Species-Specific Discrimination of Insect Meals for Aquafeeds by Direct Comparison of Tandem Mass Spectra

 ,
,  , , ,
, , ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Material
2.2. Amino Acid Analysis
2.3. Total Nitrogen
Protein Content
2.4. Proteomics Analysis
2.4.1. Extraction, Solubilization and Quantification of Proteins
Protein Extraction 1:
Protein Extraction 2:
2.4.2. In-Solution Digestion of Proteins
2.4.3. Mass Spectrometry
2.5. Proteomics Bioinformatics and Data Mining
3. Results and Discussion
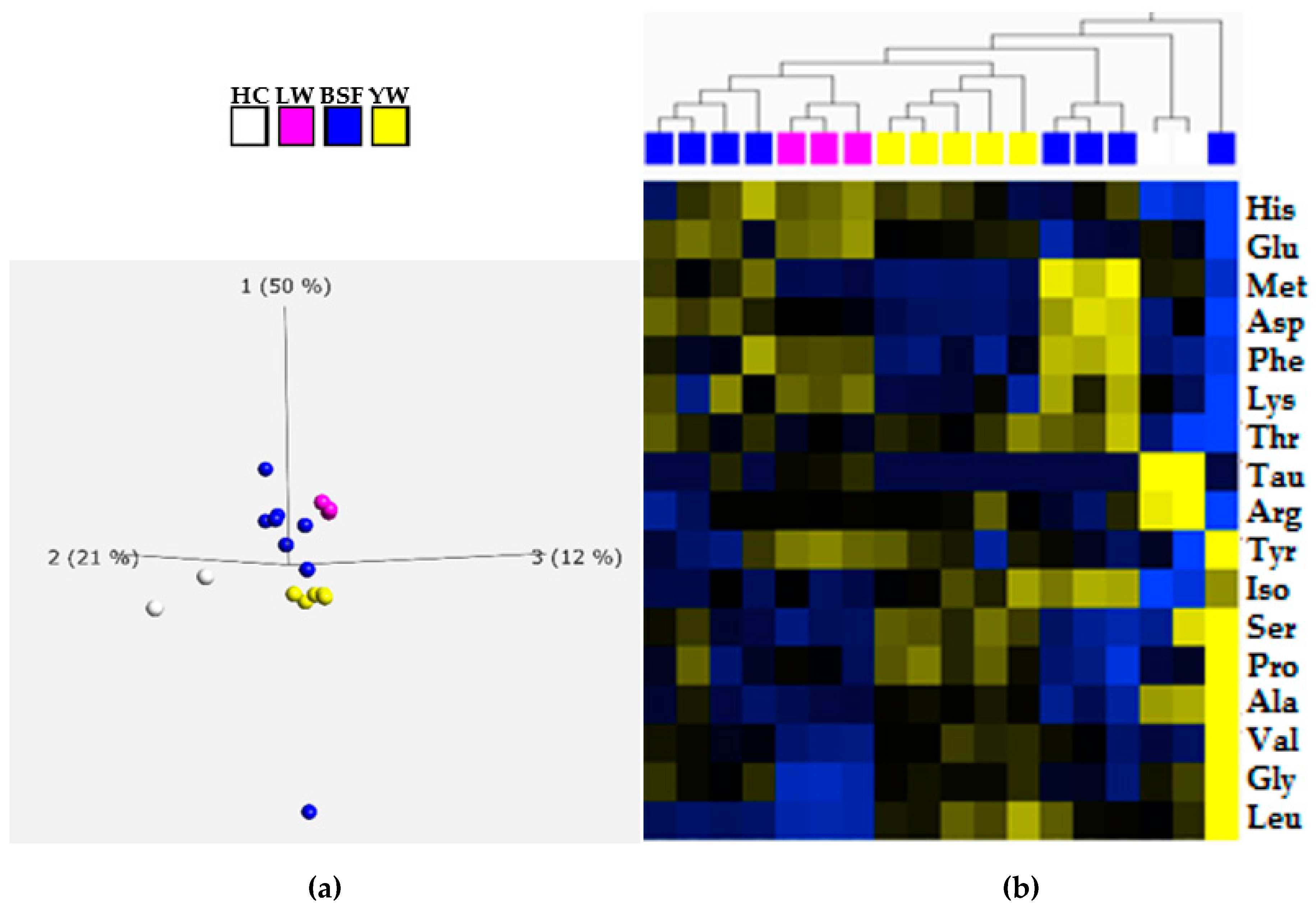
3.1. Quantification of Total Protein and Amino Acid Profiling
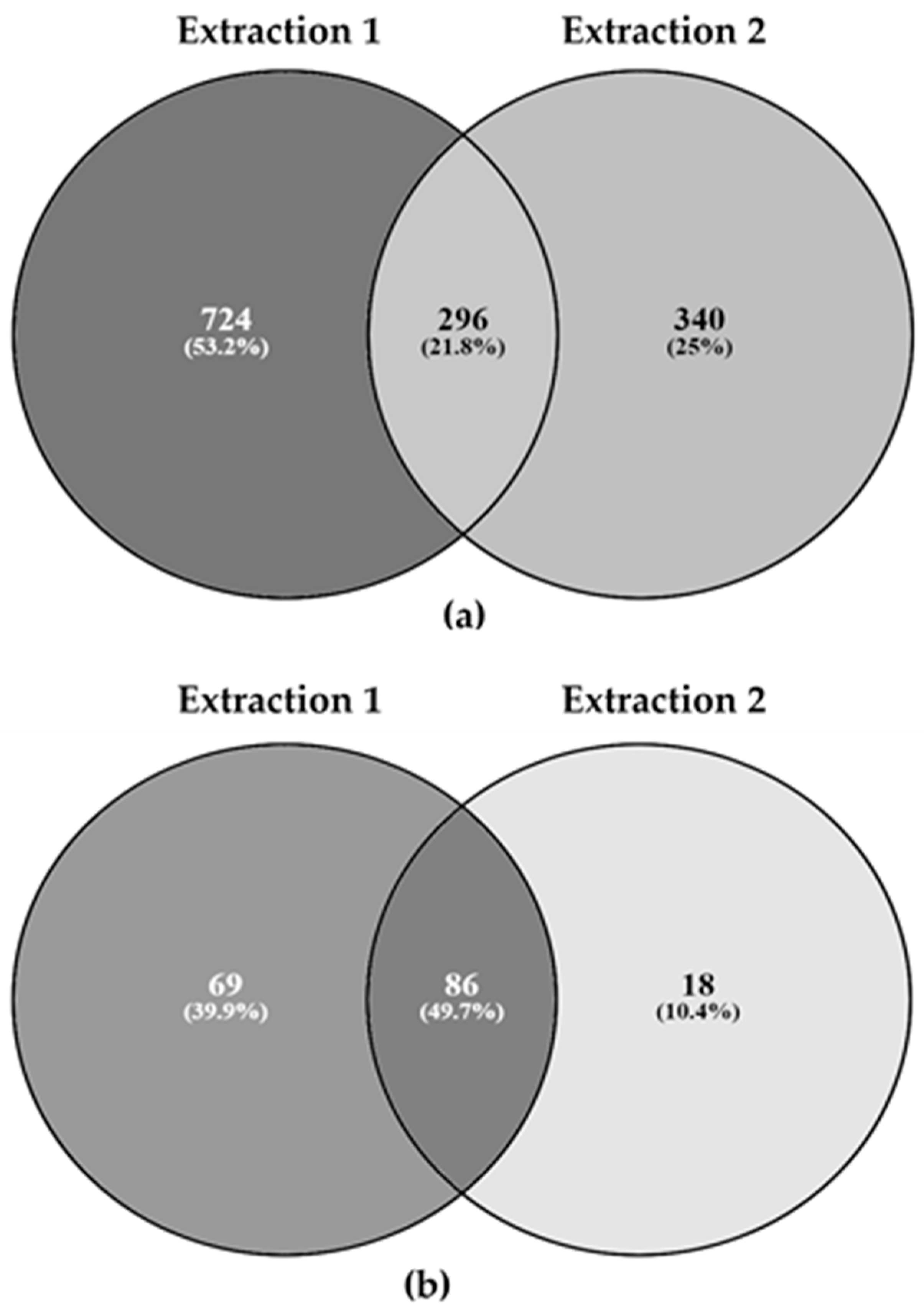
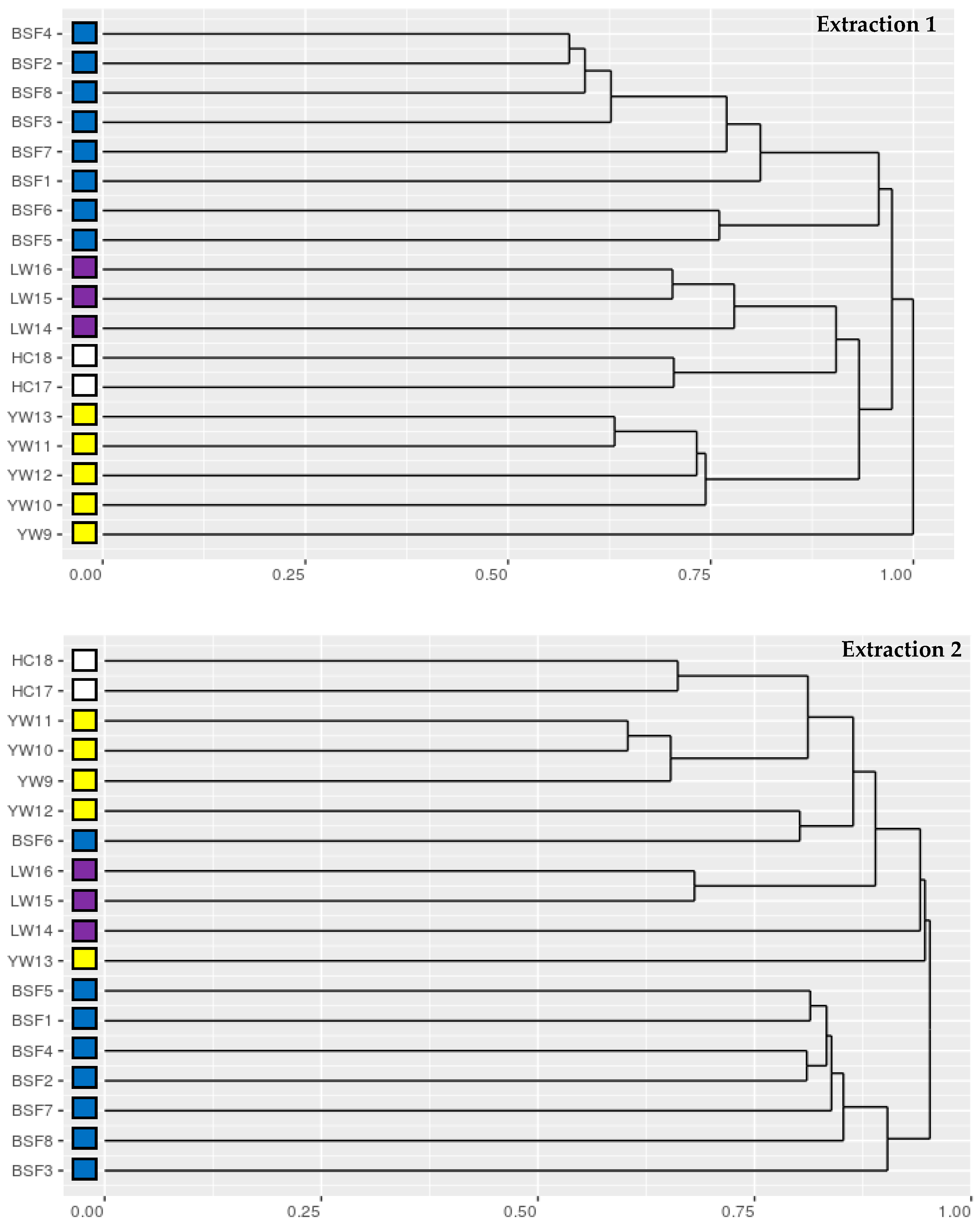
3.2. Proteomics Analysis of Insect Meal Samples
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Froehlich, H.E.; Runge, C.A.; Gentry, R.R.; Gaines, S.D.; Halpern, B.S. Comparative terrestrial feed and land use of an aquaculture-dominant world. PNAS 2018, 115, 5295–5300. [Google Scholar] [CrossRef] [PubMed]
- Tran, G.; Heuzé, V.; Makkar, H.P.S. Insects in fish diets. Anim. Front. 2015, 5, 37–44. [Google Scholar]
- Nogales-Mérida, S.; Gobbi, P.; Józefiak, D.; Mazurkiewicz, J.; Dudek, K.; Rawski, M.; Kierończyk, B.; Józefiak, A. Insect meals in fish nutrition. Rev. Aquacult. 2018, 0. [Google Scholar] [CrossRef]
- Lock, E.R.; Arsiwalla, T.; Waagbø, R. Insect larvae meal as an alternative source of nutrients in the diet of Atlantic salmon (Salmo salar) postsmolt. Aquacult. Nutr. 2016, 22, 1202–1213. [Google Scholar] [CrossRef]
- Belghit, I.; Liland, N.S.; Waagbø, R.; Biancarosa, I.; Pelusio, N.; Li, Y.; Krogdahl, Å.; Lock, E.-J. Potential of insect-based diets for Atlantic salmon (Salmo salar). Aquaculture 2018, 491, 72–81. [Google Scholar] [CrossRef]
- Magalhães, R.; Sánchez-López, A.; Leal, R.S.; Martínez-Llorens, S.; Oliva-Teles, A.; Peres, H. Black soldier fly (Hermetia illucens) pre-pupae meal as a fish meal replacement in diets for European seabass (Dicentrarchus labrax). Aquaculture 2017, 476, 79–85. [Google Scholar] [CrossRef]
- Belghit, I.; Waagbø, R.; Lock, E.-J.; Liland, N.S. Insect-based diets high in lauric acid reduce liver lipids in freshwater Atlantic salmon. Aquacult. Nutr. 2019, 25, 343–357. [Google Scholar] [CrossRef]
