
Probiotic Characterization of Lactic Acid Bacteria from Donkey Feces in China

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Preliminary Identification
2.2. Antimicrobial Activity
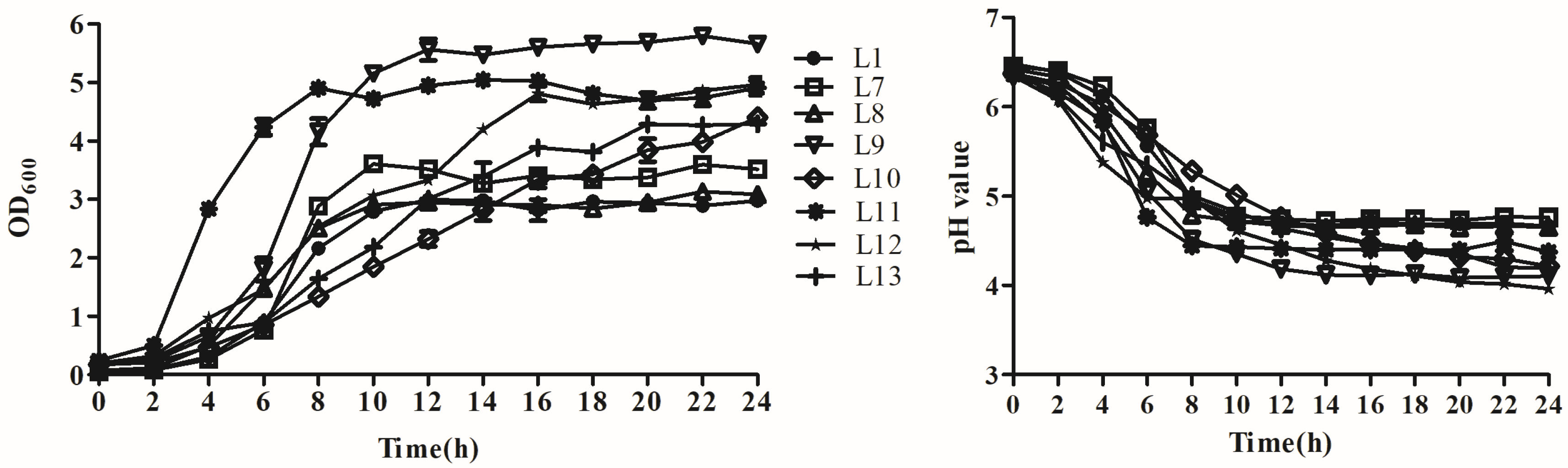
2.3. Growth Curve and Acid Production Rate
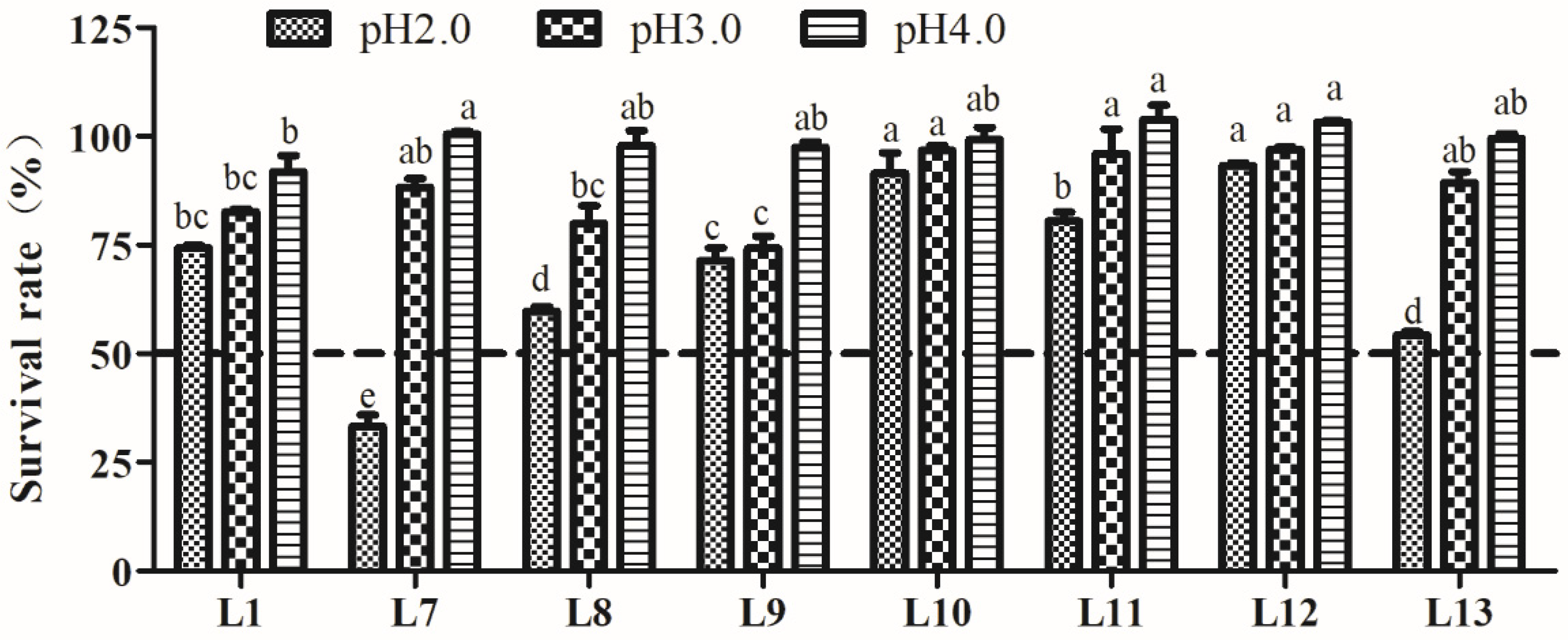
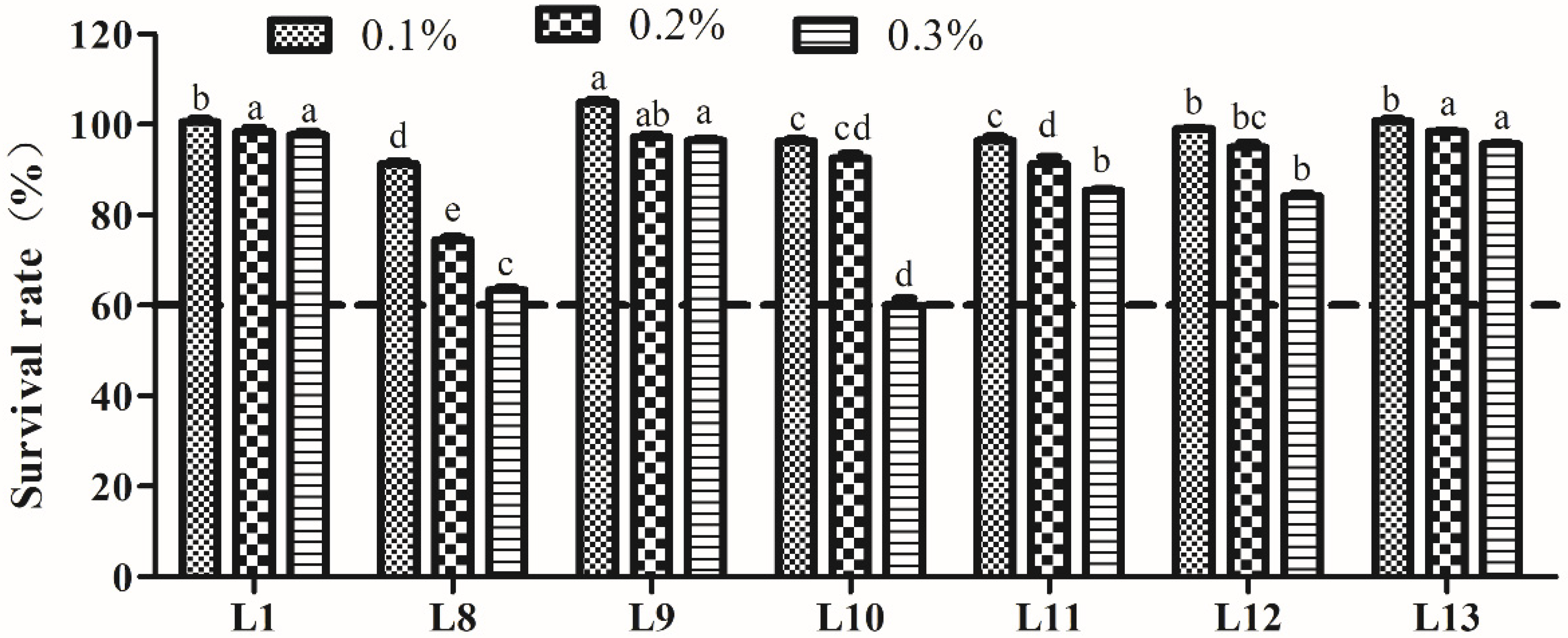
2.4. Tolerance to Acid and Bile Salts
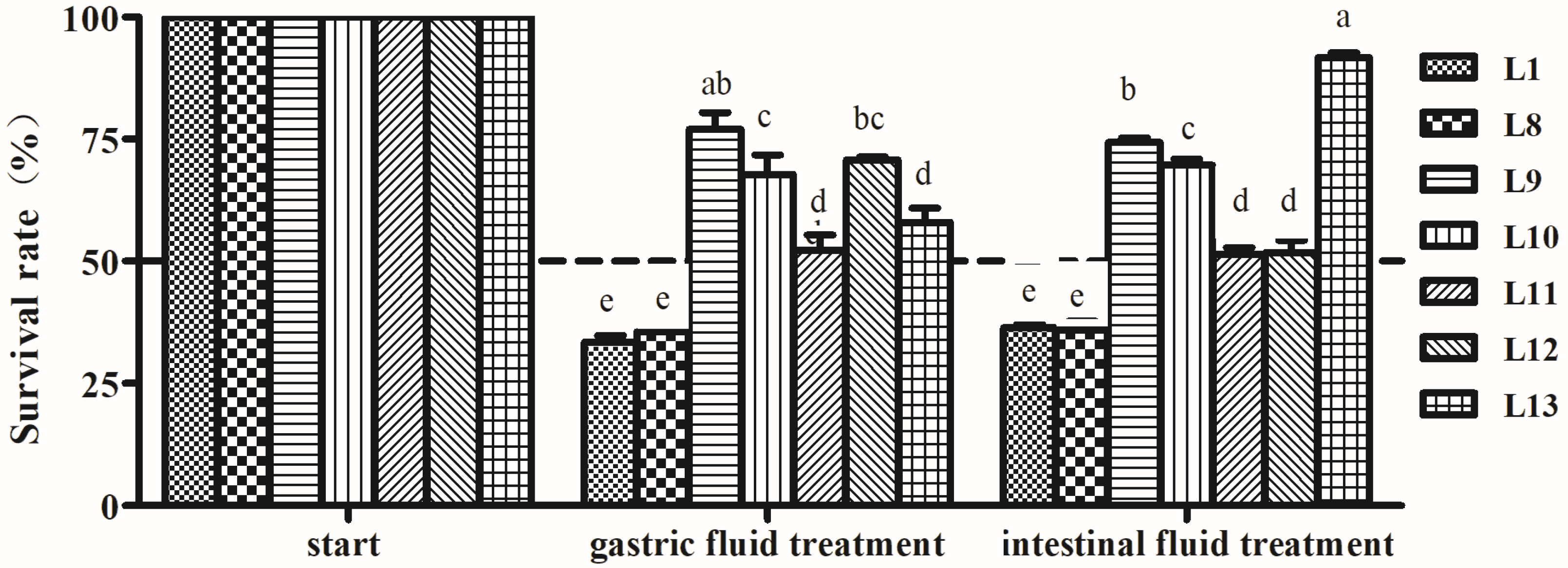
2.5. Tolerance to Artificial Gastric and Intestinal Fluids
2.6. Cell Surface Characteristics
2.6.1. Hydrophobicity
2.6.2. Auto-Aggregation Assay
2.6.3. Co-Aggregation Assay
2.7. Safety Assessment
2.7.1. Hemolytic Activity
2.7.2. Antibiotic Susceptibility
2.8. Molecular Identification
2.9. Carbon Sources Utilization
2.10. Characteristic of Antimicrobial Substances
2.11. Stastical Analysis
3. Results
3.1. Bacteria Isolation
3.2. In Vitro Antimicrobial Activity
3.3. Analysis of Growth and Acid Production
3.4. Analysis of Tolerance to the Gastrointestinal Environment
3.4.1. Acid Tolerance Analysis
3.4.2. Bile Salt Tolerance Analysis
3.4.3. Artificial Gastric and Intestinal Fluids Tolerance Analysis
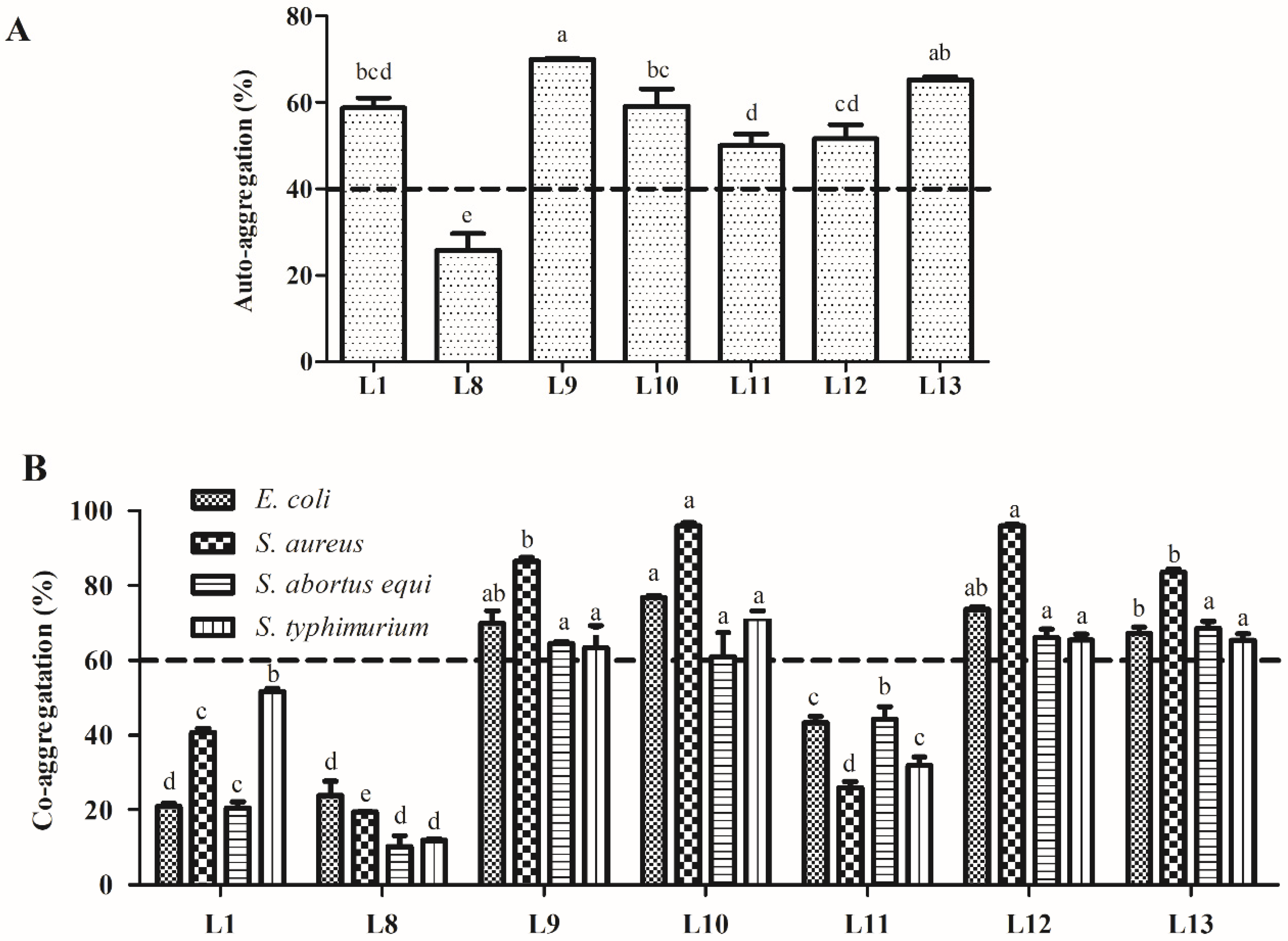
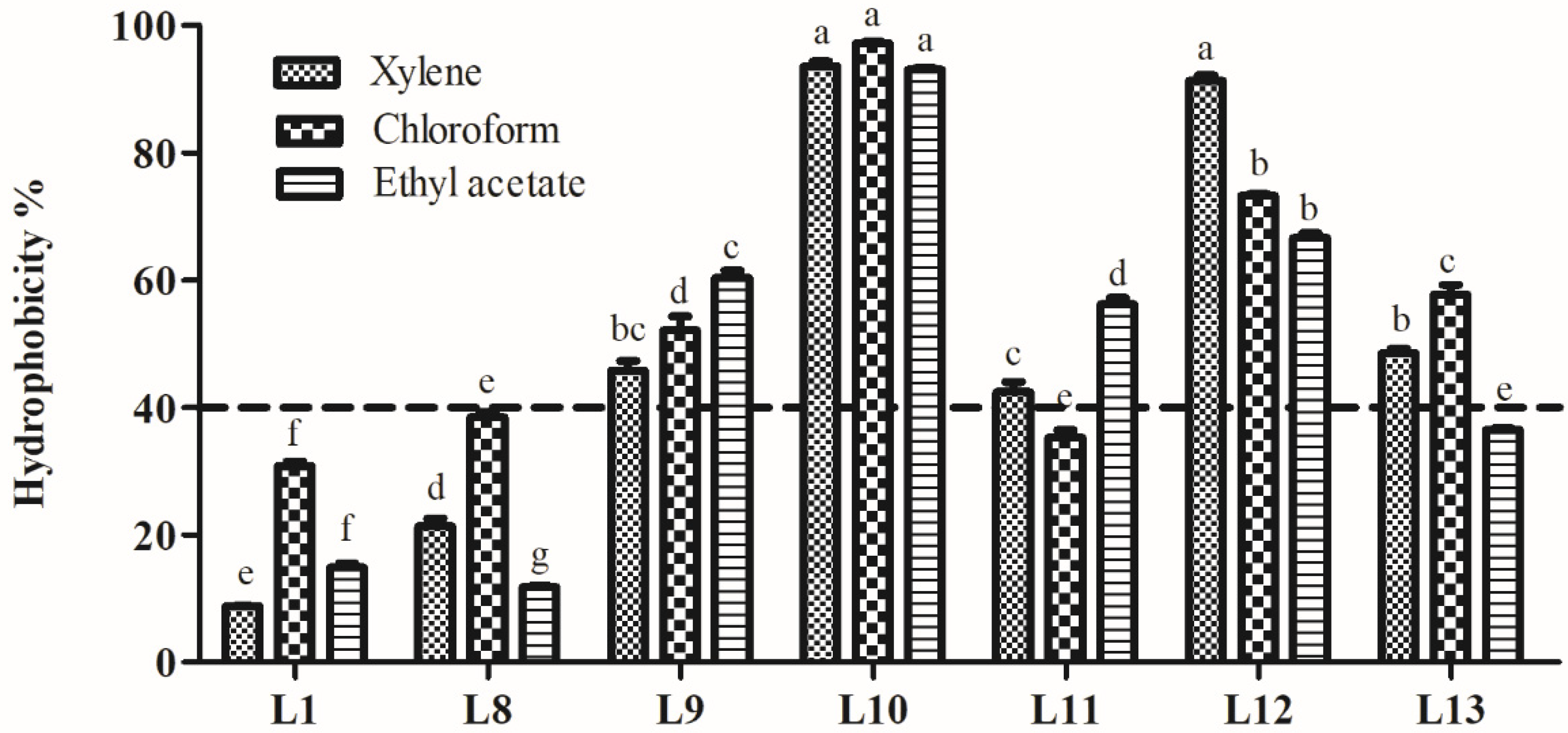
3.5. Analysis ofCell Surface Characteristics
3.5.1. Auto-Aggregation and Co-Aggregation Assay
3.5.2. Hydrophobicity Analysis
3.6. Analysis of Safety
3.7. Molecular Identification via 16S rRNA Sequencing
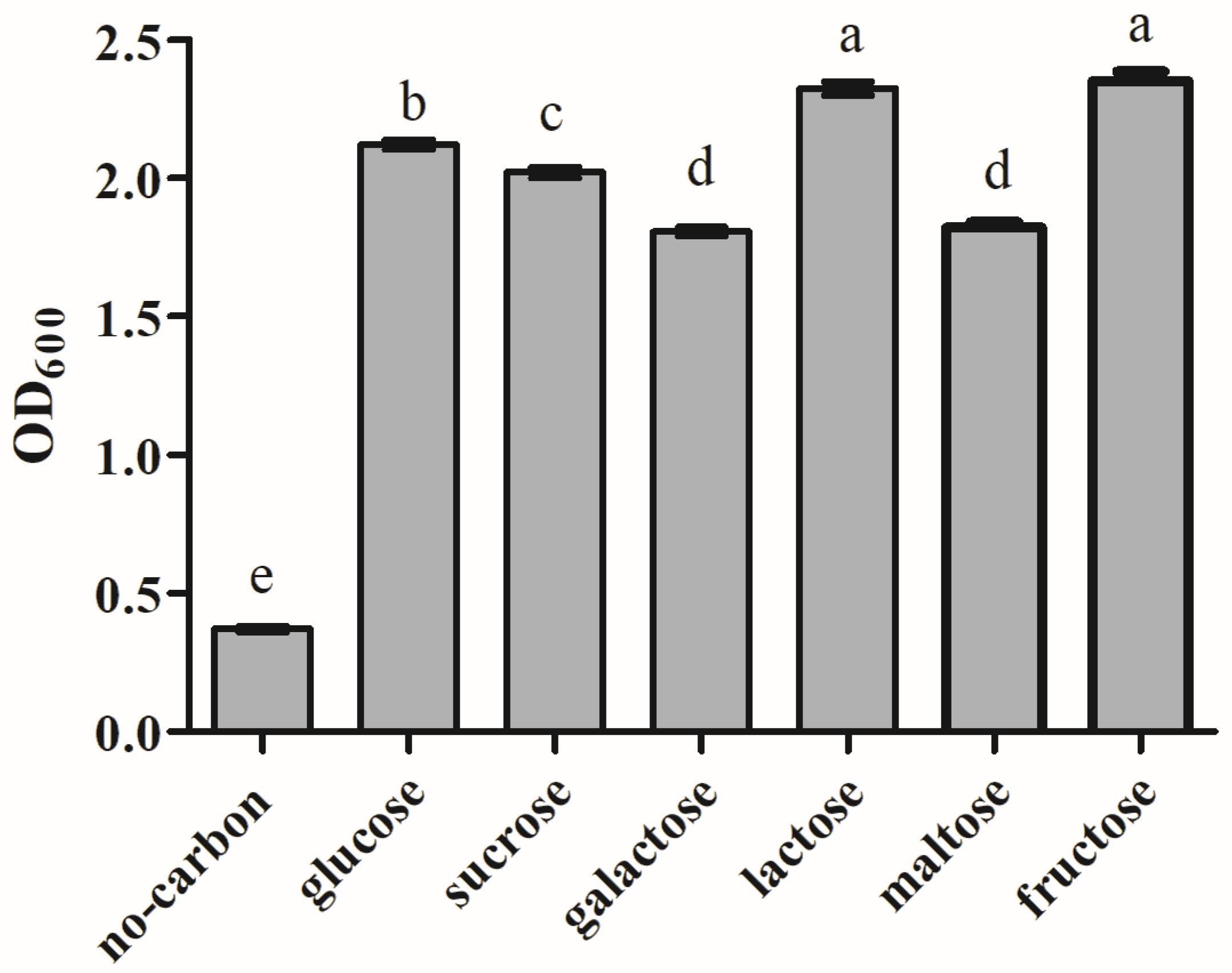
3.8. Analysis of Carbon Source Utilization
3.9. Characteristics of Antimicrobial Substances from Lactic Acid Bacteria
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, M.; Wang, Y.; Cui, H.; Li, Y.; Sun, Y.; Qiu, H.J. Characterization of lactic acid bacteria isolated from the gastrointestinal tract of a wild boar as potential probiotics. Front. Vet. Sci. 2020, 7, 49. [Google Scholar] [CrossRef]
- Duan, H.; Lu, L.; Zhang, L.; Li, J.; Gu, X.; Li, J. Effects of Lactobacillus lactis supplementation on growth performance, hematological parameters, meat quality and intestinal flora in growing-finishing pigs. Animals 2023, 13, 1247. [Google Scholar] [CrossRef]
- Kawahara, T.; Shimizu, I.; Tanaka, Y.; Tobita, K.; Tomokiyo, M.; Watanabe, I. Lactobacillus crispatus Strain KT-11 S-layer protein inhibits Rotavirus infection. Front. Microbiol. 2022, 13, 783879. [Google Scholar] [CrossRef] [PubMed]
