
Modulation of the Toll-like Receptor Pathway in Ovine Endometria During Early Pregnancy


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. RNA Extraction and RT-qPCR Assay
2.3. Western Blot Analysis
2.4. Immunohistochemistry Analysis
2.5. Statistical Analysis
3. Results
3.1. Expression of TLR2, TLR3, TLR4, TLR5, IRAK1, TRAF6, and MYD88 mRNAs in the Endometrium During Pregnancy
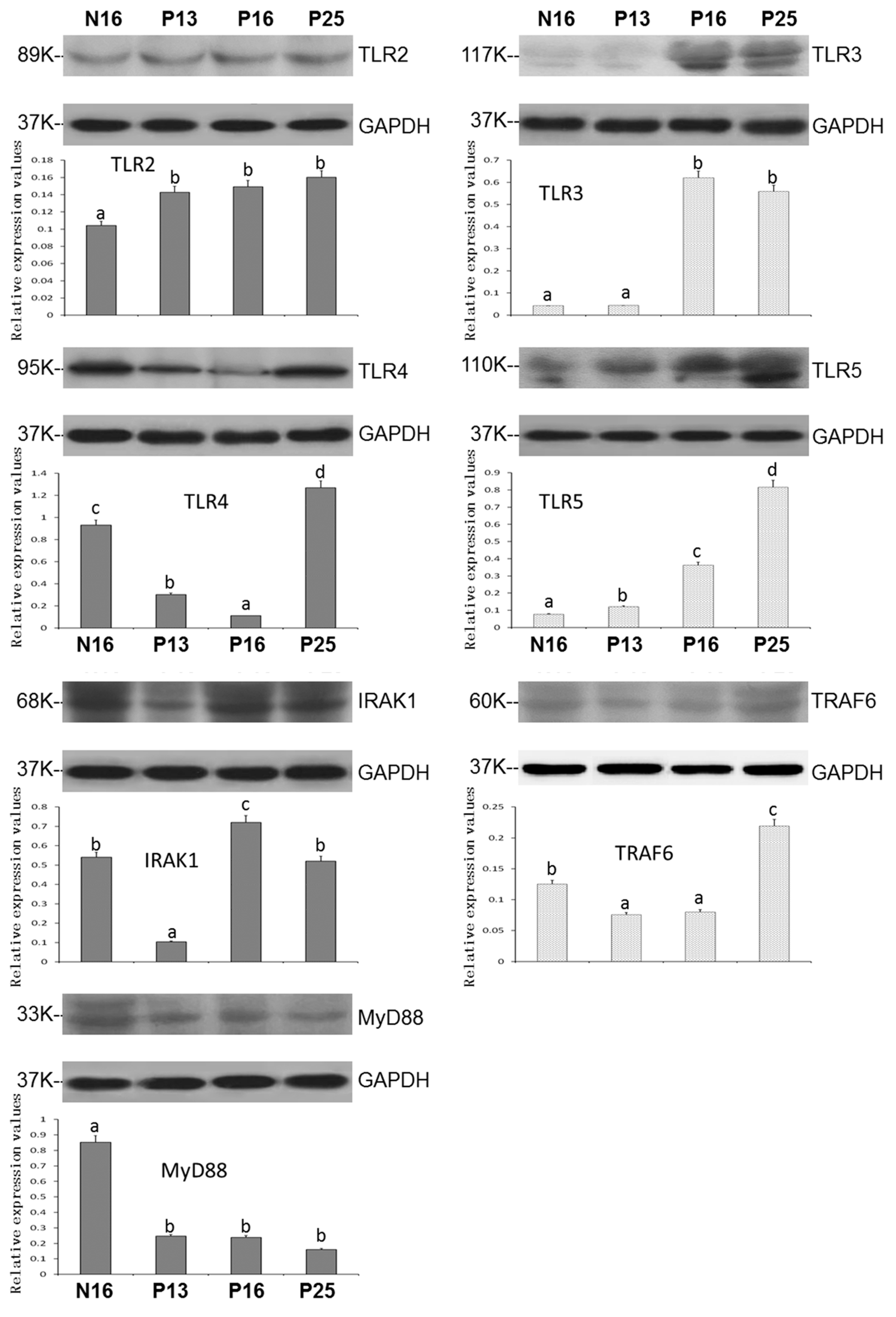
3.2. Expression of TLR2, TLR3, TLR4, TLR5, IRAK1, TRAF6, and MyD88 Proteins in the Endometrium During Pregnancy
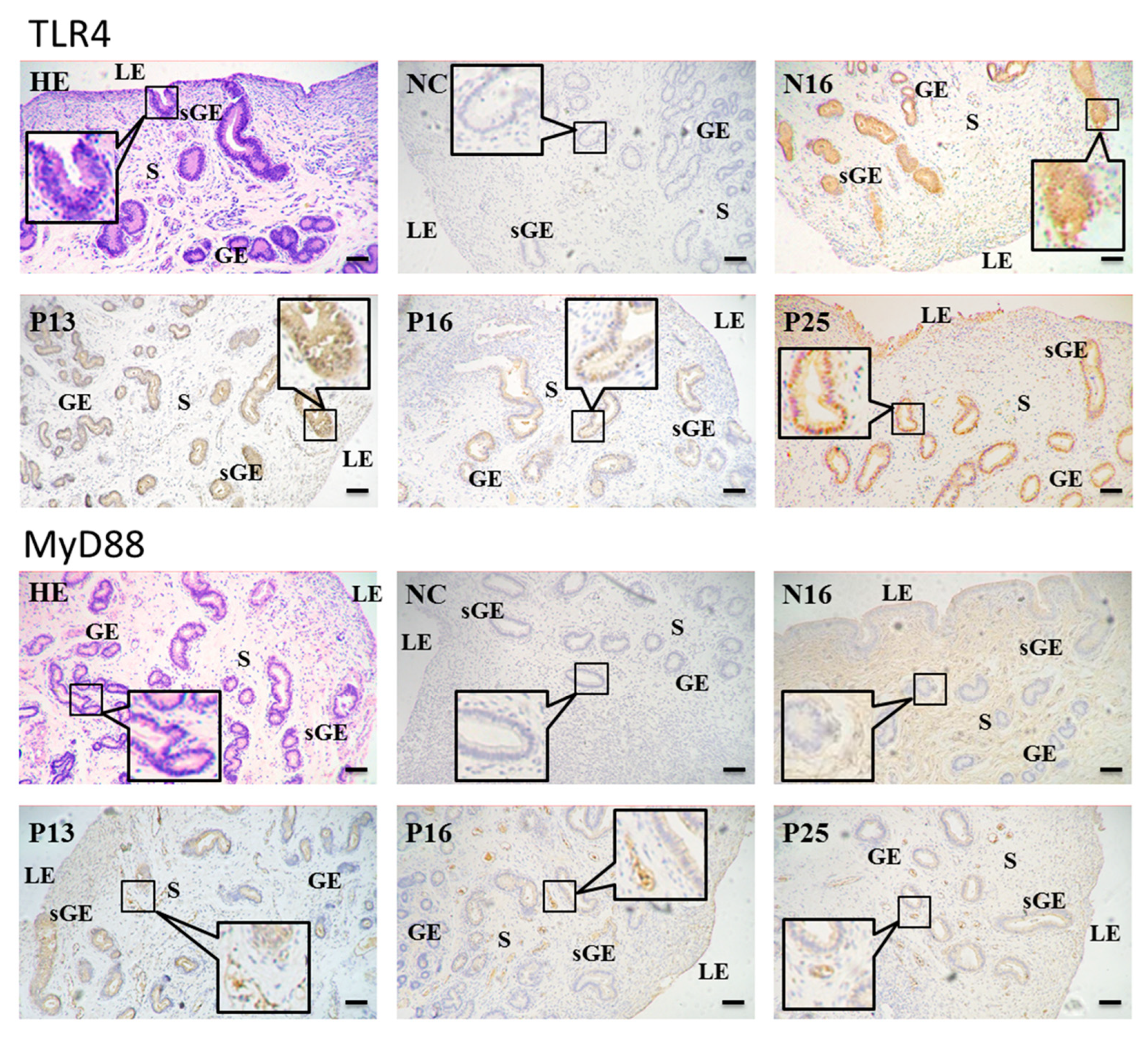
3.3. Immunohistochemistry for TLR4 and MyD88 Proteins in the Endometria
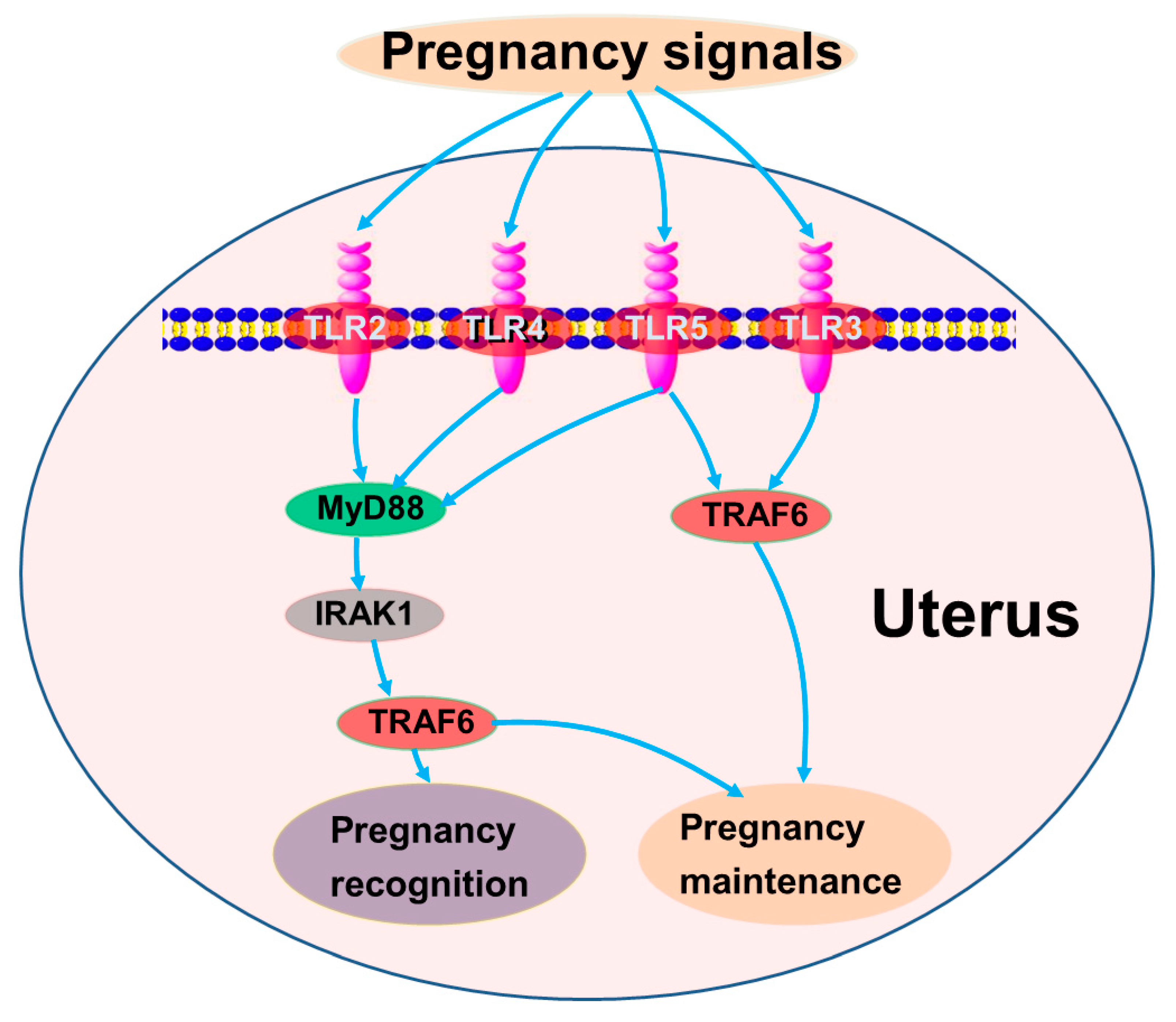
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Robertson, S.A.; Moldenhauer, L.M.; Green, E.S.; Care, A.S.; Hull, M.L. Immune determinants of endometrial receptivity: A biological perspective. Fertil. Steril. 2022, 117, 1107–1120. [Google Scholar] [CrossRef] [PubMed]
- Marozio, L.; Nuzzo, A.M.; Gullo, E.; Moretti, L.; Canuto, E.M.; Tancredi, A.; Goia, M.; Cosma, S.; Revelli, A.; Rolfo, A.; et al. Immune checkpoints in recurrent pregnancy loss: New insights into a detrimental and elusive disorder. Int. J. Mol. Sci. 2023, 24, 13071. [Google Scholar] [CrossRef]
- Wang, J.; Han, T.; Zhu, X. Role of maternal-fetal immune tolerance in the establishment and maintenance of pregnancy. Chin. Med. J. 2024, 137, 1399–1406. [Google Scholar] [CrossRef] [PubMed]
- Campanile, G.; Baruselli, P.S.; Limone, A.; D’Occhio, M.J. Local action of cytokines and immune cells in communication between the conceptus and uterus during the critical period of early embryo development, attachment and implantation—Implications for embryo survival in cattle: A review. Theriogenology 2021, 167, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Liu, J.; Wang, Y.; Zhao, W.; Wang, W.; Cui, J.; Yang, J.; Yue, Y.; Zhang, S.; Chu, M.; et al. A proteomic atlas of ligand-receptor interactions at the ovine maternal-fetal interface reveals the role of histone lactylation in uterine remodeling. J. Biol. Chem. 2022, 298, 101456. [Google Scholar] [CrossRef]
