]
- Plaizier, J.C.; Li, S.; Danscher, A.M.; Derakshani, H.; Andersen, P.H.; Khafipour, E. Changes in microbiota in rumen digesta and feces due to a grain-based subacute ruminal acidosis (SARA) challenge. Microb. Ecol. 2017, 74, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Wells, J.E. A meta-analysis of bacterial diversity in the feces of cattle. Curr. Microbiol. 2016, 72, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Hua, C.; Tian, J.; Tian, P.; Cong, R.; Luo, Y.; Geng, Y.; Tao, S.; Ni, Y.; Zhao, R. Feeding a high concentration diet induces unhealthy alterations in the composition and metabolism of ruminal microbiota and host response in a goat model. Front. Microbiol. 2017, 8, 138. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; He, Y.; Li, H.; Wu, F.; Qiu, Q.; Niu, W.; Gao, Z.; Su, H.; Cao, B. Rumen fermentation, intramuscular fat fatty acid profiles and related rumen bacterial populations of Holstein bulls fed diets with different energy levels. Appl. Microbiol. Biotechnol. 2019, 103, 4931–4942. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Wu, H.; Wang, G.; Meng, Q.; Zhou, Z. The effects of including corn silage, corn stalk silage, and corn grain in finishing ration of beef steers on meat quality and oxidative stability. Meat Sci. 2018, 139, 142–148. [Google Scholar] [CrossRef] [PubMed]
- NRC. Nutrient Requirements of Beef Cattle, 8th ed.; The National Academies Press: Washington, DC, USA, 2016; pp. 397–399. [Google Scholar]
- Broderick, G.A.; Kang, J.H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and invitro media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]
- Paz, H.A.; Anderson, C.L.; Muller, M.J.; Kononoff, P.J.; Fernando, S.C. Rumen bacterial community composition in Holstein and Jersey cows is different under same dietary condition and is not affected by sampling method. Front. Microbiol. 2016, 7, 1206. [Google Scholar] [CrossRef] [PubMed]
- Calamari, L.; Morera, P.; Bani, P.; Minuti, A.; Basirico, L.; Vitali, A.; Bernabucci, U. Effect of hot season on blood parameters, fecal fermentative parameters, and occurrence of Clostridium tyrobutyricum spores in feces of lactating dairy cows. J. Dairy Sci. 2018, 101, 4437–4447. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Khafipour, E.; Krause, D.O.; Kroeker, A.; Rodriguez-Lecompte, J.C.; Gozho, G.N.; Plaizier, J.C. Effects of subacute ruminal acidosis challenges on fermentation and endotoxins in the rumen and hindgut of dairy cows. J. Dairy Sci. 2012, 95, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Sato, H. Fecal acetone and isopropanol levels of wild sika deer (Cervus nippon yesoensis) in spring and winter. Grassl. Sci. 2010, 56, 56–58. [Google Scholar] [CrossRef]
