Physiological Changes as a Measure of Crustacean Welfare under Different Standardized Stunning Techniques: Cooling and Electroshock

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
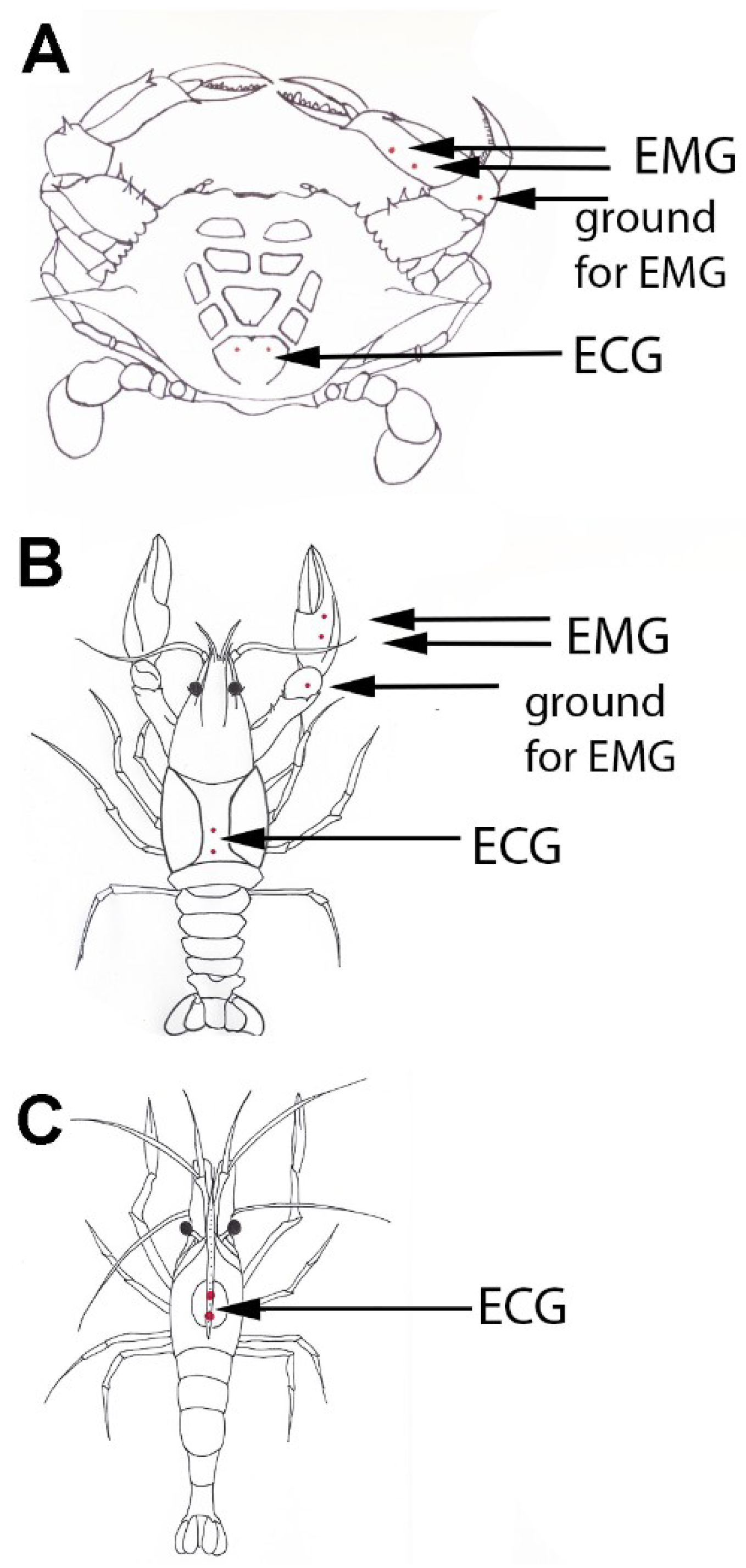
2.2. Electromyograms (EMG) and Electrocardiocrams (ECG)
2.3. Ice Slurry
2.4. Electrical Stunning
2.5. Hemolymph High Pressure Liquid Chromatography (HPLC) Samples
2.6. Statistical Analysis
3. Results
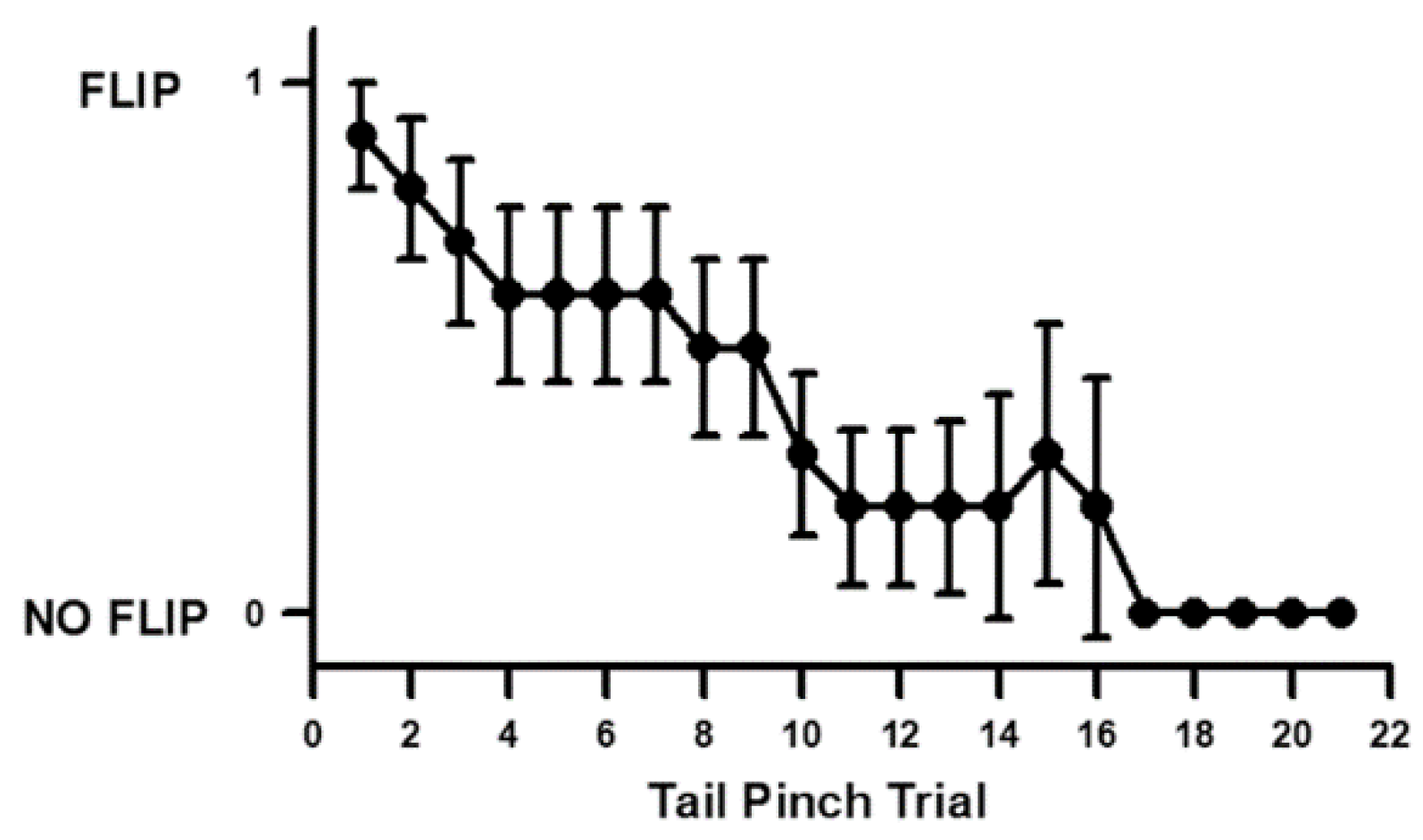
3.1. Habituation Rate in Shrimp Tail Flipping to a Stimulus
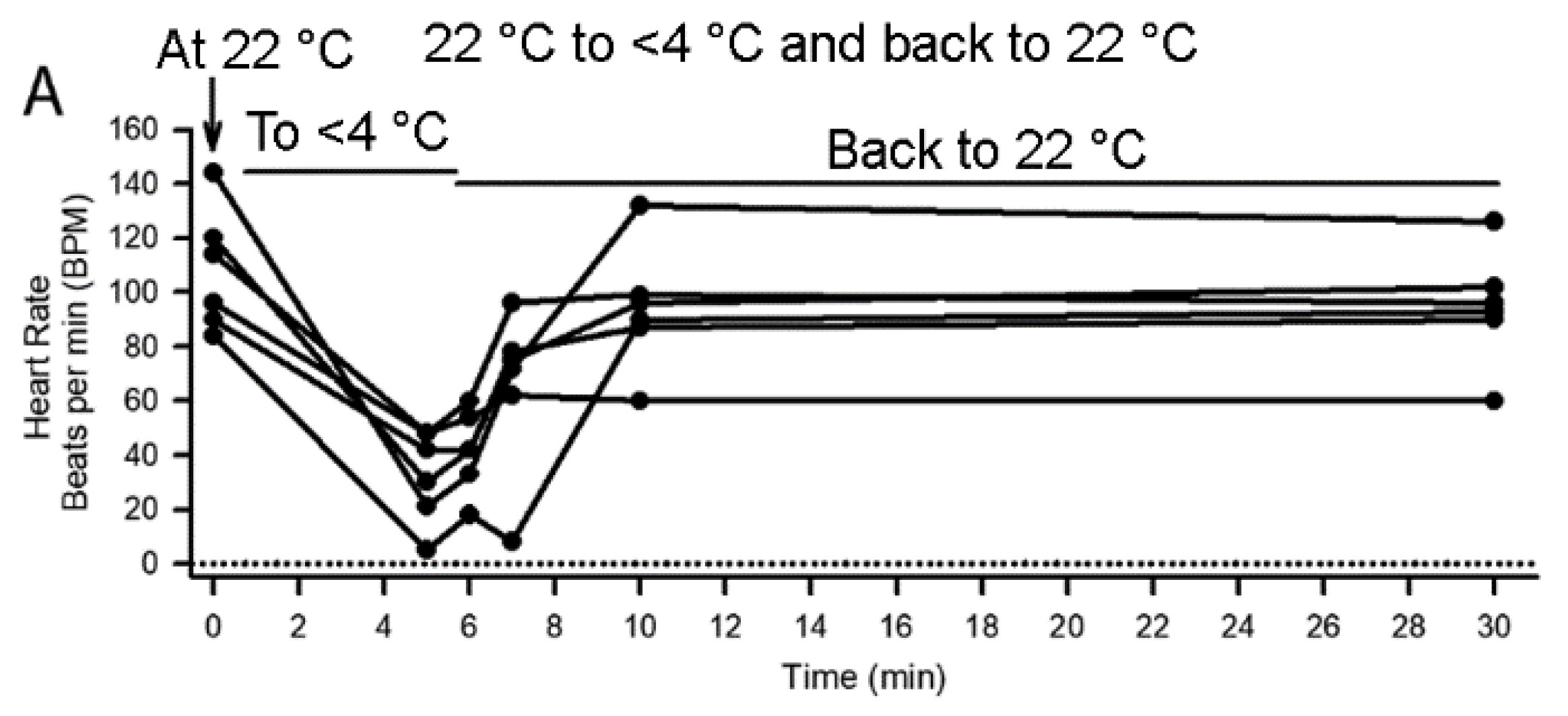
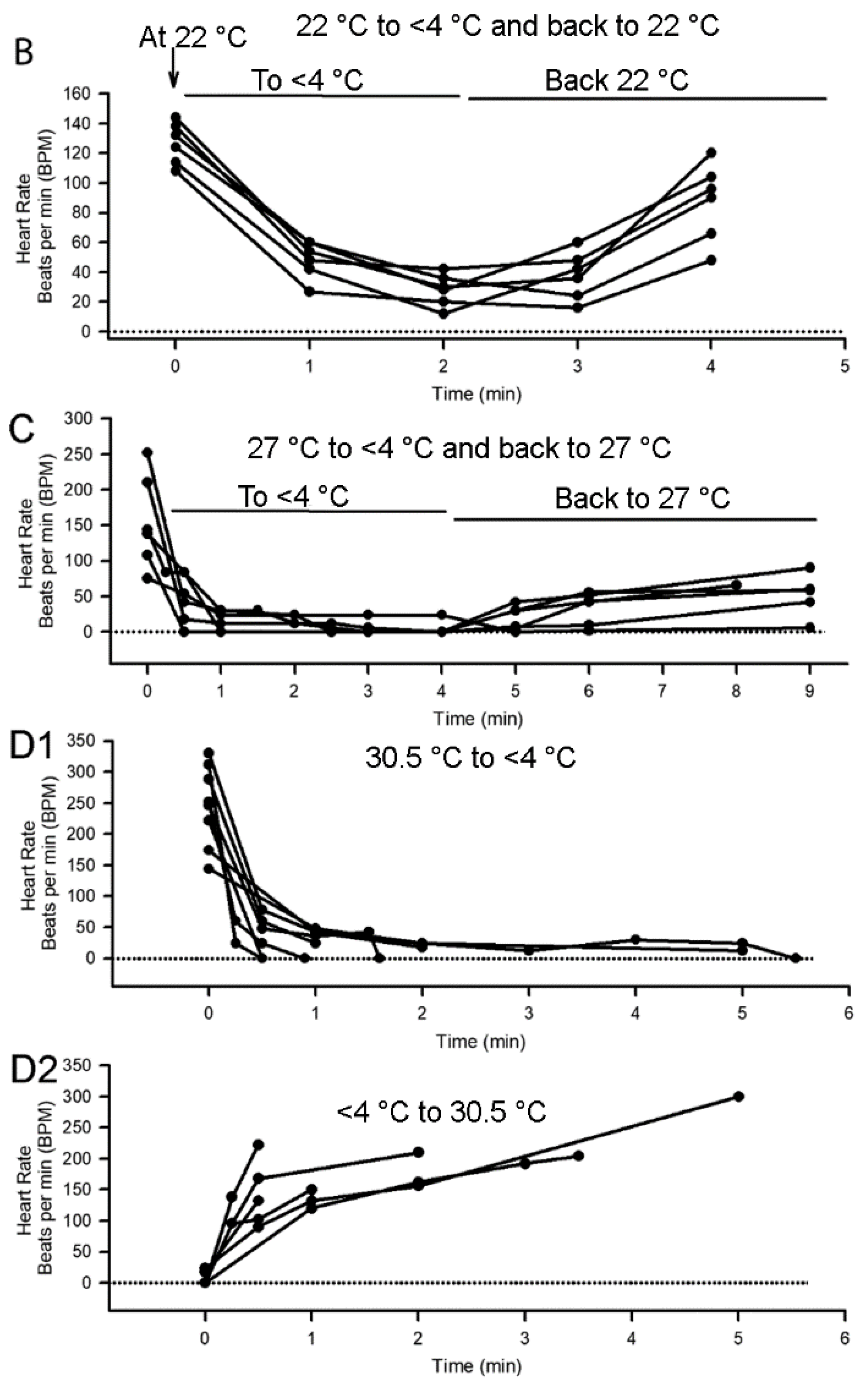
