
Effects of Varied Stimuli on Escape Behavior Diversification of Himalayan Marmots for Different Human Disturbances

 and
and {kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
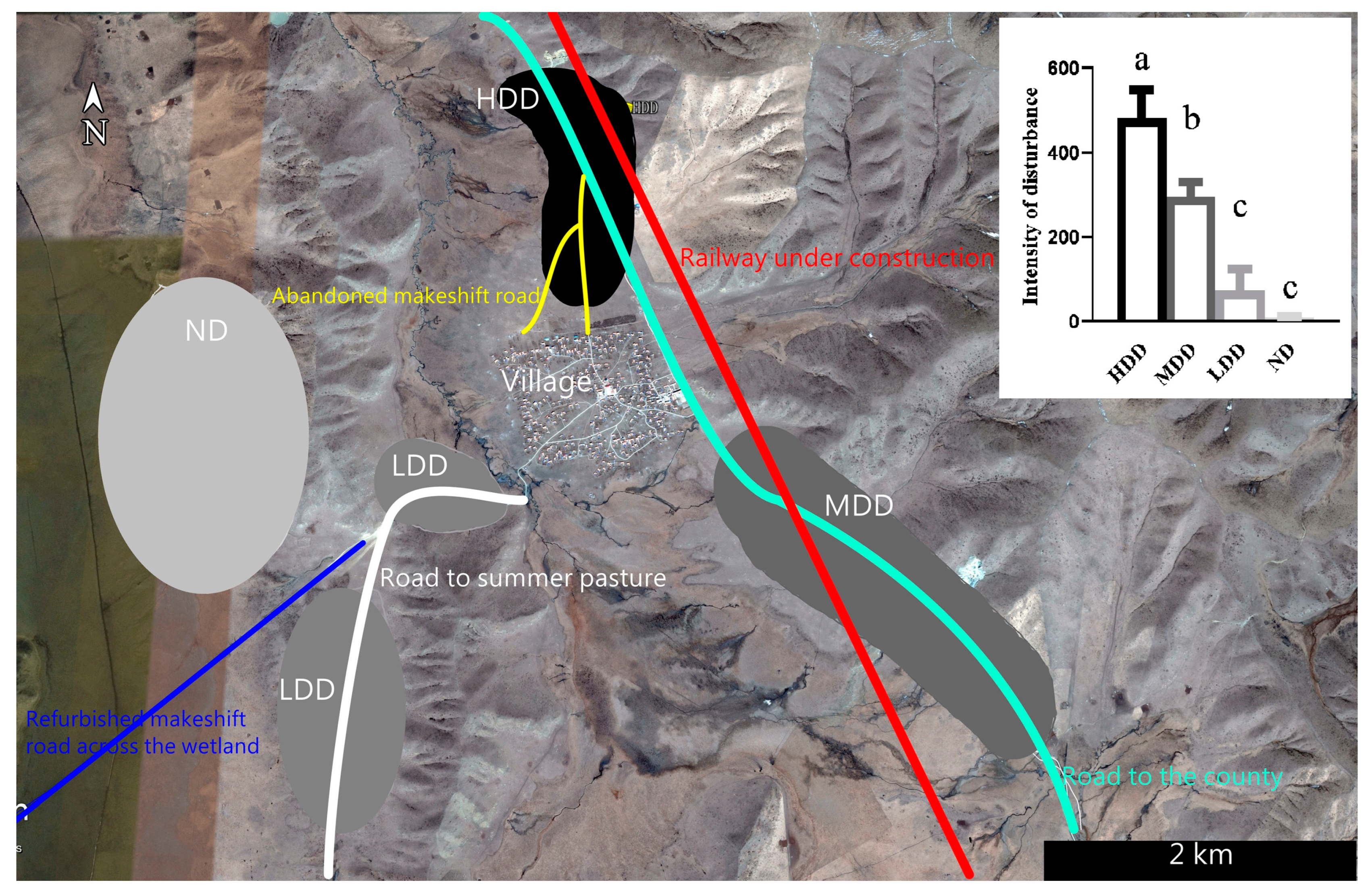
2.1. Study Site
2.2. Disturbance Intensity
2.3. Data Collection
2.4. Experimental Procedure
2.5. Data Analysis
3. Results
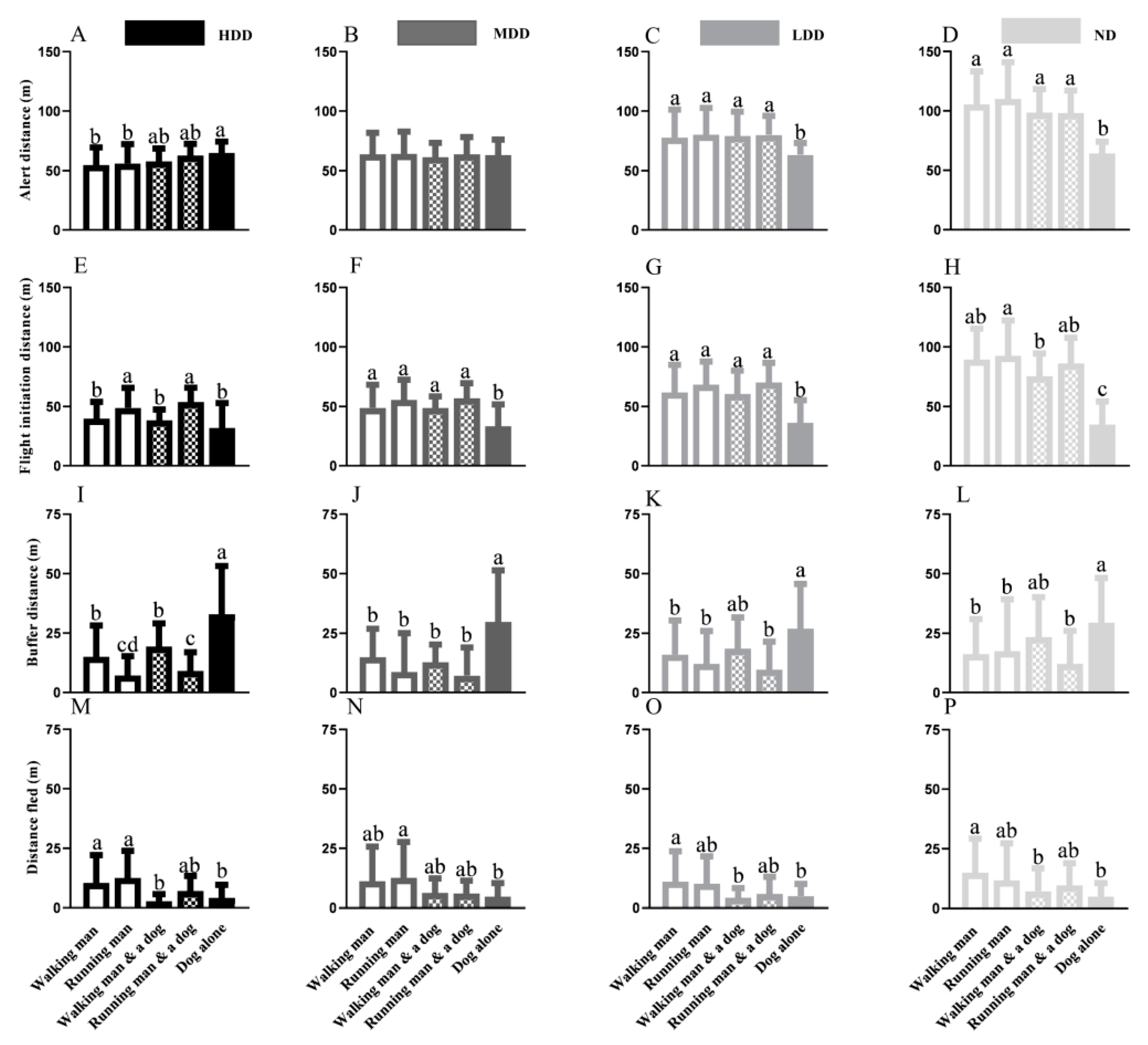
3.1. Intra-Population Variation When Disturbed by Varied Stimuli
3.2. Inter-Population Variation When Disturbed by the Same Stimulus
4. Discussions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human-induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [PubMed]
- Matthews, T.J.; Triantis, K.A.; Wayman, J.P.; Martin, T.E.; Hume, J.P.; Cardoso, P.; Faurby, S.; Mendenhall, C.D.; Dufour, P.; Rigal, F.; et al. The global loss of avian functional and phylogenetic diversity from anthropogenic extinctions. Science 2024, 386, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.X.T.; Amir, Z.; Moore, J.H.; Gaynor, K.M.; Luskin, M.S. Effects of human disturbances on wildlife behaviour and consequences for predator-prey overlap in Southeast Asia. Nat. Commun. 2024, 15, 1521. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Jimenez, C.A.; Marcus, M.; Erika, P.; Andrew, S.; Jennifer, E.S. Behavioural correlations across multiple stages of the antipredator response: Do animals that escape sooner hide longer? Anim. Behav. 2022, 185, 175–184. [Google Scholar] [CrossRef]
- Wrege, P.H.; Rowland, E.D.; Thompson, B.G.; Batruch, N. Use of acoustic tools to reveal otherwise cryptic responses of forest elephants to oil exploration. Conserv. Biol. 2010, 24, 1578–1585. [Google Scholar] [CrossRef]
- Zhang, C.; Li, Y.; Hu, X.; Ma, X.; Jia, W.; Liu, K.; Nie, Y. Human-induced behavioural changes of global threatened terrestrial mammals. Glob. Ecol. Biogeogr. 2023, 32, 1645–1659. [Google Scholar] [CrossRef]
- Guo, C.; Zhou, S.; Krzton, A.; Xiang, Z. Variation in escape response of Himalayan marmots (Marmota himalayana) under different human disturbances. Ethology 2022, 128, 143–151. [Google Scholar] [CrossRef]
- Uchida, K.; Suzuki, K.; Shimamoto, T.; Yanagawa, H.; Koizumi, I. Escaping height in a tree represents a potential indicator of fearfulness in Arboreal squirrels. Mammal Study 2017, 42, 39–43. [Google Scholar] [CrossRef]
