The Usefulness of Retinoic Acid Supplementation during In Vitro Oocyte Maturation for the In Vitro Embryo Production of Livestock: A Review

 ,
,  ,
,  ,
,  ,
,  ,
, 
 ,
, 
Simple Summary
Abstract
1. Introduction
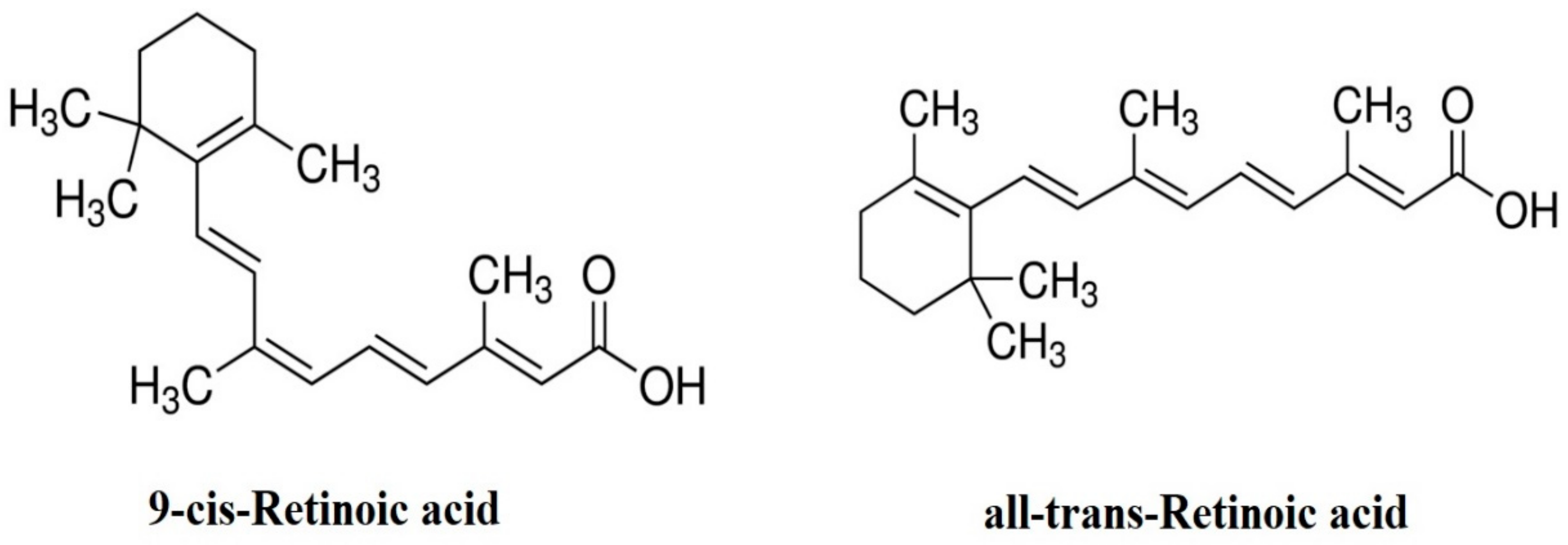
2. Forms of RA
3. Effects of RA and Retinol on Oocyte until Blastocyst Formation
3.1. Effects of RA on Oocyte Maturation Rates
3.2. Effects of RA on Embryo Cleavage Rate
3.3. Effects of RA on Blastocyst Formation
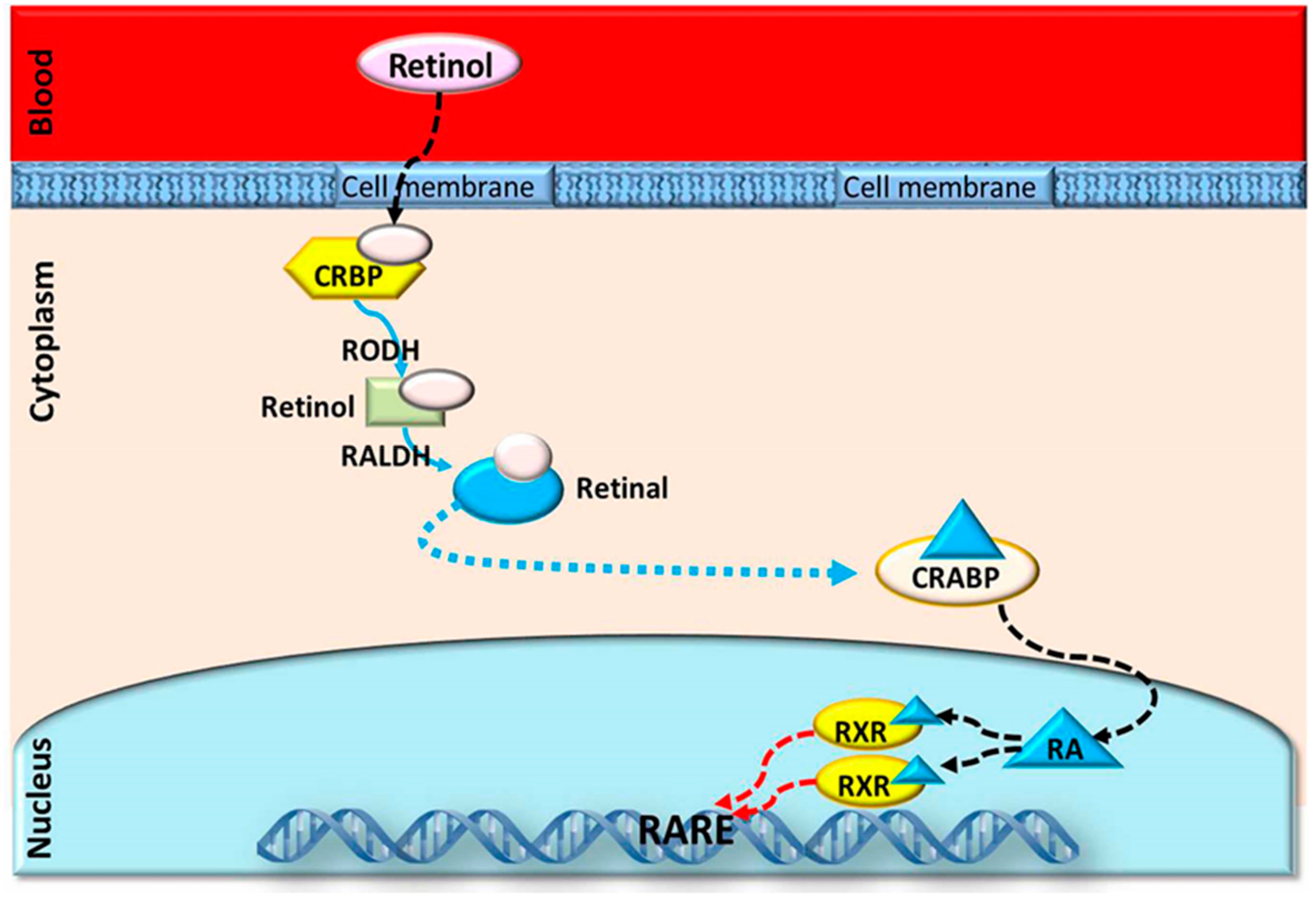
4. Different Modes of Action of RA and Retinol
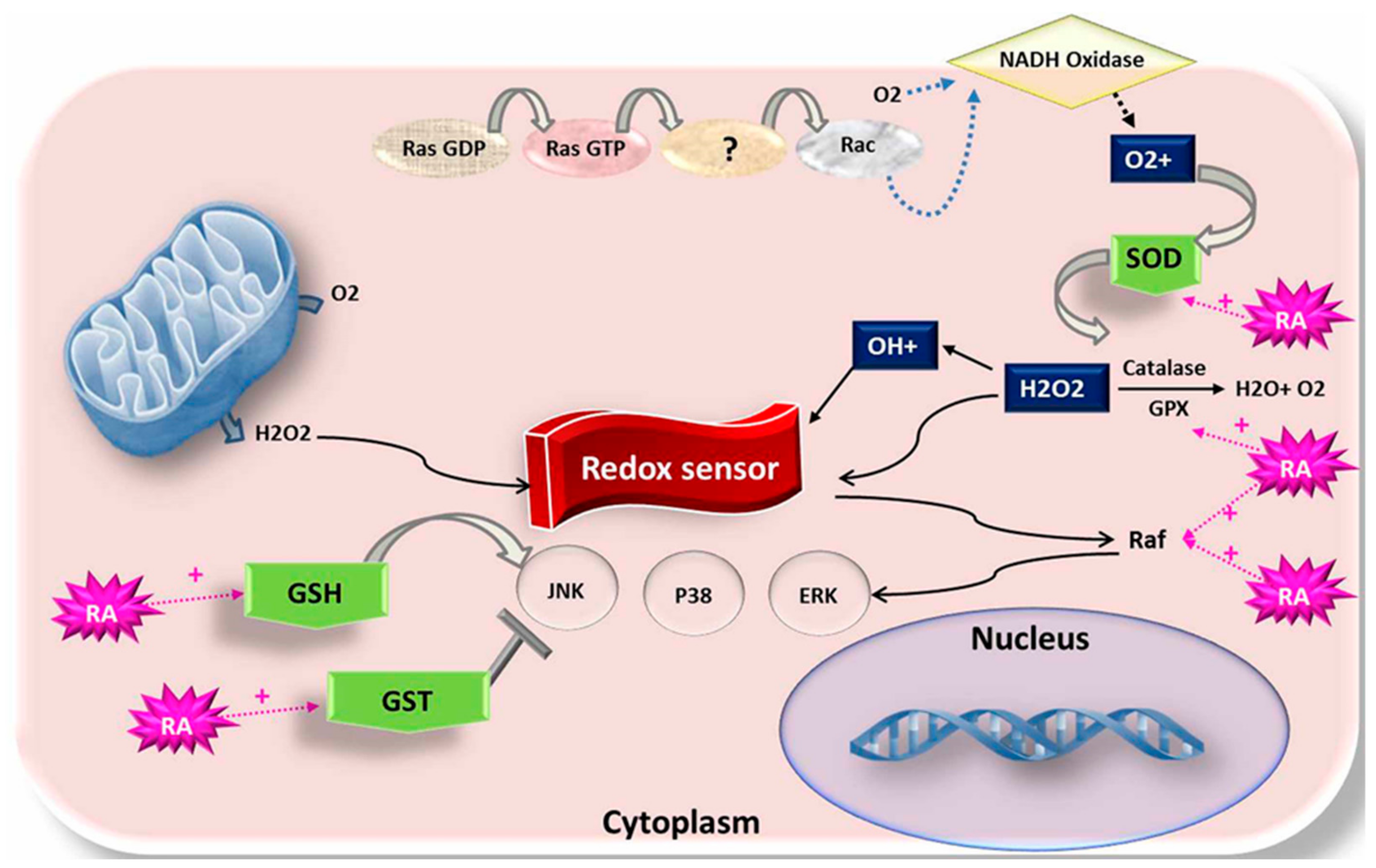
4.1. Impact of RA on Mitochondrial Membrane Potential Activity and ROS Level
4.2. Impact of RA on Apoptosis
5. Effects of RA on Antioxidant-Related Genes
6. Effects of Retinoic Acid on Prostaglandin-Endoperoxide Synthase (PTGS)
7. Conclusion and Future Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pu, Y.; Wang, Z.; Bian, Y.; Zhang, F.; Yang, P.; Li, Y.; Zhang, Y.; Liu, Y.; Fang, F.; Cao, H. All-trans retinoic acid improves goat oocyte nuclear maturation and reduces apoptotic cumulus cells during in vitro maturation. Anim. Sci. J. 2014, 85, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Gómez, E.; Royo, L.J.; Duque, P.; Carneiro, G.; Hidalgo, C.; Goyache, F.; Lorenzo, P.L.; Alvarez, I.; Facal, N.; Díez, C. 9-cis-retinoic acid during in vitro maturation improves development of the bovine oocyte and increases midkine but not IGF-I expression in cumulus-granulosa cells. Mol. Reprod. Dev. 2003, 66, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Duque, P.; Diez, C.; Royo, L.; Lorenzo, P.; Carneiro, G.; Hidalgo, C.; Facal, N.; Gomez, E. Enhancement of developmental capacity of meiotically inhibited bovine oocytes by retinoic acid. Hum. Reprod. 2002, 17, 2706–2714. [Google Scholar] [CrossRef] [PubMed]
- Gad, A.; Hamed, S.A.; Khalifa, M.; Amin, A.; El-Sayed, A.; Swiefy, S.A.; El-Assal, S. Retinoic acid improves maturation rate and upregulates the expression of antioxidant-related genes in in vitro matured buffalo (Bubalus bubalis) oocytes. Int. J. Vet. Sci. Med. 2018, 6, 279–285. [Google Scholar] [CrossRef]
- Cajuday, L.A.; Herrera, A.A.; Duran, D.H. Effect of retinoic acid on the development of water buffalo embryos in vitro. Philipp. J. Vet. Anim. Sci. 2012, 38, 107–116. [Google Scholar]
- Atikuzzaman, M.; Koo, O.J.; Kang, J.T.; Kwon, D.K.; Park, S.J.; Kim, S.J.; Gomez, M.N.L.; Oh, H.J.; Hong, S.G.; Jang, G. The 9-cis retinoic acid signaling pathway and its regulation of prostaglandin-endoperoxide synthase 2 during in vitro maturation of pig cumulus cell-oocyte complexes and effects on parthenogenetic embryo production. Biol. Reprod. 2011, 84, 1272–1281. [Google Scholar] [CrossRef] [PubMed]