- Belghit, I.; Liland, N.S.; Gjesdal, P.; Biancarosa, I.; Menchetti, E.; Li, Y.; Waagbø, R.; Krogdahl, Å.; Lock, E.-J. Black soldier fly larvae meal can replace fish meal in diets of sea-water phase Atlantic salmon (Salmo salar). Aquaculture 2019, 503, 609–619. [Google Scholar] [CrossRef]
- Veldkamp, T.; Duinkerken, G.; Huis, A.; Lakemond, C.M.M.; Ottevanger, E.; Bosch, G.; Boekel, T. Insects as a Sustainable Feed Ingredient in Pig and Poultry Diets: A Feasibility Study = Insecten Als Duurzame Diervoedergrondstof in Varkens- en Pluimveevoeders: Een Haalbaarheidsstudie; Wageningen UR Livestock Research: Lelystad, The Netherlands, 2012; Rapport 638. [Google Scholar]
- Van Huis, A. Potential of Insects as Food and Feed in Assuring Food Security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.X.; Tomberlin, J.K.; Vanlaerhoven, S. Ability of Black Soldier Fly (Diptera: Stratiomyidae) Larvae to Recycle Food Waste. Environ. Entomol. 2015, 44, 406–410. [Google Scholar] [CrossRef]
- Rasinger, J.D.; Marbaix, H.; Dieu, M.; Fumière, O.; Mauro, S.; Palmblad, M.; Raes, M.; Berntssen, M.H.G. Species and tissues specific differentiation of processed animal proteins in aquafeeds using proteomics tools. J. Proteomics 2016, 147, 125–131. [Google Scholar] [CrossRef]
- Marien, A.; Debode, F.; Aerts, C.; Ancion, C.; Francis, F.; Berben, G. Detection of Hermetia illucens by real-time PCR. J. Insects Food Feed 2018, 4, 115–122. [Google Scholar] [CrossRef]
- Debode, F.; Marien, A.; Gérard, A.; Francis, F.; Fumière, O.; Berben, G. Development of real-time PCR tests for the detection of Tenebrio molitor in food and feed. Food Addit. Contam. Part A 2017, 34, 1421–1426. [Google Scholar] [CrossRef] [PubMed]
- Ottoboni, M.; Tretola, M.; Cheli, F.; Marchis, D.; Veys, P.; Baeten, V.; Pinotti, L. Light microscopy with differential staining techniques for the characterisation and discrimination of insects versus marine arthropods processed animal proteins. Food Addit. Contam. Part A 2017, 34, 1377–1383. [Google Scholar] [CrossRef] [PubMed]
- Veys, P.; Baeten, V. Protocol for the isolation of processed animal proteins from insects in feed and their identification by microscopy. Food Control 2018, 92, 496–504. [Google Scholar] [CrossRef]
- Mandrile, L.; Fusaro, I.; Amato, G.; Marchis, D.; Martra, G.; Rossi, A.M. Detection of insect’s meal in compound feed by Near Infrared spectral imaging. Food Chem. 2018, 267, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernández Escámez, P.S.; Gironés, R.; Herman, L.; Koutsoumanis, K.; Lindqvist, R.; et al. Updated quantitative risk assessment (QRA) of the BSE risk posed by processed animal protein (PAP). EFSA J. 2018, 16, 05314. [Google Scholar]