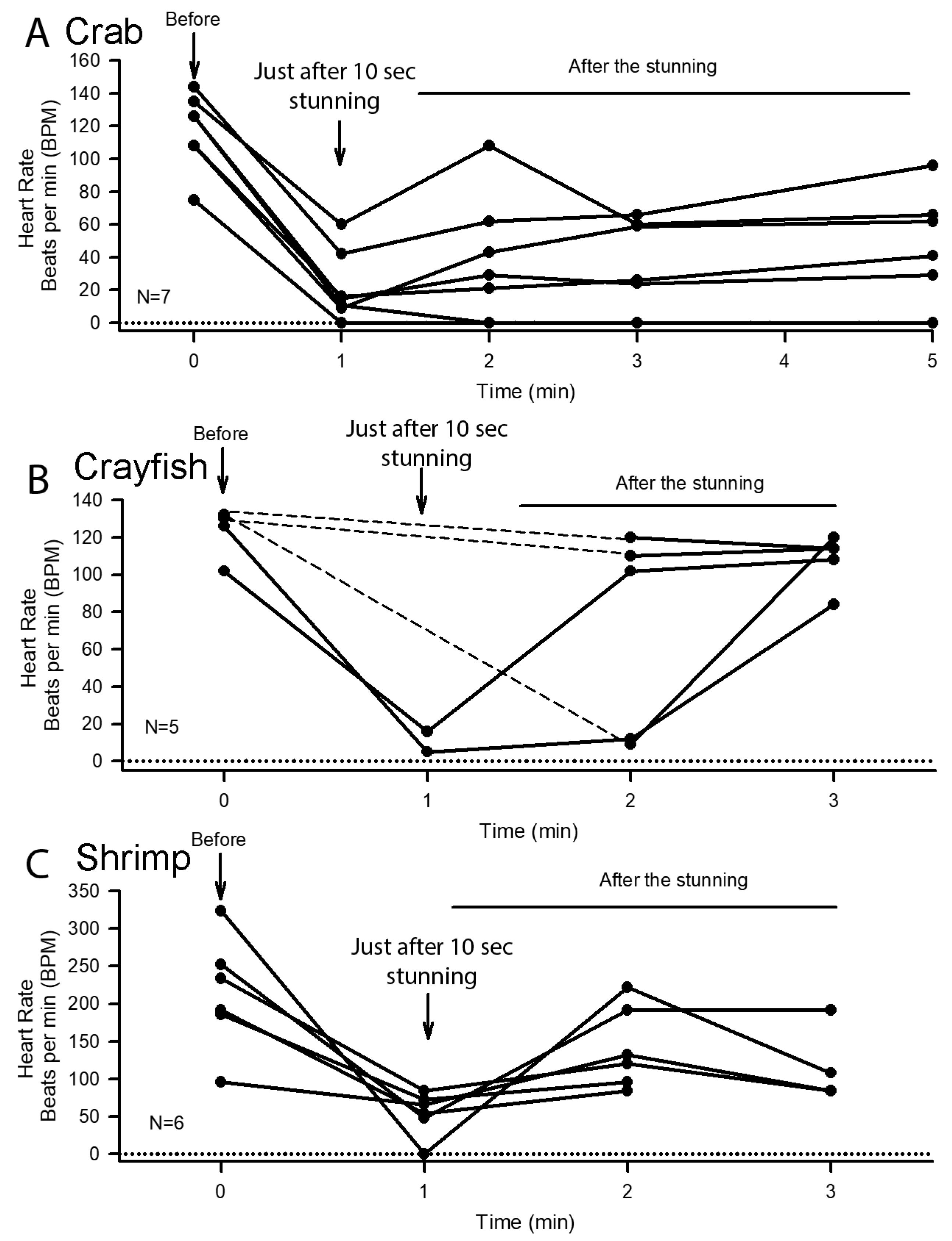
3.2. Effect of Cold Shock on Heart Rate and Response to a Sensory Stimulus
3.3. Heart Rate Changes Triggered by Sensory Stimuli before and after Cold Shock
3.4. Heart Rate Measures before and after Electric Stunning
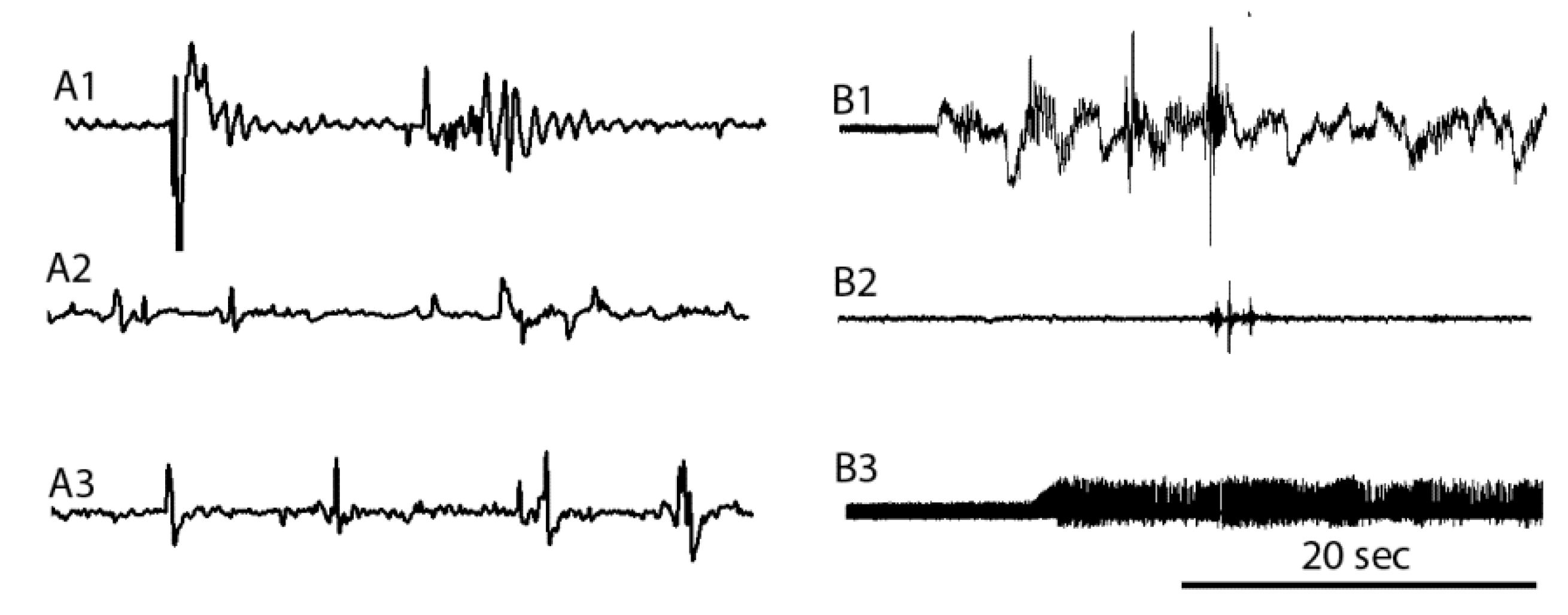
3.5. EMG Measures before, during, and after Cold Shock
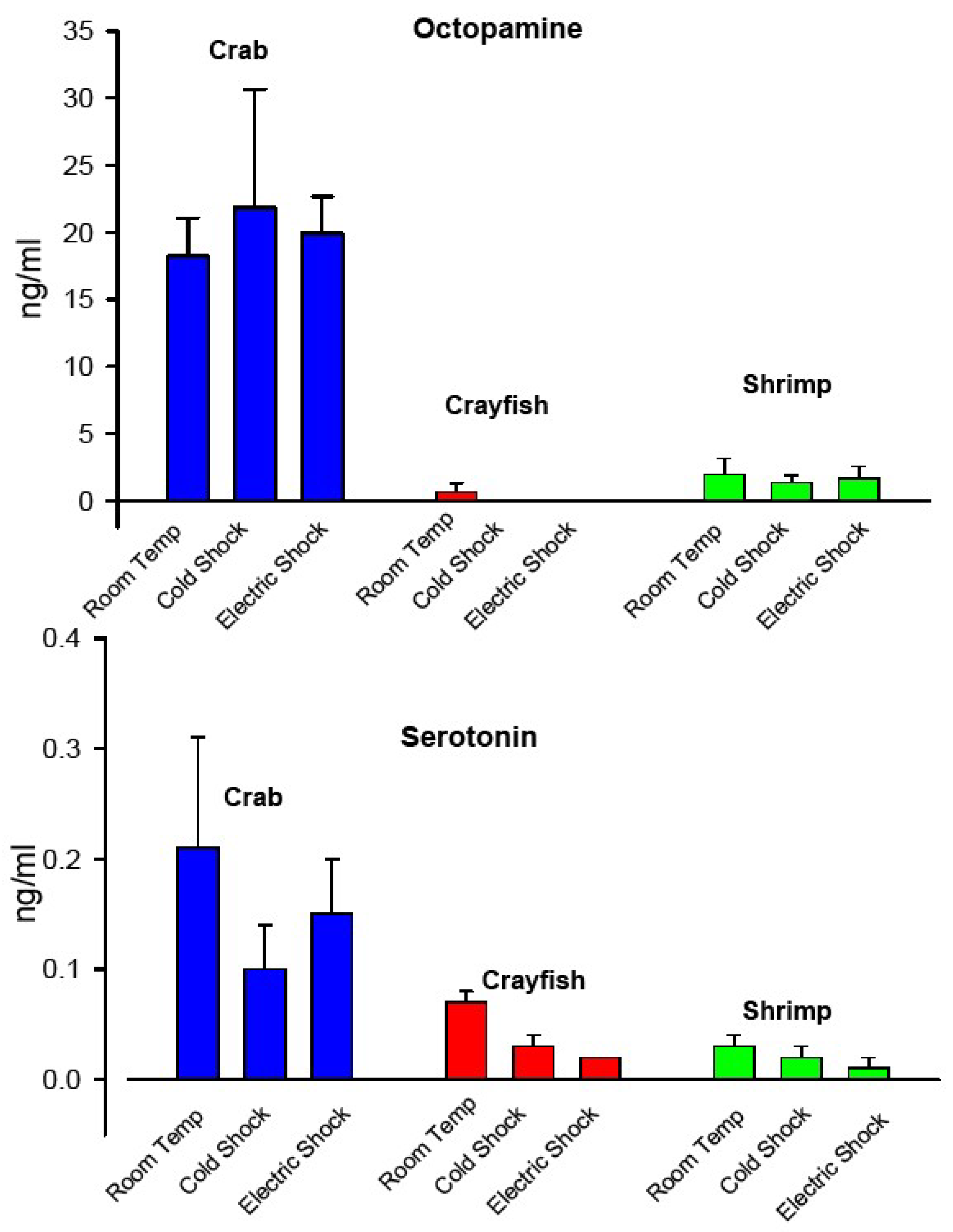
3.6. Modulators in the Hemolymph during Cold Shock and Electric Stunning
3.7. Isolated Shrimp Abdomens Show Tail Flipping
4. Discussion
Animal Welfare Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, M.D.; Roheim, C.A.; Crowder, L.B.; Halpern, B.S.; Turnipseed, M.; Anderson, J.L. Sustainability