
Comprehensive Phenotypic Characterization and Genomic Analysis Unveil the Probiotic Potential of Bacillus velezensis K12



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation, Purification, and Characterization of Strains
2.2. Determination of Strain Biology
2.2.1. Determination of the Growth Curve of Strain K12 Versus the pH Change Curve
2.2.2. Hemolysis Analysis
2.2.3. Acid Resistance Analysis
2.2.4. Bile Resistance Analysis
2.2.5. Determination of Tolerance to Simulated Gastrointestinal Fluids
2.2.6. Antibiotic Susceptibility Testing
2.2.7. Determination of Bacteriostatic Capacity
2.2.8. Whole-Genome Sequencing and Annotation
2.3. Statistical Analysis
3. Results
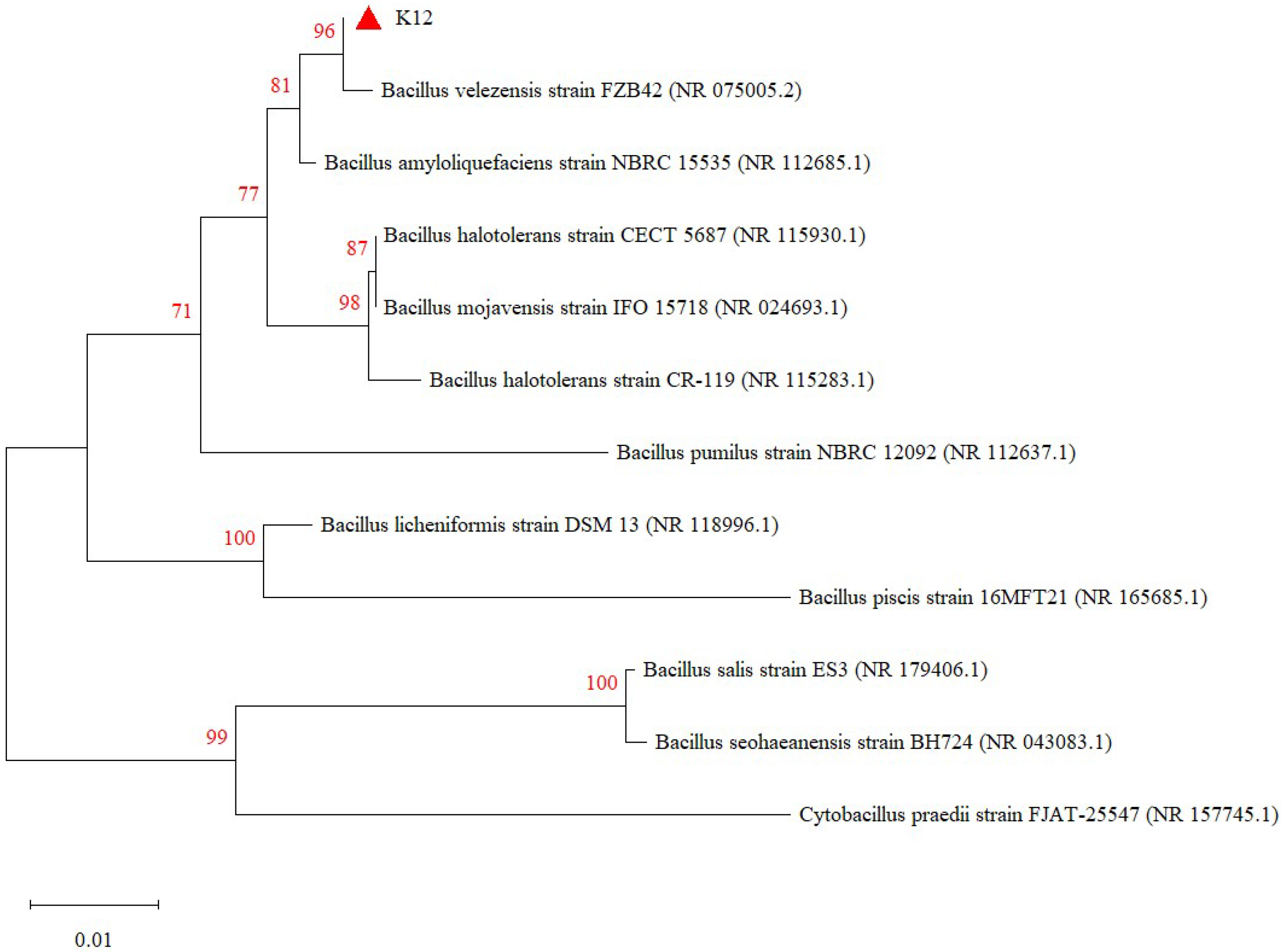
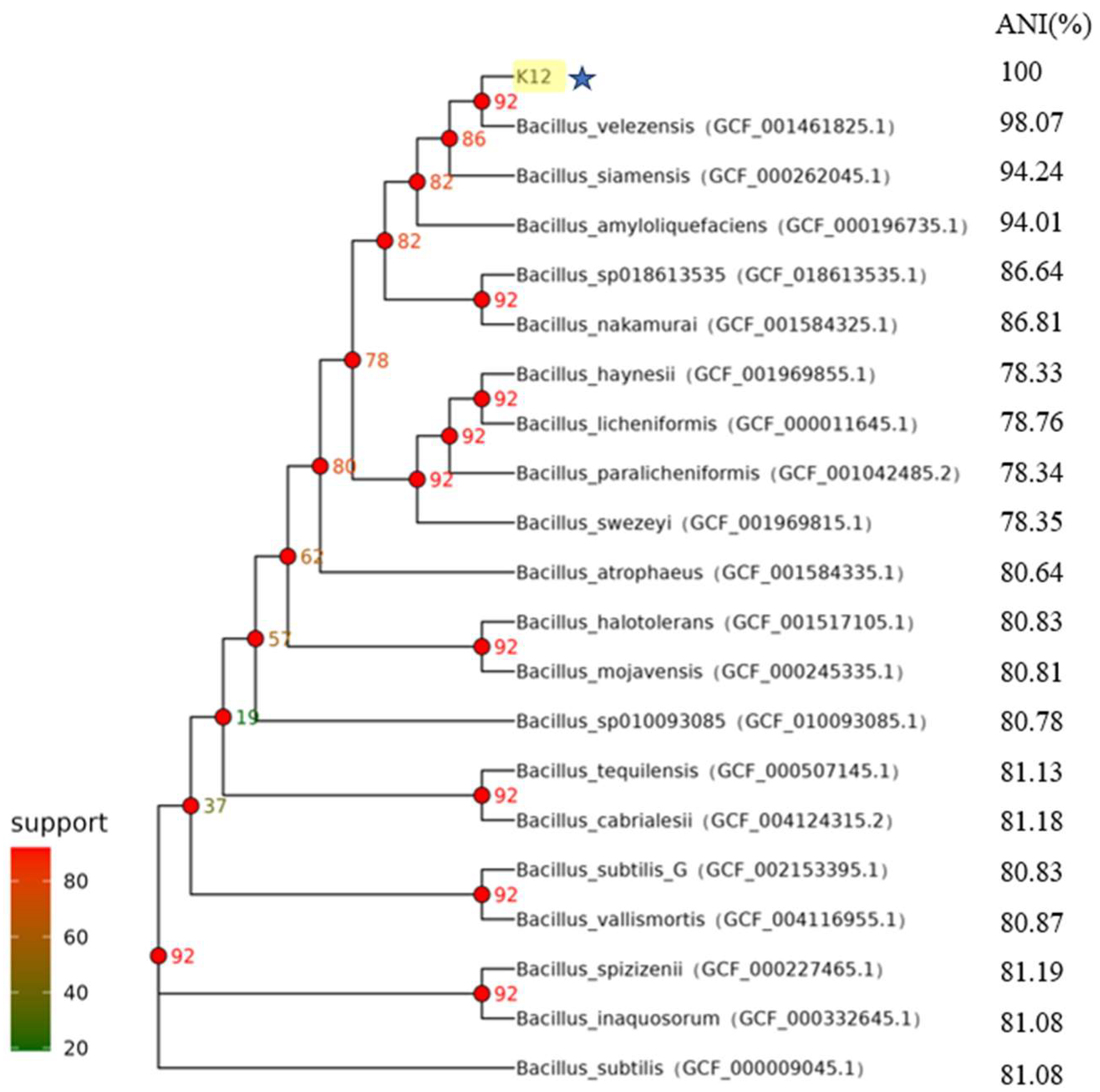
3.1. Identification of Strain K12
3.2. Characterization of the Growth of B. velezensis K12
3.3. In Vitro Bacteriostatic Activity of B. Velezensis K12
3.4. Whole-Genome Sequencing Results of B. velezensis K12
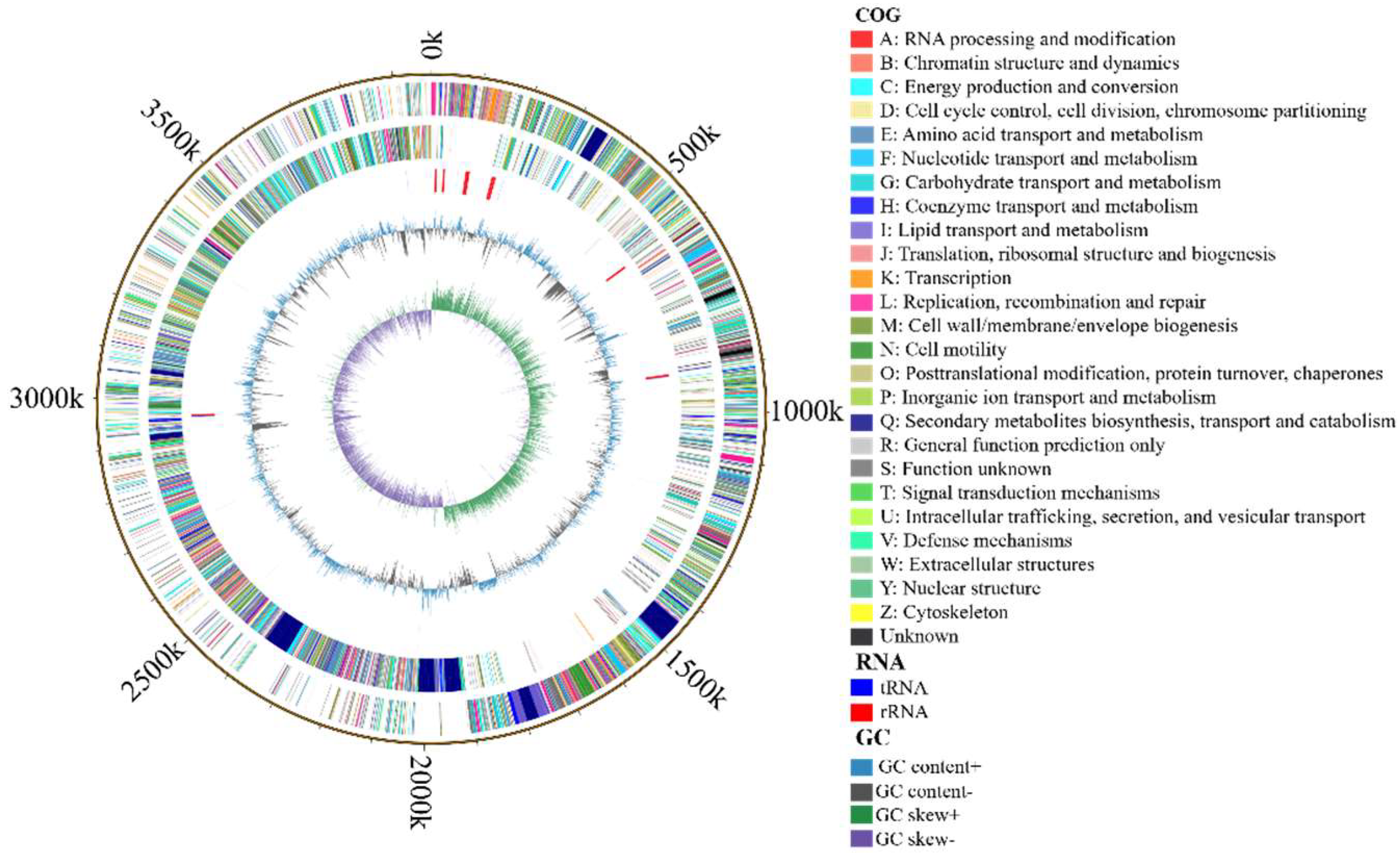
3.4.1. Genome Composition of B. velezensis K12
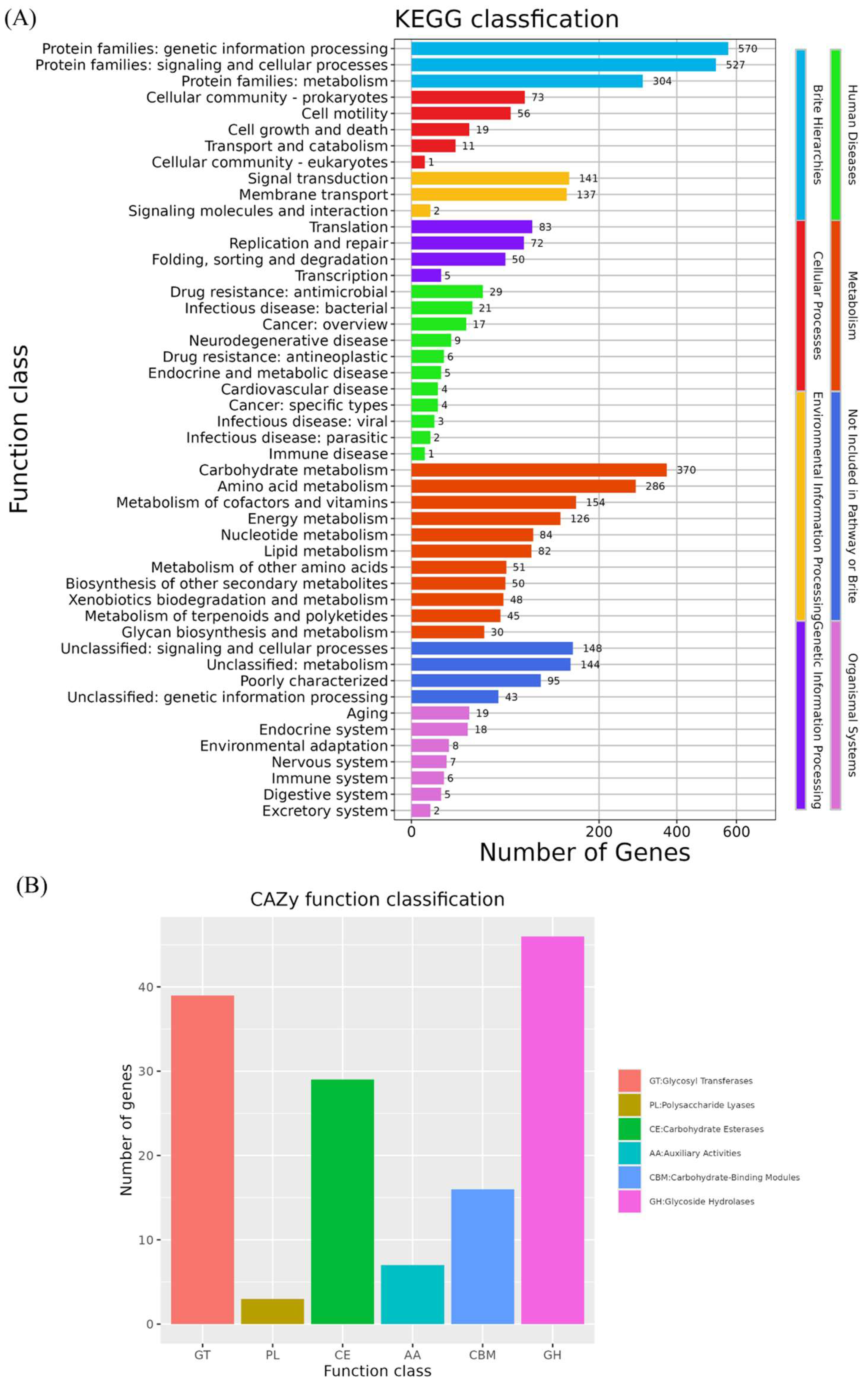
3.4.2. Functional Annotation of B. velezensis K12 Genome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ISAPP | International Scientific Association for Probiotics and Prebiotics |
| E. coli | Escherichia coli |
| LB | Luria–Bertani |
| CLSI | American Society for Clinical Laboratory Standardization |
| ANI | Average Nucleotide Identity |
| UBCG | Up-to-Date Bacterial Core Gene |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| COGs | Clusters of Orthologous Groups |
| GO | Gene Ontology |
| CAZy | Carbohydrate-Active Enzymes database |
| CARD | Comprehensive Antibiotic Research Database |
| VFDB | Virulence Factors of Pathogenic Bacteria Database |
| antiSMASH | Antibiotics and Secondary Metabolite Analysis Shell |
| ANOVA | One-way analysis of variance |
| NCBI | National Center for Biotechnology Information |
| NRPs | Non-ribosomal peptides |
| PKS | Polyketide synthases |
| RiPPs | Ribosomal synthesis and post-translationally modified peptides |
| EFSA | European Food Safety Authority |
| QPS | Qualified Presumption of Safety |
References
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Corrigan, A.; de Leeuw, M.; Penaud-Frézet, S.; Dimova, D.; Murphy, R.A. Phylogenetic and functional alterations in bacterial community compositions in broiler ceca as a result of mannan oligosaccharide supplementation. Appl. Environ. Microbiol. 2015, 81, 3460–3470. [Google Scholar] [CrossRef] [PubMed]
- de Vries, S.; Pustjens, A.M.; Kabel, M.A.; Salazar-Villanea, S.; Hendriks, W.H.; Gerrits, W.J. Processing technologies and cell wall degrading enzymes to improve nutritional value of dried distillers grain with solubles for animal feed: An in vitro digestion study. J. Agric. Food Chem. 2013, 61, 8821–8828. [Google Scholar] [CrossRef]
- Millet, S.; Maertens, L. The European ban on antibiotic growth promoters in animal feed: From challenges to opportunities. Vet. J. 2011, 187, 143–144. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Yousaf, S.; Nouman, H.M.; Ahmed, I.; Husain, S.; Waseem, M.; Nadeem, S.; Tariq, M.; Sizmaz, O.; Chudhry, M.F.Z. A Review of Probiotic Applications in Poultry: Improving Immunity and Having Beneficial Effects on Production and Health. Adv. Microbiol. 2022, 61, 115–123. [Google Scholar] [CrossRef]
- Bilal, M.; Si, W.; Barbe, F.; Chevaux, E.; Sienkiewicz, O.; Zhao, X. Effects of novel probiotic strains of Bacillus pumilus and Bacillus subtilis on production, gut health, and immunity of broiler chickens raised under suboptimal conditions. Poult. Sci. 2021, 100, 100871. [Google Scholar] [CrossRef]
- Tran, C.; Cock, I.E.; Chen, X.; Feng, Y. Antimicrobial Bacillus: Metabolites and Their Mode of Action. Antibiotics 2022, 11, 88. [Google Scholar] [CrossRef]
