Influence of Feeding Linseed on SCD Activity in Grazing Goat Mammary Glands

 ,
,  , , ,
, , ,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Diets, and Management
2.2. Milk Analysis
2.3. Statistical Analysis
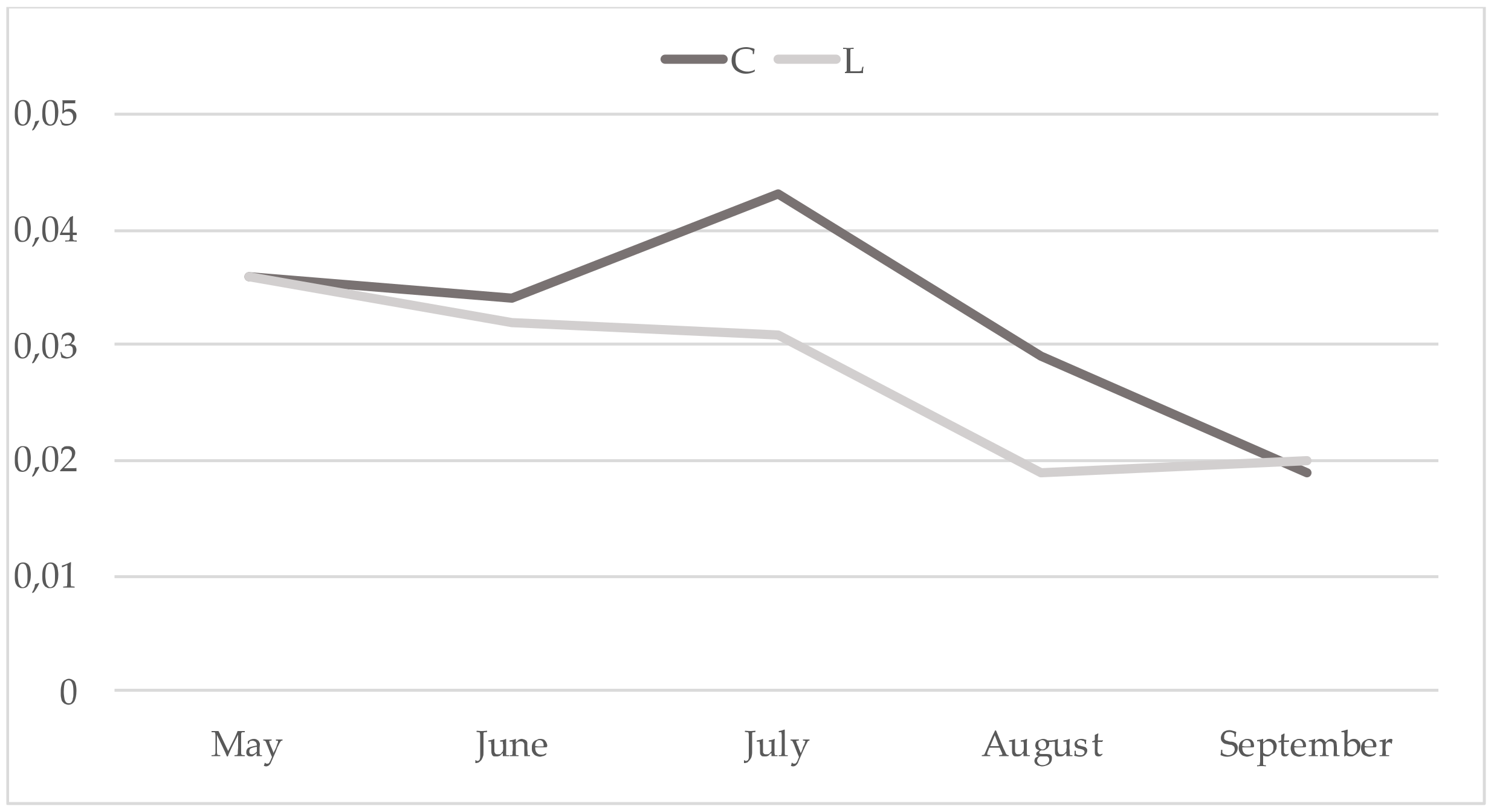
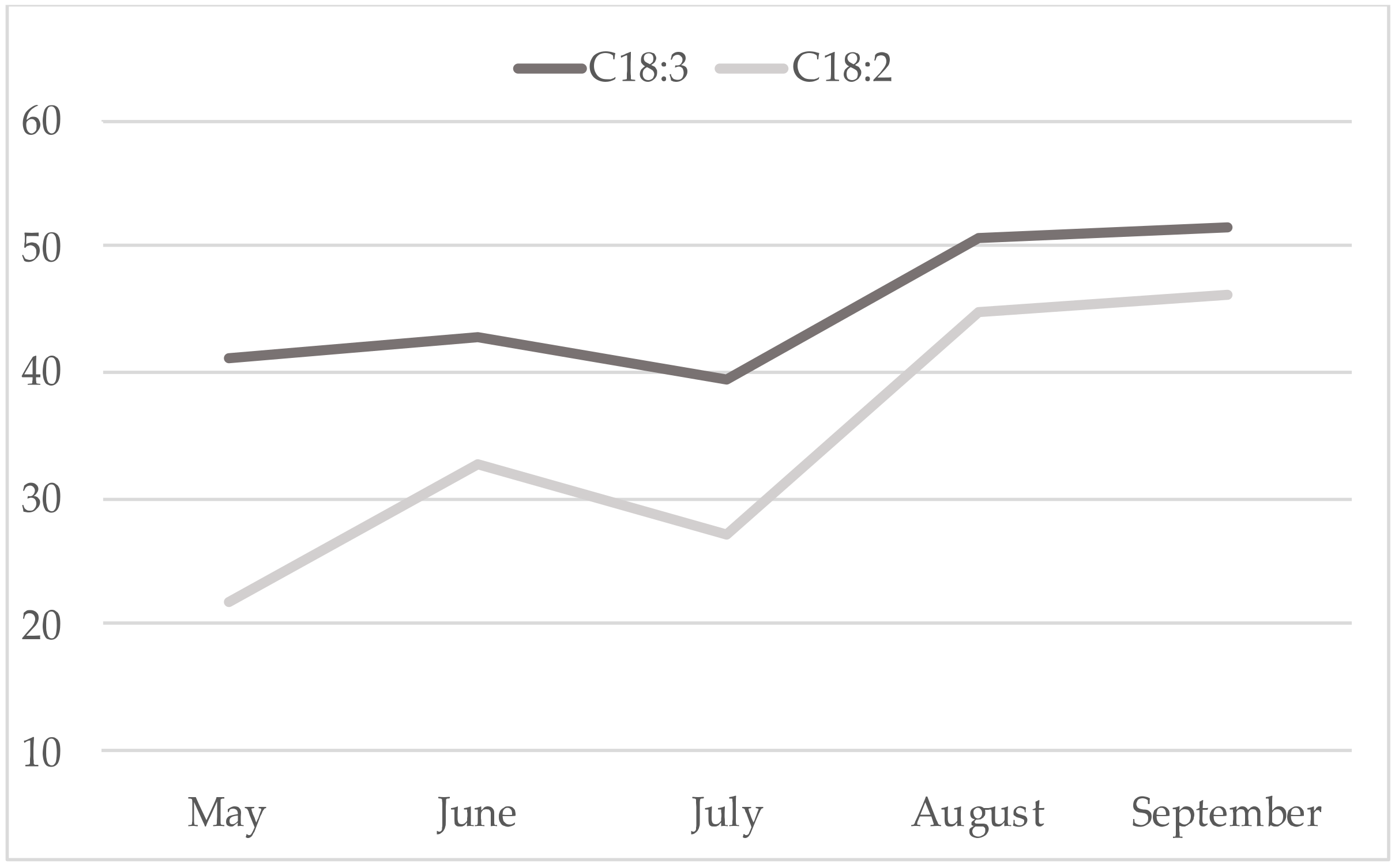
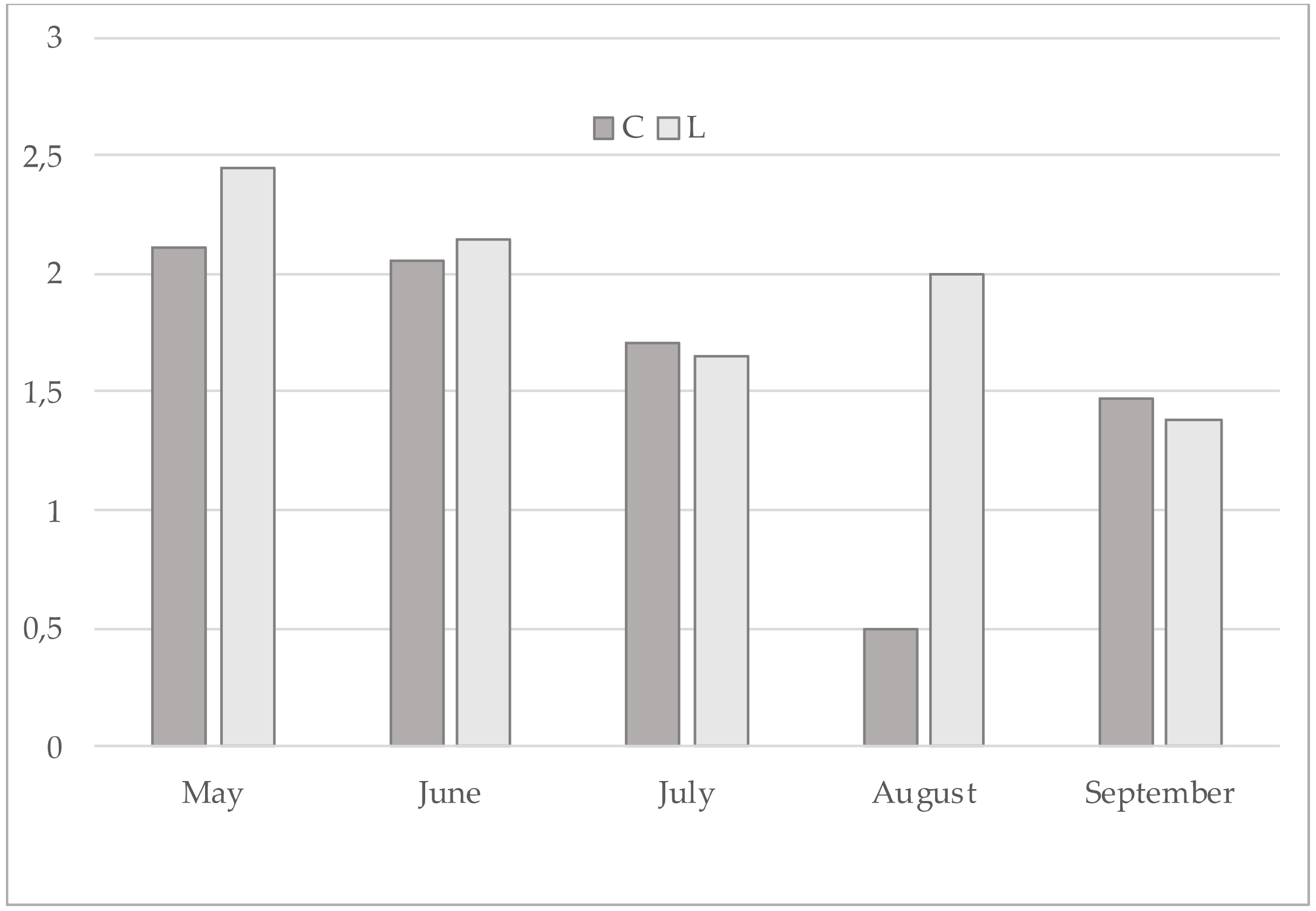
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kamaleldin, A.; Zuki, A.B.; Goh, Y.M.; Sazili, A.Q.; Abubakr, A. Effects of feeding whole linseed on ruminal fatty acid composition and microbial population in goats. Anim. Nutr. 2016, 2, 323–328. [Google Scholar]
- Lee, Y.J.; Jenkins, T.C. Biohydrogenation of linolenic acid to stearic acid by the rumen microbial population yields multiple intermediate conjugated diene isomers. J. Nutr. 2011, 141, 1445–1450. [Google Scholar] [CrossRef] [PubMed]
- Waghmare, J.S. Conjugated linoleic acid: A significant review. Indian J. Sci. Technol. 2013, 2, 8–12. [Google Scholar]