and global seafood. Science 2010, 327, 784–786. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations 2016. Available online: http://www.fao.org/fishery/topic/16140/en (accessed on 16 August 2018).
- Scientific Panel on Animal Health and Welfare 2005 Aspects of the Biology and Welfare of Animals Used for Experimental and Other Scientific Purposes. EFSA-Q-2004-105. EFSA J. 2005, 292, 1–46.
- Barr, S.; Laming, P.R.; Dick, J.T.A.; Elwood, R.W. Nociception or pain in a decapod crustacean? Anim. Behav. 2008, 75, 745–751. [Google Scholar] [CrossRef]
- Elwood, R.W. Evidence for pain in decapod crustaceans. Anim. Welf. 2012, 21, 23–27. [Google Scholar] [CrossRef]
- Elwood, R.W. Pain and suffering in invertebrates? ILAR 2011, 52, 174–184. [Google Scholar] [CrossRef]
- Roth, B.; Øines, S. Stunning and killing of edible crabs (Cancer pagurus). Anim. Welf. 2010, 19, 287–294. [Google Scholar]
- Roth, B.; Grimsbø, E. Nofima Report 18/2013 Electrical Stunning of Edible Crabs. 2013. Available online: https://nofima.no/filearchive/Rapport%2018-2013.pdf (accessed on 13 September 2018).
- Chung, Y.-S.; Cooper, R.M.; Graff, J.; Cooper, R.L. The acute and chronic effect of low temperature on survival, heart rate and neural function in crayfish (Procambarus clarkii) and prawn (Macrobrachium rosenbergii) species. Open J. Mol. Integr. Physiol. 2012, 2, 75–86. [Google Scholar] [CrossRef]
- Fregin, T.; Bickmeyer, U. Electrophysiological investigation of different methods of anesthesia in lobster and crayfish. PLoS ONE 2016, 11, e0162894. [Google Scholar] [CrossRef] [PubMed]
- Neil, D. The Effect of the Crustastun on Nerve Activity in Crabs and Lobsters. Project Report. University of Glasgow, Glasgow, UK. Available online: crustastun.com/assets/files/neil_2010_glasgow_report.pdf (accessed on 13 September 2018).