- Sato, H.; Nakajima, J. Fecal ammonia, urea, volatile fatty acid and lactate levels in dairy cows and their pathophysiological significance during diarrhea. Anim. Sci. J. 2005, 76, 595–599. [Google Scholar] [CrossRef]
- Russell, J.B. The importance of pH in the regulation of ruminal acetate to propionate ratio and methane production in vitro. J. Dairy Sci. 1998, 81, 3222–3230. [Google Scholar] [CrossRef]
- Peng, L.; He, Z.; Chen, W.; Holzman, I.R.; Lin, J. Effects of butyrate on intestinal barrier function in a Caco-2 cell monolayer model of intestinal barrier. Pediatr. Res. 2007, 61, 37–41. [Google Scholar] [CrossRef]
- Ley, R.E.; Backhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef]
- Pitta, D.W.; Pinchak, W.E.; Indugu, N.; Vecchiarelli, B.; Sinha, R.; Fulford, J.D. Metagenomic analysis of the rumen microbiome of steers with wheat-induced frothy bloat. Front. Microbiol. 2016, 7, 689. [Google Scholar] [CrossRef]
- Kang, S.H.; Evans, P.; Morrison, M.; McSweeney, C. Identification of metabolically active proteobacterial and archaeal communities in the rumen by DNA- and RNA-derived 16S rRNA gene. J. Appl. Microbiol. 2013, 115, 644–653. [Google Scholar] [CrossRef]
- Stanton, T.B.; Canaleparola, E. Treponema bryantii sp. nov., a rumen spirochete that interacts with cellulolytic bacteria. Arch. Microbiol. 1980, 127, 145–156. [Google Scholar] [CrossRef]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef]
- Danielsson, R.; Dicksved, J.; Sun, L.; Gonda, H.; Muller, B.; Schnurer, A.; Bertilsson, J. Methane production in dairy cows correlates with rumen methanogenic and bacterial community structure. Front. Microbiol. 2017, 8, 226. [Google Scholar] [CrossRef]
- Callaway, T.R.; Dowd, S.E.; Edrington, T.S.; Anderson, R.C.; Krueger, N.; Bauer, N.; Kononoff, P.J.; Nisbet, D.J. Evaluation of bacterial diversity in the rumen and feces of cattle fed different levels of dried distillers grains plus solubles using bacterial tag-encoded FLX amplicon pyrosequencing. J. Anim. Sci. 2010, 88, 3977–3983. [Google Scholar] [CrossRef]
- Azad, E.; Derakhshani, H.; Forster, R.J.; Gruninger, R.J.; Acharya, S.; McAllister, T.A.; Khafipour, E. Characterization of the rumen and fecal microbiome in bloated and non-bloated cattle grazing alfalfa pastures and subjected to bloat prevention strategies. Sci. Rep. 2019, 9, 4272. [Google Scholar] [CrossRef]
- Petri, R.M.; Schwaiger, T.; Penner, G.B.; Beauchemin, K.A.; Forster, R.J.; McKinnon, J.J.; McAllister, T.A. Characterization of the core rumen microbiome in cattle during transition from forage to concentrate as well as during and after an acidotic challenge. PLoS ONE 2013, 8, e83424. [Google Scholar] [CrossRef]