- Lam, H.; Deutsch, E.W.; Eddes, J.S.; Eng, J.K.; Stein, S.E.; Aebersold, R. Building consensus spectral libraries for peptide identification in proteomics. Nat. Met. 2008, 5, 873. [Google Scholar] [CrossRef]
- Ohana, D.; Dalebout, H.; Marissen, R.J.; Wulff, T.; Bergquist, J.; Deelder, A.M.; Palmblad, M. Identification of meat products by shotgun spectral matching. Food Chem. 2016, 203, 28–34. [Google Scholar] [CrossRef]
- Steinhilber, A.E.; Schmidt, F.F.; Naboulsi, W.; Planatscher, H.; Niedzwiecka, A.; Zagon, J.; Braeuning, A.; Lampen, A.; Joos, T.O.; Poetz, O. Mass Spectrometry-Based Immunoassay for the Quantification of Banned Ruminant Processed Animal Proteins in Vegetal Feeds. Anal. Chem. 2018, 90, 4135–4143. [Google Scholar] [CrossRef]
- Lecrenier, M.C.; Marbaix, H.; Dieu, M.; Veys, P.; Saegerman, C.; Raes, M.; Baeten, V. Identification of specific bovine blood biomarkers with a non-targeted approach using HPLC ESI tandem mass spectrometry. Food. Chem. 2016, 213, 417–424. [Google Scholar] [CrossRef]
- Sentandreu, M.A.; Fraser, P.D.; Halket, J.; Patel, R.; Bramley, P.M. A Proteomic-Based Approach for Detection of Chicken in Meat Mixes. J. Proteome Res. 2010, 9, 3374–3383. [Google Scholar] [CrossRef]
- Ortea, I.; O’Connor, G.; Maquet, A. Review on proteomics for food authentication. J. Proteom. 2016, 147, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Palmblad, M.; Deelder, A.M. Molecular phylogenetics by direct comparison of tandem mass spectra. Rapid Commun. Mass Spectrom. 2012, 26, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Espe, M.; Andersen, S.M.; Holen, E.; Rønnestad, V.; Veiseth-Kent, E.; Zerrahn, J.E.; Aksnes, A. Methionine deficiency does not increase polyamine turnover through depletion of hepatic S-denosylmethionine. Br. J. Nutr. 2014, 112, 1274–1285. [Google Scholar] [CrossRef] [PubMed]
- Liland, N.S.; Biancarosa, I.; Araujo, P.; Biemans, D.; Bruckner, C.G.; Waagbø, R.; Torstensen, B.E.; Lock, E.-J. Modulation of nutrient composition of black soldier fly (Hermetia illucens) larvae by feeding seaweed-enriched media. PLOS ONE 2017, 12, 0183188. [Google Scholar] [CrossRef]
- Dumas, J.P.; Lettre de, M. Dumas a, M. Gay-Lussac, sur le procedes de l’analyse organique. Ann. Chim. Phys. 1831, 2, 198–215. [Google Scholar]