- Saadeldin, I.M.; Swelum, A.A.A.; Elsafadi, M.; Mahmood, A.; Yaqoob, S.H.; Alfayez, M.; Alowaimer, A.N. Effects of all-trans retinoic acid on the in vitro maturation of camel (Camelus dromedarius) cumulus-oocyte complexes. J. Reprod. Develop. 2019. [Google Scholar] [CrossRef]
- Ward, S.J.; Morriss-Kay, G.M. The functional basis of tissue-specific retinoic acid signalling in embryos. Semin. Cell. Dev. Biol. 1997, 8, 429–435. [Google Scholar] [CrossRef]
- Abouzaripour, M.; Fathi, F.; Daneshi, E.; Mortezaee, K.; Rezaie, M.J.; Abdi, M. Combined effect of retinoic acid and basic fibroblast growth factor on maturation of mouse oocyte and subsequent fertilization and development. Int. J. Fertil. Steril. 2018, 12, 68. [Google Scholar]
- Brown, J.A.; Eberhardt, D.M.; Schrick, F.N.; Roberts, M.P.; Godkin, J.D. Expression of retinol-binding protein and cellular retinol-binding protein in the bovine ovary. Mol. Reprod. Develop. 2003, 64, 261–269. [Google Scholar] [CrossRef]
- Schweigert, F.; Zucker, H. Concentrations of vitamin A, β-carotene and vitamin E in individual bovine follicles of different quality. Reproduction 1988, 82, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Wellik, D.M.; DeLuca, H.F. Retinol in addition to retinoic acid is required for successful gestation in vitamin A-deficient rats. Biol. Reprod. 1995, 53, 1392–1397. [Google Scholar] [CrossRef] [PubMed]
- Besenfelder, U.; Solti, L.; Seregi, J.; Brem, G. Influence of β-carotene on fertility in rabbits when using embryo transfer programs. Theriogenology 1993, 39, 1093–1109. [Google Scholar] [CrossRef]
- Coffey, M.; Britt, J. Enhancement of sow reproductive performance by β-carotene or vitamin A. J. Animal Sci. 1993, 71, 1198–1202. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, D.M.; Will, W.A.; Godkin, J.D. Retinol administration to superovulated ewes improves in vitro embryonic viability. Biol. Reprod. 1999, 60, 1483–1487. [Google Scholar] [CrossRef] [PubMed]
- Shaw, D.; Farin, P.; Washburn, S.; Britt, J. Effect of retinol palmitate on ovulation rate and embryo quality in superovulated cattle. Theriogenology 1995, 44, 51–58. [Google Scholar] [CrossRef]
- Kedishvili, N.Y. Enzymology of retinoic acid biosynthesis and degradation. J. Lipid Res. 2013, 54, 1744–1760. [Google Scholar] [CrossRef]
- Dyce, P.W.; Tenn, N.; Kidder, G.M. Retinoic acid enhances germ cell differentiation of mouse skin-derived stem cells. J. Ovarian Res. 2018, 11, 19. [Google Scholar] [CrossRef]
- Weiss, W. Requirements of fat-soluble vitamins for dairy cows: A review. J. Dairy Sci. 1998, 81, 2493–2501. [Google Scholar] [CrossRef]
- Morriss-Kay, G.; Wardt, S. Retinoids and mammalian development. Int. Rev. Cytol. 1999, 188, 73–131. [Google Scholar]