- Pitta, D.W.; Pinchak, W.E.; Dowd, S.E.; Osterstock, J.; Gontcharova, V.; Youn, E.; Dorton, K.; Yoon, I.; Min, B.R.; Fulford, J.D.; et al. Rumen bacterial diversity dynamics associated with changing from bermudagrass hay to grazed winter wheat diets. Microb. Ecol. 2010, 59, 511–522. [Google Scholar] [CrossRef]
- Duncan, S.H.; Hold, G.L.; Harmsen, H.J.M.; Stewart, C.S.; Flint, H.J. Growth requirements and fermentation products of Fusobacterium prausnitzii, and a proposal to reclassify it as Faecalibacterium prausnitzii gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 2141–2146. [Google Scholar] [CrossRef]


{kind=link}
| Item 1 | Phase 1 | Phase 2 | Phase 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| H | C | L | H | C | L | H | C | L | |
| Ingredients, % of DM | |||||||||
| Corn | 52.94 | 41.44 | 29.94 | 58.75 | 46.77 | 34.79 | 61.99 | 50.08 | 38.17 |
| Wheat | 7.32 | 5.73 | 4.14 | 7.17 | 5.71 | 4.25 | 8.78 | 7.09 | 5.40 |
| Soybean meal | 9.41 | 7.37 | 5.32 | 9.37 | 7.46 | 5.55 | 8.97 | 7.25 | 5.53 |
| Leymus chinensis | 28.18 | 43.78 | 59.38 | 21.59 | 37.58 | 53.57 | 16.94 | 32.90 | 48.86 |
| Calcium carbonate | 0.72 | 0.56 | 0.41 | 0.78 | 0.62 | 0.46 | 0.83 | 0.67 | 0.51 |
| Sodium bicarbonate | 0.00 | 0.00 | 0.00 | 0.78 | 0.62 | 0.46 | 0.83 | 0.67 | 0.51 |
| Sodium chloride | 0.71 | 0.56 | 0.40 | 0.78 | 0.62 | 0.46 | 0.83 | 0.67 | 0.51 |
| Vitamin–mineral premix 2 | 0.72 | 0.56 | 0.41 | 0.78 | 0.62 | 0.46 | 0.83 | 0.67 | 0.51 |
| Nutrient composition, 3 % of DM | |||||||||
| ME 4, Mcal/kg | 2.71 | 2.53 | 2.35 | 2.77 | 2.58 | 2.40 | 2.82 | 2.64 | 2.46 |
| DPI 5 | 9.41 | 8.32 | 7.23 | 9.49 | 8.45 | 7.41 | 9.54 | 8.55 | 7.56 |
| RDPB 6 | 0.30 | 0.30 | 0.29 | 0.30 | 0.29 | 0.28 | 0.30 | 0.29 | 0.28 |
| NDF | 27.7 | 37.1 | 46.4 | 23.6 | 33.2 | 42.8 | 20.8 | 30.4 | 40.0 |
| ADF | 14.6 | 20.2 | 25.8 | 12.1 | 17.9 | 23.7 | 10.4 | 16.2 | 21.9 |
| Crude fat | 3.46 | 3.25 | 3.05 | 3.56 | 3.34 | 3.13 | 3.61 | 3.40 | 3.19 |
| Starch | 43.7 | 34.8 | 25.9 | 47.6 | 38.5 | 29.3 | 50.8 | 41.6 | 32.3 |
| NFC 7 | 52.0 | 43.7 | 35.4 | 54.9 | 46.7 | 38.6 | 60.3 | 51.6 | 42.9 |
| ME/DPI, Mcal/g | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 |
| Calcium | 0.47 | 0.48 | 0.50 | 0.46 | 0.48 | 0.49 | 0.45 | 0.47 | 0.49 |
| Phosphorus | 0.29 | 0.26 | 0.23 | 0.29 | 0.27 | 0.24 | 0.30 | 0.27 | 0.24 |
| Item 1 | H 2 | C 3 | L 4 | SEM 5 | p-value 6 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HP1 | HP2 | HP3 | CP1 | CP2 | CP3 | LP1 | LP2 | LP3 | Diet | Phase | D × P | ||
| pH value | 5.70 c | 5.30 d | 5.00 d | 6.26 b | 6.19 b | 6.22 b | 6.82 a | 6.83 a | 6.68 a | 0.079 | <0.001 | <0.001 | 0.001 |
| Acetate, mM | 41.2 bc | 48.3 ab | 50.0 a | 42.5 bc | 43.1 abc | 44.7 abc | 30.8 d | 39.1 c | 45.6 abc | 1.599 | <0.001 | <0.001 | 0.005 |
| Propionate, mM | 13.8 bc | 17.2 b | 25.7 a | 13.6 bc | 15.0 bc | 17.1 b | 11.8 c | 14.1 bc | 14.9 bc | 0.973 | <0.001 | <0.001 | <0.001 |