- Ruiz-García, C.; Béjar, V.; Martínez-Checa, F.; Llamas, I.; Quesada, E. Bacillus velezensis sp. nov., a surfactant-producing bacterium isolated from the river Vélez in Málaga, southern Spain. Int. J. Syst. Evol. Microbiol. 2005, 55, 191–195. [Google Scholar] [CrossRef]
- Ye, M.; Tang, X.; Yang, R.; Zhang, H.; Li, F.; Tao, F.; Li, F.; Wang, Z. Characteristics and Application of a Novel Species of Bacillus: Bacillus velezensis. ACS Chem. Biol. 2018, 13, 500–505. [Google Scholar] [CrossRef]
- Khalid, F.; Khalid, A.; Fu, Y.; Hu, Q.; Zheng, Y.; Khan, S.; Wang, Z. Potential of Bacillus velezensis as a probiotic in animal feed: A review. J. Microbiol. 2021, 59, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Johny, L.C.; Suresh, P.V. Complete genome sequencing and strain characterization of a novel marine Bacillus velezensis FTL7 with a potential broad inhibitory spectrum against foodborne pathogens. World J. Microbiol. Biotechnol. 2022, 38, 164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, Z.; Pu, Y.; Zhang, B.; Wang, B.; Xing, L.; Li, Y.; Zhang, Y.; Gu, R.; Jia, F.; et al. Antagonistic Strain Bacillus velezensis JZ Mediates the Biocontrol of Bacillus altitudinis m-1, a Cause of Leaf Spot Disease in Strawberry. Int. J. Mol. Sci. 2024, 25, 8872. [Google Scholar] [CrossRef] [PubMed]
- Dutilloy, E.; Arias, A.A.; Richet, N.; Guise, J.F.; Duban, M.; Leclere, V.; Selim, S.; Jacques, P.; Jacquard, C.; Clément, C.; et al. Bacillus velezensis BE2 controls wheat and barley diseases by direct antagonism and induced systemic resistance. Appl. Microbiol. Biotechnol. 2024, 108, 64. [Google Scholar] [CrossRef]
- Ye, Q.; Zhong, Z.; Chao, S.; Liu, L.; Chen, M.; Feng, X.; Wu, H. Antifungal Effect of Bacillus velezensis ZN-S10 against Plant Pathogen Colletotrichum changpingense and Its Inhibition Mechanism. Int. J. Mol. Sci. 2023, 24, 16694. [Google Scholar] [CrossRef]
- Yang, H.; Du, D.; Zhang, Q.; Teame, T.; Wang, A.; Hao, Q.; Liu, S.; Ding, Q.; Yao, Y.; Yang, Y.; et al. Dietary Bacillus velezensis T23 fermented products supplementation improves growth, hepatopancreas and intestine health of Litopenaeus vannamei. Fish Shellfish. Immunol. 2024, 149, 109595. [Google Scholar] [CrossRef]
- Chang, X.; Yun, L.; Liu, Z.; Shen, Y.; Feng, S.; Yang, G.; Meng, X. Antagonistic Effects and the Underlying Mechanisms of Bacillus velezensis and its Antibacterial Peptide LCI Against Aeromonas hydrophila Infection in Largemouth Bass. Probiotics Antimicrob. Proteins 2024, 1–18. [Google Scholar] [CrossRef]
- Li, C.; Li, S.; Dang, G.; Jia, R.; Chen, S.; Deng, X.; Liu, G.; Beckers, Y.; Cai, H. Screening and characterization of Bacillus velezensis LB-Y-1 toward selection as a potential probiotic for poultry with multi-enzyme production property. Front. Microbiol. 2023, 14, 1143265. [Google Scholar] [CrossRef]
- Dong, H.; Gao, R.; Dong, Y.; Yao, Q.; Zhu, H. Bacillus velezensis RC116 Inhibits the Pathogens of Bacterial Wilt and Fusarium Wilt in Tomato with Multiple Biocontrol Traits. Int. J. Mol. Sci. 2023, 24, 8527. [Google Scholar] [CrossRef]
- Brutscher, L.M.; Gebrechristos, S.; Garvey, S.M.; Spears, J.L. Genetic and Phenotypic Characterization of Bacillus velezensis Strain BV379 for Human Probiotic Applications. Microorganisms 2024, 12, 436. [Google Scholar] [CrossRef]
- Larsen, I.S.; Chenaux, M.; Collins, F.W.J.; Mandic, A.; Hansen, L.B.S.; Lauridsen, C.A.S.; Haller, R.F.; Elvig-Jørgensen, S.; Horwell, E.; Christiansen, J.; et al. Bacillus velezensis DSM 33864 reduces Clostridioides difficile colonization without disturbing commensal gut microbiota composition. Sci. Rep. 2023, 13, 14941. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Kong, Y.; Fan, Y.; Geng, C.; Peng, D.; Sun, M. Whole-genome sequencing of Bacillus velezensis LS69, a strain with a broad inhibitory spectrum against pathogenic bacteria. J. Biotechnol. 2017, 249, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Chen, H.; Zhu, L.; Song, Y.; Jiang, Q.; Zhang, Y.; Ali, Q.; Gu, Q.; Gao, X.; Borriss, R.; et al. Profiling of Antimicrobial Metabolites Synthesized by the Endophytic and Genetically Amenable Biocontrol Strain Bacillus velezensis DMW1. Microbiol. Spectr. 2023, 11, e0003823. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; He, X.; Chen, J.; Li, J.; Wu, Y.; Chen, Y.; Yang, Y. Whole-Genome Analysis of Termite-Derived Bacillus velezensis BV-10 and Its Application in King Grass Silage. Microorganisms 2023, 11, 2697. [Google Scholar] [CrossRef]