- Di Masi, A.; Leboffe, L.; De Marinis, E.; Pagano, F.; Cicconi, L.; Rochette-Egly, C.; Lo-Coco, F.; Ascenzi, P.; Nervi, C. Retinoic acid receptors: From molecular mechanisms to cancer therapy. Mol. Aspects Med. 2015, 41, 1–115. [Google Scholar] [CrossRef] [PubMed]
- Mohan, M.; Malayer, J.; Geisert, R.; Morgan, G. Expression patterns of retinoid X receptors, retinaldehyde dehydrogenase, and peroxisome proliferator activated receptor gamma in bovine preattachment embryos. Biol. Reprod. 2002, 66, 692–700. [Google Scholar] [CrossRef] [PubMed]
- Mohan, M.; Thirumalapura, N.R.; Malayer, J. Bovine cumulus-granulosa cells contain biologically active retinoid receptors that can respond to retinoic acid. Reprod. Biol. Endocrin. 2003, 1, 104. [Google Scholar] [CrossRef] [PubMed]
- Noa, N. Retinoid-binding proteins: Mediators of retinoid action. Biochem. J. 2000, 348, 481–495. [Google Scholar] [CrossRef]
- Mangelsdorf, D.; Umesona, K.; Evans, R. The retinoid receptors. In The Retinoids: Biology, Chemistry and Medicine, 2nd ed.; Sporn, M.B.R.A., Goodman, D.S., Eds.; Raven Press Ltd.: New York, NY, USA, 1994; pp. 319–350. [Google Scholar]
- Conceição, J.C.; Moura, M.T.; Ferreira-Silva, J.C.; Cantanhêde, L.F.; Chaves, R.M.; Lima, P.F.; Oliveira, M.A. Incidence of apoptosis after retinoids and insulin-like growth factor-I (IGF-I) supplementation during goat in vitro embryo production. Zygote 2016, 24, 808–813. [Google Scholar] [CrossRef]
- Nasiri, E.; Mahmoudi, R.; Bahadori, M.H.; Amiri, I. The effect of retinoic acid on in vitro maturation and fertilization rate of mouse germinal vesicle stage oocytes. Cell J. Yakhteh 2011, 13, 19. [Google Scholar]
- Tahaei, L.S.; Eimani, H.; Yazdi, P.E.; Ebrahimi, B.; Fathi, R. Effects of retinoic acid on maturation of immature mouse oocytes in the presence and absence of a granulosa cell co-culture system. J. Assist. Reprod. Genet. 2011, 28, 553–558. [Google Scholar] [CrossRef]
- Alminana, C.; Gil, M.A.; Cuello, C.; Caballero, I.; Roca, J.; Vazquez, J.M.; Gomez, E.; Martinez, E.A. In vitro maturation of porcine oocytes with retinoids improves embryonic development. Reprod. Fertil. Dev. 2008, 20, 483–489. [Google Scholar] [CrossRef]
- Lima, P.F.; Oliveira, M.A.; Santos, M.H.; Reichenbach, H.D.; Weppert, M.; Paula-Lopes, F.F.; Neto, C.C.; Goncalves, P.B. Effect of retinoids and growth factor on in vitro bovine embryos produced under chemically defined conditions. Anim. Reprod. Sci. 2006, 95, 184–192. [Google Scholar] [CrossRef]
- Arhin, S.K.; Lu, J.; Xi, H.; Jin, X. Energy requirements in mammalian oogenesis. Cell. Mol. Biol. (Noisy Le Grand) 2018, 64, 12–19. [Google Scholar] [CrossRef]
- Vahedi, V.; Zeinoaldini, S.; Kohram, H.; Farahavar, A. Retinoic acid effects on nuclear maturation of bovine oocytes in vitro. Afr. J. Biotechnol. 2009, 8, 3974–3978. [Google Scholar]
