


Multi-Sensor Integration and Machine Learning for High-Resolution Classification of Herbivore Foraging Behavior
 , , and
, , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Areas
2.2. Animals, Sensors, and Camera Deployment
2.3. Pre-Processing of GPS, Accelerometer, and Camera Data
2.4. Feature Calculations
2.5. Data Partition Strategy and Machine Learning Models
2.6. Models Performance Assessment
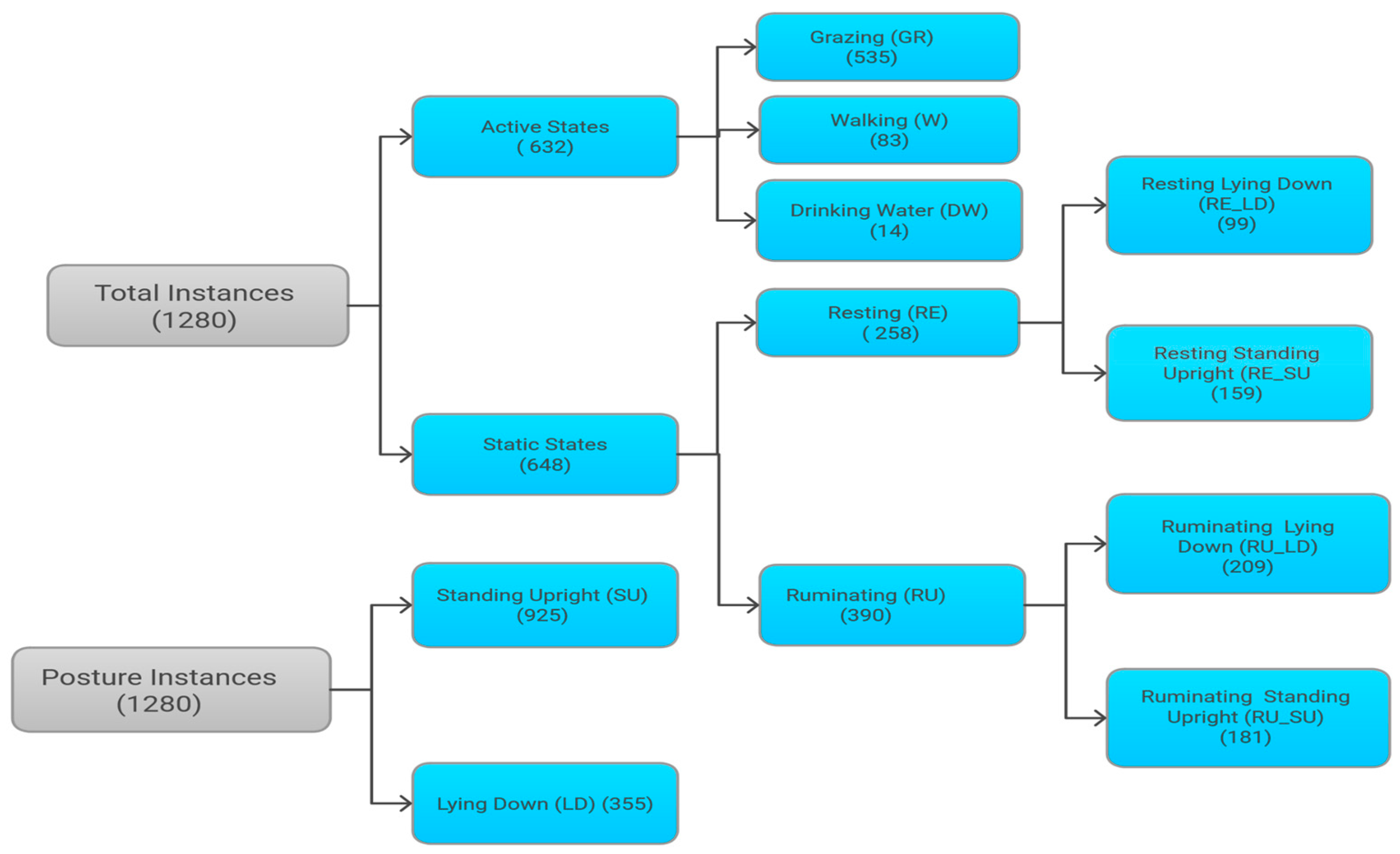
3. Results
3.1. Data Summary and Challenges
3.2. Behavior Classification Using Random Train-Test Split Method
3.2.1. Activity States Classification
3.2.2. Foraging Behaviors Classification
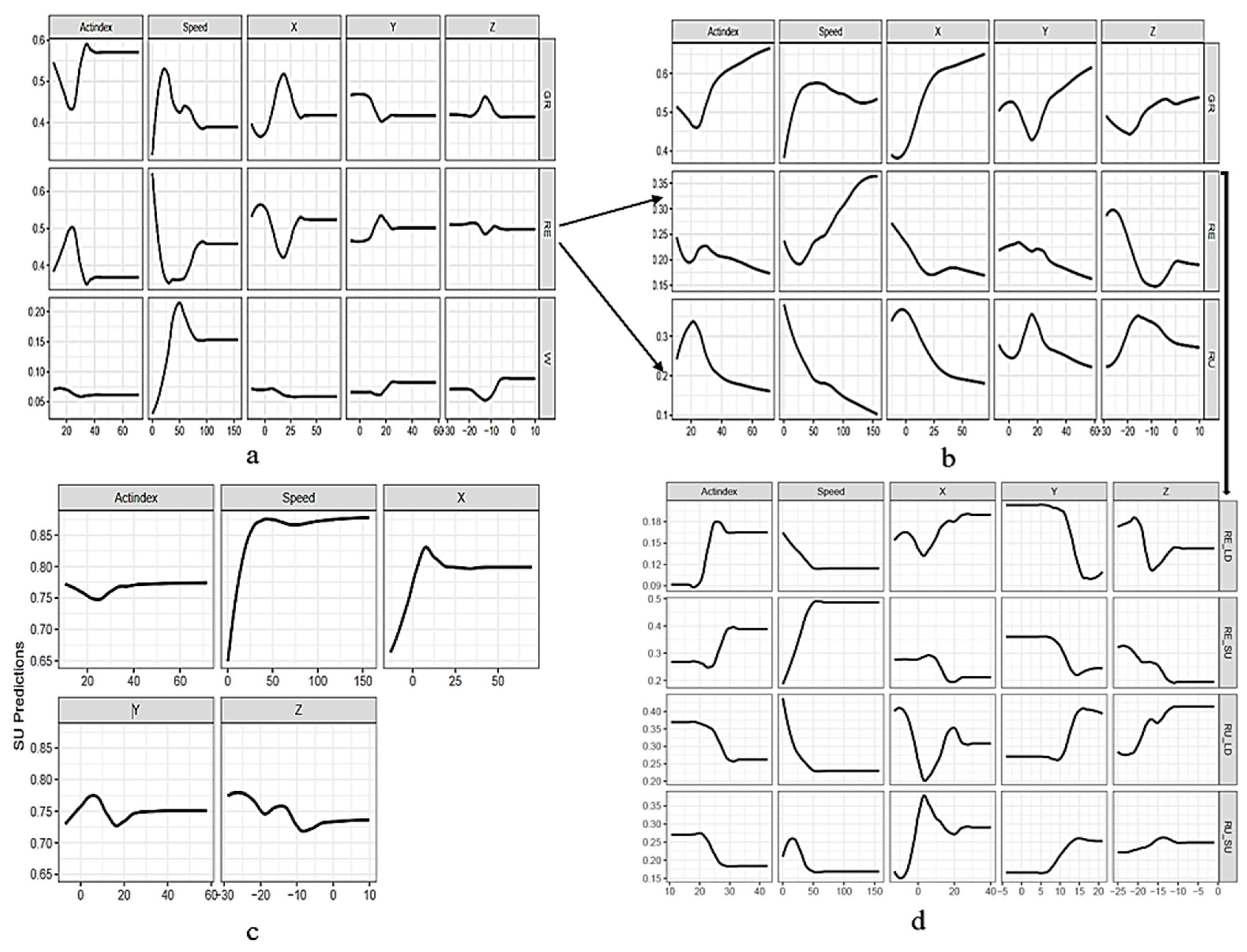
3.3. Behavior Classification Using Cross-Validation Method
3.3.1. Activity States Classification
3.3.2. Foraging Behaviors Classification and by Posture
4. Discussion
4.1. Activity State Classification
4.2. Foraging Behaviors Classification
4.3. Posture and Foraging Behaviors-by-Posture Classification
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Strömberg, C.A.E.; Staver, A.C. The History and Challenge of Grassy Biomes. Science 2022, 377, 592–593. [Google Scholar] [CrossRef] [PubMed]
- Giridhar, K.; Samireddypalle, A.; Giridhar, K.; Samireddypalle, A. Impact of climate change on forage availability for livestock. In Climate Change Impact on Livestock: Adaptation and Mitigation; Springer: New Delhi, India, 2015; pp. 97–112. [Google Scholar] [CrossRef]
- Teague, R.; Kreuter, U. Managing Grazing to Restore Soil Health, Ecosystem Function, and Ecosystem Services. Front. Sustain. Food Syst. 2020, 4, 534187. [Google Scholar] [CrossRef]