- Ablimit, N.; Zheng, F.; Wang, Y.; Wen, J.; Wang, H.; Deng, K.; Cao, Y.; Wang, Z.; Jiang, W. Bacillus velezensis strain NA16 shows high poultry feather-degrading efficiency, protease and amino acid production. Ecotoxicol. Environ. Saf. 2024, 278, 116353. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2012, 41, e1. [Google Scholar] [CrossRef]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2022, 50, D20–D26. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Renschler, M.A.; Wyatt, A.; Anene, N.; Robinson-Hill, R.; Pickerill, E.S.; Fox, N.E.; Griffith, J.A.; McKillip, J.L. Using nitrous acid-modified de Man, Rogosa, and Sharpe medium to selectively isolate and culture lactic acid bacteria from dairy foods. J. Dairy Sci. 2020, 103, 1215–1222. [Google Scholar] [CrossRef]
- Soni, R.; Keharia, H.; Shah, K.; Jain, N. Phenotypic characterization and genome analysis reveal the probiotic potential of a banyan endophyte Bacillus velezensis K1. J. Appl. Microbiol. 2023, 134, lxac057. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Cortes, J.L.; Azaola-Espinosa, A.; Lozano-Aguirre, L.; Ponce-Alquicira, E. Physiological and Genomic Analysis of Bacillus pumilus UAMX Isolated from the Gastrointestinal Tract of Overweight Individuals. Microorganisms 2021, 9, 1076. [Google Scholar] [CrossRef]
- Tanvi, S.; Ravichandra, V.; Madhur, D.S.; Agampodi Promoda, P.; Stephen, T.; Roger, S.; Rajaraman, E. Probiotic Bacillus coagulans MTCC 5856 spores exhibit excellent in-vitro functional efficacy in simulated gastric survival, mucosal adhesion and immunomodulation. J. Funct. Foods 2019, 52, 100–108. [Google Scholar] [CrossRef]
- Humphries, R.; Bobenchik, A.M.; Hindler, J.A.; Schuetz, A.N. Overview of Changes to the Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial Susceptibility Testing, M100, 31st Edition. J. Clin. Microbiol. 2021, 59, e0021321. [Google Scholar] [CrossRef] [PubMed]
- Yi-Zhou, X.; Xin-Yue, L.; Hai-Long, Z.; Jun-Yuan, C.; Lian-Bing, L.; Qi-Lin, Z. Purification and antibacterial properties of a novel bacteriocin against Escherichia coli from Bacillus subtilis isolated from blueberry ferments. LWT 2021, 146, 111456. [Google Scholar] [CrossRef]
- Mark, D.; Tairo, F.; Ndunguru, J.; Kweka, E.; Saggaf, M.; Bachwenkizi, H.; Chiunga, E.; Lusana, J.L.; Sikazwe, G.; Maghembe, R. Assessing the effect of sample storage time on viral detection using a rapid and cost-effective CTAB-based extraction method. Plant Methods 2024, 20, 64. [Google Scholar] [CrossRef]
- Safar, H.A.; Alatar, F.; Nasser, K.; Al-Ajmi, R.; Alfouzan, W.; Mustafa, A.S. The impact of applying various de novo assembly and correction tools on the identification of genome characterization, drug resistance, and virulence factors of clinical isolates using ONT sequencing. BMC Biotechnol. 2023, 23, 26. [Google Scholar] [CrossRef]
- Chen, S. Ultrafast one-pass FASTQ data preprocessing, quality control, and deduplication using fastp. iMeta 2023, 2, e107. [Google Scholar] [CrossRef]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y.; et al. SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. GigaScience 2012, 1, 18. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef] [PubMed]
- Jain, C.; Rodriguez, R.L.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef]