- Tudisco, R.; Morittu, V.M.; Addi, L.; Moniello, G.; Grossi, M.; Musco, N.; Grazioli, R.; Mastellone, V.; Pero, M.E.; Lombardi, P.; et al. Influence of pasture on stearoyl-coa desaturase and miRNA 103 expression in goat milk: Preliminary results. Animals 2019, 9, 606. [Google Scholar] [CrossRef] [PubMed]
- Lock, A.L.; Garnsworthy, P.C. Seasonal variation in milk conjugated linoleic acid and 9-desaturase activity in dairy cows. Livest. Prod. Sci. 2003, 79, 47–59. [Google Scholar] [CrossRef]
- Bernard, L.; Rouel, J.; Leroux, C.; Ferlay, A.; Faulconnier, Y.; Legrand, P.; Chilliard, Y. Mammary lipid metabolism and milk fatty acid secretion in alpine goats fed vegetable lipids. J. Dairy Sci. 88, 1478–1489. [CrossRef]
- Bernard, L.; Leroux, C.; Faulconnier, Y.; Durand, D.; Shingfield, K.J.; Chilliard, Y. Effect of sunflower-seed oil or linseed oil on milk fatty acid secretion and lipogenic gene expression in goats fed hay-based diets. J. Dairy Sci. 2009, 76, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Bernard, L.; Bonnet, M.; Leroux, C.; Shingfield, K.J.; Chilliard, Y. Effect of sunflower-seed oil and linseed oil on tissue lipid metabolism, gene expression, and milk fatty acid secretion in alpine goats fed maizesilage-based diets. J. Dairy Sci. 2009, 92, 6083–6094. [Google Scholar] [CrossRef] [PubMed]
- Bernard, L.; Leroux, C.; Bonnet, M.; Rouel, J.; Martin, P.; Chilliard, Y. Expression and nutritional regulation of lipogenic genes in mammary gland and adipose of lactating goats. J. Dairy Sci. 2005, 72, 250–255. [Google Scholar] [CrossRef]
- Tudisco, R.; Calabrò, S.; Cutrignelli, M.I.; Grossi, M.; Musco, N.; Piccolo, V.; Infascelli, F. Extruded linseed in the diet of grazing goats: Effects on milk Conjugated Linoleic Acid. In Trends in Veterinary; Boiti, C., Ferlazzo, A., Gaiti, A., Pugliese, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 181–185. [Google Scholar]
- Tudisco, R.; Chiofalo, B.; Addi, L.; Lo Presti, V.; Rao, R.; Calabro’, S.; Musco, N.; Grossi, M.; Cutrignelli, M.I.; Mastellone, V.; et al. Effect of hydrogenated palm oil dietary supplementation on milk yield and composition, fatty acids profile and Stearoyl-CoA desaturase expression in goat milk. Small Ruminant Res. 2015, 132, 72–78. [Google Scholar] [CrossRef]
- Legrand, P.; Schmitt, B.; Mourot, J.; Catheline, D.; Chesneau, G.; Mireaux, M. The consumption of food products from linseed-fed animals maintains erythrocyte omega-3 fatty acids in obese humans. Lipids 2010, 45, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Maddock, T.D.; Anderson, V.L.; Lardy, G.P. Using flax in livestock diets. NDSU Extension Service. 2005. Available online: https://www.agmrc.org/media/cms/as1283_E9EE897D14437.pdf (accessed on 2 August 2019).