| Isobutyrate, mM | 1.10 bcd | 0.89 d | 1.44 ab | 1.16 bcd | 1.28 bc | 1.70 a | 0.97 bcd | 0.93 d | 1.70 a | 0.068 | <0.001 | <0.001 | 0.025 |
| Butyrate, mM | 8.31 c | 12.0 b | 20.2 a | 7.27 c | 6.74 cd | 8.71 c | 4.49 d | 6.89 cd | 7.42 c | 0.567 | <0.001 | <0.001 | <0.001 |
| Isovalerate, mM | 0.24 ab | 0.25 ab | 0.12 ab | 0.22 ab | 0.18 ab | 0.27 a | 0.08 b | 0.07 b | 0.17 ab | 0.041 | 0.003 | 0.841 | 0.015 |
| Valerate, mM | 0.08 b | 0.43 a | 0.46 a | 0.48 a | 0.49 a | 0.53 a | 0.48 a | 0.55 a | 0.54 a | 0.033 | <0.001 | <0.001 | <0.001 |
| TVFA, mM | 65.8 c | 78.4 b | 96.4 a | 65.3 c | 66.7 c | 72.9 bc | 48.6 d | 63.2 c | 70.1 bc | 2.255 | <0.001 | <0.001 | <0.001 |
| BCVFA, mM | 1.15 | 1.11 | 1.56 | 1.34 | 1.40 | 2.02 | 1.02 | 1.04 | 1.75 | 0.097 | <0.001 | <0.001 | 0.460 |
| A/P | 2.96 a | 2.88 a | 1.82 b | 3.12 a | 2.91 a | 2.65 b | 2.60 a | 2.72 a | 2.77a | 0.123 | 0.005 | <0.001 | <0.001 |
| NH3-N, mg/dL | 3.18 | 2.92 | 2.42 | 2.38 | 2.52 | 2.92 | 2.54 | 2.71 | 2.52 | 0.422 | 0.724 | 0.955 | 0.633 |
| Item 1 | P1 | P2 | P3 | SEM 2 | p-value 3 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H | C | L | H | C | L | H | C | L | Diet | Phase | D × P | ||
| Chao1 | 925.6 | 1212.4 | 1180.9 | 654.8 | 1008.3 | 1002.5 | 637.2 | 985.7 | 890.5 | 59.3 | <0.001 | <0.001 | 0.854 |
| Observed species | 674.3 | 950.6 | 925.0 | 477.6 | 750.0 | 802.4 | 466.8 | 722.4 | 731.9 | 44.6 | <0.001 | <0.001 | 0.891 |
| PD whole tree | 52.75 | 67.33 | 67.54 | 42.68 | 58.80 | 60.51 | 43.38 | 57.57 | 54.45 | 2.62 | <0.001 | <0.000 | 0.724 |
| Shannon index | 6.31 | 7.30 | 7.34 | 5.87 | 7.08 | 7.20 | 5.81 | 6.83 | 7.07 | 0.16 | <0.001 | 0.024 | 0.880 |
| Item 1 | P1 | P2 | P3 | SEM 2 | p-value 3 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H | C | L | H | C | L | H | C | L | Diet | Phase | D × P | ||
| Ruminococcaceae_UCG-005 | 7.66 | 14.52 | 16.69 | 4.77 | 13.83 | 13.50 | 2.09 | 11.13 | 9.76 | 0.976 | 0.001 | <0.001 | 0.509 |
| Prevotella_9 | 20.35 | 1.21 | 0.44 | 21.41 | 3.53 | 0.26 | 21.71 | 9.71 | 0.26 | 7.929 | <0.001 | 0.407 | 0.592 |
| Succinivibrio | 9.48 | 3.85 | 4.41 | 7.68 | 4.87 | 2.94 | 4.17 | 5.25 | 2.34 | 1.408 | 0.008 | 0.249 | 0.248 |
| Rikenellaceae_RC9_gut_group | 0.84 | 5.80 | 8.37 | 1.03 | 5.36 | 8.42 | 1.55 | 2.88 | 6.38 | 0.841 | <0.001 | 0.079 | 0.225 |
| Bacteroides | 1.25 | 5.38 | 8.46 | 1.51 | 4.70 | 6.16 | 0.35 | 1.49 | 3.55 | 0.848 | <0.001 | <0.001 | 0.229 |
| Clostridium_sensu_stricto_1 | 2.96 | 2.53 | 2.57 | 3.36 | 2.70 | 2.17 | 3.47 | 5.58 | 3.64 | 0.879 | 0.537 | 0.060 | 0.471 |
| Ruminococcaceae_UCG-014 | 2.43 | 2.14 | 1.99 | 2.67 | 2.99 | 4.61 | 3.02 | 2.21 | 1.78 | 0.698 | 0.819 | 0.067 | 0.182 |
| Phascolarctobacterium | 2.29 | 1.54 | 1.43 | 3.63 | 3.29 | 2.26 | 3.18 | 2.95 | 2.33 | 0.604 | 0.134 | 0.043 | 0.952 |