- Na, S.I.; Kim, Y.O.; Yoon, S.H.; Ha, S.M.; Baek, I.; Chun, J. UBCG: Up-to-date bacterial core gene set and pipeline for phylogenomic tree reconstruction. J. Microbiol. 2018, 56, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. GeneMarkS: A self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef]
- Chan, P.P.; Lin, B.Y.; Mak, A.J.; Lowe, T.M. tRNAscan-SE 2.0: Improved detection and functional classification of transfer RNA genes. Nucleic Acids Res. 2021, 49, 9077–9096. [Google Scholar] [CrossRef]
- Nawrocki, E.P.; Burge, S.W.; Bateman, A.; Daub, J.; Eberhardt, R.Y.; Eddy, S.R.; Floden, E.W.; Gardner, P.P.; Jones, T.A.; Tate, J.; et al. Rfam 12.0: Updates to the RNA families database. Nucleic Acids Res. 2015, 43, D130–D137. [Google Scholar] [CrossRef] [PubMed]
- Tarailo-Graovac, M.; Chen, N. Using RepeatMasker to identify repetitive elements in genomic sequences. Curr. Protoc. Bioinform. 2009, 5, 4.10.11–14.10.14. [Google Scholar] [CrossRef]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef]
- Couvin, D.; Bernheim, A.; Toffano-Nioche, C.; Touchon, M.; Michalik, J.; Néron, B.; Rocha, E.P.C.; Vergnaud, G.; Gautheret, D.; Pourcel, C. CRISPRCasFinder, an update of CRISRFinder, includes a portable version, enhanced performance and integrates search for Cas proteins. Nucleic Acids Res. 2018, 46, W246–W251. [Google Scholar] [CrossRef]
- Akhter, S.; Aziz, R.K.; Edwards, R.A. PhiSpy: A novel algorithm for finding prophages in bacterial genomes that combines similarity- and composition-based strategies. Nucleic Acids Res. 2012, 40, e126. [Google Scholar] [CrossRef]
- Bertelli, C.; Laird, M.R.; Williams, K.P.; Lau, B.Y.; Hoad, G.; Winsor, G.L.; Brinkman, F.S.L. IslandViewer 4: Expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 2017, 45, W30–W35. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res. 2023, 51, D587–D592. [Google Scholar] [CrossRef] [PubMed]
- Cantalapiedra, C.P.; Hernández-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. eggNOG-mapper v2: Functional Annotation, Orthology Assignments, and Domain Prediction at the Metagenomic Scale. Mol. Biol. Evol. 2021, 38, 5825–5829. [Google Scholar] [CrossRef]
- Consortium, T.G.O. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar] [CrossRef]
- Ausland, C.; Zheng, J.; Yi, H.; Yang, B.; Li, T.; Feng, X.; Zheng, B.; Yin, Y. dbCAN-PUL: A database of experimentally characterized CAZyme gene clusters and their substrates. Nucleic Acids Res. 2021, 49, D523–D528. [Google Scholar] [CrossRef] [PubMed]
- Alcock, B.P.; Huynh, W.; Chalil, R.; Smith, K.W.; Raphenya, A.R.; Wlodarski, M.A.; Edalatmand, A.; Petkau, A.; Syed, S.A.; Tsang, K.K.; et al. CARD 2023: Expanded curation, support for machine learning, and resistome prediction at the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2023, 51, D690–D699. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. VFDB 2022: A general classification scheme for bacterial virulence factors. Nucleic Acids Res. 2022, 50, D912–D917. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Augustijn, H.E.; Reitz, Z.L.; Biermann, F.; Alanjary, M.; Fetter, A.; Terlouw, B.R.; Metcalf, W.W.; Helfrich, E.J.N.; et al. antiSMASH 7.0: New and improved predictions for detection, regulation, chemical structures and visualisation. Nucleic Acids Res. 2023, 51, W46–W50. [Google Scholar] [CrossRef]
- Dhanya Raj, C.T.; Suryavanshi, M.V.; Kandaswamy, S.; Ramasamy, K.P.; James, R.A. Whole genome sequence analysis and in-vitro probiotic characterization of Bacillus velezensis FCW2 MCC4686 from spontaneously fermented coconut water. Genomics 2023, 115, 110637. [Google Scholar] [CrossRef]
- McHugh, M.L. Multiple comparison analysis testing in ANOVA. Biochem. Medica 2011, 21, 203–209. [Google Scholar] [CrossRef]
- Stothard, P.; Wishart, D.S. Circular genome visualization and exploration using CGView. Bioinformatics 2005, 21, 537–539. [Google Scholar] [CrossRef]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Hilbert, F.; Lindqvist, R.; et al. Update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 11: Suitability of taxonomic units notified to EFSA until September 2019. EFSA J. Eur. Food Saf. Auth. 2020, 18, e05965. [Google Scholar] [CrossRef]