- Gómez-Cortés, P.; Bach, A.; Luna, P.; Juárez, M.; de la Fuente, M.A. Effects of extruded linseed supplementation on n-3 fatty acids and conjugated linoleic acid in milk and cheese from ewes. J. Dairy Sci. 2008, 92, 4122–4134. [Google Scholar] [CrossRef] [PubMed]
- Nudda, A.; Battacone, M.G.; Usai, G.; Fancellu, S.; Pulina, G. Supplementation with Extruded linseed cake affects concentrations of Conjugated Linoleic Acid and vaccenic acid in goat milk. J. Dairy Sci. 2006, 89, 277–282. [Google Scholar] [CrossRef]
- Renna, M.; Lussiana, C.; D’Agostino, M.; Mimosi, A.; Fortina, R. Extruded linseed supplementation in dairy goat diet: Effects on productive performance and fatty acid profile of bulk milk, fresh and ripened cheese. J. Anim. Vet. Adv. 2013, 12, 1550–1564. [Google Scholar]
- Klir, Z.; Castro-Montoya, J.M.; Novoselec, J.; Molkentin, J.; Domacinovic, M.; Mioc, B.; Dickhoefer, U.; Antunovic, Z. Influence of pumpkin seed cake and extruded linseed on milk production and milk fatty acid profile in Alpine goats. Animal 2017, 11, 1772–1778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and non starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3598. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 19th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2012. [Google Scholar]
- INRA. Alimentation des Ruminants; Jarrige, R., Ed.; INRA Editions: Versailles, France, 1978. [Google Scholar]
- Folch, J.; Lees, M.; Sloane, G.H. A simple method for isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Christie, W.W. A simple procedure of rapid transmethylation of glycerolipids and cholesteryl esters. J. Lipid Res. 1982, 23, 1072–1075. [Google Scholar]
- Chouinard, P.Y.; Corneau, L.; Barbano, D.M.; Metzger, L.E.; Bauman, D.E. Conjugated linoleic acids alter milk fatty acid composition and inhibit milk fat secretion in dairy cows. J. Nutr. 1999, 129, 1579–1584. [Google Scholar] [CrossRef]
- Tudisco, R.; Cutrignelli, M.I.; Calabrò, S.; Piccolo, G.; Bovera, F.; Guglielmelli, A.; Infascelli, F. Influence of organic systems on milk fatty acid profile and CLA in goats. Small Ruminant Res. 2010, 88, 151–155. [Google Scholar] [CrossRef]
- Hara, A.; Radin, N.S. Lipid extraction of tissues with a low-toxicity solvent. Annal. Biochem. 1978, 90, 420–426. [Google Scholar] [CrossRef] [Green Version]
- D’Urso, S.; Cutrignelli, M.I.; Calabrò, S.; Bovera, F.; Tudisco, R.; Piccolo, V.; Infascelli, F. Influence of pasture on fatty acid profile of goat milk. J. Anim. Physiol. Anim. Nutr. 2008, 92, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Tudisco, R.; Calabrò, S.; Cutrignelli, M.I.; Moniello, G.; Grossi, M.; Gonzalez, O.J.; Piccolo, V.; Infascelli, F. Influence of organic systems on Stearoyl-CoA desaturase in goat milk. Small Ruminant Res. 2012, 106, 37–42. [Google Scholar] [CrossRef]