- Kadja, L.; Dib, A.L.; Lakhdara, N.; Bouaziz, A.; Espigares, E.; Gagaoua, M. Influence of three probiotics strains, Lactobacillus rhamnosus GG, Bifidobacterium animalis subsp. lactis BB-12 and Saccharomyces boulardii CNCM I-745 on the biochemical and haematological profiles and body weight of healthy rabbits. Biology 2021, 10, 1194. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.; Liu, F.; Mao, Y.; Zhang, Y.; Zeng, H.; Ren, S.; Guo, L.; Chen, Z.; Hrabchenko, N.; et al. Mechanisms and applications of probiotics in prevention and treatment of swine diseases. Porc. Health Manag. 2023, 9, 5. [Google Scholar] [CrossRef]
- Shokryazdan, P.; Faseleh Jahromi, M.; Liang, J.B.; Ho, Y.W. Probiotics: From isolation to application. J. Am. Coll. Nutr. 2017, 36, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Özkan, E.R.; Demirci, T.; Öztürk, H.İ.; Akın, N. Screening Lactobacillus strains from artisanal Turkish goatskin casing Tulum cheeses produced by nomads via molecular and in vitro probiotic characteristics. J. Sci. Food Agric. 2021, 101, 2799–2808. [Google Scholar] [CrossRef]
- Hassan, M.U.; Nayab, H.; Shafique, F.; Williamson, M.P.; Almansouri, T.S.; Asim, N.; Shafi, N.; Attacha, S.; Khalid, M.; Ali, N.; et al. Probiotic properties of Lactobacillus helveticus and Lactobacillus plantarum isolated from traditional Pakistani yoghurt. BioMed Res. Int. 2020, 2020, 8889198. [Google Scholar] [CrossRef] [PubMed]
- Alkalbani, N.S.; Turner, M.S.; Ayyash, M.M. Isolation, identification, and potential probiotic characterization of isolated lactic acid bacteria and in vitro investigation of the cytotoxicity, antioxidant, and antidiabetic activities in fermented sausage. Microb. Cell Factories 2019, 18, 188. [Google Scholar] [CrossRef] [PubMed]
- Angmo, K.; Kumari, A.; Savitri, T.C.; Bhalla, T.C. Probiotic characterization of lactic acid bacteria isolated from fermented foods and beverage of Ladakh. LWT Food Sci. Technol. 2016, 66, 428–435. [Google Scholar] [CrossRef]
- Reuben, R.C.; Roy, P.C.; Sarkar, S.L.; Alam, A.R.U.; Jahid, I.K. Characterization and evaluation of lactic acid bacteria from indigenous raw milk for potential probiotic properties. J. Dairy Sci. 2020, 103, 1223–1237. [Google Scholar] [CrossRef]
- Abushelaibi, A.; Al-Mahadin, S.; El-Tarabily, K.; Shah, N.P.; Ayyash, M. Characterization of potential probiotic lactic acid bacteria isolated from camel milk. LWT Food Sci. Technol. 2017, 79, 316–325. [Google Scholar] [CrossRef]
- Wei, X.; Zhang, Y.; Zhou, H.; Tian, F.; Ni, Y. Antimicrobial activities and in vitro properties of cold-adapted Lactobacillus strains isolated from the intestinal tract of cold water fishes of high latitude water areas in Xinjiang, China. BMC Microbiol. 2019, 19, 247. [Google Scholar] [CrossRef]