- Larm, M.; Norén, K.; Angerbjörn, A. Temporal activity shift in arctic foxes (Vulpes lagopus) in response to human disturbance. Glob. Ecol. Conserv. 2021, 27, e01602. [Google Scholar] [CrossRef]
- Manlick, P.J.; Pauli, J.N. Human disturbance increases trophic niche overlap in terrestrial carnivore communities. Proc. Natl. Acad. Sci. USA 2020, 117, 26842–26848. [Google Scholar] [CrossRef]
- Poudel, B.S.; Spooner, P.G.; Matthews, A. Temporal shift in activity patterns of Himalayan marmots in relation to pastoralism. Behav. Ecol. 2015, 26, 1345–1351. [Google Scholar] [CrossRef]
- Stankowich, T.; Blumstein, D. Fear in animals: A meta-analysis and review of risk assessment. Proc. R. Soc. B Biol. Sci. 2005, 272, 2627–2634. [Google Scholar] [CrossRef]
- Fernández-Juricic, E.; Jimenez, M.D.; Lucas, E. Alert distance as an alternative measure of bird tolerance to human disturbance: Implications for park design. Environ. Conserv. 2001, 28, 263–269. [Google Scholar] [CrossRef]
- Williams, D.M.; Nguyen, P.-T.; Chan, K.; Krohn, M.; Blumstein, D.T. High human disturbance decreases individual variability in skink escape behavior. Curr. Zool. 2020, 66, 63–70. [Google Scholar] [CrossRef]
- Chen, X.-M.; Xie, W.-T.; Shuai, L.-Y. Flush early and avoid the rush? It may depend on where you stand. Ethology 2020, 126, 987–992. [Google Scholar] [CrossRef]
- Fernández-Juricic, E.; Jimenez, M.D.; Lucas, E. Factors affecting intra- and inter-specific variations in the difference between alert and flight distances in forested habitats. Can. J. Zool. 2002, 80, 1212–1220. [Google Scholar] [CrossRef]
- McLeod, E.M.; Guay, P.-J.; Taysom, A.J.; Robinson, R.W.; Weston, M.A. Buses, cars, bicycles and walkers: The influence of the type of human transport on the flight responses of waterbirds. PLoS ONE 2013, 8, e82008. [Google Scholar] [CrossRef]
- Stankowich, T. Ungulate flight responses to human disturbance: A review and meta-analysis. Biol. Conserv. 2008, 141, 2159–2173. [Google Scholar] [CrossRef]
- Shearer, D.J.; Beilke, E.A. Playing it by ear: Gregarious sparrows recognize and respond to isolated wingbeat sounds and predator-based cues. Anim. Cogn. 2022, 26, 357–367. [Google Scholar] [CrossRef]