- Tudisco, R.; Grossi, M.; Calabrò, S.; Cutrignelli, M.I.; Musco, N.; Addi, A.; Infascelli, F. Influence of pasture on goat milk fatty acids and Stearoyl-CoA desaturase expression in milk somatic cells. Small Ruminant Res. 2014, 122, 38–43. [Google Scholar] [CrossRef]
- Nudda, A.; Battacone, G.; Atzori, A.S.; Dimauro, C.; Rassu, S.P.G.; Nicolussi, P.; Bonelli, P.; Pulina, G. Effect of extruded linseed supplementation on blood metabolic profile and milk performance of Saanen goats. Animal 2013, 7, 1464–1471. [Google Scholar] [CrossRef] [PubMed]
- Almeida, O.C.; Ferraz, M.V.C.; Susin, I.; Gentil, R.S.; Polizel, D.M.; Ferreira, E.M.; Barroso, J.P.R.; Pires, A.V. Plasma and milk fatty acid profiles in goats fed diets supplemented with oils from soybean, linseed or fish. Small Ruminant Res. 2019, 170, 125–130. [Google Scholar] [CrossRef]
- Rubino, R. L’allevamento caprino; ASSONAPA: Roma, Italy, 1996. [Google Scholar]
- Loor, J.J.; Ferlay, A.; Ollier, A.; Doreau, M.; Chilliard, Y. Relationship among trans and conjugated fatty acids and bovine milk fat yield due to dietary concentrate and linseed oil. J. Dairy Sci. 2005, 88, 726–740. [Google Scholar] [CrossRef]
- Castro, T.; Manso, T.; Jimeno, V.; Del Alamo, M.; Mantecón, A.R. Effects of dietary sources of vegetable fats on performance of dairy ewes and conjugated linoleic acid (CLA) in milk. Small Ruminant Res. 2009, 84, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Di Trana, A.; Cifuni, G.F.; Fedele, V.; Braghieri, A.; Claps, S.; Rubino, R. The grazing system and season affect CLA, ω-3 and trans fatty acid contents in goat milk. Prog. Nutr. 2004, 6, 109–115. [Google Scholar]
- Palmquist, D.L. and Conrad, H.R. High fat rations for dairy cows. Effects on feed intake, milk and fat production, and plasma metabolites. J. Dairy Sci. 1978, 61, 890–901. [Google Scholar] [CrossRef]
- Chilliard, Y.; Ferlay, A.; Leroux, C. A review of nutritional and physiological factors affecting goat milk lipid synthesis and lipolysis. J. Dairy Sci. 2003, 86, 1751–1770. [Google Scholar] [CrossRef]
- Caroprese, M.; Ciliberti, M.G.; Santillo, A.; Marino, R.; Sevi, A.; Albenzio, M. Immune response, productivity and quality of milk from grazing goats as affected by dietary polyunsaturated fatty acid supplementation. Res. Vet. Sci. 2016, 105, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Griinari, J.M.; Corl, B.A.; Lacy, S.H.; Chouinard, P.Y.; Nurmela, K.V.; Bauman, D.E. GrinariJ Conjugated linoleic acid is synthesized endogenously in lactating dairy cows by Delta (9)-desaturase. J. Nutr. 2000, 130, 2285–2291. [Google Scholar] [CrossRef] [PubMed]
