

Identification of Hybrid Sturgeon (Acipenser baerii × Acipenser schrenckii) from Their Parents Using Germplasm

 , , , and
, , , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Sampling
2.2. Sequencing and Genotyping
2.3. Statistical Analyses
3. Results
3.1. Genetic Variation Among the Hybrid Sturgeon and Their Parents
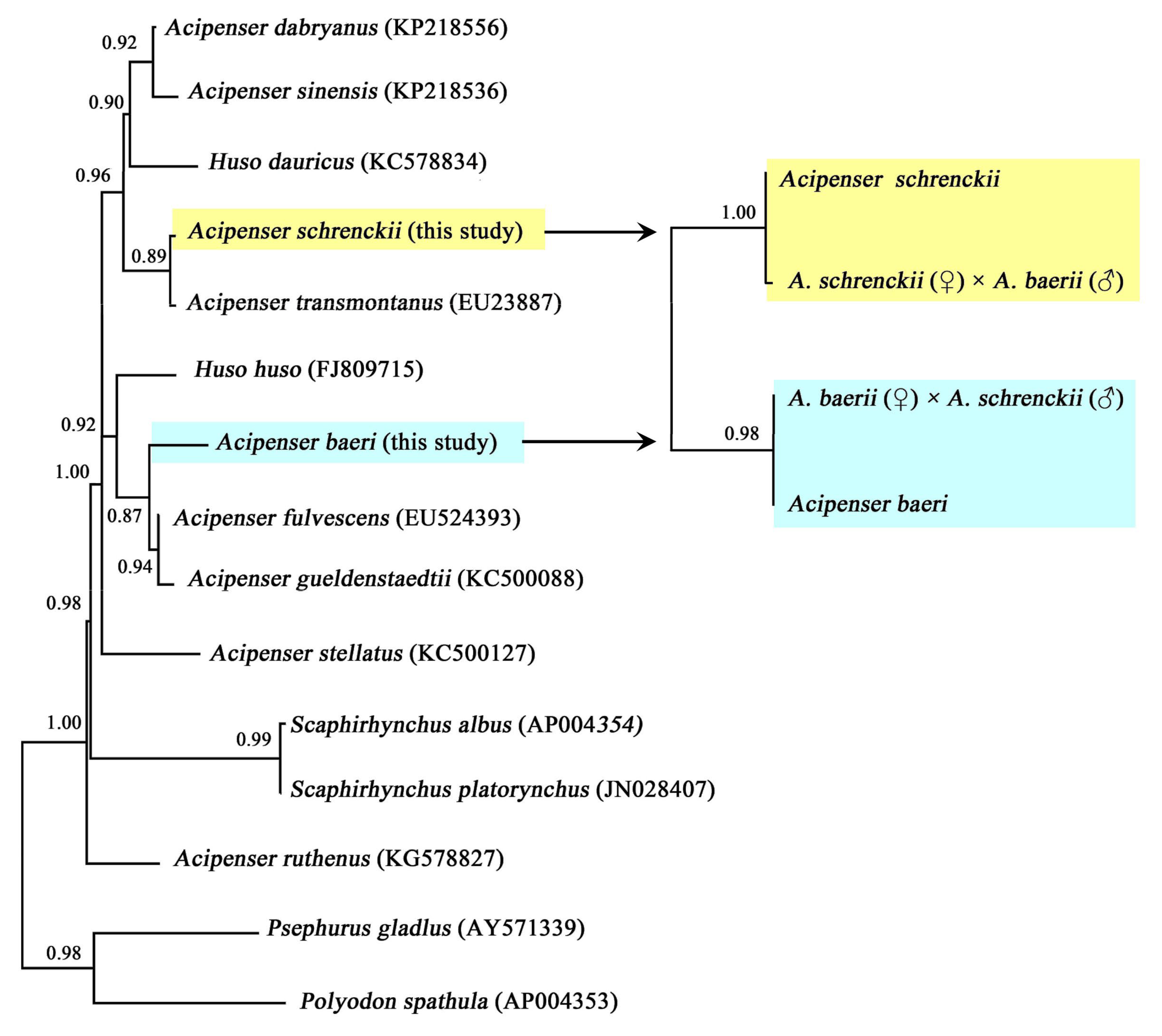
3.2. The Hybrid Sturgeons Sepearted from Their Male Parent by Phylogenetic Analysis
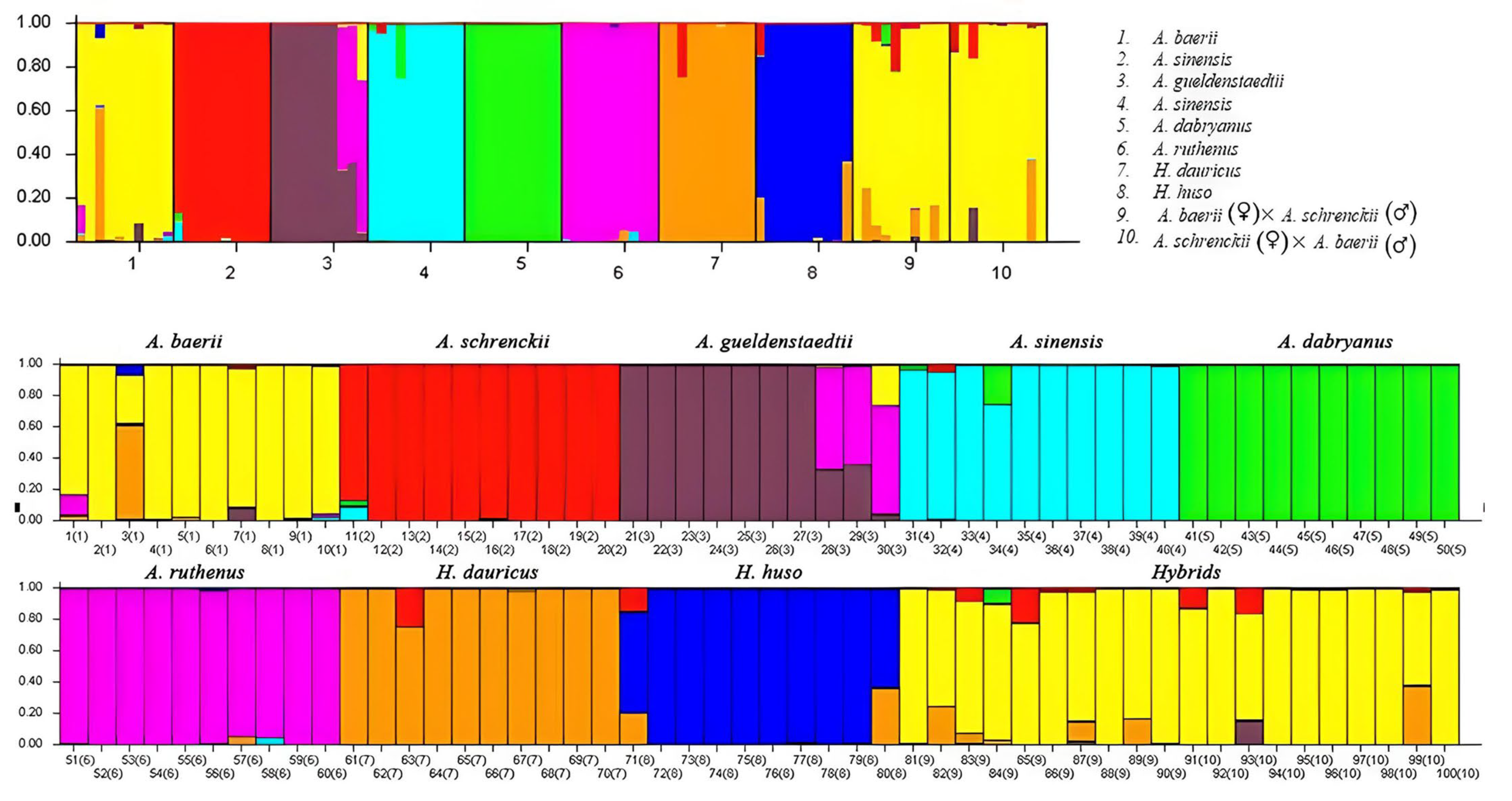
3.3. The Hybrid Sturgeons Were Separated from A. sinesis by Structure Analysis
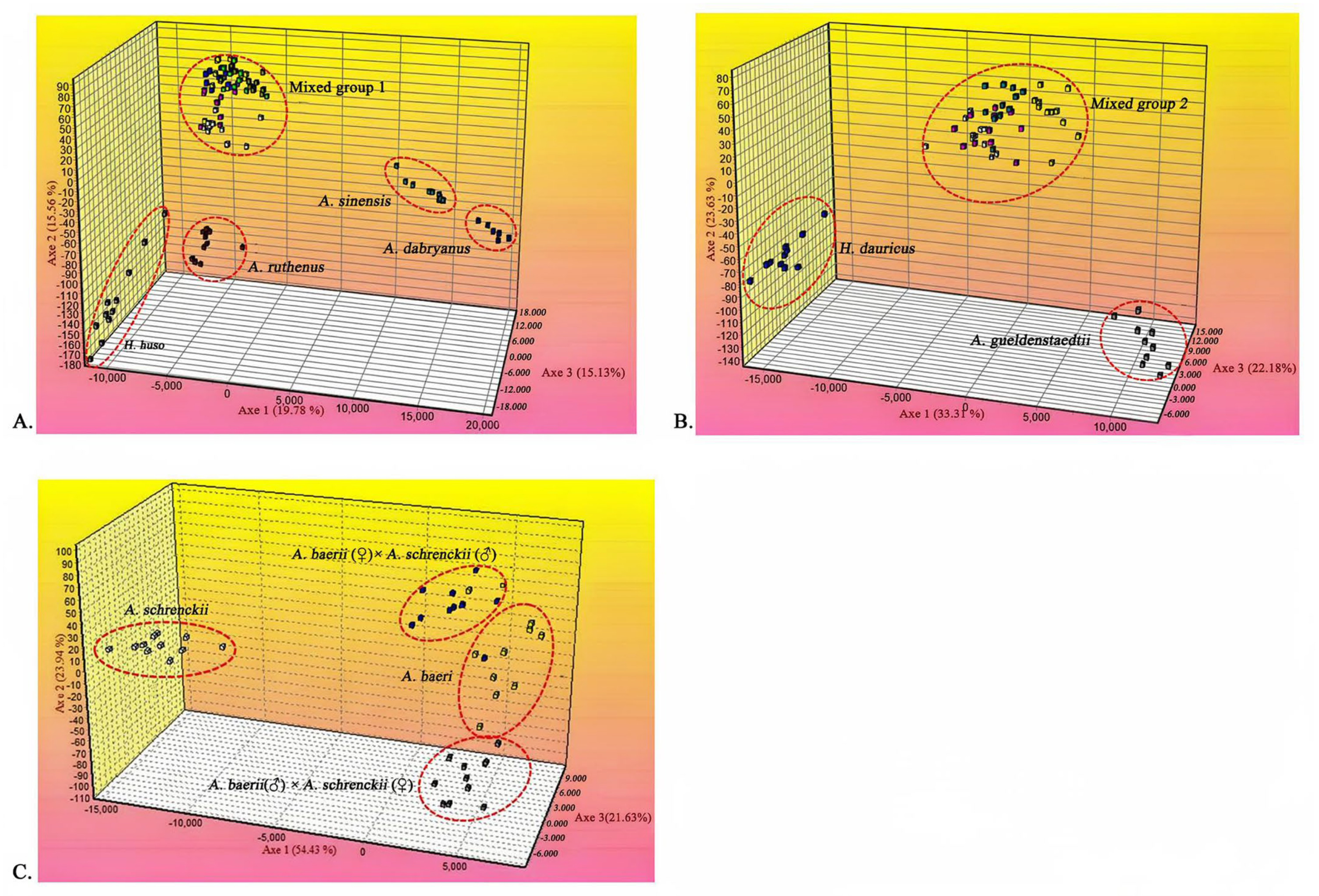
3.4. The Hybrid Sturgeons Were Successfully Separated from Their Parents by FCA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, J.; Ma, J.; Wu, H. The current situation and development trend of Chinese sturgeon caviar industry. China Fish. 2011, 7, 25–27. [Google Scholar]
- Quattro, M.; Greig, W.; Coykendall, K.; Bowen, W.; Baldwin, D. Genetic issues in aquatic species management: The shortnose sturgeon (Acipenser brevirostrum) in the southeastern united states. Conserv. Genet. 2002, 3, 155–166. [Google Scholar]
- Farasati, S.; Khoshkholgh, M.; Yarmohammadi, M. Genetic diversity of cultured beluga sturgeon (Huso huso) brood stocks by using microsatellite method. Aquat. Physiol. Biotechnol. 2020, 8, 55–72. [Google Scholar]
- Schwartz, S.; Beheregaray, B. Using genotype simulations and Bayesian analyses to identify individuals of hybrid origin in Australian bass: Lessons for fisheries management. J. Fish Biol. 2010, 72, 435–450. [Google Scholar]
- Jobling, M. Salmonid fishes: Population biology, genetics and management. Aquaculture 2002, 204, 180–182. [Google Scholar]
- Huang, X.; Zhong, L.; Fan, W.; Feng, Y.; Xiong, G.; Liu, S.; Wang, K.; Geng, Y.; Ouyang, P.; Chen, D.; et al. Enteritis in hybrid sturgeon (Acipenser schrenckii♂×Acipenser baeri♀) caused by intestinal microbiota disorder. Aquac. Rep. 2020, 18, 100456. [Google Scholar]