- Reuben, R.C.; Roy, P.C.; Sarkar, S.L.; Alam, R.U.; Jahid, I.K. Isolation, characterization, and assessment of lactic acid bacteria toward their selection as poultry probiotics. BMC Microbiol. 2019, 19, 253. [Google Scholar] [CrossRef] [PubMed]
- El-Deeb, W.M.; Fayez, M.; Elsohaby, I.; Ghoneim, I.; Al-Marri, T.; Kandeel, M.; Elgioushy, M. Isolation and characterization of vaginal Lactobacillus spp. in dromedary camels (Camelus dromedarius): In vitro evaluation of probiotic potential of selected isolates. PeerJ 2020, 8, e8500. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.H.; Han, S.H.; Kim, Y.; Paek, N.S.; So, J.S. In vitro probiotic properties of Lactobacillus salivarius MG242 isolated from human vagina. Probiot. Antimicrob. Proteins 2018, 10, 343–349. [Google Scholar] [CrossRef]
- Dan, T.; Hu, H.; Tian, J.; He, B.; Tai, J.; He, Y. Influence of different ratios of Lactobacillus delbrueckii subsp. bulgaricus and Streptococcus thermophilus on fermentation characteristics of yogurt. Molecules 2023, 28, 2123. [Google Scholar] [CrossRef] [PubMed]
- Bengoa, A.A.; Iraporda, C.; Acurcio, L.B.; de Cicco Sandes, S.H.; Costa, K.; Moreira Guimarães, G.; Esteves Arantes, R.M.; Neumann, E.; Cantini Nunes, Á.; Nicoli, J.R.; et al. Physicochemical, immunomodulatory and safety aspects of milks fermented with Lactobacillus paracasei isolated from kefir. Food Res. Int. 2019, 123, 48–55. [Google Scholar] [CrossRef]
- Ruiz-Moyano, S.; Martín, A.; Benito, M.J.; Hernández, A.; Casquete, R.; de Guia Córdoba, M. Application of Lactobacillus fermentum HL57 and Pediococcus acidilactici SP979 as potential probiotics in the manufacture of traditional Iberian dry-fermented sausages. Food Microbiol. 2011, 28, 839–847. [Google Scholar] [CrossRef]
- Jin, Y.; Cao, X.; Shi, C.; Feng, B.; Huang, H.; Jiang, Y.; Wang, J.; Yang, G.; Yang, W.; Wang, C. Lactobacillus rhamnosus GG promotes early B lineage development and IgA production in the lamina propria in piglets. J. Immunol. 2021, 207, 2179–2191. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yao, B.; Gao, H.; Zang, J.; Tao, S.; Zhang, S.; Huang, S.; He, B.; Wang, J. Combined supplementation of Lactobacillus fermentum and Pediococcus acidilactici promoted growth performance, alleviated inflammation, and modulated intestinal microbiota in weaned pigs. BMC Vet. Res. 2019, 15, 239. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, E.; Hemmerling, A.; Miller, S.; Burke, K.E.; Newmann, S.J.; Morris, S.R.; Reno, H.; Huibner, S.; Kulikova, M.; Nagelkerke, N.; et al. Sustained effect of LACTIN-V (Lactobacillus crispatus CTV-05) on genital immunology following standard bacterial vaginosis treatment: Results from a randomised, placebo-controlled trial. Lancet Microbe 2022, 3, e435–e442. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; An, M.; Heo, H.; Park, J.Y.; Lee, J.; Kang, C.H. Limosilactobacillus fermentum MG4294 and Lactiplantibacillus plantarum MG5289 ameliorates nonalcoholic fatty liver disease in high-fat diet-induced mice. Nutrients 2023, 15, 2005. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Yu, L.; Tian, F.; Chen, W.; Zhai, Q. The potential therapeutic role of Lactobacillaceae rhamnosus for treatment of inflammatory bowel disease. Foods 2023, 12, 692. [Google Scholar] [CrossRef]
- Wang, S.; Peng, Q.; Jia, H.M.; Zeng, X.F.; Zhu, J.L.; Hou, C.L.; Liu, X.T.; Yang, F.J.; Qiao, S.Y. Prevention of Escherichia coli infection in broiler chickens with Lactobacillus plantarum B1. Poult. Sci. 2017, 96, 2576–2586. [Google Scholar] [CrossRef]
- Kumar, M.; Kala, A.; Chaudhary, L.C.; Agarwal, N.; Kochewad, S.A. Microencapsulated and lyophilized Lactobacillus acidophilus improved gut health and immune status of preruminant calves. Probiot. Antimicrob. Proteins 2022, 14, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Yang, K.; Zhang, A.; Chang, W.; Zheng, A.; Chen, Z.; Cai, H.; Liu, G. Effects of Lactobacillus acidophilus on the growth performance, immune response, and intestinal barrier function of broiler chickens challenged with Escherichia coli O157. Poult. Sci. 2021, 100, 101323. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Xiao, F.; Li, X.; Li, Y.; Wang, X.; Yu, G.; Zhang, T.; Wang, Y. Pediococcus pentosaceus CECT 8330 protects DSS-induced colitis and regulates the intestinal microbiota and immune responses in mice. J. Transl. Med. 2022, 20, 33. [Google Scholar] [CrossRef] [PubMed]
- Zou, Q.; Meng, W.; Wang, T.; Liu, X.; Li, D. Effect of multi-strain probiotics on the performance of AA+ male broilers. Front. Vet. Sci. 2022, 9, 1098807. [Google Scholar] [CrossRef]
- Neveling, D.P.; Dicks, L.M.T. Probiotics: An antibiotic replacement strategy for healthy broilers and productive rearing. Probiot. Antimicrob. Proteins 2021, 13, 1–11. [Google Scholar] [CrossRef]
- Sharma, A.N.; Kumar, S.; Tyagi, A.K. Effects of mannan-oligosaccharides andLactobacillus acidophilus supplementation on growth performance, nutrient utilization and faecal characteristics in Murrah buffalo calves. J. Anim. Physiol. Anim. Nutr. 2018, 102, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Lauková, A.; Kandričáková, A.; Ščerbová, J. Use of bacteriocin-producing, probiotic strain Enterococcus faecium AL41 to control intestinal microbiota in farm ostriches. Lett. Appl. Microbiol. 2015, 60, 531–535. [Google Scholar] [CrossRef]