- Martinez Marín, A.L.; Gómez-Cortés, P.; Castro, G.G.; Juarez, M.; Alba, L.P.; Hernandez, M.P.; de la Fuente, M.A. Effects of feeding increasing dietary levels of high oleic or regular sunflower or linseed oil on fatty acid profile of goat milk. J. Dairy Sci. 2012, 95, 1942–1955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mele, M.; Contarini, G.; Cercaci, L.; Serra, A.; Buccioni, A.; Conte, P.M.G.; Funaro, A.; Banni, S.; Lercker, G.; Secchiari, P. Enrichment of Pecorino cheese with conjugated linoleic acid by feeding dairy ewes with extruded linseed: Effect on fatty acid and triglycerides composition and on oxidative stability. Int. Dairy J. 2011, 21, 365–372. [Google Scholar] [CrossRef]
- Jeffcoat, R.; James, A.T. The control of stearoyl-CoA desaturase by dietary linoleic acid. FEBS Lett. 1978, 85, 114–118. [Google Scholar] [CrossRef] [Green Version]
- Bernard, L.; Leroux, C.; Chilliard, Y. Expression and nutritional regulation of Stearoyl-CoA desaturase genes in the ruminant mammary gland: Relationship with milk fatty acid composition. In Stearoyl-CoA Desaturase. Genes in Lipid Metabolism; Ntambi, J.M., Ed.; Springer Science+Business Media: New York, NY, USA, 2013; pp. 161–193. [Google Scholar]
- Bragaglio, A.; Braghieri, A.; Napolitano, F.; De Rosa, G.; Riviezzi, A.M.; Surianello, F.; Pacelli, C. Omega-3 supplementation, milk quality and cow immune-competence. Ital. J. Agron. 2015, 10, 9–14. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| Chemical Characteristics | CC* | LC** | Pasture |
|---|---|---|---|
| Crude protein | 180.0 | 182.0 | 164.0 |
| Ether extract | 30.0 | 71.0 | 21.0 |
| NDF | 264.0 | 270.0 | 489.0 |
| ADF | 101.0 | 105.0 | 343.0 |
| ADL | 28.0 | 29.0 | 47.0 |
| UFL***/kg DM | 1.05 | 1.16 | 0.77 |
| Fatty acid profile (% of total FA) | |||
| SFA | 29.0 | 26.6 | 18.2 |
| MUFA | 14.4 | 12.0 | 6.0 |
| PUFA | 56.6 | 61.4 | 75.8 |
| C18:2 | 46.0 | 35.8 | 28.1 |
| C18:3 | 3.2 | 18.9 | 43.0 |
| Milk Characteristics | Yield | Fat | Protein | Lactose | ||||
|---|---|---|---|---|---|---|---|---|
| Group | C | L | C | L | C | L | C | L |
| 1133.6 | 1372.7 | 4.20 | 4.38 | 3.53 | 3.58 | 4.00 | 4.02 | |
| Group effect | ** | ** | NS | NS | ||||
| Sampling effect | ** | ** | NS | ** | ||||
| G x S | NS | NS | NS | NS | ||||
| SEM | 300.1 | 0.621 | 0.246 | 0.24 | ||||
| Fatty Acids Profile | C | L | Group Effect | Sampling Effect | G x S | SEM |
|---|---|---|---|---|---|---|
| C4:0 | 1.42 | 1.53 | * | * | NS | 0.225 |
| C6:0 | 1.91 | 2.07 | NS | * | NS | 0.23 |
| C8:0 | 2.53 | 2.67 | NS | ** | NS | 0.31 |
| C10:0 | 9.00 | 8.76 | NS | ** | NS | 0.96 |
| C11:0 | 0.158 | 0.130 | * | ** | ** | 0.03 |
| C12:0 | 3.57 | 3.26 | NS | ** | NS | 0.53 |
| C14:0 | 10.24 | 8.74 | ** | ** | NS | 0.83 |
| C14:1 cis 9 | 0.33 | 0.24 | ** | ** | ** | 0.04 |