- Krykhtin, L.; Svirskii, G. Endemic sturgeons of the Amur River: Kaluga, Huso dauricus, and Amur sturgeon, Acipenser schrenckii. Environ. Biol. Fishes 1997, 48, 231–239. [Google Scholar]
- Birstein, J.; Doukakis, P.; Sorkin, B.; Desalle, R. Population aggregation analysis of three caviar-producing species of sturgeons and implications for the species identification of black caviar. Conserv. Biol. 1998, 12, 766–775. [Google Scholar]
- Eenennaam, V.; Eenennaam, V.; Medrano, F.; Doroshov, I. Rapid verification of meiotic gynogenesis and polyploidy in white sturgeon (Acipenser transmontanus Richardson). Aquaculture 1996, 147, 177–189. [Google Scholar]
- Chelomina, N.; Rozhkovan, V.; Ivanov, A. Discrimination of interspecific hybrids in natural populations of amur sturgeon fishes using multilocus RAPD-PCR markers. Cytol. Genet. 2008, 42, 342–350. [Google Scholar] [CrossRef]
- Costa, O.; Jeremy, R.; Boutillier, A.; Ratnasingham, S.; Dooh, R.; Hajibabaei, M. Biological identifications through DNA barcodes: The case of the crustacea. Can. J. Fish. Aquat. Sci. 2007, 64, 272–295. [Google Scholar]
- Havelka, M.; Arai, K.; Boscari, E.; Congiu, L.; Sergeev, A.; Mugue, N. A new marker, isolated by ddRAD sequencing, detects Siberian and Russian sturgeon in hybrids. Anim. Genet. 2019, 50, 115–116. [Google Scholar] [PubMed]
- Havelka, M.; Fujimoto, T.; Hagihara, S.; Adachi, S.; Arai, K. Nuclear DNA markers for identification of Beluga and Sterlet sturgeons and their interspecific Bester hybrid. Sci. Rep. 2017, 7, 1694. [Google Scholar]
- Boscari, E.; Barmintseva, A.; Zhang, S.; Yue, H.; Li, C.; Shedko, S.V.; Lieckfeldt, D.; Ludwig, A.; Wei, Q.W.; Mugue, N.S.; et al. Genetic identification of the caviar-producing Amur and Kaluga sturgeons revealed a high level of concealed hybridization. Food Control 2017, 82, 243–250. [Google Scholar]
- Yan, X.; Dong, Y.; Dong, T.; Song, H.; Wang, W.; Hu, H. InDel DNA Markers Potentially Unique to Kaluga Sturgeon Huso dauricus Based on Whole-Genome Resequencing Data. Diversity 2023, 15, 689. [Google Scholar] [CrossRef]