- Diggles, B.K. Review of some scientific issues related to crustacean welfare. ICES J. Mar. Sci. 2018, fsy058. [Google Scholar] [CrossRef]
- Stevens, E.D.; Arlinghaus, R.; Browman, H.I.; Cooke, S.J.; Cowx, I.G.; Diggles, B.K. Stress is not pain. Comment on Elwood and Adams (2015) ‘Electric shock causes physiological stress responses in shore crabs, consistent with prediction of pain’. Biol. Lett. 2016, 12. [Google Scholar] [CrossRef] [PubMed]
- Albalat, A.; Sinclair, S.; Laurie, J.; Taylor, A.; Neil, D.M. Targeting the live market: recovery of Norway lobsters Nephrops norvegicus (L.) from trawl-capture as assessed by stress-related parameters and nucleotide breakdown. J. Exp. Mar. Biol. Ecol. 2010, 395, 206–214. [Google Scholar]
- Fotedar, S.; Evans, L. Health management during handling and live transport of crustaceans: A review. J. Invertebr. Pathol. 2011, 106, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Neil, D.M. Ensuring crustacean product quality in the post-harvest phase. J. Invertebr. Pathol. 2012, 110, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Roth, B.; Grimsbø, E. Electrical stunning of edible crabs (Cancer pagurus): From single experiments to commercial practice. Anim. Welf. 2016, 25, 489–497. [Google Scholar] [CrossRef]
- Van DeVis, H.; Kestin, S.; Robb, D.; Oehlenschläger, J.; Lambooij, B.; Münkner, W. Is humane slaughter of fish possible for industry? Aquacult. Res. 2003, 34, 211–220. [Google Scholar] [CrossRef]
- Tania, M.; Kuramotoa, T. Cool-sensitive neurons in the ventral nerve cord of crustaceans. Comp. Biochem. Physiol. 1998, 119, 845–852. [Google Scholar] [CrossRef]
- Tang, L.S.; Goeritz, M.L.; Caplan, J.S.; Taylor, A.L.; Fisek, M.; Marder, E. Precise temperature compensation of phase in a rhythmic motor pattern. PLoS Biol. 2010, 8, e1000469. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.S.; Taylor, A.L.; Rinberg, A.; Marder, E. Robustness of a rhythmic circuit to short- and long-term temperature changes. J. Neurosci. 2012, 32, 10075–20008. [Google Scholar] [CrossRef] [PubMed]
- Marder, E. Variability Compensation, and modulation in neurons and circuits. Proc. Natl. Acad. Sci USA 2011, 108, 15542–15548. [Google Scholar] [CrossRef] [PubMed]
- RSPCA Australia. (nd) Humane Killing and Processing of Crustaceans for Human Consumption. 2016. Available online: http://kb.rspca.org.au/what-is-the-most-humane-way-to-kill-crustaceans-for-human-consumption_625.html (accessed on 23 August 2018).
- Fishcount, UK. Reducing Suffering in Fisheries. Available online: http://fishcount.org.uk/ (accessed on 21 August 2018).
- Rose, J.D.; Arlinghaus, R.; Cooke, S.J.; Diggles, B.K.; Sawynok, W.; Stevens, E.D.; Wynne, C.D.L. Can fish really feel pain? Fish Fish. 2014, 15, 97–133. [Google Scholar] [CrossRef]
- Baker, J.R. Experiments on the humane killing of crabs. J. Mar. Biol. Assoc. UK 1955, 34, 15–24. [Google Scholar] [CrossRef]
- Gunter, G. Painless Killing of crabs and other large crustaceans. Science 1961, 133, 327. [Google Scholar] [CrossRef] [PubMed]
- Puri, S.; Faulkes, Z. Do decapod crustaceans have nociceptors for extreme pH? PLoS ONE 2010, 5, e10244. [Google Scholar] [CrossRef] [PubMed]
- Bierbower, S.M.; Cooper, R.L. The effects of acute carbon dioxide on behavior and physiology in Procambarus clarkii. J. Exp. Zool. 2010, 313A, 484–497. [Google Scholar] [CrossRef] [PubMed]
- McMahon, B.R. Integrated neural and neurohormonal control of respiratory and circulatory function in crustaceans: Is there evidence for an ‘autonomic’ control system? Verh. Dtsch. Zool. Ges. 1995, 88, 87–101. [Google Scholar]
- Orlov, Y. Das Magenganglion des Fluβkrebses, Ein Beitrag zur vergleichenden Histologis des sympathischen Nervensystem. Zeitschrift fur Mikroskopisch–Anatomische Forschung 1927, 8, 67–102. [Google Scholar]
- Shuranova, Z.P.; Burmistrov, Y.M.; Strawn, J.P.; Cooper, R.L. Evidence for an autonomic nervous system in decapod crustaceans. Int. J. Zool. Res. 2006, 2, 242–283. [Google Scholar]
- Zavarzin, A.A. Ocherki po evol utsionnoj gistologii nervnoj sistemy. In Essays on the Evolutionary Histology of the Nervous System; Zavarzin, A.A., Ed.; Izbrannye trudy (Selected Works), Tom III; Izdatel stvo AN SSSR: Leningrad, Russia, 1941. (In Russian) [Google Scholar]
