
Effects of Dietary Protein and Fat Levels on Growth Performance, Nutrient Digestibility, Serum Indexes, and Rectal Fecal Microbiota of Sika Deer (Cervus nippon) Fawns in Early Wintering Period




Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Weight Determination and Feed Intake
2.2. Sample Collection and Chemical Analysis (Diet Ingredients and Excreta)
2.3. Serum Collection and Analysis
2.4. Rectal Fecal Sample Collection for Microbiome Analysis
2.5. DNA Extraction, PCR Amplification, and Sequencing of Gastrointestinal Microbiota
2.6. Statistical Analyses
3. Results
3.1. Effects of Dietary Protein and Fat Levels on Growth Performance of Sika Deer Fawns
3.2. Effect of Dietary Protein and Fat Levels on Nutrients Apparent Digestibility of Sika Deer Fawns
3.3. Effect of Dietary Protein and Fat Levels on Serum Indices of Sika Deer Fawns
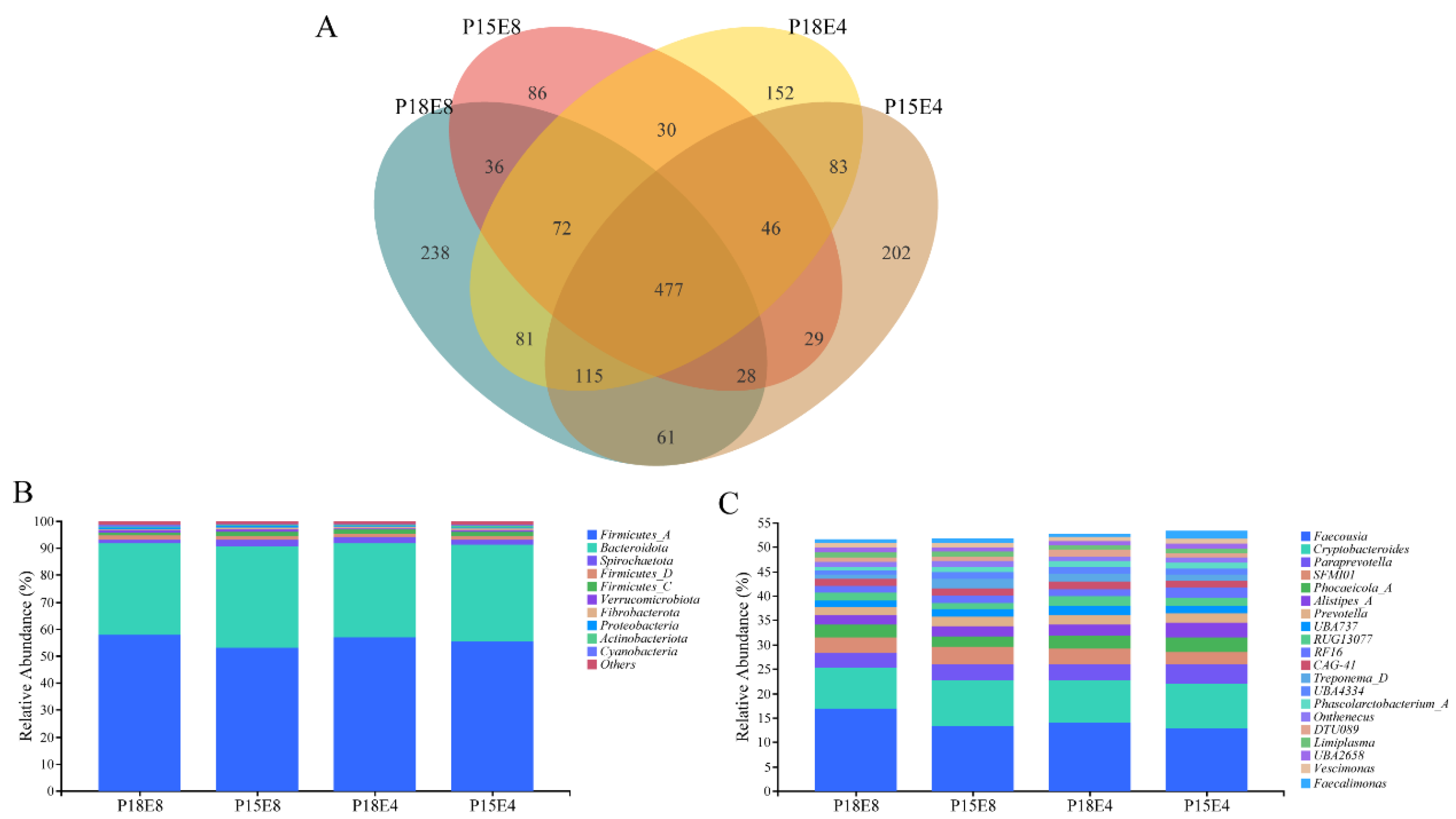
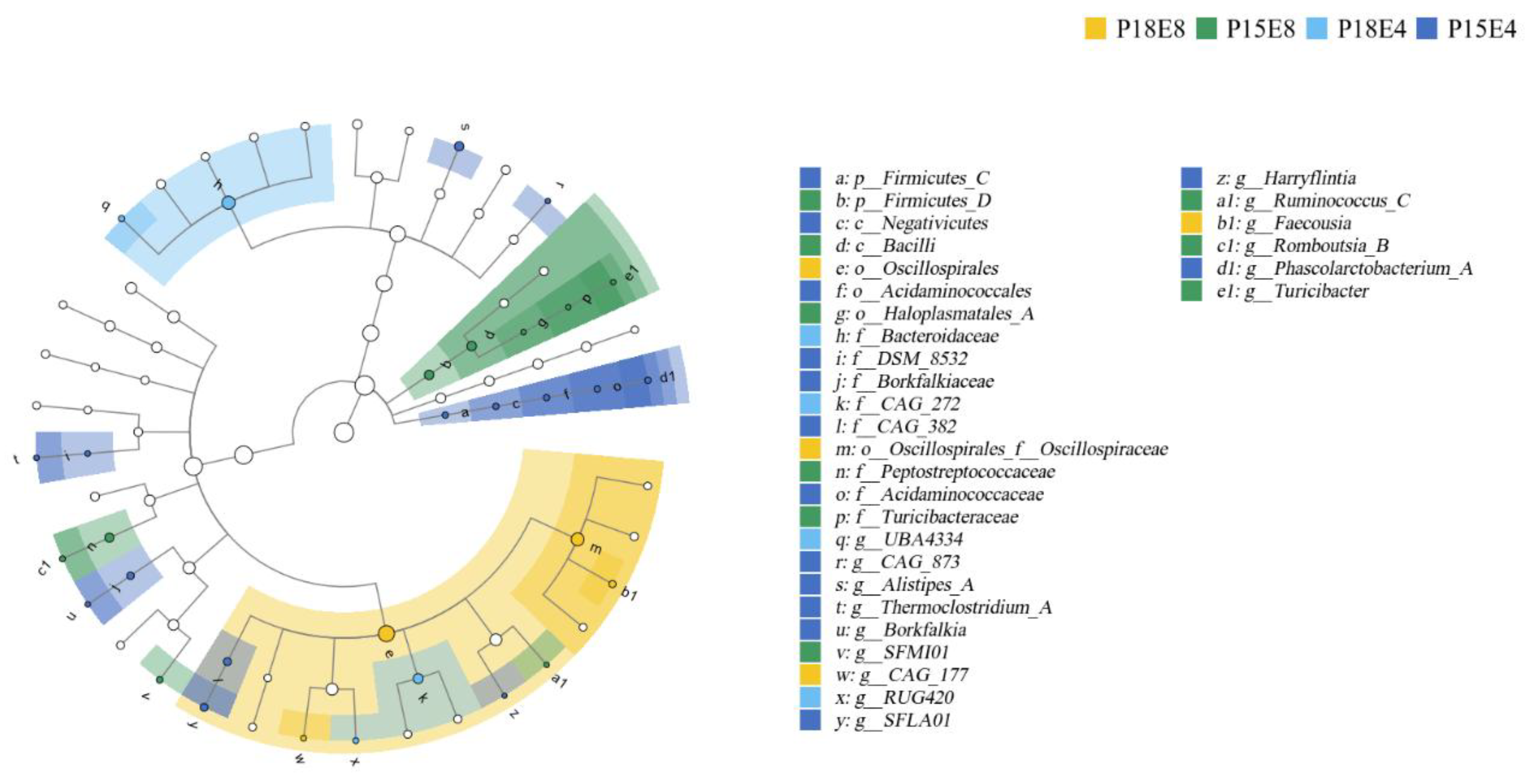
3.4. Effects of Dietary Protein and Fat Levels on Rectal Fecal Flora of Sika Deer Fawns
4. Discussion
4.1. Growth Performance
4.2. Nutrient Digestibility
4.3. Serum Biochemical Indicators
4.4. Intestinal Microbiota
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Qian, W.; Gao, X. Research Progress on Nutritional Requirements and Digestive Physiological Characteristics of Antler Deer. Chin. J. Anim. Nutr. 2020, 32, 4770–4778. [Google Scholar]
- Poławska, E.; Cooper, R.G.; Jóźwik, A.; Pomianowski, J. Meat from Alternative Species—Nutritive and Dietetic Value, and Its Benefit for Human Health—A Review. CyTA—J. Food 2013, 11, 37–42. [Google Scholar] [CrossRef]
- Liu, S. Effects of Lysine and Methionine Supplementation on Growth Performance of Weaner Sika Deer. Master’s Thesis, Jilin Agricultural University, Changchun, China, 2018. [Google Scholar]
- Li, R.; Zhang, S.; Li, Y.; Zhu, Y.; Zhao, Y.; Si, F.; Li, H.Z. Microbiota and Metabolite Profiles in the Feces of Juvenile Sika Deer (Cervus nippon) from Birth to Weaning. Animals 2024, 14, 432. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Bao, K.; Wang, X.; Shun, W.; Liu, H.; Cui, X.; Ning, W.; Li, G. Effects of Dietary Fat Level on Nutrient Digestibility, Serum Biochemical Indexes and Volatile Fatty Acid Composition in Rumen Fluid of 3-Year-Old Sika Deer during Velvet Antler Growth Period. Chin. J. Anim. Nutr. 2019, 31, 4162–4169. [Google Scholar]