- Valcarcel, A.; Fernández-Juricic, E. Antipredator strategies of house finches: Are urban habitats safe spots from predators even when humans are around? Behav. Ecol. Sociobiol. 2009, 63, 673–685. [Google Scholar] [CrossRef]
- Griffin, S.C.; Valois, T.; Taper, M.L.; Scott Mills, L. Effects of tourists on behavior and demography of Olympic marmots. Conserv. Biol. 2007, 21, 1070–1081. [Google Scholar] [CrossRef] [PubMed]
- Díaz, E.A.; Saenz, C.M.S.; Vega, Y.Y.M.; Rubio, E.; González, G.; Zug, R.; Zapata-Ríos, G. Dog and cat-related attacks on wildlife in the Metropolitan district of Quito, Ecuador: An integrative approach to reduce the impact. Ecosyst. People 2023, 19, 2191735. [Google Scholar] [CrossRef]
- Cavalli, M.; Baladrón, A.V.; Isacch, J.P.; Biondi, L.M.; Bó, M.S. Differential risk perception of rural and urban Burrowing owls exposed to humans and dogs. Behav. Process. 2016, 124, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Zaman, M.; Tolhurst, B.A.; Zhu, M.; Jiang, G. Increased flight initiation distance (FID) in golden marmots (Marmota caudata aurea) responding to domestic dogs in a landscape of human disturbance. Animals 2019, 9, 605. [Google Scholar] [CrossRef]
- Minini, B.; Neuhaus, P.; Ingold, I. Behaviour of marmots marmota marmota under the influence of different hiking activities. Biol. Conserv. 1993, 64, 161. [Google Scholar] [CrossRef]
- Holderness-Roddam, B.; McQuillan, P.B. Domestic dogs (Canis familiaris) as a predator and disturbance agent of wildlife in Tasmania. Australas. J. Environ. Manag. 2014, 21, 441–452. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Shrestha, T. Marmota himalayana (errata Version Published in 2017). The IUCN Red List of Threatened Species 2016: E.T12826A115106426. Available online: https://www.iucnredlist.org/species/12826/115106426 (accessed on 10 February 2025).
- Bao, Z.; Guo, C.; Chen, Y.; Li, C.; Lei, T.; Zhou, S.; Qi, D.; Xiang, Z. Fatty acid metabolization and insulin regulation prevent liver injury from lipid accumulation in Himalayan marmots. Cell Rep. 2023, 42, 112718. [Google Scholar] [CrossRef]
- Cooper, W. Dynamic risk assessment: Prey rapidly adjust flight initiation distance to changes in predator approach speed. Ethology 2006, 112, 858–864. [Google Scholar] [CrossRef]