| C15:0 | 0.82 | 0.62 | ** | ** | ** | 0.07 |
| C15:1 | 0.20 | 0.14 | * | ** | ** | 0.02 |
| C16:0 | 28.53 | 23.50 | ** | ** | NS | 1.83 |
| C16:1 cis9 | 0.42 | 0.33 | ** | ** | ** | 0.10 |
| C17:0 | 0.70 | 0.55 | ** | ** | ** | 0.12 |
| C17:1 | 0.22 | 0.14 | ** | NS | NS | 0.03 |
| C18:0 | 13.42 | 16.31 | ** | ** | NS | 1.61 |
| C18:1 cis 9 | 2.09 | 4.79 | ** | NS | NS | 0.61 |
| C18:1 trans 11 | 20.17 | 20.97 | NS | * | NS | 1.92 |
| LA, C18:2 trans 9 trans 12 omega 6 | 0.19 | 0.55 | ** | * | ** | 0.15 |
| C18:2 cis 9 cis 12 omega 6 | 1.36 | 1.45 | NS | ** | ** | 0.37 |
| C20:0 | 0.55 | 0.50 | * | ** | NS | 0.06 |
| ALA, C18:3 omega 3 | 1.16 | 1.41 | NS | ** | ** | 0.18 |
| C22:0 | 0.22 | 0.29 | NS | ** | ** | 0.11 |
| C24:0 | 0.20 | 0.16 | NS | ** | * | 0.05 |
| C22:6 omega 6 | 0.16 | 0.14 | NS | NS | NS | 0.02 |
| cis-9 trans-11 CLA | 0.34 | 0.63 | * | * | ** | 0.16 |
| trans-10 cis-12 CLA | 0.09 | 0.08 | NS | ** | ** | 0.03 |
| SFA | 73.29 | 69.12 | ** | * | NS | 2.52 |
| MUFA | 23.44 | 26.62 | ** | ** | NS | 2.09 |
| PUFA | 3.29 | 4.27 | ** | ** | ** | 0.64 |
| ∑ CLA | 0.43 | 0.71 | * | * | ** | 0.14 |
| PUFA omega 3 | 1.16 | 1.41 | NS | ** | ** | 0.18 |
| PUFA omega 6 | 1.79 | 2.22 | ** | ** | ** | 0.43 |
| C14:1/C14:0 | 0.033 | 0.028 | ** | ** | NS | 0.01 |
| C16:1/C16:0 | 0.015 | 0.013 | NS | ** | * | 0.01 |
| C18:1/C18:0 | 0.161 | 0.299 | ** | * | NS | 0.08 |
| cis-9 trans-11 CLA / C18:1 trans-11 | 0.016 | 0.030 | ** | ** | ** | 0.01 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tudisco, R.; Chiofalo, B.; Lo Presti, V.; Morittu, V.M.; Moniello, G.; Grossi, M.; Musco, N.; Grazioli, R.; Mastellone, V.; Lombardi, P.; et al. Influence of Feeding Linseed on SCD Activity in Grazing Goat Mammary Glands. Animals 2019, 9, 786. https://doi.org/10.3390/ani9100786
Tudisco R, Chiofalo B, Lo Presti V, Morittu VM, Moniello G, Grossi M, Musco N, Grazioli R, Mastellone V, Lombardi P, et al. Influence of Feeding Linseed on SCD Activity in Grazing Goat Mammary Glands. Animals. 2019; 9(10):786. https://doi.org/10.3390/ani9100786
Chicago/Turabian StyleTudisco, Raffaella, Biagina Chiofalo, Vittorio Lo Presti, Valeria Maria Morittu, Giuseppe Moniello, Micaela Grossi, Nadia Musco, Raffaella Grazioli, Vincenzo Mastellone, Pietro Lombardi, and et al. 2019. "Influence of Feeding Linseed on SCD Activity in Grazing Goat Mammary Glands" Animals 9, no. 10: 786. https://doi.org/10.3390/ani9100786
APA StyleTudisco, R., Chiofalo, B., Lo Presti, V., Morittu, V. M., Moniello, G., Grossi, M., Musco, N., Grazioli, R., Mastellone, V., Lombardi, P., & Infascelli, F. (2019). Influence of Feeding Linseed on SCD Activity in Grazing Goat Mammary Glands. Animals, 9(10), 786. https://doi.org/10.3390/ani9100786