- Weese, J.S.; Anderson, M.E.; Lowe, A.; Penno, R.; da Costa, T.M.; Button, L.; Goth, K.C. Screening of the equine intestinal microflora for potential probiotic organisms. Equine Vet, J. 2004, 36, 351–355. [Google Scholar] [CrossRef]
- Khusro, A.; Arasu, M.V.; Sahibzada, M.U.K.; Salem, A.Z.M.; Al-Dhabi, N.A.; Rivas-Caceres, R.R.; Seidel, V.; Choi, K.C. Assessment on in vitro probiotic attributes of Lactobacillus plantarum isolated from horse feces. J. Equine Vet. Sci. 2021, 107, 103769. [Google Scholar] [CrossRef] [PubMed]
- Pei, L.; Liu, J.; Huang, Z.; Iqbal, M.; Shen, Y. Effects of lactic acid bacteria isolated from Equine on Salmonella-infected gut mouse model. Probiot. Antimicrob. Proteins 2023, 15, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Xiao, M.; Liu, M.; Wang, Z.; Liu, F.; Guo, L.; Meng, H.; Zhang, H.; Yang, J.; Deng, D.; et al. Coupling metagenomics with cultivation to select host-specific probiotic micro-organisms for subtropical aquaculture. J. Appl. Microbiol. 2017, 123, 1274–1285. [Google Scholar] [CrossRef] [PubMed]
- Carminati, D.; Tidona, F.; Fornasari, M.E.; Rossetti, L.; Meucci, A.; Giraffa, G. Biotyping of cultivable lactic acid bacteria isolated from donkey milk. Lett. Appl. Microbiol. 2014, 59, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Soto Del Rio Mde, L.; Andrighetto, C.; Dalmasso, A.; Lombardi, A.; Civera, T.; Bottero, M.T. Isolation and characterisation of lactic acid bacteria from donkey milk. J. Dairy Res. 2016, 83, 383–386. [Google Scholar] [CrossRef]
- Xia, Y.; Qin, S.K.; Shen, Y.Q. Probiotic potential of Weissella strains isolated from horse feces, a probable equine probiotic. Microb. Pathog. 2019, 132, 117–123. [Google Scholar] [CrossRef]
- Azat, R.; Liu, Y.; Li, W.; Kayir, A.; Lin, D.B.; Zhou, W.W.; Zheng, X.D. Probiotic properties of lactic acid bacteria isolated from traditionally fermented Xinjiang cheese. J. Zhejiang Univ. Sci. B 2016, 17, 597–609. [Google Scholar] [CrossRef]
- Baccouri, O.; Boukerb, A.M.; Farhat, L.B.; Zébré, A.; Zimmermann, K.; Domann, E.; Cambronel, M.; Barreau, M.; Maillot, O.; Rincé, I.; et al. Probiotic potential and safety evaluation of Enterococcus faecalis OB14 and OB15, isolated from traditional Tunisian Testouri cheese and Rigouta, using physiological and genomic analysis. Front. Microbiol. 2019, 10, 881. [Google Scholar] [CrossRef]