- Biancarosa, I.; Espe, M.; Bruckner, C.G.; Heesch, S.; Liland, N.; Waagbø, R.; Torstensen, B.; Lock, E.J. Amino acid composition, protein content, and nitrogen-to-protein conversion factors of 21 seaweed species from Norwegian waters. J. Appl. Phycol. 2016, 29, 1001–1009. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359. [Google Scholar] [CrossRef]
- Distler, U.; Kuharev, J.; Navarro, P.; Tenzer, S. Label-free quantification in ion mobility–enhanced data-independent acquisition proteomics. Nat. Protocols 2016, 11, 795. [Google Scholar] [CrossRef]
- An R Package for Proteomics Data Quality Control. R package version 1.18.1. Available online: https://githubcom/wenbostar/proteoQC (accessed on 2 May 2019).
- R: A Language and Environment for Statistical Computing. Available online: https://wwwR-projectorg/ (accessed on 2 May 2019).
- RStudio: Integrated Development for R. Available online: http://wwwrstudiocom/ (accessed on 2 May 2019).
- Craig, R.; Beavis, R.C. TANDEM: Matching proteins with tandem mass spectra. Bioinformatics 2004, 20, 1466–1467. [Google Scholar] [CrossRef] [PubMed]
- Nesvizhskii, A.I.; Keller, A.; Kolker, E.; Aebersold, R. A statistical model for identifying proteins by tandem mass spectrometry. Anal. Chem. 2003, 75, 4646–4658. [Google Scholar] [CrossRef]
- Rieder, V.; Blank-Landeshammer, B.; Stuhr, M.; Schell, T.; Biß, K.; Kollipara, L.; Meyer, A.; Pfenninger, M.; Westphal, H.; Sickmann, A.; et al. DISMS2: A flexible algorithm for direct proteome- wide distance calculation of LC-MS/MS runs. BMC Bioinformatics 2017, 18, 148. [Google Scholar] [CrossRef]
- Venny. An Interactive Tool for Comparing Lists with Venn’s Diagrams. Available online: http://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 2 May 2019).
- Janssen, R.H.; Vincken, J.-P.; Van den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-Protein Conversion Factors for Three Edible Insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food. Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef] [PubMed]