- Durand, J.D.; Hubert, N.; Shen, K.N.; Borsa, P. DNA barcoding grey mullets. Rev. Fish Biol. Fish. 2017, 27, 1–11. [Google Scholar] [CrossRef]
- Ghadirneja, H.; Aziza, S.; Salehi, A.; Aghili, K.; Kamaruddin, K.; Jamsari, A.; Chiun, L. Barcoding of five sturgeon species in Iran. J. Mol. Genet. 2009, 2, 29–34. [Google Scholar]
- Hu, J.; Wang, D.; Wei, Q.; Shen, L. Molecular identification of amur sturgeon (Acipenser schrenckii), kaluga (Huso dauricus) and their reciprocal hybrids. J. Fish. Sci. China 2010, 17, 21–30. [Google Scholar]
- Esselink, D.; Nybom, H.; Vosman, B. Assignment of allelic configuration in polyploids using the MAC-PR (microsatellite DNA allele counting-peak ratios) method. Theor. Appl. Genet. 2004, 109, 402–408. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731. [Google Scholar]
- Rozas, J.; Sánchez-Delbarrio, C.; Messeguer, X.; Rozas, R. DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics 2003, 19, 2496–2497. [Google Scholar] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MrBayes: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar]
- Zhu, H.; Hu, H.; Wang, W.; Dong, Y.; Yao, Z.; Li, L. Comparative experiment on seed cultivation between Siberian sturgeon and hybrid sturgeon. China Fish. 2014, 8, 73–74. [Google Scholar]
- Qi, X.; Liu, H.; Li, Z. Comparison of reproductive performance, growth performance, and disease resistance of amur sturgeon, siberian sturgeon, and their hybrid. J. Jiangxi Agric. Univ. 2017, 39, 376–383. [Google Scholar]
- Wang, N.; Yang, G.; Peng, T.; Qu, Q.; Sun, D.; Wu, W.; Wang, G.; Zhang, Y.; Liu, X. Comparison of offspring survival of three kinds of sturgeons purebreds and crossbreds. J. Sichuan Agric. Univ. 2010, 28, 507–511. [Google Scholar]
- Hebert, D.; Ratnasingham, S.; Dewaard, R. Barcoding animal life: Cytochrome C oxidase subunit I divergences among closely related species. Proc. R. Soc. B Biol. Sci. 2003, 270, 96–99. [Google Scholar]