- Yuan, C.; Ju, G.; Zhang, A. Research progress on nutritional requirement of skir deer. J. Inn. Mong. Agric. Univ. 2017, 38, 97–102. [Google Scholar]
- Paster, B.J.; Russell, J.B.; Yang, C.M.; Chow, J.M.; Woese, C.R.; Tanner, R. Phylogeny of the Ammonia-Producing Ruminal Bacteria Peptostreptococcus anaerobius, Clostridium sticklandii, and Clostridium aminophilum Sp. Nov. Int. J. Syst. Evol. Microbiol. 1993, 43, 107–110. [Google Scholar] [CrossRef]
- Dewhurst, R.J.; Newbold, J.R. Effect of Ammonia Concentration on Rumen Microbial Protein Production in Vitro. Br. J. Nutr. 2022, 127, 847–849. [Google Scholar] [CrossRef]
- Rotger, A.; Ferret, A.; Calsamiglia, S.; Manteca, X. Effects of Nonstructural Carbohydrates and Protein Sources on Intake, Apparent Total Tract Digestibility, and Ruminal Metabolism in Vivo and in Vitro with High-Concentrate Beef Cattle Diets. J. Anim. Sci. 2006, 84, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Atasoglu, C.; Newbold, C.J.; Wallace, R.J. Incorporation of [15N] Ammonia by the Cellulolytic Ruminal Bacteria Fibrobacter succinogenes BL2, Ruminococcus albus SY3, and Ruminococcus flavefaciens 17. Appl. Environ. Microbiol. 2001, 67, 2819–2822. [Google Scholar] [CrossRef]
- Russell, J.B.; Rychlik, J.L. Factors That Alter Rumen Microbial Ecology. Science 2001, 292, 1119–1122. [Google Scholar] [CrossRef]
- Nagaraja, T.G.; Titgemeyer, E.C. Ruminal Acidosis in Beef Cattle: The Current Microbiological and Nutritional Outlook. J. Dairy Sci. 2007, 90 (Suppl. S1), E17–E38. [Google Scholar] [PubMed]
- Dai, R.; Ma, X.; Dingkao, R.; Huang, C.; La, Y.; Li, X.; Ma, X.; Wu, X.; Chu, M.; Guo, X.; et al. Effects of Dietary Crude Protein Levels in the Concentrate Supplement after Grazing on Rumen Microbiota and Metabolites by Using Metagenomics and Metabolomics in Jersey-Yak. Front. Microbiol. 2023, 14, 1124917. [Google Scholar] [CrossRef] [PubMed]
- Sajid, Q.U.A.; Wilk, M.; Asghar, M.U. Analysis of Crude Protein Utilisation in Ruminant Rations: Supplementation of Limiting Amino Acids and Their Effect on the Environment—An Updated Review. J. Anim. Feed Sci. 2023, 33, 3–12. [Google Scholar] [CrossRef]
- Gao, X.; Wang, F. Research on suitable energy and protein levels of concentrate for one-year-old male sika deer in wintering period. Spec. Wild Econ. Anim. Plant Res. 1996, 25–27. [Google Scholar] [CrossRef]