- Bilotta, G.S.; Brazier, R.E.; Haygarth, P.M. The Impacts of Grazing Animals on the Quality of Soils, Vegetation, and Surface Waters in Intensively Managed Grasslands. Adv. Agron. 2007, 94, 237–280. [Google Scholar] [CrossRef]
- Papanastasis, V.P. Restoration of Degraded Grazing Lands through Grazing Management: Can It Work? Restor. Ecol. 2009, 17, 441–445. [Google Scholar] [CrossRef]
- Afonso, B.C.; Swanepoel, L.H.; Rosa, B.P.; Marques, T.A.; Rosalino, L.M.; Santos-Reis, M.; Curveira-Santos, G. Patterns and Drivers of Rodent Abundance across a South African Multi-Use Landscape. Animals 2021, 11, 2618. [Google Scholar] [CrossRef]
- Andreychev, A.V.; Zhalilov, A.B.; Kuznetsov, V.A. The State of Local Steepe Woodchuck (Marmota Bobak) Populations in the Republic of Mordovia. Zool Zhurnal 2015, 94, 723–730. [Google Scholar] [CrossRef]
- Sanz, A.; Blanco-Penedo, I.; Quintans, G.; Álvarez-Rodríguez, J. Mother-Offspring Bonding Revisited: A Blueprint for the Future of Beef Cattle Farming. Appl. Anim. Behav. Sci. 2024, 277, 106346. [Google Scholar] [CrossRef]
- Derner, J.D.; Hunt, L.; Filho, K.E.; Ritten, J.; Capper, J.; Han, G. Livestock Production Systems. In Rangeland Systems: Processes, Management and Challenges; Springer International Publishing: Cham, Switzerland, 2017; pp. 347–372. [Google Scholar] [CrossRef]
- Bailey, D.W.; Trotter, M.G.; Knight, C.W.; Thomas, M.G. Use of GPS Tracking Collars and Accelerometers for Rangeland Livestock Production Research. Transl. Anim. Sci. 2018, 2, 81–88. [Google Scholar] [CrossRef]
- Nyamuryekung’e, S. Transforming Ranching: Precision Livestock Management in the Internet of Things Era. Rangelands 2024, 46, 13–22. [Google Scholar] [CrossRef]