- Zhang, L.; Cai, J.; Wang, X.; Yang, Z.; Ding, H.; Yang, L. Effects of early pregnancy on NOD-like receptor expression in the ovine endometrium. Front. Vet. Sci. 2024, 11, 1384386. [Google Scholar] [CrossRef]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like receptors and the control of immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef]
- Koga, K.; Izumi, G.; Mor, G.; Fujii, T.; Osuga, Y. Toll-like receptors at the maternal-fetal interface in normal pregnancy and pregnancy complications. Am. J. Reprod. Immunol. 2014, 72, 192–205. [Google Scholar] [CrossRef]
- Kaya, M.S.; Kose, M.; Guzeloglu, A.; Kıyma, Z.; Atli, M.O. Early pregnancy-related changes in toll-like receptors expression in ovine trophoblasts and peripheral blood leukocytes. Theriogenology 2017, 93, 40–45. [Google Scholar] [CrossRef]
- Afkham, A.; Eghbal-Fard, S.; Heydarlou, H.; Azizi, R.; Aghebati-Maleki, L.; Yousefi, M. Toll-like receptors signaling network in pre-eclampsia: An updated review. J. Cell. Physiol. 2019, 234, 2229–2240. [Google Scholar] [CrossRef]
- Yoo, I.; Han, J.; Lee, S.; Jung, W.; Kim, J.H.; Kim, Y.W.; Kim, H.J.; Hong, M.; Ka, H. Analysis of stage-specific expression of the toll-like receptor family in the porcine endometrium throughout the estrous cycle and pregnancy. Theriogenology 2019, 125, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Mathew, D.J.; Peterson, K.D.; Senn, L.K.; Oliver, M.A.; Ealy, A.D. Ruminant conceptus-maternal interactions: Interferon-tau and beyond. J. Anim. Sci. 2022, 100, skac123. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Sui, L.; Miao, K.; An, L.; Wang, D.; Hou, Z.; Wang, R.; Guo, M.; Wang, Z.; Xu, J.; et al. Comparative analysis between endometrial proteomes of pregnant and non-pregnant ewes during the peri-implantation period. J. Anim. Sci. Biotechnol. 2015, 6, 18. [Google Scholar] [CrossRef]
- Kose, M.; Hitit, M.; Kaya, M.S.; Kırbas, M.; Dursun, S.; Alak, I.; Atli, M.O. Expression pattern of microRNAs in ovine endometrium during the peri-implantation. Theriogenology 2022, 191, 35–46. [Google Scholar] [CrossRef]