- Gerrits, W.J.; Tolman, G.H.; Schrama, J.W.; Tamminga, S.; Bosch, M.W.; Verstegen, M.W. Effect of Protein and Protein-Free Energy Intake on Protein and Fat Deposition Rates in Preruminant Calves of 80 to 240 Kg Live Weight. J. Anim. Sci. 1996, 74, 2129–2139. [Google Scholar]
- Zanton, G.I.; Heinrichs, A.J. Analysis of Nitrogen Utilization and Excretion in Growing Dairy Cattle. J. Dairy Sci. 2008, 91, 1519–1533. [Google Scholar] [PubMed]
- Latimer, G.W., Jr.; Latimer, G.W., Jr. (Eds.) Official Methods of Analysis. In Official Methods of Analysis of AOAC International, 22nd ed.; Oxford University Press: Oxford, UK, 2023. [Google Scholar]
- Guo, Y.; Zhao, X.; Xu, D.; Sun, C.; Yang, H.; Wei, Q.; Si, Q.; Wang, H.; Zhang, K.T. Effect of Methionine Supplementation on Serum Metabolism and the Rumen Bacterial Community of Sika Deer (Cervus nippon). Animals 2022, 12, 1950. [Google Scholar] [CrossRef]
- Gao, X.; Jin, S.; Yang, F.; Jin, D. Effects of Different Levels of Dietary Energy on the Yearling Sika Deer During the Antler Growth Period. J. Econ. Anim. 1997, 1, 20–25. [Google Scholar]
- Wang, X.; Li, G.; Cui, X.; Bao, K.; Ning, H. Investigation about Appropriate Proteins of Total Mixed Rations on Young Male Sika Cavles over Winter. China Anim. Husb. Vet. Med. 2011, 38, 23–26. [Google Scholar]
- Ruvalcaba-Gómez, J.M.; Villaseñor-González, F.; Espinosa-Martínez, M.A.; Gómez-Godínez, L.J.; Rojas-Anaya, E.; Villagrán, Z.; Anaya-Esparza, L.M.; Buendía-Rodríguez, G.; Arteaga-Garibay, R.I. Growth Performance and Fecal Microbiota of Dairy Calves Supplemented with Autochthonous Lactic Acid Bacteria as Probiotics in Mexican Western Family Dairy Farming. Animals 2023, 13, 2841. [Google Scholar] [CrossRef]
- Jia, P.; Cui, K.; Ma, T.; Wan, F.; Wang, W.; Yang, D.; Wang, Y.; Guo, B.; Zhao, L.; Diao, Q. Influence of Dietary Supplementation with Bacillus Licheniformis and Saccharomyces Cerevisiae as Alternatives to Monensin on Growth Performance, Antioxidant, Immunity, Ruminal Fermentation and Microbial Diversity of Fattening Lambs. Sci. Rep. 2018, 8, 16712. [Google Scholar]
- Buttery, P.J.; Dawson, J.M. Growth Promotion in Farm Animals. Proc. Nutr. Soc. 1990, 49, 459–466. [Google Scholar] [PubMed]
- Ghasemi, E.; Azad-Shahraki, M.; Khorvash, M. Effect of Different Fat Supplements on Performance of Dairy Calves during Cold Season. J. Dairy Sci. 2017, 100, 5319–5328. [Google Scholar]