- Nyamuryekung’e, S.; Cibils, A.F.; Estell, R.E.; VanLeeuwen, D.; Spiegal, S.; Steele, C.; González, A.L.; McIntosh, M.M.; Gong, Q.; Cao, H. Movement, Activity, and Landscape Use Patterns of Heritage and Commercial Beef Cows Grazing Chihuahuan Desert Rangeland. J. Arid Environ. 2022, 199, 104704. [Google Scholar] [CrossRef]
- Brennan, J.; Johnson, P.; Olson, K. Classifying Season Long Livestock Grazing Behavior with the Use of a Low-Cost GPS and Accelerometer. Comput. Electron. Agric. 2021, 181, 105957. [Google Scholar] [CrossRef]
- Cabezas, J.; Yubero, R.; Visitación, B.; Navarro-García, J.; Algar, M.J.; Cano, E.L.; Ortega, F. Analysis of Accelerometer and GPS Data for Cattle Behaviour Identification and Anomalous Events Detection. Entropy 2022, 24, 336. [Google Scholar] [CrossRef] [PubMed]
- Arcidiacono, C.; Porto, S.M.C.; Mancino, M.; Cascone, G. Development of a Threshold-Based Classifier for Real-Time Recognition of Cow Feeding and Standing Behavioural Activities from Accelerometer Data. Comput. Electron. Agric. 2017, 134, 124–134. [Google Scholar] [CrossRef]
- Hounslow, J.L.; Fossette, S.; Chong, W.; Bali, R.; Tucker, A.D.; Whiting, S.D.; Gleiss, A.C. Behaviour-Specific Spatiotemporal Patterns of Habitat Use by Sea Turtles Revealed Using Biologging and Supervised Machine Learning. J. Appl. Ecol. 2023, 60, 1828–1840. [Google Scholar] [CrossRef]
- Bailey, D.W.; Trotter, M.G.; Tobin, C.; Thomas, M.G. Opportunities to Apply Precision Livestock Management on Rangelands. Front. Sustain. Food Syst. 2021, 5, 611915. [Google Scholar] [CrossRef]
- Ganskopp, D.C.; Johnson, D.D. GPS Error in Studies Addressing Animal Movements and Activities. Rangel. Ecol. Manag. 2007, 60, 350–358. [Google Scholar] [CrossRef]