- Gao, M.; Cai, C.; Han, X.; Wang, L.; Zhang, W.; Zhang, L.; Yang, L. The early stage of pregnancy modulates toll-like receptor signaling in the ovine liver. J. Appl. Anim. Res. 2021, 49, 374–381. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- McCoy, M.G.; Nascimento, D.W.; Veleeparambil, M.; Murtazina, R.; Gao, D.; Tkachenko, S.; Podrez, E.; Byzova, T.V. Endothelial TLR2 promotes proangiogenic immune cell recruitment and tumor angiogenesis. Sci. Signal. 2021, 14, eabc5371. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, Q.; Zhang, L.; Feng, P.; Gao, M.; Zhao, Z.; Yang, L. Toll-like receptor signaling is changed in ovine lymph node during early pregnancy. Anim. Sci. J. 2021, 92, e13541. [Google Scholar] [CrossRef]
- Hirata, T.; Osuga, Y.; Hamasaki, K.; Hirota, Y.; Nose, E.; Morimoto, C.; Harada, M.; Takemura, Y.; Koga, K.; Yoshino, O.; et al. Expression of toll-like receptors 2, 3, 4, and 9 genes in the human endometrium during the menstrual cycle. J. Reprod. Immunol. 2007, 74, 53–60. [Google Scholar] [CrossRef]
- Wira, C.R.; Rodriguez-Garcia, M.; Patel, M.V. The role of sex hormones in immune protection of the female reproductive tract. Nat. Rev. Immunol. 2015, 15, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Atli, M.O.; Kose, M.; Hitit, M.; Kaya, M.S.; Bozkaya, F. Expression patterns of Toll-like receptors in the ovine corpus luteum during the early pregnancy and prostaglandin F2α-induced luteolysis. Theriogenology 2018, 111, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Gong, H.; Liu, L.; Ramos-Solis, N.; Seye, C.I.; Derbigny, W.A. TLR3 deficiency exacerbates the loss of epithelial barrier function during genital tract Chlamydia muridarum infection. PLoS ONE 2019, 14, e0207422. [Google Scholar] [CrossRef]
- Gonzalez, J.M.; Xu, H.; Ofori, E.; Elovitz, M.A. Toll-like receptors in the uterus, cervix, and placenta: Is pregnancy an immunosuppressed state? Am. J. Obstet. Gynecol. 2007, 197, 296.e1–296.e6. [Google Scholar] [CrossRef] [PubMed]