- Zinn, R.A.; Brazle, F.; White, T. Effects of Excessive Supplemental Fat on Feedlot Cattle Growth Performance and Digestive Function. Prof. Anim. Sci. 1994, 10, 66–72. [Google Scholar]
- Sahib, Q.S.; Aafaq, I.; Ahmed, H.A.; Sheikh, G.G.; Ganai, I.A. Mitigating Cold Stress in Livestock by Nutritional Interventions: A Comprehensive Review. Indian J. Anim. Res. 2024, 58, 353–363. [Google Scholar]
- Panadi, M.; Mat, K.; Rahman, M.M.; Khan, M.A.K.G.; Balakrishnan, M.; Rusli, N.D. Nutrient Intake Growth Performance and Nutrient Digestibility of Pre- and Post-Weaning Dorper Lambs Fed Varying Crude Protein Level. Trop. Anim. Health Prod. 2021, 53, 515. [Google Scholar]
- Wang, X. The Effect of the Optimal Protein Levels of Formula Feed on the Digestion and Metabolism of Sika Calves. Master’s Thesis, Jiangsu University of Science and Technology, Zhenjiang, China, 2012. [Google Scholar]
- Cho, J.; Jeong, H.; Kang, S.K.; Lee, M.; Jeon, S.; Kang, H.; Seo, S. Effects of Dietary Crude Protein Level of Concentrate Mix on Growth Performance, Rumen Characteristics, Blood Metabolites, and Methane Emissions in Fattening Hanwoo Steers. Animals 2024, 14, 469. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Dai, C.; Li, J.; Huang, P.; Li, Y.; Ding, X.; Huang, J.; Hussain, T.; Yang, H. Effects of Dietary Energy on Growth Performance, Carcass Characteristics, Serum Biochemical Index, and Meat Quality of Female Hu Lambs. Anim. Nutr. 2020, 6, 499–506. [Google Scholar]
- Garcia-Bojalil, C.M.; Staples, C.R.; Risco, C.A.; Savio, J.D.; Thatcher, W.W. Protein Degradability and Calcium Salts of Long-Chain Fatty Acids in the Diets of Lactating Dairy Cows: Productive Responses. J. Dairy Sci. 1998, 81, 1374–1384. [Google Scholar] [CrossRef]
- Zhao, T. Effect and the Initial Mechanism Research of Dietary Oil Sources on Fatty Acid and Intramucular Fat Contents of Lamb. Ph.D. Thesis, China Agricultural University, Beijing, China, 2014. [Google Scholar]
- De Souza, C.; Messana, D.; Batista, D.; Alves, L.G.C.; Titgemeyer, K.; Pires, V.; Junior, V.F.; Silva, G.; Negrão, A.; Costa, E.; et al. Effects of Protein Sources and Inclusion Levels on Nitrogen Metabolism and Urea Kinetics of Nellore Feedlot Steers Fed Concentrate-Based Diets. J. Anim. Sci. 2021, 99, skab185. [Google Scholar]
- Guangyu. Research on the Laws of Nutrition, Serum concentrations of lGF-1 and Antler Growth in sika deer and Chinese Wapiti. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2005.