- Pritchard, K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Ludwig, A.; Kirschbaum, F. Comparison of mitochondrial DNA sequences between the european and the adriatic sturgeon. J. Fish Biol. 1998, 52, 1289–1291. [Google Scholar]
- Zhang, S.; Deng, H.; Wang, D.; Wu, Q. Study on the genetic relationships of seven species of Acipenseriformes by random amplified polymorphic DNA. Prog. Nat. Sci. 1999, 9, 818–823. [Google Scholar]
- Gaston, K.J.; Oneill, M.A. Automated species identification: Why not? Philos. Trans. Biol. Sci. 2004, 359, 655–667. [Google Scholar]
- Iswarya-Deepti, V.; Kandula, S.; Khedkar, G.D. DNA barcoding of five species of groupers (Pisces: Serranidae) off Visakhapatnam, central eastern coast of India. Mitochondrial DNA A DNA Mapp. Seq. Anal. 2018, 29, 659–663. [Google Scholar] [PubMed]
- Wu, W. DNA Barcoding Studing and Molecular Identification of Sturgeons and Their Hybrids; Inner Mongolia Agricultural University: Hohhot, China, 2010. [Google Scholar]
- May, B.; Krueger, C.C.; Kincaid, H.L. Genetic variation at microsatellite loci in sturgeon: Primer sequence homology in Acipenser and Scaphirhynchus. Can. J. Fish. Aquat. Sci. 1997, 54, 1542–1547. [Google Scholar] [CrossRef]
- Jenneckens, I.; Meyer, J.N.; Horstgen-Schwark, G.; May, B.; Debus, L.; Wedekind, H.; Ludwing, A. A fixed allele at microsatellite locus LS-39 exhibiting species-specificity for the black caviar producer Acipenser stellatus. J. Appl. Ichthyol. 2010, 17, 39–42. [Google Scholar] [CrossRef]
- Chassaing, O.; Hnni, C.; Beerbi, P. Distinguishing species of european sturgeons Acipenser spp. using microsatellite allele sequences. J. Fish Biol. 2011, 78, 208–226. [Google Scholar]
- Dudu, A.; Suciu, R.; Paraschiv, M.; Emil Georgescu, S.; Costache, M.; Berrebi, P. Nuclear markers of Danube sturgeons hybridization. Int. J. Mol. Sci. 2011, 12, 6796–6809. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, C.; Li, C.; Cheng, L.; Wu, W.; Zhao, Y.; Sun, X. Genetic identification of amur sturgeon, kaluga and their hybrids based on microsatellite markers. Chin. J. Fish. 2015, 28, 18–23. [Google Scholar]