- Deb, G.K.; Dey, S.R.; Bang, J.I.; Cho, S.J.; Park, H.C.; Lee, J.G.; Kong, I.K. 9-cis retinoic acid improves developmental competence and embryo quality during in vitro maturation of bovine oocytes through the inhibition of oocyte tumor necrosis factor-alpha gene expression. J. Anim. Sci. 2011, 89, 2759–2767. [Google Scholar] [CrossRef] [PubMed]
- Rashedi, M.; Batavani, A.; Najafi, R. Improvement of cortical granules migration and in vitro embryo production of vitrified bovine oocyte by 9-cis retinoic acid. Asian Pac. J. Reprod. 2018, 7, 123–128. [Google Scholar] [CrossRef]
- Old, R.W.; Smith, D.P.; Mason, C.S.; Marklew, S.; Jones, E.A. Effects of retinoic acid on Xenopus embryos. Biochem. Soc. Symp. 1996, 62, 157–174. [Google Scholar] [PubMed]
- Imam, A.; Hoyos, B.; Swenson, C.; Levi, E.; Chua, R.; Viriya, E.; Hammerling, U. Retinoids as ligands and coactivators of protein kinase C alpha. FASEB J. 2001, 15, 28–30. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Kang, J.; Jin, H.; Liu, X.; Li, J.; Li, S.; Lu, Y.; Wang, W.; Yin, X.J. The influence of 9-cis-retinoic acid on nuclear and cytoplasmic maturation and gene expression in canine oocytes during in vitro maturation. Theriogenology 2012, 77, 1198–1205. [Google Scholar] [CrossRef]
- Hattori, M.; Takesue, K.; Nishida, N.; Kato, Y.; Fujihara, N. Inhibitory effect of retinoic acid on the development of immature porcine granulosa cells to mature cells. J. Mol. Endocrin. 2000, 25, 53–61. [Google Scholar] [CrossRef]
- Livingston, T.; Eberhardt, D.; Edwards, J.L.; Godkin, J. Retinol improves bovine embryonic development in vitro. Reprod. Biol. Endocrin. 2004, 2, 83. [Google Scholar] [CrossRef]
- Ahmed, J.; Dutta, D.; Nashiruddullah, N. Comparative efficacy of antioxidant retinol, melatonin, and zinc during in vitro maturation of bovine oocytes under induced heat stress. Turk. J. Vet. Anim. Sci. 2016, 40, 365–373. [Google Scholar] [CrossRef]
- Islam, R.; Deb, G.; Kabir, M.; Miraz, M.F.; Nurun Nahar, T.; M Jahangir Hossain, S.; Paul, S. Supplementation of 9-cis retinoic acid in the in vitro maturation medium increase blastocyst development rate and quality. Asian J. Med. Biol. Res. 2017, 3, 516–520. [Google Scholar] [CrossRef]
- Gomez, E.; Rodriguez, A.; Goyache, F.; Diez, C.; Jose Royo, L.; Moreira, P.N.; Nestor Caamano, J.; Moran, E.; Gutierrez-Adan, A. Retinoid-dependent mRNA expression and poly-(A) contents in bovine oocytes meiotically arrested and/or matured in vitro. Mol. Reprod. Dev. 2004, 69, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Guerin, P.; El Mouatassim, S.; Menezo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, C.O.; Diez, C.; Duque, P.; Facal, N.; Gomez, E. Pregnancies