- Gierman, L.M.; Silva, G.B.; Pervaiz, Z.; Rakner, J.J.; Mundal, S.B.; Thaning, A.J.; Nervik, I.; Elschot, M.; Mathew, S.; Thomsen, L.C.V.; et al. TLR3 expression by maternal and fetal cells at the maternal-fetal interface in normal and preeclamptic pregnancies. J. Leukoc. Biol. 2021, 109, 173–183. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, G.; Zhang, Q.; Feng, P.; Gao, M.; Yang, L. Early pregnancy affects expression of Toll-like receptor signaling members in ovine spleen. Anim. Reprod. 2021, 18, e20210009. [Google Scholar] [CrossRef]
- Allhorn, S.; Böing, C.; Koch, A.A.; Kimmig, R.; Gashaw, I. TLR3 and TLR4 expression in healthy and diseased human endometrium. Reprod. Biol. Endocrinol. 2008, 6, 40. [Google Scholar] [CrossRef]
- Firmal, P.; Shah, V.K.; Chattopadhyay, S. Insight into TLR4-mediated immunomodulation in normal pregnancy and related disorders. Front. Immunol. 2020, 11, 807. [Google Scholar] [CrossRef]
- Barboza, R.; Lima, F.A.; Reis, A.S.; Murillo, O.J.; Peixoto, E.P.M.; Bandeira, C.L.; Fotoran, W.L.; Sardinha, L.R.; Wunderlich, G.; Bevilacqua, E.; et al. TLR4-mediated placental pathology and pregnancy outcome in experimental malaria. Sci. Rep. 2017, 7, 8623. [Google Scholar] [CrossRef]
- Zhong, Y.; Zhang, Y.; Liu, W.; Zhao, Y.; Zou, L.; Liu, X. TLR4 Modulates Senescence and Paracrine Action in Placental Mesenchymal Stem Cells via Inhibiting Hedgehog Signaling Pathway in Preeclampsia. Oxid. Med. Cell. Longev. 2022, 2022, 7202837. [Google Scholar] [CrossRef]
- de Oliveira, A.A.; Elder, E.; Spaans, F.; Graton, M.E.; Quon, A.; Kirschenman, R.; Wooldridge, A.L.; Cooke, C.M.; Davidge, S.T. Excessive hypercholesterolemia in pregnancy impairs rat uterine artery function via activation of Toll-like receptor 4. Clin. Sci. 2024, 138, 137–151. [Google Scholar] [CrossRef]
- Pandya, Y.; Marta, A.; Barateiro, A.; Bandeira, C.L.; Dombrowski, J.G.; Costa, J.; Marinho, C.R.F.; Penha-Gonçalves, C. TLR4-endothelin axis controls syncytiotrophoblast motility and confers fetal protection in placental malaria. Infect. Immun. 2021, 89, e0080920. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.P.; Costa, E.A.; Macêdo, A.A.; Martins Tda, M.; Borges, A.M.; Paixão, T.A.; Santos, R.L. Transcription of pattern recognition receptors and abortive agents induced chemokines in the bovine pregnant uterus. Vet. Immunol. Immunopathol. 2012, 145, 248–256. [Google Scholar] [CrossRef]