- Zhou, S.; Krzton, A.; Gao, S.; Guo, C.; Xiang, Z. Effects of human activity on the habitat utilization of Himalayan marmot (Marmota himalayana) in Zoige wetland. Ecol. Evol. 2021, 11, 8957–8968. [Google Scholar] [CrossRef]
- Guo, C.; Gao, S.; Zhou, S.; Zhang, L.; Xiang, Z. Burrowing and anti-predator requirements determine the microhabitat selection of Himalayan marmot in Zoige wetland. Zool. Sci. 2020, 37, 554–562. [Google Scholar] [CrossRef]
- Samia, D.; Blumstein, D.; Stankowich, T.; Cooper, W. Fifty years of chasing lizards: New insights advance optimal escape theory. Biol. Rev. 2015, 91, 349–366. [Google Scholar] [CrossRef] [PubMed]
- Muposhi, V.; Gandiwa, E.; Makuza, S.; Bartels, P. Trophy hunting and perceived risk in closed ecosystems: Flight behaviour of three gregarious African ungulates in a semi-arid tropical savanna. Austral Ecol. 2016, 41, 809–818. [Google Scholar] [CrossRef]
- Baskin, L.M.; Hjältén, J. Fright and flight behavior of reindeer. Alces 2001, 37, 435–445. [Google Scholar]
- Uchida, K.; Blumstein, D. Habituation or sensitization? Long-term responses of yellow-bellied marmots to human disturbance. Behav. Ecol. 2021, 32, arab016. [Google Scholar] [CrossRef]
- Kittendorf, A.; Dantzer, B. Urban fox squirrels exhibit tolerance to humans but respond to stimuli from natural predators. Ethology 2021, 127, 697–709. [Google Scholar] [CrossRef]
- Gómez-Serrano, M. Four-legged foes: Dogs disturb nesting plovers more than people do on tourist beaches. Ibis 2020, 163, 338–352. [Google Scholar] [CrossRef]
- Zukerman, Y.; Berger-Tal, O. Exposure of Nubian ibex (Capra nubiana) to humans reduces behavioural responses to potential threats. People Nat. 2024, 6, 562–572. [Google Scholar] [CrossRef]
- Jorgensen, J.; Dinan, L.; Brown, M. Flight initiation distances of nesting Piping plovers (Charadrius melodus) in response to human disturbance. Avian Conserv. Ecol. 2016, 11, 5. [Google Scholar] [CrossRef]
- Ota, K. Fight, fatigue and flight: Narrowing of attention to a threat compensates for decreased anti-predator vigilance. J. Exp. Biol. 2018, 221, jeb168047. [Google Scholar] [CrossRef]