- Alexandrowicz, J.S. Zur Kenntnis des sympathischen Nervensystems der Crustaceae. Jenaische Zeitschrift fur Naturwissenschaft 1909, 45, 395–444. (In German) [Google Scholar]
- Elwood, R.W.; Appel, M. Pain experience in hermit crabs? Anim. Behav. 2009, 77, 1243–1246. [Google Scholar] [CrossRef]
- Elwood, R.W.; Adams, L. Electric shock causes physiological stress responses in shore crabs, consistent with prediction of pain. Biol. Lett. 2015, 11, 20150800. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Spaziani, E. Stress reduces hemolymph ecdysteroid levels in the crab: mediation by the eyestalks. J. Exp. Zool. 1985, 234, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Pagé, M.-P.; Cooper, R.L. Novelty stress and reproductive state alters responsiveness to sensory stimuli and 5-HT neuromodulation. Comp. Biochem. Physiol. Part A Mol. Integr. Phys. 2004, 139, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Sneddon, L.U.; Taylor, A.C.; Huntingford, F.A.; Watson, D.G. Agonistic behavior and biogenic amines in shore crabs Carcinus maenas. J. Exp. Biol. 2000, 203, 537–545. [Google Scholar] [PubMed]
- Fingerman, M. Endocrine mechanisms in crayfish, with emphasis on reproduction and neurotransmitter regulation of hormone release. Am. Zool. 1995, 35, 68–78. [Google Scholar] [CrossRef]
- Mello, J.J.; Cromarty, S.I.; Kass-Simon, G. Increased aggressiveness in gravid American Lobsters, Homarus americanus. Aggress. Behav. 1999, 25, 451–472. [Google Scholar] [CrossRef]
- Tilden, A.R.; Brauch, R.; Ball, R.; Janze, A.M.; Ghaffari, A.H.; Sweeney, K.; Yurek, J.C.; Cooper, R.L. Modulatory effects of melatonin on behavior, hemolymph metabolites, and neurotransmitter release in crayfish. Brain Res. 2003, 992, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Monastirioti, M. Biogenic amine systems in the fruit fly Drosophila melanogaster. Microsc. Res. Tech. 1999, 45, 106–121. [Google Scholar] [CrossRef]
- Goosey, M.; Candy, D. The d-octopamine content of the haemolymph of the locust, Schistocerca Americana gregaria and its elevation during flight. Insect Biochem. 1980, 10, 393–397. [Google Scholar] [CrossRef]
- Zhu, Y.-C.; Yocum, E.; Sifers, J.; Uradu, H.; Cooper, R.L. Modulatory effects on Drosophila larva hearts in room temperature, acute and chronic cold stress. J. Comp. Physiol. B 2016, 186, 829–841. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y. The Effect of Cold on the Physiology of Drosophila Larva Heart and on Synaptic Transmission at Crayfish Neuromuscular Junctions. Theses and Dissertations Biology 2017, 50. Available online: https://uknowledge.uky.edu/biology_etds/50 (accessed on 13 September 2018).
- Cooke, I.M. Studies on the crustacean cardiac ganglion. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 1988, 91, 205–218. [Google Scholar] [CrossRef]
- Avnimelech, Y. Biofloc Technology; A Practical Guide Book; The World Aquaculture Society: Baton Rouge, LA, USA, 2009. [Google Scholar]
- McRae, T. Chemical removal of nitrite and chlorinating agents from municipal water supplies used for crayfish and aquarium finfish culture. Freshw. Crayfish 1999, 12, 727–732. [Google Scholar]
- Bierbower, S.M.; Cooper, R.L. Measures of heart and ventilatory rates in freely moving crayfish. J. Vis. Exp. 2009, 32. Available online: www.jove.com/video/1594/measures-of-heart-and-ventilatory-rates-in-freely-moving-crayfish (accessed on 13 September 2018).
- Schapker, H.; Breithaupt, T.; Shuranova, Z.; Burmistrov, Y.; Cooper, R.L. Heart rate and ventilatory correlative measures in crayfish during environmental disturbances & social interactions. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 131, 397–407. [Google Scholar]
- Listerman, L.; Deskins, J.; Bradacs, H.; Cooper, R.L. Measures of heart rate during social interactions in crayfish and effects of 5-HT. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2000, 125, 251–264. [Google Scholar] [CrossRef]
- Wycoff, S.; Weineck, K.; Conlin, S.; Grau, E.; Bradley, A.; Cantrell, D. Investigating Potential Effects of Clove Oil (Eugenol) in Model Crustaceans. IMPULSE 2018, 1–21. Available online: https://impulse.appstate.edu/articles/2018/effects-clove-oil-eugenol-proprioceptive-neurons-heart-rate-and-behavior-model-crustac (accessed on 13 September 2018).