- Schiavone, A.; De Marco, M.; Martínez, S.; Dabbou, S.; Renna, M.; Madrid, J.; Hernandez, F.; Rotolo, L.; Costa, P.; Gai, F.; et al. Nutritional value of a partially defatted and a highly defatted black soldier fly larvae (Hermetia illucens L.) meal for broiler chickens: apparent nutrient digestibility, apparent metabolizable energy and apparent ileal amino acid digestibility. J. Anim. Sci. 2017, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Espe, M.; Rathore, R.M.; Du, Z.-Y.; Liaset, B.; El-Mowafi, A. Methionine limitation results in increased hepatic FAS activity, higher liver 18:1 to 18:0 fatty acid ratio and hepatic TAG accumulation in Atlantic salmon, Salmo salar. Amino Acids 2010, 39, 449–460. [Google Scholar] [CrossRef]
- Belghit, I.; Skiba-Cassy, S.; Geurden, I.; Dias, K.; Surget, A.; Kaushik, S.; Panserat, S.; Seiliez, I. Dietary methionine availability affects the main factors involved in muscle protein turnover in rainbow trout (Oncorhynchus mykiss). Br. J. Nut. 2014, 112, 493–503. [Google Scholar] [CrossRef]
- Finke, M.D. Complete nutrient composition of commercially raised invertebrates used as food for insectivores. Zoo Biol. 2002, 21, 269–285. [Google Scholar] [CrossRef]
- Finke, M.D. Complete nutrient content of four species of commercially available feeder insects fed enhanced diets during growth. Zoo Biol. 2015, 34, 554–564. [Google Scholar] [CrossRef]
- Salze, G.P.; Davis, D.A. Taurine: A critical nutrient for future fish feeds. Aquaculture 2015, 437, 215–229. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Fish and Shrimp; The National Academies Press: Washington, DC, USA, 2011; p. 363. [Google Scholar]
- Njaa, L.; Utne, F. A comparison of the amino acid composition of fifteen species of whole fish. Fisk. Dir. Skr. Ernaering 1982, 2, 25–33. [Google Scholar]
- Bußler, S.; Rumpold, B.A.; Jander, E.; Rawel, H.M.; Schlüter, O.K. Recovery and techno-functionality of flours and proteins from two edible insect species: Meal worm (Tenebrio molitor) and black soldier fly (Hermetia illucens) larvae. Heliyon 2016, 2, 00218. [Google Scholar] [CrossRef]
- Ericsson, C.; Peredo, I.; Nister, M. Optimized protein extraction from cryopreserved brain tissue samples. Acta Oncol. 2007, 46, 10–20. [Google Scholar] [CrossRef]
- Cilia, M.; Fish, T.; Yang, X.; McLaughlin, M.; Thannhauser, T.W.; Gray, S. A comparison of protein extraction methods suitable for gel-based proteomic studies of aphid proteins. J. Biomol. Tech. 2009, 20, 201–215. [Google Scholar]
- Gatto, L.; Breckels, L.M.; Naake, T.; Gibb, S. Visualization of proteomics data using R and bioconductor. Proteomics 2015, 15, 1375–1389. [Google Scholar] [CrossRef]