- Kopchick, J.J.; Berryman, D.E.; Puri, V.; Lee, K.Y.; Jorgensen, J.O.L. The Effects of Growth Hormone on Adipose Tissue: Old Observations, New Mechanisms. Nat. Rev. Endocrinol. 2020, 16, 135–146. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, X.; Liu, H.; Brown, M.A.; Qiao, S. Dietary Protein and Gut Microbiota Composition and Function. Curr. Protein Pept. Sci. 2019, 20, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Sommer, F.; Bäckhed, F. The Gut Microbiota—Masters of Host Development and Physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef]
- Hsu, C.L.; Schnabl, B. The Gut–Liver Axis and Gut Microbiota in Health and Liver Disease. Nat. Rev. Microbiol. 2023, 21, 719–733. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, X.; Si, T.; Xu, H.; Wright, C.A.-D.G.; Li, G. Heterogeneous Development of Methanogens and the Correlation with Bacteria in the Rumen and Cecum of Sika Deer (Cervus nippon) during Early Life Suggest Different Ecology Relevance. BMC Microbiol. 2019, 19, 129. [Google Scholar]
- Castro, J.J.; Gomez, A.; White, B.; Loften, J.R.; Drackley, J.K. Changes in the Intestinal Bacterial Community, Short-Chain Fatty Acid Profile, and Intestinal Development of Preweaned Holstein Calves. 2. Effects of Gastrointestinal Site and Age. J. Dairy Sci. 2016, 99, 9703–9715. [Google Scholar] [CrossRef]
- Klein-Jöbstl, D.; Schornsteiner, E.; Mann, E.; Wagner, M.; Drillich, M.; Schmitz-Esser, S. Pyrosequencing Reveals Diverse Fecal Microbiota in Simmental Calves during Early Development. Front. Microbiol. 2014, 5, 622. [Google Scholar]
- Zhang, X.; Li, S. Research Progress on Ruminant Gut Microbiota Function and Health Control Strategy. Chin. J. Anim. Sci. 2024, 60, 1–6. [Google Scholar] [CrossRef]
- Zhen, J.; Yuan, X.; Tao, L.; Zhang, H.; Ren, Y.; Xie, S.; Wang, L.; Shen, H.; Chen, Y. Intestinal Ecology Changes in Diarrheic Père David’s Deer Revealed by Gut Microbiota and Fecal Metabolites Analysis. Animals 2022, 12, 3366. [Google Scholar] [CrossRef]
- Bai, T.; Cui, H.; Zhao, L.; Wang, Y.; Guo, X.; Zhou, X. Effects of Adding Enterococcus faecalis, Bacillus subtilis and Their Compound Bacteria on Growth Performance, Nutrient Apparent Digestibility, Rumen Fermentation Indexes and Rumen Microflora of Sheep. Chin. J. Anim. Nutr. 2022, 34, 5190–5205. [Google Scholar]
- Song, B.; Jiang, P.; Cai, F.; Gu, Z.; Gao, H.; Li, H.; Liang, B.; Qin, C.; Zhang, W. Large-Scale Metagenomic Assembly Provide New Insights into the Genetic Evolution of Gut Microbiomes in Plateau Ungulates. NPJ Biofilms Microbiomes 2024, 10, 120. [Google Scholar]
- Mackie, R.I.; Aminov, R.I.; Hu, W.; Klieve, A.V.; Ouwerkerk, D.; Sundset, M.A.; Kamagata, Y. Ecology of Uncultivated Oscillospira Species in the Rumen of Cattle, Sheep, and Reindeer as Assessed by Microscopy and Molecular Approaches. Appl. Environ. Microbiol. 2003, 69, 6808–6815. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, Y.; Wen, Z.; Liu, W.; Meng, L.; Huang, H. Oscillospira—A Candidate for the next-Generation Probiotics. Gut Microbes 2021, 13, 1987783. [Google Scholar] [CrossRef]