- Bradacs, H.; Cooper, R.L.; Msghina, M.; Atwood, H.L. Differential physiology and morphology of phasic and tonic motor axons in a crayfish limb extensor muscle. J. Exp. Biol 1997, 200 Pt 4, 677–691. [Google Scholar]
- Eckert, B. Über das Zusammenwirken des erregenden und des hemmenden Neurons des M. abductor der Krebsschere beim Ablauf von Reflexen des myotatischen Typus. Z. Vgl. Physiol. 1959, 41, 500–526. [Google Scholar] [CrossRef]
- Wiens, T.J.; Gerstein, G.L. Reflex pathways of the crayfish claw. J. Comp. Physiol. A 1976, 107, 309–326. [Google Scholar] [CrossRef]
- Wilkens, J.L.; Mercier, A.J.; Evans, J. Cardiac and ventilatory responses to stress and to neurohormonal modulators by the shore crab Carcinus maenas. Comp. Biochem. Physiol. C 1985, 82, 337–343. [Google Scholar] [CrossRef]
- Wilkens, J.L.; Wilkens, L.A.; McMahon, B.R. Central control of cardiac and scaphognathite pacemakers in the crab, Cancer magister. J. Comp. Physiol. 1974, 90, 89–104. [Google Scholar] [CrossRef]
- Kramer, A.P.; Krasne, F.B. Crayfish escape behavior: production of tailflips without giant fiber activity. J. Neurophysiol. 1984, 52, 189–211. [Google Scholar] [CrossRef] [PubMed]
- Krasne, F.B.; Wine, J.J. The Production of Crayfish Tailflip Escape Responses. In Neural Mechanisms of Startle Behavior; Eaton, R.C., Ed.; Plenum Press: New York, NY, USA, 1984; pp. 179–211. [Google Scholar]
- Reichert, H.; Wine, J.J. Neural mechanisms for serial order in a stereotyped behaviour sequence. Nature 1982, 296, 86–87. [Google Scholar] [CrossRef] [PubMed]
- Wine, J.J. The structural basis of an innate behavioural pattern. J. Exp. Biol. 1984, 112, 283–319. [Google Scholar]
- Edwards, D.H.; Heitler, W.J.; Krasne, F.B. Fifty years of a command neuron: The neurobiology of escape behavior in the crayfish. Trends Neurosci. 1999, 22, 153–160. [Google Scholar] [CrossRef]
- Faulkes, Z. Turning loss into opportunity: The key deletion of an escape circuit in decapod crustaceans. Brain Behav. Evol. 2008, 72, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Astley, C.A.; Smith, O.A.; Ray, R.D.; Golanov, E.V.; Chesney, M.A.; Chalyan, V.G. Integrating behavior and cardiovascular responses: The code. Am. J. Physiol. 1991, 261, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Richardson, R.; Wang, P.; Campbell, B.A. Developmental and pharmacological analysis of the cardiac response to an acoustic startle stimulus. Psychophysiology 1996, 33, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Smith, O.A.; Astley, C.A.; Spelman, F.A. Cardiovascular responses in anticipation of changes in posture and locomotion. Brain Res. Bull. 2000, 53, 69–76. [Google Scholar] [CrossRef]
- Invasive Species Compendium (nd). Compendium Coordinator. CAB International. Available online: https://www.cabi.org/isc/datasheet/71097 (accessed on 13 September 2018).
- Milliken, M.; Williams, A. Synopsis of Biological data on the Blue Crab, Callinectes Sapidus Rathbun. NOAA Tech. Rep. NMFS 1984, 1, 1–39. Available online: http://aquaticcommons.org/2821/1/tr1opt.pdf (accessed on 13 September 2018).
- Shuranova, Z.P.; Burmistrov, Y.M.; Cooper, R.L. Activity of the ventilatory muscles in the crayfish. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2003, 134, 461–469. [Google Scholar] [CrossRef]
- Chen, R.; Xiao, M.; Buchberger, A.; Li, L. Quantitative neuropeptidomics study of the effects of temperature change in the crab Cancer borealis. J. Proteome Res. 2014, 13, 5767–5776. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.S. Stressed-out lobsters: Crustacean hyperglycemic hormone and stress proteins. Intergr. Comp. Biol. 2005, 45, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Webster, S.G. Measurement of crustacean hyperglycaemic hormone levels in the edible crab Cancer pagurus during emersion stress. J. Exp. Biol. 1996, 199, 1579–1585. [Google Scholar] [PubMed]
- Lorenzon, S.; Edomi, P.; Giulianini, P.G.; Mettulio, R.; Ferrero, E.A. Role of biogenic amines and CHH in the crustacean hyperglycemic stress response. J. Exp. Biol. 2005, 208, 3341–3347. [Google Scholar] [CrossRef] [PubMed]
- Elwood, R.W.; Barr, S.; Patterson, L. Pain and stress in crustaceans? Appl. Anim. Behav. Sci. 2009, 118, 128–136. [Google Scholar] [CrossRef]
- Puri, S.; Faulkes, Z. Can crayfish take the heat? Procambarus clarkii show nociceptive behaviour to high temperature stimuli, but not low temperature or chemical stimuli. Biol. Open 2015, 4, 441–448. [Google Scholar] [CrossRef]
- Sneddon, L.U. Comparative physiology of nociception and pain. Physiology 2018, 33, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Walters, E.T. Nociceptive biology of molluscs and arthropods: Evolutionary clues about functions and mechanisms potentially related to pain. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]










© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weineck, K.; Ray, A.J.; Fleckenstein, L.J.; Medley, M.; Dzubuk, N.; Piana, E.; Cooper, R.L. Physiological Changes as a Measure of Crustacean Welfare under Different Standardized Stunning Techniques: Cooling and Electroshock. Animals 2018, 8, 158. https://doi.org/10.3390/ani8090158
Weineck K, Ray AJ, Fleckenstein LJ, Medley M, Dzubuk N, Piana E, Cooper RL. Physiological Changes as a Measure of Crustacean Welfare under Different Standardized Stunning Techniques: Cooling and Electroshock. Animals. 2018; 8(9):158. https://doi.org/10.3390/ani8090158
Chicago/Turabian StyleWeineck, Kristin, Andrew J. Ray, Leo J. Fleckenstein, Meagan Medley, Nicole Dzubuk, Elena Piana, and Robin L. Cooper. 2018. "Physiological Changes as a Measure of Crustacean Welfare under Different Standardized Stunning Techniques: Cooling and Electroshock" Animals 8, no. 9: 158. https://doi.org/10.3390/ani8090158
APA StyleWeineck, K., Ray, A. J., Fleckenstein, L. J., Medley, M., Dzubuk, N., Piana, E., & Cooper, R. L. (2018). Physiological Changes as a Measure of Crustacean Welfare under Different Standardized Stunning Techniques: Cooling and Electroshock. Animals, 8(9), 158. https://doi.org/10.3390/ani8090158