- MassIVE Dataset: MSV000083737. Available online: https://massive.ucsd.edu/ProteoSAFe/dataset.jsp?accession=MSV000083737 (accessed on 6 May 2019). [CrossRef]
- Yin, C.; Shen, G.; Guo, D.; Wang, S.; Ma, X.; Xiao, H.; Liu, J.; Zhang, Z.; Liu, Y.; Zhang, Y.; et al. InsectBase: A resource for insect genomes and transcriptomes. Nucleic Acids Res. 2016, 44, D801–D807. [Google Scholar] [CrossRef] [PubMed]
- Wulff, T.; Nielsen, M.E.; Deelder, A.M.; Jessen, F.; Palmblad, M. Authentication of Fish Products by Large-Scale Comparison of Tandem Mass Spectra. J. Proteome Res. 2013, 12, 5253–5259. [Google Scholar] [CrossRef]
- D’Alessandro, A.; Zolla, L. We Are What We Eat: Food Safety and Proteomics. J. Proteome Res. 2012, 11, 26–36. [Google Scholar] [CrossRef]
- Nessen, M.A.; van der Zwaan, D.J.; Grevers, S.; Dalebout, H.; Staats, M.; Kok, E.; Palmblad, M. Authentication of Closely Related Fish and Derived Fish Products Using Tandem Mass Spectrometry and Spectral Library Matching. J. Agric. Food Chem. 2016, 64, 3669–3677. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Samples | Species | Latin Name | Order-Family |
|---|---|---|---|
| BSF1-BSF8 | Black soldier fly | Hermetia illucens | Diptera- Stratiomyidae |
| YW9-YW13 | Yellow mealworm | Tenebrio molitor | Coleoptera-Tenebrionidae |
| LW14-LW16 | Lesser mealworm | Alphitobius diapernius | Coleoptera-Tenebrionidae |
| HC17-HC18 | House cricket | Acheta domesticus | Orthoptera- Gryllidae |
| Species | Taxon Identifier | Number of Proteins (UNIPROT) |
|---|---|---|
| H. illucens | 343,691 | 71 (1) |
| T. molitor | 7067 | 532 (26) |
| A. diaperinus | 27,448 | 28 (0) |
| A. domesticus | 6997 | 131 (4) |
| Species | Ala | Arg | Asp | Glu | Gly | His | Ile | Leu | Lys | Met | Phe | Pro | Ser | Tau | Thr | Tyr | Val |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BSF 1 | 9.5 | 6.2 | 12.4 | 16.0 | 6.7 | 3.8 | 5.2 | 8.8 | 6.9 | 2.0 | 5.2 | 8.8 | 6.0 | <LoD | 5.2 | 8.1 | 7.4 |
| BSF 2 | 7.7 | 6.8 | 12.3 | 14.2 | 7.1 | 4.2 | 5.2 | 8.7 | 7.7 | 2.4 | 6.5 | 7.7 | 5.5 | <LoD | 5.2 | 9.4 | 7.1 |
| BSF 3 | 8.1 | 5.6 | 12.6 | 15.2 | 6.7 | 3.3 | 5.2 | 8.5 | 8.1 | 2.1 | 5.6 | 7.4 | 5.6 | <LoD | 5.2 | 8.1 | 7.4 |
| BSF 4 | 8.2 | 6.7 | 12.4 | 15.5 | 6.7 | 3.9 | 5.5 | 8.8 | 8.8 | 2.1 | 5.2 | 7.0 | 5.5 | <LoD | 5.2 | 7.9 | 7.0 |
| BSF 5 | 8.1 | 6.1 | 13.9 | 13.9 | 6.4 | 3.6 | 5.6 | 9.4 | 8.1 | 2.6 | 6.4 | 6.9 | 5.3 | <LoD | 5.3 | 8.6 | 7.2 |