- Zhang, S.; Yang, H.; Xin, M.; Wu, J.; Dai, Z.; Du, H.; Liu, Z.; Wei, Q. External morphology and molecular identification of wild juvenile Acipenser sinensis newly found in the Jiangsu Xupu section of the Yangtze River in 2015. J. Fish. Sci. China 2016, 23, 1–9. [Google Scholar]
- Fain, R.; Straughan, J.; Hamlin, B.; Hoesch, M.; Lemay, P. Forensic genetic identification of sturgeon caviars traveling in world trade. Conserv. Genet. 2013, 14, 855–874. [Google Scholar]
- Zhang, X.; Wu, W.; Li, L.; Ma, X.; Chen, J. Genetic variation and relationships of seven sturgeon species and ten interspecific hybrids. Genet. Sel. Evol. 2013, 45, 21. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Microsatellite Markers | Primer Sequences | Repetitive Sequence | Allele Size Range (bp) | Origin of the Primers |
|---|---|---|---|---|
| Ab3 | F:CGCTTCACAGATGTCTCAGTCAGTT | CA | 195–286 | This study |
| R:TCCTGAGTGGGGTTGTTCAATAAGA | ||||
| Ab5 | F:TTTTCTCCACGAATGCCC | CA | 107–175 | This study |
| R:TCAAAGTCAAGCCAAAGC | ||||
| As1 | F: AACAAGCGACGAACAGTGTG | ATTG | 242–301 | This study |
| R: CACAAATCGGACAGAAAGCA | ||||
| LS19 | F: CATCTTAGCCGTCTGTGGTAC | TTG | 116–153 | Hu et al. [18] |
| R: CAGGTCCCTAATACAATGGC | ||||
| HLJSX7 | F: GAAAGGACACCAGCAGTG | GT | 212–254 | Hu et al. [18] |
| R: AACCCATTAACAATTACAGC | ||||
| HLJSX30 | F: GGGGAGAAAACTGGGGTAAA | CTAT | 180–284 | Hu et al. [18] |
| R: CACGTGGATGCGAGAAATAC |
| Species | T% | C% | A% | G% | C | V | Pi | S |
|---|---|---|---|---|---|---|---|---|
| A. schrenckii | 26.0 | 30.5 | 22.5 | 21.0 | 659 | 25 | 3 | 22 |
| A. baeri (♂) × A. schrenckii (♀) | 25.8 | 30.7 | 22.2 | 21.3 | 662 | 1 | 0 | 1 |
| A. baeri | 26.7 | 30.2 | 22.8 | 20.3 | 661 | 0 | 0 | 0 |
| A. baeri (♀) × A. schrenckii (♂) | 26.7 | 30.2 | 22.8 | 20.3 | 660 | 0 | 0 | 0 |