- Karen, A.; Beckers, J.F.; Sulon, J.; de Sousa, N.M.; Szabados, K.; Reczigel, J.; Szenci, O. Early pregnancy diagnosis in sheep by progesterone and pregnancy-associated glycoprotein tests. Theriogenology 2003, 59, 1941–1948. [Google Scholar] [CrossRef] [PubMed]
- Gottipati, S.; Rao, N.L.; Fung-Leung, W.P. IRAK1: A critical signaling mediator of innate immunity. Cell. Signal. 2008, 20, 269–276. [Google Scholar] [CrossRef]
- Jain, V.G.; Kong, F.; Kallapur, S.G.; Presicce, P.; Senthamaraikannnan, P.; Cappelletti, M.; Chougnet, C.A.; Bhattacharyya, S.; Pasare, C.; Muglia, L.J. IRAK1 is a critical mediator of inflammation-induced preterm birth. J. Immunol. 2020, 204, 2651–2660. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Tan, X.; Deng, L.; Zhong, L.; Wei, C.; Ruan, H.; Lu, Y.; Pang, L. miR-146b-5p downregulates IRAK1 and ADAM19 to suppress trophoblast proliferation, invasion, and migration in miscarriage. Biol. Reprod. 2023, 109, 938–953. [Google Scholar] [CrossRef]
- Ruiz-González, I.; Xu, J.; Wang, X.; Burghardt, R.C.; Dunlap, K.A.; Bazer, F.W. Exosomes, endogenous retroviruses and toll-like receptors: Pregnancy recognition in ewes. Reproduction 2015, 149, 281–291. [Google Scholar] [CrossRef]
- Yamamoto, M.; Gohda, J.; Akiyama, T.; Inoue, J.I. TNF receptor-associated factor 6 (TRAF6) plays crucial roles in multiple biological systems through polyubiquitination-mediated NF-κB activation. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2021, 97, 145–160. [Google Scholar] [CrossRef]
- Ding, J.; Zhang, Y.; Cai, X.; Zhang, Y.; Yan, S.; Wang, J.; Zhang, S.; Yin, T.; Yang, C.; Yang, J. Extracellular vesicles derived from M1 macrophages deliver miR-146a-5p and miR-146b-5p to suppress trophoblast migration and invasion by targeting TRAF6 in recurrent spontaneous abortion. Theranostics 2021, 11, 5813–5830. [Google Scholar] [CrossRef]