- Jiao, J.; Lu, Q.; Forster, R.J.; Zhou, C.; Wang, M.; Kang, J.; Tan, Z. Age and Feeding System (Supplemental Feeding versus Grazing) Modulates Colonic Bacterial Succession and Host Mucosal Immune Maturation in Goats. J. Anim. Sci. 2016, 94, 2506–2518. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Items | P18E8 | P15E8 | P18E4 | P15E4 |
|---|---|---|---|---|
| Ingredients (%) | ||||
| Corn | 29.00 | 40.00 | 33.00 | 46.00 |
| Soybean meal | 6.00 | 1.00 | 6.00 | 0.60 |
| Corn germ oil meal | 5.80 | 8.00 | 5.80 | 6.60 |
| Distillers dried grains with soluble | 9.40 | 1.20 | 9.40 | 1.00 |
| Soybean oil | 4.00 | 4.00 | 0 | 0 |
| Molasses | 3.00 | 3.00 | 3.00 | 3.00 |
| Alfalfa (CP 17%) | 40.00 | 40.00 | 40.00 | 40.00 |
| Calcium bicarbonate·2H2O | 0.60 | 0.60 | 0.60 | 0.60 |
| Mountain flour | 0.30 | 0.30 | 0.30 | 0.30 |
| NaCl | 0.50 | 0.50 | 0.50 | 0.50 |
| Baking soda | 0.70 | 0.70 | 0.70 | 0.70 |
| Premix 1 | 0.70 | 0.70 | 0.70 | 0.70 |
| Total | 100.00 | 100.00 | 100.00 | 100.00 |
| Nutrient levels (% DM) | ||||
| RDP | 9.25 | 7.19 | 9.45 | 7.18 |
| RUP | 9.35 | 8.32 | 8.61 | 8.04 |
| RDP/RUP | 0.99 | 0.86 | 1.10 | 0.89 |
| NFE | 40.36 | 44.98 | 44.67 | 45.80 |
| GE (MJ/kg DM) | 18.48 | 18.28 | 18.01 | 17.63 |
| CP | 18.60 | 15.51 | 18.06 | 15.22 |
| EE | 8.41 | 8.06 | 4.76 | 4.33 |
| CF | 11.58 | 9.54 | 10.94 | 12.47 |
| Ca | 0.83 | 0.73 | 1.03 | 0.85 |
| P | 0.52 | 0.47 | 0.61 | 0.49 |
| NDF | 49.78 | 47.02 | 48.81 | 55.62 |
| ADF | 15.81 | 13.04 | 14.28 | 17.25 |
| Items | IW, kg | FW, kg | TWG, kg | ADWG, g/d | ADFI, kg/d | F/W, % | |
|---|---|---|---|---|---|---|---|
| Individual treatment means 1 | P18E8 | 43.88 ± 1.80 | 48.61 ± 1.82 | 4.73 ± 0.38 yz | 78.85 ± 6.30 yz | 1.69 ± 0.05 | 22.50 ± 1.95 y |
| P15E8 | 43.03 ± 2.29 | 48.94 ± 1.91 | 5.91 ± 0.83 xy | 98.57 ± 13.81 xy | 1.68 ± 0.06 | 18.55 ± 1.84 y | |
| P18E4 | 42.10 ± 1.69 | 48.74 ± 1.87 | 6.64 ± 0.39 x | 110.71 ± 6.49 x | 1.68 ± 0.04 | 15.52 ± 0.95 y | |
| P15E4 | 40.83 ± 1.59 | 44.47 ± 1.79 | 3.64 ± 0.62 z | 60.71 ± 10.29 z | 1.66 ± 0.04 | 31.90 ± 4.76 x | |
| Means of main effects 2 | |||||||
| Protein level | 18% | 43.05 ± 1.73 | 48.67 ± 0.87 | 5.62 ± 0.41 | 93.72 ± 3.58 | 1.69 ± 0.05 | 19.24 ± 1.56 b |
| 15% | 41.93 ± 1.13 | 46.71 ± 1.25 | 4.78 ± 0.20 | 79.64 ± 6.66 | 1.67 ± 0.04 | 25.22 ± 1.49 a | |
| Fat level | 8% | 43.48 ± 1.37 | 48.77 ± 0.92 | 5.28 ± 0.37 | 88.06 ± 4.89 | 1.69 ± 0.08 | 20.65 ± 1.44 |
| 4% | 41.46 ± 1.22 | 46.61 ± 1.15 | 5.14 ± 0.40 | 85.71 ± 3.93 | 1.67 ± 0.06 | 23.71 ± 2.18 | |
| p-values | Protein | 0.574 | 0.298 | 0.125 | 0.125 | 0.883 | 0.031 |
| Fat | 0.296 | 0.252 | 0.756 | 0.756 | 0.882 | 0.253 | |
| Interaction | 0.911 | 0.226 | 0.001 | 0.001 | 0.925 | <0.001 |