- Bonneau, M.; Vayssade, J.A.; Troupe, W.; Arquet, R. Outdoor Animal Tracking Combining Neural Network and Time-Lapse Cameras. Comput. Electron. Agric. 2020, 168, 105150. [Google Scholar] [CrossRef]
- Aquilani, C.; Confessore, A.; Bozzi, R.; Sirtori, F.; Pugliese, C. Review: Precision Livestock Farming Technologies in Pasture-Based Livestock Systems. Animal 2022, 16, 100429. [Google Scholar] [CrossRef]
- Herlin, A.; Brunberg, E.; Hultgren, J.; Högberg, N.; Rydberg, A.; Skarin, A.; Siniscalchi, M. Animal Welfare Implications of Digital Tools for Monitoring and Management of Cattle and Sheep on Pasture. Animals 2021, 11, 829. [Google Scholar] [CrossRef]
- Russel, N.S.; Selvaraj, A. Decoding Cow Behavior Patterns from Accelerometer Data Using Deep Learning. J. Vet. Behav. 2024, 74, 68–78. [Google Scholar] [CrossRef]
- Mao, A.; Huang, E.; Wang, X.; Liu, K. Deep Learning-Based Animal Activity Recognition with Wearable Sensors: Overview, Challenges, and Future Directions. Comput. Electron. Agric. 2023, 211, 108043. [Google Scholar] [CrossRef]
- Liu, J.; Bailey, D.W.; Cao, H.; Son, T.C.; Tobin, C.T. Development of a Novel Classification Approach for Cow Behavior Analysis Using Tracking Data and Unsupervised Machine Learning Techniques. Sensors 2024, 24, 4067. [Google Scholar] [CrossRef]
- Augustine, D.J.; Derner, J.D. Assessing Herbivore Foraging Behavior with GPS Collars in a Semiarid Grassland. Sensors 2013, 13, 3711–3723. [Google Scholar] [CrossRef]
- Climate United States—Monthly Averages. Available online: https://www.usclimatedata.com/ (accessed on 11 February 2025).