- Wu, S.; Han, L.; Zhou, M.; Li, X.; Luo, L.; Wang, Z.; Yan, S.; Li, F.; Chen, J.; Yang, J. LncRNA AOC4P recruits TRAF6 to regulate EZH2 ubiquitination and participates in trophoblast glycolysis and M2 macrophage polarization which is associated with recurrent spontaneous abortion. Int. Immunopharmacol. 2023, 125, 111201. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Wang, Y.; Tian, W.; Liu, Y.; Gu, M.; Jiang, X.; Cai, Y.; Huo, R.; Li, Y.; Li, L.; et al. Exosomal miR-146a-5p derived from human umbilical cord mesenchymal stem cells can alleviate antiphospholipid antibody-induced trophoblast injury and placental dysfunction by regulating the TRAF6/NF-κB axis. J. Nanobiotechnol. 2023, 21, 419. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Li, Z.; Zong, Y.; Xia, B.; Luo, S.P.; Deng, G.P.; Gao, J. Exosome-delivered miR-410-3p reverses epithelial-mesenchymal transition, migration and invasion of trophoblasts in spontaneous abortion. J. Cell. Mol. Med. 2024, 28, e18097. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Hu, Z.; Wang, C.; Zhang, Y. The TLR/MyD88 signalling cascade in inflammation and gastric cancer: The immune regulatory network of Helicobacter pylori. J. Mol. Med. 2023, 101, 767–781. [Google Scholar] [CrossRef] [PubMed]
- Xuemei, L.; Qiu, S.; Chen, G.; Liu, M. Myrtenol alleviates oxidative stress and inflammation in diabetic pregnant rats via TLR4/MyD88/NF-κB signaling pathway. J. Biochem. Mol. Toxicol. 2021, 35, e22904. [Google Scholar] [CrossRef]
- Li, Y.; Xie, H.; Zhang, H. Protective effect of sinomenine against inflammation and oxidative stress in gestational diabetes mellitus in female rats via TLR4/MyD88/NF-κB signaling pathway. J. Food. Biochem. 2021, 45, e13952. [Google Scholar] [CrossRef]
- Tang, H.; Li, H.; Li, D.; Peng, J.; Zhang, X.; Yang, W. The gut microbiota of pregnant rats alleviates fetal growth restriction by inhibiting the TLR9/MyD88 pathway. J. Microbiol. Biotechnol. 2023, 33, 1213–1227. [Google Scholar] [CrossRef]
- Panja, S.; Benjamin, J.T.; Paria, B.C. Maternal neutrophil depletion fails to avert systemic lipopolysaccharide-induced early pregnancy defects in mice. Int. J. Mol. Sci. 2021, 22, 7932. [Google Scholar] [CrossRef]