| BSF 6 | 13.1 | 4.5 | 8.6 | 10.0 | 8.6 | 2.8 | 5.5 | 10.3 | 5.2 | 1.4 | 4.1 | 10.7 | 6.6 | <LoD | 4.5 | 10.7 | 9.3 |
| BSF 7 | 7.1 | 6.9 | 13.6 | 13.8 | 6.0 | 3.8 | 5.5 | 9.3 | 9.0 | 2.7 | 6.4 | 6.2 | 5.2 | <LoD | 5.5 | 7.9 | 6.7 |
| BSF 8 | 7.3 | 6.4 | 13.2 | 12.7 | 6.4 | 3.4 | 5.5 | 9.5 | 9.1 | 2.7 | 6.4 | 7.0 | 5.5 | <LoD | 5.2 | 8.9 | 7.3 |
| YW 9 | 9.0 | 6.9 | 10.8 | 14.7 | 6.7 | 3.7 | 5.5 | 9.6 | 7.5 | 1.7 | 5.1 | 8.2 | 5.9 | <LoD | 5.1 | 9.0 | 7.6 |
| YW 10 | 9.2 | 6.7 | 10.8 | 14.6 | 6.7 | 3.8 | 5.4 | 9.2 | 7.4 | 1.7 | 4.6 | 8.7 | 5.9 | <LoD | 5.1 | 9.0 | 7.2 |
| YW 11 | 9.2 | 6.8 | 10.8 | 14.5 | 6.6 | 3.7 | 5.3 | 9.2 | 7.4 | 1.7 | 4.7 | 8.7 | 6.1 | <LoD | 5.3 | 9.5 | 7.4 |
| YW 12 | 9.0 | 6.7 | 11.0 | 15.0 | 6.9 | 3.3 | 5.6 | 10.0 | 6.7 | 1.8 | 5.2 | 8.1 | 6.0 | <LoD | 5.4 | 8.8 | 7.5 |
| YW 13 | 9.5 | 7.6 | 10.8 | 15.1 | 6.8 | 3.8 | 5.4 | 9.7 | 8.1 | 1.7 | 4.6 | 8.6 | 6.2 | <LoD | 5.4 | 7.6 | 7.6 |
| LW 14 | 8.1 | 6.7 | 11.4 | 16.0 | 5.6 | 4.0 | 5.1 | 8.4 | 8.6 | 1.8 | 5.6 | 7.2 | 5.3 | 0.3 | 5.1 | 9.3 | 6.5 |
| LW 15 | 8.1 | 6.7 | 11.6 | 15.8 | 5.6 | 4.0 | 5.1 | 8.4 | 8.4 | 1.8 | 5.8 | 7.7 | 5.3 | 0.2 | 5.1 | 9.8 | 6.7 |
| LW 16 | 8.1 | 6.7 | 11.9 | 15.8 | 5.6 | 4.0 | 5.3 | 8.6 | 8.8 | 1.8 | 5.8 | 7.9 | 5.3 | 0.2 | 5.1 | 9.8 | 6.7 |
| HC 17 | 11.1 | 8.7 | 11.7 | 14.3 | 7.0 | 3.0 | 5.0 | 9.3 | 7.2 | 2.1 | 4.6 | 7.6 | 6.5 | 1.0 | 4.8 | 6.5 | 6.7 |
| HC 18 | 10.7 | 8.3 | 10.5 | 14.9 | 6.8 | 2.9 | 4.9 | 9.3 | 7.8 | 2.1 | 4.6 | 7.3 | 5.4 | 0.8 | 5.1 | 8.5 | 7.1 |
| FM * | 6.1 | 5.2 | 9.3 | 13.1 | 6.6 | 2.1 | 5.1 | 6.5 | 10.1 | 3.1 | 3.7 | 4.2 | 4.4 | 1.0 | 3.5 | 2.9 | 4.5 |
| SP * | 3.7 | 5.1 | 11.5 | 20.7 | 3.7 | 1.3 | 4.0 | 6.1 | 3.7 | 1.4 | 3.4 | 5.1 | 5.3 | - | 3.7 | 3.1 | 4.5 |
| Abbr | CP | TP | ∆% | N-Prot Factor |
|---|---|---|---|---|
| BSF1 | 72 | 42 | 28 | 4.51 |
| BSF2 | 76 | 31 | 24 | 4.74 |
| BSF3 | 73 | 27 | 27 | 4.59 |
| BSF4 | 73 | 33 | 27 | 4.55 |
| BSF5 | 78 | 36 | 22 | 4.85 |
| BSF6 | 67 | 29 | 33 | 4.21 |
| BSF7 | 78 | 42 | 21 | 4.91 |
| BSF8 | 80 | 44 | 20 | 5.01 |
| YW9 | 76 | 51 | 24 | 4.73 |
| YW10 | 75 | 39 | 25 | 4.67 |
| YW11 | 77 | 38 | 23 | 4.81 |
| YW12 | 78 | 52 | 22 | 4.86 |
| YW13 | 74 | 37 | 26 | 4.64 |
| LW14 | 78 | 43 | 22 | 4.89 |
| LW15 | 81 | 43 | 19 | 5.05 |
| LW16 | 80 | 43 | 20 | 4.98 |
| HC 17 | 73 | 46 | 27 | 4.53 |
| HC18 | 77 | 41 | 23 | 4.80 |
| Species | Extraction 1 | Extraction 2 | ||||||