| Species | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | |
| 2 | 0.0034 | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | |
| 3 | 0.0298 | 0.0298 | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | |
| 4 | 0.0595 | 0.0595 | 0.0520 | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | |
| 5 | 0.0501 | 0.0501 | 0.0427 | 0.0428 | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | |
| 6 | 0.0652 | 0.0690 | 0.0575 | 0.0614 | 0.0209 | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | |
| 7 | 0.0596 | 0.0633 | 0.0557 | 0.0595 | 0.0521 | 0.0596 | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | |
| 8 | 0.0446 | 0.0446 | 0.0282 | 0.0617 | 0.0541 | 0.0654 | 0.0636 | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | |
| 9 | 0.0464 | 0.0501 | 0.0426 | 0.0539 | 0.0354 | 0.0501 | 0.0521 | 0.0540 | n/c | n/c | n/c | n/c | n/c | n/c | n/c | n/c | |
| 10 | 0.1298 | 0.1341 | 0.1274 | 0.1317 | 0.1391 | 0.1366 | 0.1280 | 0.1391 | 0.1392 | n/c | n/c | n/c | n/c | n/c | n/c | n/c | |
| 11 | 0.1463 | 0.1463 | 0.1354 | 0.1403 | 0.1403 | 0.1514 | 0.1400 | 0.1449 | 0.1493 | 0.1041 | n/c | n/c | n/c | n/c | n/c | n/c | |
| 12 | 0.1018 | 0.1018 | 0.0978 | 0.1040 | 0.1040 | 0.1124 | 0.1078 | 0.1021 | 0.1017 | 0.1928 | 0.1694 | n/c | n/c | n/c | n/c | n/c | |
| 13 | 0.0996 | 0.0996 | 0.0956 | 0.1017 | 0.1017 | 0.1102 | 0.1055 | 0.0999 | 0.0994 | 0.1899 | 0.1667 | 0.0017 | 0.0016 | n/c | n/c | n/c | |
| 14 | 0.0341 | 0.0341 | 0.0075 | 0.0470 | 0.0470 | 0.0657 | 0.0639 | 0.0360 | 0.0470 | 0.1325 | 0.1363 | 0.1046 | 0.1024 | 0.0000 | n/c | n/c | |
| 15 | 0.0334 | 0.0334 | 0.0069 | 0.0463 | 0.0463 | 0.0650 | 0.0633 | 0.0354 | 0.0463 | 0.1339 | 0.1376 | 0.1019 | 0.0997 | 0.0040 | 0.0000 | n/c | |
| 16 | 0.0483 | 0.0483 | 0.0445 | 0.0052 | 0.0052 | 0.0262 | 0.0539 | 0.0559 | 0.0372 | 0.1391 | 0.1360 | 0.1060 | 0.1038 | 0.0488 | 0.0482 | 0.0000 | |
| 17 | 0.0483 | 0.0483 | 0.0445 | 0.0052 | 0.0052 | 0.0262 | 0.0539 | 0.0559 | 0.0372 | 0.1391 | 0.1360 | 0.1060 | 0.1038 | 0.0488 | 0.0482 | 0.0000 |