- Quach, N.T.; Vu, T.H.N.; Nguyen, N.A.; Nguyen, V.T.; Bui, T.L.; Ky, S.C.; Le, T.L.; Hoang, H.; Ngo, C.C.; Le, T.T.M.; et al. Phenotypic features and analysis of genes supporting probiotic action unravel underlying perspectives of Bacillus velezensis VTX9 as a potential feed additive for swine. Ann. Microbiol. 2021, 71, 36. [Google Scholar] [CrossRef]
- Twinkle, B.; Bhargab, G.; Ankita, K.; Madhurjya, G.; Aparoop, D.; Debajit, B. Probiotic characterization of indigenous Bacillus velezensis strain DU14 isolated from Apong, a traditionally fermented rice beer of Assam. Biocatal. Agric. Biotechnol. 2019, 18, 101008. [Google Scholar] [CrossRef]
- Żbikowska, K.; Michalczuk, M.; Dolka, B. The Use of Bacteriophages in the Poultry Industry. Animals 2020, 10, 872. [Google Scholar] [CrossRef]
- Liu, Z.; Zhou, Y.; Wang, H.; Liu, C.; Wang, L. Recent advances in understanding the fitness and survival mechanisms of Vibrio parahaemolyticus. Int. J. Food Microbiol. 2024, 417, 110691. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wang, Y.; Liu, Y.; Jia, K.; Zhang, Z.; Dong, Q. Cereulide and Emetic Bacillus cereus: Characterizations, Impacts and Public Precautions. Foods 2023, 12, 833. [Google Scholar] [CrossRef]
- Wu, L.; Wu, H.; Chen, L.; Yu, X.; Borriss, R.; Gao, X. Difficidin and bacilysin from Bacillus amyloliquefaciens FZB42 have antibacterial activity against Xanthomonas oryzae rice pathogens. Sci. Rep. 2015, 5, 12975. [Google Scholar] [CrossRef]
- Li, Y.; Gu, Y.; Li, J.; Xu, M.; Wei, Q.; Wang, Y. Biocontrol agent Bacillus amyloliquefaciens LJ02 induces systemic resistance against cucurbits powdery mildew. Front. Microbiol. 2015, 6, 883. [Google Scholar] [CrossRef]
- Sabaté, D.C.; Audisio, M.C. Inhibitory activity of surfactin, produced by different Bacillus subtilis subsp. subtilis strains, against Listeria monocytogenes sensitive and bacteriocin-resistant strains. Microbiol. Res. 2013, 168, 125–129. [Google Scholar] [CrossRef]
- Romero-Tabarez, M.; Jansen, R.; Sylla, M.; Lünsdorf, H.; Häussler, S.; Santosa, D.A.; Timmis, K.N.; Molinari, G. 7-O-malonyl macrolactin A, a new macrolactin antibiotic from Bacillus subtilis active against methicillin-resistant Staphylococcus aureus, vancomycin-resistant enterococci, and a small-colony variant of Burkholderia cepacia. Antimicrob. Agents Chemother. 2006, 50, 1701–1709. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Overbeck, T.J.; Skebba, V.L.P.; Gandhi, N.N. Genomic and Phenotypic Safety Assessment of Probiotic Bacillus coagulans Strain JBI-YZ6.3. Probiotics Antimicrob. Proteins 2024, 1–11. [Google Scholar] [CrossRef]
- Fujisawa, M.; Kusumoto, A.; Wada, Y.; Tsuchiya, T.; Ito, M. NhaK, a novel monovalent cation/H+ antiporter of Bacillus subtilis. Arch. Microbiol. 2005, 183, 411–420. [Google Scholar] [CrossRef]
- Xiong, Z.Q.; Wang, Q.H.; Kong, L.H.; Song, X.; Wang, G.Q.; Xia, Y.J.; Zhang, H.; Sun, Y.; Ai, L.Z. Short communication: Improving the activity of bile salt hydrolases in Lactobacillus casei based on in silico molecular docking and heterologous expression. J. Dairy Sci. 2017, 100, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Mazhar, S.; Khokhlova, E.; Colom, J.; Simon, A.; Deaton, J.; Rea, K. In vitro and in silico assessment of probiotic and functional properties of Bacillus subtilis DE111®. Front. Microbiol. 2022, 13, 1101144. [Google Scholar] [CrossRef]
- Prasad, J.; McJarrow, P.; Gopal, P. Heat and osmotic stress responses of probiotic Lactobacillus rhamnosus HN001 (DR20) in relation to viability after drying. Appl. Environ. Microbiol. 2003, 69, 917–925. [Google Scholar] [CrossRef]
- Lipke, P.N.; Ragonis-Bachar, P. Sticking to the Subject: Multifunctionality in Microbial Adhesins. J. Fungi 2023, 9, 419. [Google Scholar] [CrossRef]