and improved early embryonic development with bovine oocytes matured in vitro with 9-cis-retinoic acid. Reproduction 2003, 125, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.J.; Huang, Y.; Duan, Y.P.; Zhang, M.; Lu, Y.Q.; Lu, K.H. Translocation of active mitochondria during buffalo (Bubalus bubalis) oocytes in vitro maturation, fertilization and preimplantation embryo development. Reprod. Domest. Anim. 2012, 47, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.; Picton, H. Metabolism of follicles and oocytes during growth and maturation. Vitr. Matur. Hum. Oocytes, In in Vitro Maturation of Human Oocytes; Tan, S., Chian, R., Bucket, W., Eds.; Informa UK Ltd.: London, UK, 2006; pp. 15–36. [Google Scholar]
- Amoushahi, M.; Salehnia, M. Reactive oxygen species level, mitochondrial transcription factor A gene expression and succinate dehydrogenase activity in metaphase II oocytes derived from in vitro cultured vitrified mouse ovaries. Vet. Res. Forum 2018, 9, 145–152. [Google Scholar] [PubMed]
- Khazaei, M.; Aghaz, F. Reactive Oxygen Species Generation and Use of Antioxidants during In Vitro Maturation of Oocytes. Int. J. Fertil. Steril. 2017, 11, 63–70. [Google Scholar] [PubMed]
- De Oliveira, M.R. Vitamin A and Retinoids as Mitochondrial Toxicants. Oxid. Med. Cell. Longev. 2015, 2015, 13. [Google Scholar] [CrossRef]
- Lucas, C.G.; Remiao, M.H.; Komninou, E.R.; Domingues, W.B.; Haas, C.; Leon, P.M.; Campos, V.F.; Ourique, A.; Guterres, S.S.; Pohlmann, A.R.; et al. Tretinoin-loaded lipid-core nanocapsules decrease reactive oxygen species levels and improve bovine embryonic development during in vitro oocyte maturation. Reprod. Toxicol. 2015, 58, 131–139. [Google Scholar] [CrossRef]
- Ikeda, S.; Imai, H.; Yamada, M. Apoptosis in cumulus cells during in vitro maturation of bovine cumulus-enclosed oocytes. Reproduction 2003, 125, 369–376. [Google Scholar] [CrossRef]
- Depalo, R.; Nappi, L.; Loverro, G.; Bettocchi, S.; Caruso, M.L.; Valentini, A.M.; Selvaggi, L. Evidence of apoptosis in human primordial and primary follicles. Hum. Reprod. 2003, 18, 2678–2682. [Google Scholar] [CrossRef]
- Ribarov, S.R.; Benov, L.C. Relationship between the hemolytic action of heavy metals and lipid peroxidation. Biochim. Biophys. Acta 1981, 640, 721–726. [Google Scholar] [CrossRef]
- Khafaga, A.F.; El-Sayed, Y.S. All-trans-retinoic acid ameliorates doxorubicin-induced cardiotoxicity: in vivo potential involvement of oxidative stress, inflammation and apoptosis via caspase-3 and p53 down-expression. Naunyn Schmiedeberg’s Arch. Pharmacol. 2018, 391, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Amann, P.M.; Eichmuller, S.B.; Schmidt, J.; Bazhin, A.V. Regulation of gene expression by retinoids. Curr. Med. Chem. 2011, 18, 1405–1412. [Google Scholar] [CrossRef] [PubMed]