| Items | Nutrients Apparent Digestibility % | |||||||
|---|---|---|---|---|---|---|---|---|
| DM | CP | EE | Ca | P | NDF | ADF | ||
| Individual treatment means 1 | P18E8 | 75.44 ± 0.88 x | 65.08 ± 1.39 y | 88.53 ± 0.57 w | 39.58 ± 1.12 x | 73.37 ± 2.38 w | 61.83 ± 2.85 x | 18.85 ± 1.34 x |
| P15E8 | 75.65 ± 0.34 x | 69.86 ± 1.16 x | 89.04 ± 0.56 w | 35.53 ± 1.45 y | 71.62 ± 1.67 w | 60.41 ± 1.96 x | 17.53 ± 1.54 x | |
| P18E4 | 79.47 ± 0.66 w | 80.46 ± 0.58 w | 89.18 ± 0.31 w | 56.13 ± 0.94 w | 75.92 ± 1.55 w | 75.10 ± 0.71 w | 29.18 ± 1.43 w | |
| P15E4 | 72.09 ± 0.16 y | 58.14 ± 0.95 z | 86.30 ± 0.87 x | 30.17 ± 1.72 z | 51.86 ± 4.62 x | 52.71 ± 1.10 y | 14.09 ± 1.01 y | |
| Means of main effects 2 | ||||||||
| Protein level | 18% | 77.46 ± 0.18 a | 72.77 ± 1.46 a | 88.85 ± 0.30 a | 47.86 ± 1.40 a | 74.65 ± 0.65 a | 68.46 ± 0.29 a | 24.02 ± 1.11 a |
| 15% | 73.87 ± 0.25 b | 64.00 ± 0.71 b | 87.67 ± 0.92 b | 32.85 ± 0.15 b | 61.74 ± 1.94 b | 56.56 ± 0.90 b | 15.81 ± 0.98 b | |
| Fat level | 8% | 75.55 ± 0.54 | 67.47 ± 0.28 | 88.78 ± 0.57 A | 37.56 ± 0.11 B | 72.50 ± 0.68 A | 61.12 ± 0.82 B | 18.19 ± 0.41 B |
| 4% | 75.78 ± 0.52 | 69.30 ± 1.73 | 87.74 ± 0.72 B | 43.15 ± 1.95 A | 63.89 ± 1.60 B | 63.90 ± 1.84 A | 21.63 ± 1.43 A | |
| p-values | Protein | <0.001 | <0.001 | 0.003 | <0.001 | <0.001 | <0.001 | <0.001 |
| Fat | 0.598 | 0.097 | 0.002 | <0.001 | 0.005 | <0.001 | <0.001 | |
| Interaction | <0.001 | <0.001 | 0.022 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Items | TG, mmol/L | CHO, mmol/L | HDL-C, mmol/L | LDL-C, mmol/L | TP, g/L | ALB, g/L | GLU, mmol/L | Urea, mmol/L | |
|---|---|---|---|---|---|---|---|---|---|
| Individual treatment means 1 | P18E8 | 0.16 ± 0.02 | 1.90 ± 0.07 z | 1.11 ± 0.06 z | 0.27 ± 0.03 | 59.29 ± 0.41 | 31.23 ± 0.34 | 9.31 ± 1.33 | 10.12 ± 0.38 y |
| P15E8 | 0.18 ± 0.02 | 2.44 ± 0.10 y | 1.56 ± 0.08 y | 0.28 ± 0.04 | 58.94 ± 0.69 | 30.93 ± 1.16 | 10.62 ± 0.43 | 9.65 ± 0.27 y | |
| P18E4 | 0.16 ± 0.03 | 1.74 ± 0.08 z | 0.99 ± 0.05 z | 0.25 ± 0.03 | 62.01 ± 1.18 | 32.39 ± 0.45 | 10.83 ± 0.34 | 7.06 ± 0.13 z | |
| P15E4 | 0.14 ± 0.01 | 1.60 ± 0.17 z | 0.94 ± 0.11 z | 0.22 ± 0.03 | 57.56 ± 2.04 | 28.87 ± 2.04 | 8.68 ± 0.55 | 10.09 ± 0.47 y | |
| Means of main effects 2 | |||||||||
| Protein level | 18% | 0.16 ± 0.03 | 1.82 ± 0.07 | 1.05 ± 0.07 b | 0.26 ± 0.03 | 60.65 ± 0.80 | 31.81 ± 0.52 | 10.07 ± 0.79 | 8.59 ± 0.13 b |
| 15% | 0.16 ± 0.03 | 2.02 ± 0.08 | 1.25 ± 0.04 a | 0.25 ± 0.01 | 58.25 ± 1.22 | 29.90 ± 1.18 | 9.65 ± 0.66 | 9.87 ± 0.13 a | |
| Fat level | 8% | 0.17 ± 0.03 | 2.17 ± 0.06 A | 1.34 ± 0.03 A | 0.28 ± 0.04 | 59.12 ± 0.19 | 31.08 ± 0.24 | 9.97 ± 0.77 | 9.89 ± 0.17 A |
| 4% | 0.15 ± 0.02 | 1.67 ± 0.05 B | 0.96 ± 0.04 B | 0.23 ± 0.03 | 59.79 ± 1.12 | 30.63 ± 0.67 | 9.76 ± 0.68 | 8.57 ± 0.17 B | |
| p-values | Protein | 0.764 | 0.083 | 0.026 | 0.725 | 0.065 | 0.194 | 0.784 | <0.001 |