- DigitalCommons, U.; All Graduate Decker, T. Targeted-Grazing as a Fuels Reduction Treatment: Evaluation of Targeted-Grazing as a Fuels Reduction Treatment: Evaluation of Vegetation Dynamics and Utilization Levels Vegetation Dynamics and Utilization Levels. Master’s Thesis, Utah State University, Logan, UT, USA, 2018. [Google Scholar]
- Liu, Z.; Lin, Y.; Hoover, J.; Beene, D.; Charley, P.H.; Singer, N. Individual Level Spatial-Temporal Modelling of Exposure Potential of Livestock in the Cove Wash Watershed, Arizona. Ann. GIS 2023, 29, 87–107. [Google Scholar] [CrossRef]
- Versluijs, E.; Niccolai, L.J.; Spedener, M.; Zimmermann, B.; Hessle, A.; Tofastrud, M.; Devineau, O.; Evans, A.L. Classification of Behaviors of Free-Ranging Cattle Using Accelerometry Signatures Collected by Virtual Fence Collars. Front. Anim. Sci. 2023, 4, 1083272. [Google Scholar] [CrossRef]
- Sivakumar, M.; Parthasarathy, S.; Padmapriya, T. Trade-off between Training and Testing Ratio in Machine Learning for Medical Image Processing. PeerJ Comput. Sci. 2024, 10, e2245. [Google Scholar] [CrossRef]
- Jain, E.; Neeraja, J.; Banerjee, B.; Ghosh, P. A Diagnostic Approach to Assess the Quality of Data Splitting in Machine Learning. arXiv 2022, arXiv:2206.11721. [Google Scholar]
- Li, Y.; Shu, H.; Bindelle, J.; Xu, B.; Zhang, W.; Jin, Z.; Guo, L.; Wang, W. Classification and Analysis of Multiple Cattle Unitary Behaviors and Movements Based on Machine Learning Methods. Animals 2022, 12, 1060. [Google Scholar] [CrossRef]
- Ghojogh, B.; Ca, B.; Crowley, M.; Ca, M. The Theory Behind Overfitting, Cross Validation, Regularization, Bagging and Boosting: Tutorial. arXiv 2019, arXiv:1905.12787. [Google Scholar]
- Li, G.; Chai, L. AnimalAccML: An Open-Source Graphical User Interface for Automated Behavior Analytics of Individual Animals Using Triaxial Accelerometers and Machine Learning. Comput. Electron. Agric. 2023, 209, 107835. [Google Scholar] [CrossRef]
- Ibrahim, T.; Isaac, K.B.; Francis, B.; Lule, E.; Hellen, N.; Chongomweru, H.; Marvin, G. Interpretable Machine Learning Techniques for Predictive Cattle Behavior Monitoring. In Proceedings of the 2nd International Conference on Sustainable Computing and Smart Systems (ICSCSS), Coimbatore, India, 10–12 July 2024; pp. 1219–1224. [Google Scholar] [CrossRef]
- Deepak, D.; D’Mello, D.A.; Divakarla, U. Advancements in Automated Livestock Monitoring: A Concise Review of Deep Learning-Based Cattle Activity Recognition. In Proceedings of the 10th International Conference on Advanced Computing and Communication Systems (ICACCS), Coimbatore, India, 14–15 March 2024; pp. 321–327. [Google Scholar] [CrossRef]
- Najafabadi, M.M.; Villanustre, F.; Khoshgoftaar, T.M.; Seliya, N.; Wald, R.; Muharemagic, E. Deep Learning Applications and Challenges in Big Data Analytics. J. Big Data 2015, 2, 1–21. [Google Scholar] [CrossRef]
- Hey, T.; Butler, K.; Jackson, S.; Thiyagalingam, J. Machine Learning and Big Scientific Data. Philos. Trans. R. Soc. A 2020, 378, 20190054. [Google Scholar] [CrossRef]
- Mladenova, T.; Valova, I.; Evstatiev, B.; Valov, N.; Varlyakov, I.; Markov, T.; Stoycheva, S.; Mondeshka, L.; Markov, N. Evaluation of the Efficiency of Machine Learning Algorithms for Identification of Cattle Behavior Using Accelerometer and Gyroscope Data. AgriEngineering 2024, 6, 2179–2197. [Google Scholar] [CrossRef]