- Roth, Z. Symposium review: Reduction in oocyte developmental competence by stress is associated with alterations in mitochondrial function. J. Dairy Sci. 2018, 101, 3642–3654. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.; Nagy, R.; El-Asheeri, A.K.; Eid, L.N. Developmental and molecular responses of buffalo (Bubalus bubalis) cumulus-oocyte complex matured in vitro under heat shock conditions. Zygote 2018, 26, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Ahlemeyer, B.; Bauerbach, E.; Plath, M.; Steuber, M.; Heers, C.; Tegtmeier, F.; Krieglstein, J. Retinoic acid reduces apoptosis and oxidative stress by preservation of SOD protein level. Free Radic. Biol. Med. 2001, 30, 1067–1077. [Google Scholar] [CrossRef]
- Wang, F.; Tian, X.; Zhang, L.; Gao, C.; He, C.; Fu, Y.; Ji, P.; Li, Y.; Li, N.; Liu, G. Beneficial effects of melatonin on in vitro bovine embryonic development are mediated by melatonin receptor 1. J. Pineal Res. 2014, 56, 333–342. [Google Scholar] [CrossRef]
- Mehaisen, G.M.; Saeed, A.M.; Gad, A.; Abass, A.O.; Arafa, M.; El-Sayed, A. Antioxidant Capacity of Melatonin on Preimplantation Development of Fresh and Vitrified Rabbit Embryos: Morphological and Molecular Aspects. PLoS ONE 2015, 10, e0139814. [Google Scholar] [CrossRef]
- Imai, H.; Nakagawa, Y. Biological significance of phospholipid hydroperoxide glutathione peroxidase (PHGPx, GPx4) in mammalian cells. Free Radic. Biol. Med. 2003, 34, 145–169. [Google Scholar] [CrossRef]
- Borchert, A.; Wang, C.C.; Ufer, C.; Schiebel, H.; Savaskan, N.E.; Kuhn, H. The role of phospholipid hydroperoxide glutathione peroxidase isoforms in murine embryogenesis. J. Biol. Chem. 2006, 281, 19655–19664. [Google Scholar] [CrossRef]
- Yant, L.J.; Ran, Q.; Rao, L.; Van Remmen, H.; Shibatani, T.; Belter, J.G.; Motta, L.; Richardson, A.; Prolla, T.A. The selenoprotein GPX4 is essential for mouse development and protects from radiation and oxidative damage insults. Free Radic. Biol. Med. 2003, 34, 496–502. [Google Scholar] [CrossRef]
- Schneider, M.; Forster, H.; Boersma, A.; Seiler, A.; Wehnes, H.; Sinowatz, F.; Neumuller, C.; Deutsch, M.J.; Walch, A.; Hrabe de Angelis, M.; et al. Mitochondrial glutathione peroxidase 4 disruption causes male infertility. FASEB J. 2009, 23, 3233–3242. [Google Scholar] [CrossRef] [PubMed]
- Meseguer, M.; de los Santos, M.J.; Simon, C.; Pellicer, A.; Remohi, J.; Garrido, N. Effect of sperm glutathione peroxidases 1 and 4 on embryo asymmetry and blastocyst quality in oocyte donation cycles. Fertil. Steril. 2006, 86, 1376–1385. [Google Scholar] [CrossRef] [PubMed]