- Liu, Z.J.; Xu, C.; Tian, R.; Wang, W.; Ma, J.G.; Gu, L.Y.; Liu, F.; Jiang, Z.M.; Hou, J.C. Screening beneficial bacteriostatic lactic acid bacteria in the intestine and studies of bacteriostatic substances. J. Zhejiang Univ. Sci. B. 2021, 22, 533–547. [Google Scholar] [CrossRef]
- Solieri, L.; Bianchi, A.; Mottolese, G.; Lemmetti, F.; Giudici, P. Tailoring the probiotic potential of non-starter Lactobacillus strains from ripened Parmigiano Reggiano cheese by in vitro screening and principal component analysis. Food Microbiol. 2014, 38, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Charpteris, W.P.; Kelly, P.M.; Morelli, L.; Collins, J.K. Antibiotic Susceptibility of Potentially Probiotic Lactobacillus Species. J. Food Prot. 1998, 12, 1636–1643. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 4, 2758–2782. [Google Scholar] [CrossRef] [PubMed]
- Uzzau, S.; Brown, D.J.; Wallis, T.; Rubino, S.; Leori, G.; Bernard, S.; Casadesús, J.; Platt, D.J.; Olsen, J.E. Host adapted serotypes of Salmonella enterica. Epidemiol. Infect. 2000, 125, 229–255. [Google Scholar] [CrossRef]
- Niwa, H.; Hobo, S.; Kinoshita, Y.; Muranaka, M.; Ochi, A.; Ueno, T.; Oku, K.; Hariu, K.; Katayama, Y. Aneurysm of the cranial mesenteric artery as a site of carriage of Salmonella enterica subsp. enterica serovar Abortusequi in the horse. J. Vet. Diagn. Investig. 2016, 28, 440–444. [Google Scholar] [CrossRef]
- de Almeida Júnior, W.L.G.; Ferrari, W.; de Souza, J.V.; da Silva, C.D.A.; da Costa, M.M.; Dias, F.S. Characterization and evaluation of lactic acid bacteria isolated from goat milk. Food Control 2015, 53, 96–103. [Google Scholar] [CrossRef]
- Tsai, C.C.; Hsih, H.Y.; Chiu, H.H.; Lai, Y.Y.; Liu, J.H.; Yu, B.; Tsen, H.Y. Antagonistic activity against Salmonella infection in vitro and in vivo for two Lactobacillus strains from swine and poultry. Int. J. Food Microbiol. 2005, 102, 185–194. [Google Scholar] [CrossRef]
- Tang, H.; Qian, B.; Xia, B.; Zhuan, Y.; Yao, Y.; Gan, R.; Zhang, J. Screening of lactic acid bacteria isolated from fermented Cornus officinalis fruits for probiotic potential. J. Food Saf. 2018, 38, e12565. [Google Scholar] [CrossRef]
- Del Re, B.; Sgorbati, B.; Miglioli, M.; Palenzona, D. Adhesion, autoaggregation and hydrophobicity of 13 strains of Bifidobacterium longum. Lett. Appl. Microbiol. 2000, 31, 438–442. [Google Scholar] [CrossRef]
- Sharma, K.; Attri, S.; Goel, G. Selection and evaluation of probiotic and functional characteristics of autochthonous lactic acid bacteria isolated from fermented wheat flour dough Babroo. Probiot. Antimicrob. Proteins 2019, 11, 774–784. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, A.; Nawaz, M.; Rabbani, M.; Mushtaq, M.H. In vitro characterization of probiotic potential of Limosilactobacillus fermentum against Salmonella gallinarum causing fowl typhoid. Animals 2023, 13, 1284. [Google Scholar] [CrossRef]