- Oshana Iddi Dissanayake, A.; McPherson, S.E.; Allyndrée, J.; Kennedy, E.; Cunningham, P.; Riaboff, L. Accelerometer-Based Multivariate Time-Series Dataset for Calf Behavior Classification. Zenodo 2024. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, R.; Li, X.; Chen, R.; Liu, J. Grazing Regime Rather than Grazing Intensity Affect the Foraging Behavior of Cattle. Ecol. Inform. 2025, 85, 102996. [Google Scholar] [CrossRef]
- Grinsztajn, L.; Oyallon, E.; Varoquaux, G. Why Do Tree-Based Models Still Outperform Deep Learning on Tabular Data? Adv. Neural Inf. Process. Syst. 2022, 35, 507–520. [Google Scholar]
- Putun, A.; Yilmaz, D. Classification of Cattle Behavior Leveraging Accelerometer Data and Machine Learning. In Proceedings of the 8th International Artificial Intelligence and Data Processing Symposium (IDAP), Malatya, Turkiye, 21–22 September 2024. [Google Scholar] [CrossRef]
- Chen, J.M.; Schütz, K.E.; Tucker, C.B. Technical Note: Comparison of Instantaneous Sampling and Continuous Observation of Dairy Cattle Behavior in Freestall Housing. J. Dairy Sci. 2016, 99, 8341–8346. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Okubo, Y.; Deguchi, Y.; Koshikawa, S.; Takahashi, M.; Chida, Y.; Okada, K. Dairy Cattle Behavior Classifications Based on Decision Tree Learning Using 3-Axis Neck-Mounted Accelerometers. Anim. Sci. J. 2019, 90, 589–596. [Google Scholar] [CrossRef]
- Zhang, H.; Ma, Y.; Shi, Y.; Qiao, Y.; Wang, M. A Framework for Automatic Analysis of Sheep Ruminating Behavior Based on Keypoint Detection. In Proceedings of the International Conference on Mechatronics and Machine Vision in Practice (M2VIP), Leeds, UK, 3–5 October 2024. [Google Scholar] [CrossRef]
- Biau, G.; Fr, G.B. Analysis of a Random Forests Model. J. Mach. Learn. Res. 2012, 13, 1063–1095. [Google Scholar]
- Kleanthous, N.; Hussain, A.; Khan, W.; Sneddon, J.; Mason, A. Feature Extraction and Random Forest to Identify Sheep Behavior from Accelerometer Data. In Proceedings of the Lecture Notes in Computer Science (Including Subseries Lecture Notes in Artificial Intelligence and Lecture Notes in Bioinformatics), 2–5 October 2020; Springer Science and Business Media Deutschland GmbH: New Delhi, India, 2020; Volume 12465 LNAI, pp. 408–419. [Google Scholar]
- Chen, T.; Guestrin, C. XGBoost: A Scalable Tree Boosting System. In Proceedings of the 22nd ACM Sigkdd International Conference on Knowledge Discovery and Data Mining, San Francisco, CA, USA, 13–17 August 2016. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Model | Model Accuracy (%) | STATES | |||||
|---|---|---|---|---|---|---|---|
| Active | Static | ||||||
| Precision (%) | Recall (%) | F1 Score (%) | Precision (%) | Recall (%) | F1 Score (%) | ||
| Perceptron | 63.8 | 68 | 52 | 59 | 62 | 76 | 68 |
| Logistic Regression | 72.4 | 76 | 65 | 70 | 70 | 79 | 74 |
| Support Vector | 71.1 | 73 | 67 | 70 | 70 | 75 | 72 |
| K-Nearest Neighbor | 74 | 73 | 74 | 74 | 74 | 74 | 74 |
| Random Forest | 73.2 | 73 | 73 | 73 | 73 | 74 | 74 |
| XGBoost | 74.2 | 77 | 69 | 73 | 72 | 79 | 75 |
| Classification | Model | Foraging Behaviors | Precision (%) | Recall (%) | F1 Score (%) | Model Accuracy (%) |
|---|---|---|---|---|---|---|
| General foraging behaviors | Perceptron | GR | 50 | 64 | 57 | 45.8 |
| RE | 54 | 35 | 42 | |||