- Li, N.; Wang, L.; Cao, N.; Zhang, L.; Han, X.; Yang, L. Early pregnancy affects the expression of toll-like receptor pathway in ovine thymus. Reprod. Biol. 2020, 20, 547–554. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Number | Source | Concentrations |
|---|---|---|---|
| Rabbit anti-TLR2 polyclonal antibody | ab191458 | Abcam, Cambridge, UK | 0.2 µg/mL |
| Mouse anti-TLR3 monoclonal antibody | sc-32232 | Santa Cruz Biotechnology, Santa Cruz, CA, USA | 0.2 µg/mL |
| Mouse anti-TLR4 monoclonal antibody | sc-293072 | Santa Cruz Biotechnology, Santa Cruz, CA, USA | 0.2 µg/mL |
| Mouse anti-TLR5 monoclonal antibody | sc-517439 | Santa Cruz Biotechnology, Santa Cruz, CA, USA | 0.2 µg/mL |
| Mouse anti-MyD88 monoclonal antibody | sc-136970 | Santa Cruz Biotechnology, Santa Cruz, CA, USA | 0.2 µg/mL |
| Mouse anti-TRAF6 monoclonal antibody | sc-8409 | Santa Cruz Biotechnology, Santa Cruz, CA, USA | 0.2 µg/mL |
| Rabbit anti-IRAK1 polyclonal antibody | ab137327 | Abcam, Cambridge, UK | 0.2 µg/mL |
| Mouse anti-GAPDH monoclonal antibody | sc-47724 | Santa Cruz Biotechnology, Santa Cruz, CA, USA | 0.2 µg/mL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, G.; Song, M.; Wu, C.; Ma, X.; Zhang, L.; Yang, L. Modulation of the Toll-like Receptor Pathway in Ovine Endometria During Early Pregnancy. Animals 2025, 15, 917. https://doi.org/10.3390/ani15070917
Yu G, Song M, Wu C, Ma X, Zhang L, Yang L. Modulation of the Toll-like Receptor Pathway in Ovine Endometria During Early Pregnancy. Animals. 2025; 15(7):917. https://doi.org/10.3390/ani15070917
Chicago/Turabian StyleYu, Guoao, Mengyao Song, Chenxu Wu, Xiaoxin Ma, Leying Zhang, and Ling Yang. 2025. "Modulation of the Toll-like Receptor Pathway in Ovine Endometria During Early Pregnancy" Animals 15, no. 7: 917. https://doi.org/10.3390/ani15070917
APA StyleYu, G., Song, M., Wu, C., Ma, X., Zhang, L., & Yang, L. (2025). Modulation of the Toll-like Receptor Pathway in Ovine Endometria During Early Pregnancy. Animals, 15(7), 917. https://doi.org/10.3390/ani15070917