- Potočnjak, M.; Pušić, P.; Frece, J.; Abram, M.; Janković, T.; Gobin, I. Three new Lactobacillus plantarum Strains in the probiotic toolbox against gut pathogen Salmonella enterica serotype Typhimurium. Food Technol. Biotechnol. 2017, 55, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Boris, S.; Suárez, J.E.; Vázquez, F.; Barbés, C. Adherence of human vaginal lactobacilli to vaginal epithelial cells and interaction with uropathogens. Infect. Immun. 1998, 66, 1985–1989. [Google Scholar] [CrossRef]
- Lin, W.C.; Ptak, C.P.; Chang, C.Y.; Ian, M.K.; Chia, M.Y.; Chen, T.H.; Kuo, C.J. Autochthonous lactic acid bacteria isolated from dairy cow feces exhibiting promising probiotic properties and in vitro antibacterial activity against foodborne pathogens in cattle. Front. Vet. Sci. 2020, 7, 239. [Google Scholar] [CrossRef]
- Zhang, S.; Oh, J.H.; Alexander, L.M.; Özçam, M.; van Pijkeren, J.P. d-Alanyl-d-Alanine ligase as a broad-host-range counterselection marker in vancomycin-resistant lactic acid bacteria. J. Bacteriol. 2018, 200, e00607-17. [Google Scholar] [CrossRef] [PubMed]
- Caggia, C.; De Angelis, M.; Pitino, I.; Pino, A.; Randazzo, C.L. Probiotic features of Lactobacillus strains isolated from Ragusano and Pecorino Siciliano cheeses. Food Microbiol. 2015, 50, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Indo, Y.; Kitahara, S.; Tomokiyo, M.; Araki, S.; Islam, M.A.; Zhou, B.; Albarracin, L.; Miyazaki, A.; Ikeda-Ohtsubo, W.; Nochi, T.; et al. Ligilactobacillus salivarius Strains isolated from the porcine gut modulate innate immune responses in epithelial cells and improve protection against intestinal viral-bacterial superinfection. Front. Immunol. 2021, 12, 652923. [Google Scholar] [CrossRef]
- Lukasik, J.; Dierikx, T.; Besseling-van Der Vaart, I.; De Meij, T.; Szajewska, H. Multispecies probiotic for the prevention of antibiotic-associated diarrhea in children: A randomized clinical trial. JAMA Pediatr. 2022, 176, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, S.; Mittal, V.; Singh, A. In vitro evaluation of probiotic potential and safety assessment of Lactobacillus mucosae strains isolated from donkey’s lactation. Probiot. Antimicrob. Proteins 2020, 12, 1045–1056. [Google Scholar] [CrossRef]