|---|---|---|---|---|---|---|---|---|
| tSpectra | iSpectra | Peptides | Proteins | tSpectra | iSpectra | Peptides | Proteins | |
| BSF1 | 14,656 | 4 | 4 | 4 | 16,235 | 4 | 4 | 1 |
| BSF2 | 14,620 | 35 | 23 | 10 | 15,712 | 13 | 8 | 2 |
| BSF3 | 14,182 | 6 | 5 | 4 | 15,724 | 12 | 11 | 1 |
| BSF4 | 14,514 | 21 | 15 | 9 | 16,616 | 5 | 4 | 1 |
| BSF5 | 11,671 | 25 | 14 | 10 | 15,992 | 5 | 4 | 2 |
| BSF6 | 11,856 | 73 | 23 | 7 | 11,909 | 4 | 1 | 1 |
| BSF7 | 13,527 | 19 | 9 | 7 | 14,608 | 7 | 5 | 4 |
| BSF8 | 14,499 | 18 | 14 | 8 | 16,458 | 7 | 7 | 3 |
| YW9 | 17,416 | 1 | 1 | 1 | 15,469 | 1483 | 365 | 54 |
| YW10 | 13,671 | 1422 | 406 | 84 | 14,551 | 1256 | 304 | 62 |
| YW11 | 14,343 | 1987 | 548 | 103 | 11,778 | 4 | 2 | 2 |
| YW12 | 13,920 | 2138 | 431 | 69 | 12,986 | 758 | 140 | 33 |
| YW13 | 15,042 | 1627 | 524 | 92 | 16,837 | 958 | 414 | 72 |
| LW14 | 13,955 | 0 | 0 | 0 | 15,074 | 0 | 0 | 0 |
| LW15 | 12,654 | 0 | 0 | 0 | 14,267 | 0 | 0 | 0 |
| LW16 | 13,341 | 0 | 0 | 0 | 14,392 | 0 | 0 | 0 |
| HC17 | 13,732 | 39 | 16 | 5 | 15,327 | 132 | 22 | 3 |
| HC18 | 13,131 | 33 | 11 | 6 | 14,520 | 83 | 14 | 2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belghit, I.; Lock, E.-J.; Fumière, O.; Lecrenier, M.-C.; Renard, P.; Dieu, M.; Berntssen, M.H.G.; Palmblad, M.; Rasinger, J.D. Species-Specific Discrimination of Insect Meals for Aquafeeds by Direct Comparison of Tandem Mass Spectra. Animals 2019, 9, 222. https://doi.org/10.3390/ani9050222
Belghit I, Lock E-J, Fumière O, Lecrenier M-C, Renard P, Dieu M, Berntssen MHG, Palmblad M, Rasinger JD. Species-Specific Discrimination of Insect Meals for Aquafeeds by Direct Comparison of Tandem Mass Spectra. Animals. 2019; 9(5):222. https://doi.org/10.3390/ani9050222
Chicago/Turabian StyleBelghit, Ikram, Erik-Jan Lock, Olivier Fumière, Marie-Caroline Lecrenier, Patricia Renard, Marc Dieu, Marc H. G. Berntssen, Magnus Palmblad, and Josef D. Rasinger. 2019. "Species-Specific Discrimination of Insect Meals for Aquafeeds by Direct Comparison of Tandem Mass Spectra" Animals 9, no. 5: 222. https://doi.org/10.3390/ani9050222
APA StyleBelghit, I., Lock, E.-J., Fumière, O., Lecrenier, M.-C., Renard, P., Dieu, M., Berntssen, M. H. G., Palmblad, M., & Rasinger, J. D. (2019). Species-Specific Discrimination of Insect Meals for Aquafeeds by Direct Comparison of Tandem Mass Spectra. Animals, 9(5), 222. https://doi.org/10.3390/ani9050222