- Chae, S.; Lee, H.K.; Kim, Y.K.; Jung Sim, H.; Ji, Y.; Kim, C.; Ismail, T.; Park, J.W.; Kwon, O.S.; Kang, B.S.; et al. Peroxiredoxin1, a novel regulator of pronephros development, influences retinoic acid and Wnt signaling by controlling ROS levels. Sci. Rep. 2017, 7, 8874. [Google Scholar] [CrossRef] [PubMed]
- Neumann, C.A.; Krause, D.S.; Carman, C.V.; Das, S.; Dubey, D.P.; Abraham, J.L.; Bronson, R.T.; Fujiwara, Y.; Orkin, S.H.; Van Etten, R.A. Essential role for the peroxiredoxin Prdx1 in erythrocyte antioxidant defence and tumour suppression. Nature 2003, 424, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.J.; Kim, S.; Park, J.G.; Jung, I.H.; Lee, M.N.; Jeon, S.; Kweon, H.Y.; Yu, D.Y.; Lee, S.H.; Jang, Y.; et al. Prdx1 (peroxiredoxin 1) deficiency reduces cholesterol efflux via impaired macrophage lipophagic flux. Autophagy 2018, 14, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Farombi, E.O.; Surh, Y.J. Heme oxygenase-1 as a potential therapeutic target for hepatoprotection. J. Biochem. Mol. Biol. 2006, 39, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A.L. The Keap1-Nrf2 pathway: Mechanisms of activation and dysregulation in cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Zenclussen, M.L.; Jensen, F.; Rebelo, S.; El-Mousleh, T.; Casalis, P.A.; Zenclussen, A.C. Heme oxygenase-1 expression in the ovary dictates a proper oocyte ovulation, fertilization, and corpora lutea maintenance. Am. J. Reprod. Immunol. 2012, 67, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Gomez, E.; Caamano, J.N.; Rodriguez, A.; De Frutos, C.; Facal, N.; Diez, C. Bovine early embryonic development and vitamin A. Reprod. Domest. Anim. 2006, 41, 63–71. [Google Scholar] [CrossRef]
- Williams, C.S.; DuBois, R.N. Prostaglandin endoperoxide synthase: Why two isoforms? Am. J. Physiol. 1996, 270, G393–G400. [Google Scholar] [CrossRef] [PubMed]
- Nuttinck, F.; Reinaud, P.; Tricoire, H.; Vigneron, C.; Peynot, N.; Mialot, J.P.; Mermillod, P.; Charpigny, G. Cyclooxygenase-2 is expressed by cumulus cells during oocyte maturation in cattle. Mol. Reprod. Dev. 2002, 61, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Algire, J.E.; Srikandakumar, A.; Guilbault, L.A.; Downey, B.R. Preovulatory changes in follicular prostaglandins and their role in ovulation in cattle. Can. J. Vet. Res. 1992, 56, 6–69. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Items | Species | Effective Dose | Mechanism of Action | References |
|---|---|---|---|---|
| 1. Oocyte maturation | Water buffalo | 5 nM (9-cisRA) |
| [4] |
| Goat | 10 or 100 nM (All-trans RA) |
| [1] | |
| Camel | 20 µM (RA) |
| [7] | |
| Mouse | 2 and 4 µM (All-trans RA) |
| [27] | |
| Mouse | 2 µM (RA) |
| [9] | |
| Mouse | 2 µM (All-trans RA) |
| [28] | |
| Bovine | 5 nM 9-cis RA |
| [33] | |
| Porcine | 5 and 50 nM RA 125 and 1250 nM ROH |
| [29] | |