- Sam-On, M.F.S.; Mustafa, S.; Mohd Hashim, A.; Yusof, M.T.; Zulkifly, S.; Malek, A.Z.A.; Roslan, M.A.H.; Mohd Asrore, M.S. Mining the genome of Bacillus velezensis FS26 for probiotic markers and secondary metabolites with antimicrobial properties against aquaculture pathogens. Microb. Pathog. 2023, 181, 106161. [Google Scholar] [CrossRef] [PubMed]
- da Costa, T.M.; Viljoen, A.; Towell, A.M.; Dufrêne, Y.F.; Geoghegan, J.A. Fibronectin binding protein B binds to loricrin and promotes corneocyte adhesion by Staphylococcus aureus. Nat. Commun. 2022, 13, 2517. [Google Scholar] [CrossRef]
- Butorac, K.; Novak, J.; Bellich, B.; Terán, L.C.; Banić, M.; Leboš Pavunc, A.; Zjalić, S.; Cescutti, P.; Šušković, J.; Kos, B. Lyophilized alginate-based microspheres containing Lactobacillus fermentum D12, an exopolysaccharides producer, contribute to the strain’s functionality in vitro. Microb. Cell Factories 2021, 20, 85. [Google Scholar] [CrossRef]
- Lunderberg, J.M.; Liszewski Zilla, M.; Missiakas, D.; Schneewind, O. Bacillus anthracis tagO Is Required for Vegetative Growth and Secondary Cell Wall Polysaccharide Synthesis. J. Bacteriol. 2015, 197, 3511–3520. [Google Scholar] [CrossRef] [PubMed]
- Yun, L.; Kang, M.; Shen, Y.; Feng, J.; Yang, G.; Zhang, J.; Meng, X.; Chang, X. Dietary Bacillus velezensis R-71003 and sodium gluconate improve antioxidant capacity, immune response and resistance against Aeromonas hydrophila in common carp. Fish Shellfish Immunol. 2023, 139, 108921. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Su, W.; Li, W.; Wen, C.; Du, S.; He, H.; Zhang, Y.; Gong, T.; Wang, X.; Wang, Y.; et al. Bacillus amyloliquefaciens 40 regulates piglet performance, antioxidant capacity, immune status and gut microbiota. Anim. Nutr. (Zhongguo Xu Mu Shou Yi Xue Hui) 2023, 12, 116–127. [Google Scholar] [CrossRef]
- Wei-Chuan, C.; Ruey-Shin, J.; Yu-Hong, W. Applications of a lipopeptide biosurfactant, surfactin, produced by microorganisms. Biochem. Eng. J. 2015, 103, 158–169. [Google Scholar] [CrossRef]
- Holzapfel, W.; Arini, A.; Aeschbacher, M.; Coppolecchia, R.; Pot, B. Enterococcus faecium SF68 as a model for efficacy and safety evaluation of pharmaceutical probiotics. Benef. Microbes 2018, 9, 375–388. [Google Scholar] [CrossRef]
- Chen, B.; Zhou, Y.; Duan, L.; Gong, X.; Liu, X.; Pan, K.; Zeng, D.; Ni, X.; Zeng, Y. Complete genome analysis of Bacillus velezensis TS5 and its potential as a probiotic strain in mice. Front. Microbiol. 2023, 14, 1322910. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, R.; Jessberger, N.; Ehling-Schulz, M.; Märtlbauer, E.; Granum, P.E. The Food Poisoning Toxins of Bacillus cereus. Toxins 2021, 13, 98. [Google Scholar] [CrossRef]
- Fu, R.; Chen, D.; Tian, G.; Zheng, P.; Mao, X.; Yu, J.; He, J.; Huang, Z.; Luo, Y.; Yu, B. Effect of dietary supplementation of Bacillus coagulans or yeast hydrolysates on growth performance, antioxidant activity, cytokines and intestinal microflora of growing-finishing pigs. Anim. Nutr. (Zhongguo Xu Mu Shou Yi Xue Hui) 2019, 5, 366–372. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Simulated Gastrointestinal Fluids | Time/h | Survival Rate/% | ||
|---|---|---|---|---|
| 0 | 3 | |||
| Bacterial concentration (CFU/mL) | Simulated gastric fluid | 1.1 × 108 | 1.1 × 104 | 0.01% |
| Simulated intestinal fluid | 8.6 × 107 | 3.5 × 107 | 40.70% | |
| Antibiotics | Inhibitory Circle Diameter/mm |
|---|---|
| Florfenicol | 29.08 ± 1.17 |
| Ciprofloxacin | 27.69 ± 0.11 |
| Cotrimoxazole | 22.94 ± 0.04 |
| Gentamycin | 23.54 ± 0.11 |
| Doxycycline | 21.59 ± 1.04 |
| Cefotaxime | 22.33 ± 5.43 |
| Dilution Factor | E. coli CVCC25922 | E. coli K88 | Staphylococcus aureus CVCC1822 | Salmonella CVCC519 | Bacillus cereus CICC21290 | Clostridium perfringens CVCC66 | Vibrio parahaemolyticus CICC23924 |
|---|---|---|---|---|---|---|---|
| Control | 8.00 ± 0.00 e | 8.00 ± 0.00 d | 8.00 ± 0.00 g | 8.00 ± 0.00 f | 8.00 ± 0.00 e | 8.00 ± 0.00 d | 8.00 ± 0.00 g |