| W | 4 | 8 | 5 | |||
| Logistic Regression | GR | 60 | 53 | 56 | 61.2 | |
| RE | 62 | 76 | 68 | |||
| W | 0 | 0 | 0 | |||
| SVM | GR | 62 | 55 | 58 | 62.5 | |
| RE | 63 | 77 | 69 | |||
| W | 100 | 4 | 8 | |||
| K-Nearest Neighbor | GR | 55 | 64 | 59 | 60.4 | |
| RE | 65 | 65 | 65 | |||
| W | 100 | 4 | 8 | |||
| Random Forest | GR | 63 | 64 | 64 | 65.9 | |
| RE | 68 | 75 | 71 | |||
| W | 50 | 4 | 7 | |||
| XGBoost | GR | 63 | 62 | 62 | 63.3 | |
| RE | 67 | 72 | 69 | |||
| W | 13 | 8 | 10 | |||
| Fine foraging behaviors | Perceptron | GR | 70 | 63 | 66 | 53.5 |
| RE | 25 | 12 | 16 | |||
| RU | 46 | 68 | 55 | |||
| Logistic Regression | GR | 62 | 76 | 68 | 56.1 | |
| RE | 0 | 0 | 0 | |||
| RU | 49 | 66 | 56 | |||
| SVM | GR | 66 | 70 | 68 | 58 | |
| RE | 53 | 12 | 19 | |||
| RU | 50 | 72 | 59 | |||
| K-Nearest Neighbor | GR | 64 | 71 | 67 | 54.9 | |
| RE | 37 | 30 | 33 | |||
| RU | 50 | 50 | 50 | |||
| Random Forest | GR | 67 | 73 | 70 | 59.7 | |
| RE | 38 | 30 | 34 | |||
| RU | 60 | 62 | 61 | |||
| XGBoost | GR | 67 | 78 | 72 | 61.7 | |
| RE | 46 | 31 | 37 | |||
| RU | 59 | 59 | 59 |
| Classification | Method | Model Accuracy (%) | Behaviors | Precision (%) | Recall (%) | F1 (%) |
|---|---|---|---|---|---|---|
| General foraging behaviors | Random Forest | 68.51 | GR | 65.2 | 67.9 | 66.5 |
| RE | 71.8 | 77.5 | 74.5 | |||
| W | 22.2 | 2.4 | 4.3 | |||
| XGBoost | 69.38 | GR | 67.1 | 67.2 | 67.2 | |
| RE | 72 | 77.9 | 74.9 | |||
| W | 48.3 | 16.9 | 25 | |||
| Fine foraging behaviors | Random Forest | 62.38 | GR | 66.6 | 80.9 | 73.1 |
| RE | 47.1 | 18.6 | 26.7 | |||
| RU | 59.6 | 65.9 | 62.6 | |||
| XGBoost | 60.35 | GR | 67.2 | 76.4 | 71.5 | |
| RE | 36.5 | 20.9 | 26.6 | |||
| RU | 58.9 | 64.4 | 61.5 | |||
| Posture | Random Forest | 83.94 | LD | 79.9 | 47.7 | 59.8 |
| SU | 84.7 | 96 | 90 | |||
| XGBoost | 83.7 | LD | 76.4 | 50.3 | 60.7 | |
| SU | 85.1 | 94.8 | 89.7 | |||
| Foraging behavior-by-posture | Random Forest | 58.87 | RE_LD | 30.6 | 15.2 | 20.3 |
| RE_SU | 46.2 | 34 | 39.1 | |||
| RU_LD | 50.9 | 52.6 | 51.8 | |||
| RU_SU | 52.2 | 39.9 | 45.2 | |||
| XGBoost | 58.78 | RE_LD | 47.6 | 10.1 | 16.7 | |
| RE_SU | 43.1 | 13.8 | 21 | |||
| RU_LD | 52.3 | 55.5 | 53.8 | |||
| RU_SU | 64.4 | 25.7 | 36.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muzzo, B.I.; Bladen, K.; Perea, A.; Nyamuryekung’e, S.; Villalba, J.J. Multi-Sensor Integration and Machine Learning for High-Resolution Classification of Herbivore Foraging Behavior. Animals 2025, 15, 913. https://doi.org/10.3390/ani15070913
Muzzo BI, Bladen K, Perea A, Nyamuryekung’e S, Villalba JJ. Multi-Sensor Integration and Machine Learning for High-Resolution Classification of Herbivore Foraging Behavior. Animals. 2025; 15(7):913. https://doi.org/10.3390/ani15070913
Chicago/Turabian StyleMuzzo, Bashiri Iddy, Kelvyn Bladen, Andres Perea, Shelemia Nyamuryekung’e, and Juan J. Villalba. 2025. "Multi-Sensor Integration and Machine Learning for High-Resolution Classification of Herbivore Foraging Behavior" Animals 15, no. 7: 913. https://doi.org/10.3390/ani15070913
APA StyleMuzzo, B. I., Bladen, K., Perea, A., Nyamuryekung’e, S., & Villalba, J. J. (2025). Multi-Sensor Integration and Machine Learning for High-Resolution Classification of Herbivore Foraging Behavior. Animals, 15(7), 913. https://doi.org/10.3390/ani15070913