- Bateman, P.; Fleming, P. Who are you looking at? Hadeda ibises use direction of gaze, head orientation and approach speed in their risk assessment of a potential predator. J. Zool. 2011, 285, 316–323. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, J.; Zhang, C.; Liang, W. Face masks matter: Eurasian tree sparrows show reduced fear responses to people wearing face masks during the COVID-19 pandemic. Glob. Ecol. Conserv. 2020, 24, e01277. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wei, Y.; Liu, Y.; Liang, W. Responses of Black-headed gulls (Chroicocephalus ridibundus) to observers holding popguns: Adult and juvenile gulls differ. J. Nat. Conserv. 2025, 84, 126824. [Google Scholar] [CrossRef]
- Cooper, W.E., Jr.; Stankowich, T. Prey or predator? Body size of an approaching animal affects decisions to attack or escape. Behav. Ecol. 2010, 21, 1278–1284. [Google Scholar] [CrossRef]
- Zhou, B.; Liang, W. Avian escape responses to observers wearing clothing of different colors: A comparison of urban and rural populations. Glob. Ecol. Conserv. 2020, 22, e00921. [Google Scholar] [CrossRef]
- Feng, C.; Liang, W. Living together: Waterbirds distinguish between local fishermen and casual outfits. Glob. Ecol. Conserv. 2020, 22, e00994. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, T.; Peng, H.; Zhang, H.; Ban, Y.; Zaman, M.; Xiang, Z.; Guo, C. Effects of Varied Stimuli on Escape Behavior Diversification of Himalayan Marmots for Different Human Disturbances. Animals 2025, 15, 935. https://doi.org/10.3390/ani15070935
Lei T, Peng H, Zhang H, Ban Y, Zaman M, Xiang Z, Guo C. Effects of Varied Stimuli on Escape Behavior Diversification of Himalayan Marmots for Different Human Disturbances. Animals. 2025; 15(7):935. https://doi.org/10.3390/ani15070935
Chicago/Turabian StyleLei, Tao, Hua Peng, Han Zhang, Ying Ban, Muhammad Zaman, Zuofu Xiang, and Cheng Guo. 2025. "Effects of Varied Stimuli on Escape Behavior Diversification of Himalayan Marmots for Different Human Disturbances" Animals 15, no. 7: 935. https://doi.org/10.3390/ani15070935
APA StyleLei, T., Peng, H., Zhang, H., Ban, Y., Zaman, M., Xiang, Z., & Guo, C. (2025). Effects of Varied Stimuli on Escape Behavior Diversification of Himalayan Marmots for Different Human Disturbances. Animals, 15(7), 935. https://doi.org/10.3390/ani15070935