| 10 | 22.06 ± 2.24 a | 18.11 ± 2.84 a | 20.89 ± 0.19 a | 19.50 ± 0.44 a | 17.00 ± 0.00 a | 13.11 ± 0.19 a | 21.02 ± 0.03 a |
| 100 | 18.89 ± 1.92 b | 16.66 ± 2.32 ab | 18.36 ± 0.70 b | 17.06 ± 0.42 b | 14.11 ± 0.19 b | 11.56 ± 0.51 b | 19.50 ± 0.50 b |
| 1000 | 16.33 ± 2.08 c | 15.23 ± 2.17 abc | 14.33 ± 0.44 c | 15.45 ± 0.39 c | 12.67 ± 0.34 c | 10.89 ± 0.35 c | 17.83 ± 0.58 c |
| 4000 | 14.17 ± 0.73 cd | 14.19 ± 2.01 abc | 13.07 ± 0.09 d | 13.50 ± 0.44 d | 10.94 ± 0.10 d | 8.00 ± 0.00 d | 16.83 ± 0.29 d |
| 8000 | 13.28 ± 1.11 d | 12.72 ± 2.33 bc | 12.06 ± 0.05 e | 13.06 ± 0.10 d | 8.00 ± 0.00 e | 8.00 ± 0.00 d | 15.67 ± 0.58 e |
| 10,000 | 12.44 ± 0.96 d | 11.33 ± 3.51 cd | 11.00 ± 0.00 f | 11.78 ± 0.39 e | 8.00 ± 0.00 e | 8.00 ± 0.00 d | 14.50 ± 0.87 f |
| Line p-value | <0.001 | <0.001 | 0.057 | <0.001 | 0.256 | <0.001 | <0.001 |
| Sample | E. coli CVCC25922 | E. coli K88 | Staphylococcus aureus CVCC1822 | Salmonella CVCC519 | Bacillus cereus CICC21290 | Clostridium perfringens CVCC66 | Vibrio parahaemolyticus CICC23924 |
|---|---|---|---|---|---|---|---|
| Sterile water | 8.00 ± 0.00 b | 8.00 ± 0.00 c | 8.00 ± 0.00 b | 8.00 ± 0.00 c | 8.00 ± 0.00 d | 8.00 ± 0.00 c | 8.00 ± 0.00 d |
| Whole bacteria | 16.83 ± 1.61 a | 16.20 ± 0.26 a | 16.83 ± 1.61 a | 18.67 ± 1.15 a | 18.78 ± 0.84 a | 13.00 ± 1.00 a | 17.83 ± 0.29 a |
| Supernatant | 8.00 ± 0.00 b | 15.03 ± 0.50 b | 8.00 ± 0.00 b | 8.00 ± 0.00 c | 12.11 ± 0.19 c | 11.00 ± 1.00 b | 14.87 ± 0.32 c |
| Sediment | 15.50 ± 1.50 a | 15.07 ± 1.01 b | 15.50 ± 1.50 a | 17.17 ± 1.04 b | 15.56 ± 0.51 b | 11.33 ± 1.15 b | 17.37 ± 0.23 b |
| LB | 8.00 ± 0.00 b | 8.00 ± 0.00 c | 8.00 ± 0.00 b | 8.00 ± 0.00 c | 8.00 ± 0.00 d | 8.00 ± 0.00 c | 8.00 ± 0.00 d |
| PBS | 8.00 ± 0.00 b | 8.00 ± 0.00 c | 8.00 ± 0.00 b | 8.00 ± 0.00 c | 8.00 ± 0.00 d | 8.00 ± 0.00 c | 8.00 ± 0.00 d |
| Samples | E. coli K88 |
|---|---|
| Whole bacteria | 16.56 ± 0.51 a |
| Boiled | 8.00 ± 0.00 b |
| High-pressure treatment | 8.00 ± 0.00 b |
| LB | 8.00 ± 0.00 b |
| Genome | Value |
|---|---|
| Seq length (bp) | 3,973,405 |
| Seq type | Circular |
| G + C content (%) | 46.69 |
| N20 length (bp) | 28,206 |
| N50 length (bp) | 9802 |
| N90 length (bp) | 1825 |
| Max length (bp) | 174,061 |
| Min length (bp) | 1 |
| Total number of genes | 4123 |
| Number of coding sequences (CDS) | 3913 |
| Average length (bp) | 900.47 |
| Gene/genome (%) | 88.68 |
| Prophages | 19 |
| Genomic islands | 21 |
| Number of RNA genes | 210 |
| Gene Number | Resistant Gene | Antibiotic Resistance | Resistance Mechanism |
|---|---|---|---|
| chr_263 | lmrB | lincosamide antibiotic; nucleoside antibiotic | Antibiotic efflux |
| chr_519 | clbA | lincosamide antibiotic; oxazolidinone antibiotic; phenicol antibiotic; pleuromutilin antibiotic; streptogramin A antibiotic; streptogramin antibiotic | Antibiotic target alteration |
| chr_1328 | ykkD | aminoglycoside antibiotic; phenicol antibiotic; tetracycline antibiotic | Antibiotic efflux |
| chr_124 | EF-Tu | elfamycin antibiotic | Antibiotic target alteration |
| chr_118 | rpoB | peptide antibiotic; rifamycin antibiotic | Antibiotic target alteration; antibiotic target replacement |
| chr_142 | rpsE | aminoglycoside antibiotic | Antibiotic target alteration |
| Gene Number | VF Id (gb Number) | Virulence Gene | VF Function | Identity/% |
|---|---|---|---|---|
| chr 1907 | VFG016303 (gb|NP 389723) | dep/capD | Immune modulation | 84.615 |
| chr 2046 | VFG016235 (gb|WP 003182805) | hlyIII | Exotoxin | 80.189 |
| chr 2967 | VFG050021 (gb|NP 391078) | dhbE | Nutritional/Metabolic factor | 81.051 |
| chr 3389 | VFG016299 (gb|NP 391471) | capB | Immune modulation | 93.13 |
| Type | Start | End | Most Similar Known Cluster | Similarity |
|---|---|---|---|---|
| lanthipeptide-class-iii | 193,784 | 216,399 | andalusicin A; andalusicin B RiPP:Lanthipeptide | 100% |