- Benameur, F.; Belkaaloul, K.; Kheroua, O. Isolation of 60 strains from fermented milk of mares and donkeys in Algeria and identification by 16S rRNA sequencing of lactobacilli: Assessment of probiotic skills of important strains and aromatic productivity power. Vet. World 2024, 17, 829–841. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Antimicrobial Activity | |||
|---|---|---|---|---|
| E. coli ATCC 25922 | S. aureus ATCC 25923 | S. typhimurium ATCC 14028 | S. abortus equi CGMCC 18046 | |
| L1 | 16.1 ± 0.1 | 16.5 ± 0.5 | 15.7 ± 0.1 | 11.2 ± 0.3 |
| L2 | 16.4 ± 0.2 | 15.4 ± 0.2 | 16.8 ± 0.2 | - |
| L3 | 16.1 ± 0.1 | 14.1 ± 0.2 | 11.9 ± 0.1 | - |
| L4 | 15.8 ± 0.3 | - | 11.0 ± 0.0 | - |
| L5 | 16.0 ± 0.2 | 15.7 ± 0.3 | 14.6 ± 0.2 | - |
| L6 | 17.2 ± 0.2 | 13.4 ± 0.2 | 15.0 ± 0.1 | - |
| L7 | 16.7 ± 0.1 | 13.0 ± 0.1 | 16.5 ± 0.6 | 11.2 ± 0.3 |
| L8 | 15.3 ± 0.3 | 13.3 ± 0.3 | 13.9 ± 0.1 | 13.4 ± 0.3 |
| L9 | 18.1 ± 0.3 | 21.5 ± 0.2 | 20.2 ± 0.1 | 19.7 ± 0.5 |
| L10 | 20.3 ± 0.8 | 21.1 ± 0.5 | 19.5 ± 0.1 | 19.4 ± 0.1 |
| L11 | 16.7 ± 0.3 | 13.9 ± 0.1 | 13.8 ± 0.3 | 13.5 ± 0.3 |
| L12 | 19.5 ± 0.1 | 18.9 ± 0.5 | 18.2 ± 0.2 | 16.9 ± 0.5 |
| L13 | 17.1 ± 0.1 | 18.2 ± 0.3 | 14.6 ± 0.1 | 16.2 ± 0.1 |
| Strains | Hemolytic | Antibiotic Sensitivity | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TC | GM | P | VA | CC | K | E | CRD | CIP | CRO | C | NOR | ||
| L9 | No | S | R | S | R | S | R | S | S | S | S | S | MS |
| L10 | No | R | R | S | S | S | R | S | R | R | S | S | R |
| L12 | No | R | R | MS | S | S | R | S | R | R | S | S | R |
| Lactic Acid Bacteria Strain | Treatment | Zone of Inhibition (mm) | |||
|---|---|---|---|---|---|
| E. coli | S. aureus | S. typhimurium | S. abortus equi | ||
| Untreated | +++ | +++ | +++ | ++ | |
| Heat-treated | +++ | +++ | +++ | ++ | |
| L9 | Neutralized | - | - | - | - |
| Catalase-treated | +++ | +++ | +++ | ++ | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Yue, S.; Yu, J.; Bian, F.; Chen, G.; Zhang, Y. Probiotic Characterization of Lactic Acid Bacteria from Donkey Feces in China. Animals 2025, 15, 207. https://doi.org/10.3390/ani15020207
Wu Y, Yue S, Yu J, Bian F, Chen G, Zhang Y. Probiotic Characterization of Lactic Acid Bacteria from Donkey Feces in China. Animals. 2025; 15(2):207. https://doi.org/10.3390/ani15020207
Chicago/Turabian StyleWu, Yanqiu, Shousong Yue, Jinhui Yu, Fei Bian, Gao Chen, and Yan Zhang. 2025. "Probiotic Characterization of Lactic Acid Bacteria from Donkey Feces in China" Animals 15, no. 2: 207. https://doi.org/10.3390/ani15020207
APA StyleWu, Y., Yue, S., Yu, J., Bian, F., Chen, G., & Zhang, Y. (2025). Probiotic Characterization of Lactic Acid Bacteria from Donkey Feces in China. Animals, 15(2), 207. https://doi.org/10.3390/ani15020207