| Fat | 0.284 | <0.001 | <0.001 | 0.183 | 0.112 | 0.125 | 0.594 | <0.001 | |
| Interaction | 0.371 | 0.004 | 0.005 | 0.542 | 0.596 | 0.711 | 0.133 | <0.001 |
| Items | GH, ng/mL | IGF, ng/mL | |
|---|---|---|---|
| Individual treatment means 1 | P18E8 | 5.63 ± 0.11 y | 119.27 ± 2.68 |
| P15E8 | 4.91 ± 0.13 z | 117.69 ± 3.86 | |
| P18E4 | 4.77 ± 0.16 z | 114.51 ± 2.84 | |
| P15E4 | 4.72 ± 0.15 z | 114.17 ± 1.68 | |
| Means of main effects 2 | |||
| Protein level | 18% | 5.20 ± 0.05 a | 116.89 ± 1.45 |
| 15% | 4.82 ± 0.04 b | 115.93 ± 1.94 | |
| Fat level | 8% | 5.27 ± 0.05 A | 118.48 ± 1.41 |
| 4% | 4.75 ± 0.04 B | 114.34 ± 1.65 | |
| p-values | Protein | 0.011 | 0.745 |
| Fat | <0.001 | 0.835 | |
| Interaction | 0.024 | 0.172 |
| Items | Chao1 | Observed Species | Shannon | |
|---|---|---|---|---|
| Individual treatment means 1 | P18E8 | 2814.07 ± 121.83 | 2716.21 ± 131.95 | 10.14 ± 0.16 |
| P15E8 | 2541.11 ± 324.36 | 2447.47 ± 309.52 | 9.91 ± 0.24 | |
| P18E4 | 2713.96 ± 222.28 | 2604.11 ± 206.98 | 10.10 ± 0.14 | |
| P15E4 | 2639.61 ± 386.87 | 2510.05 ± 384.57 | 9.98 ± 0.37 | |
| Means of main effects 2 | ||||
| Protein level | 18% | 2764.01 ± 180.71 | 2660.16 ± 177.39 | 10.12 ± 0.15 a |
| 15% | 2587.46 ± 347.35 | 2476.92 ± 337.10 | 9.94 ± 0.30 b | |
| Fat level | 8% | 2669.56 ± 280.75 | 2573.94 ± 273.19 | 10.02 ± 0.23 |
| 4% | 2676.78 ± 307.21 | 2557.08 ± 302.27 | 10.04 ± 0.27 | |
| p-values | Protein | 0.090 | 0.070 | 0.045 |
| Fat | 0.994 | 0.799 | 0.863 | |
| Interaction | 0.324 | 0.373 | 0.503 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Z.; Tian, J.; Zhang, Q.; Sun, H.; Jiang, Q.; Zhang, T. Effects of Dietary Protein and Fat Levels on Growth Performance, Nutrient Digestibility, Serum Indexes, and Rectal Fecal Microbiota of Sika Deer (Cervus nippon) Fawns in Early Wintering Period. Animals 2025, 15, 908. https://doi.org/10.3390/ani15070908
Gao Z, Tian J, Zhang Q, Sun H, Jiang Q, Zhang T. Effects of Dietary Protein and Fat Levels on Growth Performance, Nutrient Digestibility, Serum Indexes, and Rectal Fecal Microbiota of Sika Deer (Cervus nippon) Fawns in Early Wintering Period. Animals. 2025; 15(7):908. https://doi.org/10.3390/ani15070908
Chicago/Turabian StyleGao, Zuer, Jiaxin Tian, Qiaoru Zhang, Haoran Sun, Qingkui Jiang, and Tietao Zhang. 2025. "Effects of Dietary Protein and Fat Levels on Growth Performance, Nutrient Digestibility, Serum Indexes, and Rectal Fecal Microbiota of Sika Deer (Cervus nippon) Fawns in Early Wintering Period" Animals 15, no. 7: 908. https://doi.org/10.3390/ani15070908
APA StyleGao, Z., Tian, J., Zhang, Q., Sun, H., Jiang, Q., & Zhang, T. (2025). Effects of Dietary Protein and Fat Levels on Growth Performance, Nutrient Digestibility, Serum Indexes, and Rectal Fecal Microbiota of Sika Deer (Cervus nippon) Fawns in Early Wintering Period. Animals, 15(7), 908. https://doi.org/10.3390/ani15070908