| NRPS | 314,996 | 380,403 | surfactin NRP:Lipopeptide | 95% |
| RRE-containing, LAP | 708,772 | 731,949 | plantazolicin RiPP:LAP | 91% |
| terpene | 1,097,917 | 1,118,657 | - | - |
| transAT-PKS | 1,421,088 | 1,509,330 | macrolactin H Polyketide | 100% |
| transAT-PKS, T3PKS, NRPS | 1,731,593 | 1,841,698 | bacillaene Polyketide + NRP | 100% |
| NRPS, transAT-PKS, betalactone | 1,898,733 | 2,036,597 | fengycin NRP | 100% |
| terpene | 2,061,882 | 2,083,765 | - | - |
| T3PKS | 2,143,224 | 2,184,324 | - | - |
| transAT-PKS | 2,300,233 | 2,406,424 | difficidin Polyketide | 100% |
| NRPS | 2,892,056 | 2,942,291 | bacillothiazol | 100% |
| NRP-metallophore, NRPS, RiPP-like | 3,039,145 | 3,090,934 | bacillibactin NRP | 100% |
| Other | 3,624,730 | 3,666,148 | bacilysin other | 100% |
| Functional Category | Gene | Description/Function | Gene Number |
|---|---|---|---|
| Acid stress | atp | ATP synthase | chr 3493, chr 3495, chr 3487, chr 3488, chr 3489, chr 3490, chr 3492, chr 3494 |
| atpH | F(1)F(0)-ATP synthase | chr 3491 | |
| nhaC | Na+/H+ antiporter | chr 2382 | |
| nhaC 1 | Na+/H+ antiporter | chr 530 | |
| mleN | Na+/H+ antiporter | chr 2225 | |
| yuiF | Na+/H+ antiporter | chr 2973 | |
| ycgA | Na+/H+ antiporter | chr 291 | |
| - | Na+/H+ antiporter | chr 904 | |
| mrp | Na+ (K+, Li+ and/or alkali)/H+ antiporter | chr 164, chr 2931, chr 2932, chr 2933, chr 2934, chr 2935, chr 2936, chr 2937 | |
| Bile tolerance | bshC | Glucosaminyl-malate cysteine ligase | chr 1545 |
| bshA | N-acetyl-alpha-D-glucosaminyl L-malate synthase | chr 2114 | |
| bshB1 | Acetylglucosamine-malate deacetylase | chr 2115 | |
| pyrG | CTP synthase | chr 3523 | |
| dnaK | Chaperone protein | chr 2440 | |
| dnaJ | Chaperone protein | chr 2439 | |
| eno | Enolase C/Enolase N | chr 3202 | |
| clp | CLP protease | chr 97, chr 1386, chr 2596, chr 3264 | |
| oppA | ABC transporter substrate-binding protein | chr 1159 | |
| nagB | Glucosamine-6-phosphate deaminase | chr 3301 | |
| - | Sodium bile acid symporter | chr 526 | |
| Adhesion | fbp | Firmicute fructose-1,6-bisphosphatase | chr 3790, chr 3797 |
| eps | Glycosyltransferases | chr 3232, chr 3233, chr 3234, chr 3235, chr 3236, chr 3237, chr 3238, chr 3239, chr 3240, chr 3241, chr 3242, chr 3243 | |
| lip | Lipase | chr 266 | |
| galE | NAD(P)-dependent epimerase dehydratase | chr 1228 | |
| lspA | Peptidase | chr 1575 | |
| glnH | Bacterial solute-binding protein | chr 2519 | |
| tuf | GTP EFTU | chr 124 | |
| tasA 1 | Cell division protein | chr 2356 | |
| tagO | UDP-N-acetylglucosamine-1-phosphate transferase | chr 3351 | |
| Oxidative | kat | Catalase | chr 914, chr 3685, chr 3712 |
| ahp | Alkyl hydroperoxide reductase | chr 3786, chr 3787 | |
| bsaA | glutathione peroxidase | chr 2056 | |
| sodF | Superoxide dismutase | chr 1962 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Li, T.; Huang, Y.; Wu, L.; Liu, X.; Yue, R.; Yuan, J. Comprehensive Phenotypic Characterization and Genomic Analysis Unveil the Probiotic Potential of Bacillus velezensis K12. Animals 2025, 15, 798. https://doi.org/10.3390/ani15060798
Tang Y, Li T, Huang Y, Wu L, Liu X, Yue R, Yuan J. Comprehensive Phenotypic Characterization and Genomic Analysis Unveil the Probiotic Potential of Bacillus velezensis K12. Animals. 2025; 15(6):798. https://doi.org/10.3390/ani15060798
Chicago/Turabian StyleTang, Yingying, Tian Li, Yihong Huang, Liangliang Wu, Xiaobo Liu, Ruichao Yue, and Jianmin Yuan. 2025. "Comprehensive Phenotypic Characterization and Genomic Analysis Unveil the Probiotic Potential of Bacillus velezensis K12" Animals 15, no. 6: 798. https://doi.org/10.3390/ani15060798
APA StyleTang, Y., Li, T., Huang, Y., Wu, L., Liu, X., Yue, R., & Yuan, J. (2025). Comprehensive Phenotypic Characterization and Genomic Analysis Unveil the Probiotic Potential of Bacillus velezensis K12. Animals, 15(6), 798. https://doi.org/10.3390/ani15060798