| Prevotella_2 | 7.05 | 0.23 | 0.00 | 6.12 | 0.53 | 0.00 | 6.61 | 2.34 | 0.01 | 1.007 | <0.001 | 0.598 | 0.722 |
| Alloprevotella | 1.89 | 1.90 | 0.25 | 2.65 | 1.67 | 1.49 | 5.26 | 2.37 | 1.61 | 0.875 | 0.020 | 0.070 | 0.433 |
| Faecalibacterium | 4.49 | 0.19 | 0.01 | 5.47 | 0.70 | 0.05 | 6.62 | 0.96 | 0.01 | 0.943 | <0.001 | 0.493 | 0.874 |
| Prevotellaceae_UCG-003 | 0.28 b | 7.14 a | 1.09 b | 0.51 b | 1.86 ab | 2.54 ab | 0.71 b | 2.18 ab | 2.72 ab | 1.130 | 0.007 | 0.441 | 0.043 |
| Treponema_2 | 0.02 | 3.29 | 3.58 | 0.00 | 0.20 | 2.27 | 0.01 | 1.43 | 8.18 | 1.666 | 0.007 | 0.185 | 0.250 |
| Christensenellaceae_R-7_group | 0.73 | 1.74 | 2.97 | 0.29 | 1.47 | 2.67 | 0.14 | 0.89 | 3.49 | 0.305 | <0.001 | 0.384 | 0.125 |
| Ruminococcaceae_UCG-013 | 0.17 | 2.59 | 3.15 | 0.08 | 0.48 | 2.59 | 0.01 | 0.43 | 3.18 | 0.558 | <0.001 | 0.148 | 0.292 |
| Eubacterium_coprostanoligenes_group | 0.39 | 2.26 | 2.27 | 0.55 | 0.82 | 2.04 | 0.27 | 0.65 | 2.91 | 0.397 | <0.001 | 0.331 | 0.076 |
| Ruminococcaceae_UCG-010 | 0.34 | 1.92 | 2.87 | 0.19 | 1.11 | 2.26 | 0.02 | 0.66 | 2.33 | 0.288 | <0.001 | 0.025 | 0.513 |
| Romboutsia | 1.50 | 1.37 | 1.97 | 0.49 | 1.05 | 1.46 | 0.71 | 1.55 | 1.44 | 0.253 | 0.007 | 0.027 | 0.373 |
| Turicibacter | 0.67 | 0.55 | 0.97 | 0.32 | 0.73 | 0.95 | 2.42 | 1.89 | 1.92 | 0.280 | 0.620 | <0.001 | 0.266 |
| Anaerostipes | 0.99 | 2.33 | 0.16 | 1.18 | 2.72 | 1.99 | 0.47 | 0.01 | 0.01 | 0.568 | 0.104 | 0.001 | 0.176 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, Q.; Zhu, Y.; Qiu, X.; Gao, C.; Wang, J.; Wang, H.; He, Y.; Rahman, M.A.u.; Cao, B.; Su, H. Dynamic Variations in Fecal Bacterial Community and Fermentation Profile of Holstein Steers in Response to Three Stepwise Density Diets. Animals 2019, 9, 560. https://doi.org/10.3390/ani9080560
Qiu Q, Zhu Y, Qiu X, Gao C, Wang J, Wang H, He Y, Rahman MAu, Cao B, Su H. Dynamic Variations in Fecal Bacterial Community and Fermentation Profile of Holstein Steers in Response to Three Stepwise Density Diets. Animals. 2019; 9(8):560. https://doi.org/10.3390/ani9080560
Chicago/Turabian StyleQiu, Qinghua, Yangxiang Zhu, Xinjun Qiu, Chaoyu Gao, Jingjing Wang, Haibo Wang, Yang He, Muhammad Aziz ur Rahman, Binghai Cao, and Huawei Su. 2019. "Dynamic Variations in Fecal Bacterial Community and Fermentation Profile of Holstein Steers in Response to Three Stepwise Density Diets" Animals 9, no. 8: 560. https://doi.org/10.3390/ani9080560
APA StyleQiu, Q., Zhu, Y., Qiu, X., Gao, C., Wang, J., Wang, H., He, Y., Rahman, M. A. u., Cao, B., & Su, H. (2019). Dynamic Variations in Fecal Bacterial Community and Fermentation Profile of Holstein Steers in Response to Three Stepwise Density Diets. Animals, 9(8), 560. https://doi.org/10.3390/ani9080560