| Specie | Number of Alleles (A) | ||||||
|---|---|---|---|---|---|---|---|
| As1 | Ab3 | Ab5 | LS19 | HLJSX07 | HLJSX30 | Average | |
| A. baerii | 6 | 4 | 5 | 6 | 5 | 6 | 5.3 |
| A. schrenckii | 1 | 4 | 3 | 3 | 4 | 4 | 3.2 |
| A. baerii (♀) × A. schrenckii (♂) | 6 | 5 | 4 | 5 | 3 | 4 | 4.5 |
| A. baerii (♂) × A. schrenckii (♀) | 5 | 7 | 4 | 4 | 3 | 5 | 4.7 |
| H. dauricus | 8 | 4 | 3 | 2 | 1 | 6 | 4.0 |
| H. huso | 6 | 6 | 5 | 6 | 2 | 4 | 4.8 |
| A. gueldenstaedtii | 5 | 4 | 3 | 6 | 3 | 3 | 4.0 |
| A. sinensis | 3 | 7 | 4 | 3 | 1 | 3 | 3.5 |
| A. dabryanus | 2 | 2 | 2 | 2 | 1 | 2 | 1.8 |
| A. ruthenus | 11 | 12 | 4 | 1 | 2 | 7 | 6.2 |
| Total number | 53 | 55 | 37 | 38 | 25 | 44 | 42.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Zhao, Z.; Xu, Z.; Liu, Y.; Jiang, M.; Fu, L.; Zhang, J.; Jing, Z.; Pang, X.; Shao, W.; et al. Identification of Hybrid Sturgeon (Acipenser baerii × Acipenser schrenckii) from Their Parents Using Germplasm. Animals 2025, 15, 907. https://doi.org/10.3390/ani15070907
Yang S, Zhao Z, Xu Z, Liu Y, Jiang M, Fu L, Zhang J, Jing Z, Pang X, Shao W, et al. Identification of Hybrid Sturgeon (Acipenser baerii × Acipenser schrenckii) from Their Parents Using Germplasm. Animals. 2025; 15(7):907. https://doi.org/10.3390/ani15070907
Chicago/Turabian StyleYang, Shiyong, Zhongmeng Zhao, Zihan Xu, Ya Liu, Menghao Jiang, Lin Fu, Jin Zhang, Zhaoxin Jing, Xiaojian Pang, Wuyuntana Shao, and et al. 2025. "Identification of Hybrid Sturgeon (Acipenser baerii × Acipenser schrenckii) from Their Parents Using Germplasm" Animals 15, no. 7: 907. https://doi.org/10.3390/ani15070907
APA StyleYang, S., Zhao, Z., Xu, Z., Liu, Y., Jiang, M., Fu, L., Zhang, J., Jing, Z., Pang, X., Shao, W., Zhang, C., Li, Y., Du, X., & Wu, J. (2025). Identification of Hybrid Sturgeon (Acipenser baerii × Acipenser schrenckii) from Their Parents Using Germplasm. Animals, 15(7), 907. https://doi.org/10.3390/ani15070907