| Canine | 5 nM 9-cisRA |
| [37] | |
| Porcine | 5 nM 9-cisRA |
| [6] | |
| Bovine | 1 μM (All-trans RA) |
| [32] | |
| Water buffalo | 5 µM RA |
| [5] | |
| Bovine | 5 nM 9-cisRA 500 nM RA in TCM199 is toxic |
| [2] | |
| Bovine | 500 nM RA in potassium simplex optimization medium (KSOM) |
| [30] | |
| Bovine | 5 nM of 9-cisRA |
| [34] | |
| Bovine | 7.5 µM retinol |
| [40] | |
| 2. Embryo cleavage rate | Water buffalo | 5 or 50 nM (9-cisRA) |
| [4] |
| Bovine | 5 nM retinol |
| [39] | |
| Porcine | 5 nM 9-cisRA |
| [6] | |
| Bovine | 7.5 µM retinol |
| [40] | |
| Water buffalo | (1or 5 nM) RA |
| [5] | |
| Bovine | 5 nM RA |
| [41] | |
| Porcine | 125 nM all-trans retinol |
| [29] | |
| 3. Blastocyst formation | Bovine | 5 µM retinol |
| [39] |
| Porcine | 5 nM 9-cisRA |
| [6] | |
| Bovine | 5 nM 9-cisRA |
| [41] | |
| Porcine | 1 µM retinoid |
| [38] | |
| Bovine | 7.5 µM retinol |
| [40] | |
| Bovine | 5 nM 9-cisRA 500 nM RA in TCM199 is toxic |
| [2] | |
| Porcine | 5 nM RA |
| [29] | |
| Bovine | 500 nM 9-cisRA in potassium simplex optimization medium (KSOM) |
| [30] | |
| Water buffalo | 5 µM RA |
| [5] | |
| Bovine | 0.7 μM (All-trans RA) |
| [72] | |
| Bovine | 5 nM (9-cisRA) |
| [44] | |
| Goat | 1000 nM (All-trans RA) |
| [1] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelnour, S.A.; Abd El-Hack, M.E.; Swelum, A.A.-A.; Saadeldin, I.M.; Noreldin, A.E.; Khafaga, A.F.; Al-Mutary, M.G.; Arif, M.; Hussein, E.-S.O.S. The Usefulness of Retinoic Acid Supplementation during In Vitro Oocyte Maturation for the In Vitro Embryo Production of Livestock: A Review. Animals 2019, 9, 561. https://doi.org/10.3390/ani9080561
Abdelnour SA, Abd El-Hack ME, Swelum AA-A, Saadeldin IM, Noreldin AE, Khafaga AF, Al-Mutary MG, Arif M, Hussein E-SOS. The Usefulness of Retinoic Acid Supplementation during In Vitro Oocyte Maturation for the In Vitro Embryo Production of Livestock: A Review. Animals. 2019; 9(8):561. https://doi.org/10.3390/ani9080561
Chicago/Turabian StyleAbdelnour, Sameh A., Mohamed E. Abd El-Hack, Ayman Abdel-Aziz Swelum, Islam M. Saadeldin, Ahmed E. Noreldin, Asmaa F. Khafaga, Mohsen G. Al-Mutary, Muhammad Arif, and El-Sayed O. S. Hussein. 2019. "The Usefulness of Retinoic Acid Supplementation during In Vitro Oocyte Maturation for the In Vitro Embryo Production of Livestock: A Review" Animals 9, no. 8: 561. https://doi.org/10.3390/ani9080561
APA StyleAbdelnour, S. A., Abd El-Hack, M. E., Swelum, A. A.-A., Saadeldin, I. M., Noreldin, A. E., Khafaga, A. F., Al-Mutary, M. G., Arif, M., & Hussein, E.-S. O. S. (2019). The Usefulness of Retinoic Acid Supplementation during In Vitro Oocyte Maturation for the In Vitro Embryo Production of Livestock: A Review. Animals, 9(8), 561. https://doi.org/10.3390/ani9080561