
A Comparative Study of the Effect of Including Full-Fat Tenebrio molitor for Replacing Conventional Ingredients in Practical Diets for Dicentrarchus labrax Juveniles

 ,
,  ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Meal and Experimental Diets
2.2. Fish and Experimental Designs
2.3. Fish Sampling
2.4. Growth Performance and Nutrient Utilization
2.5. Biochemical Analysis
2.5.1. Proximate Compositions of Diets and Tissues
2.5.2. Total Lipids, Lipid Classes, and FA Analyses
2.6. Statistical Analysis
3. Results
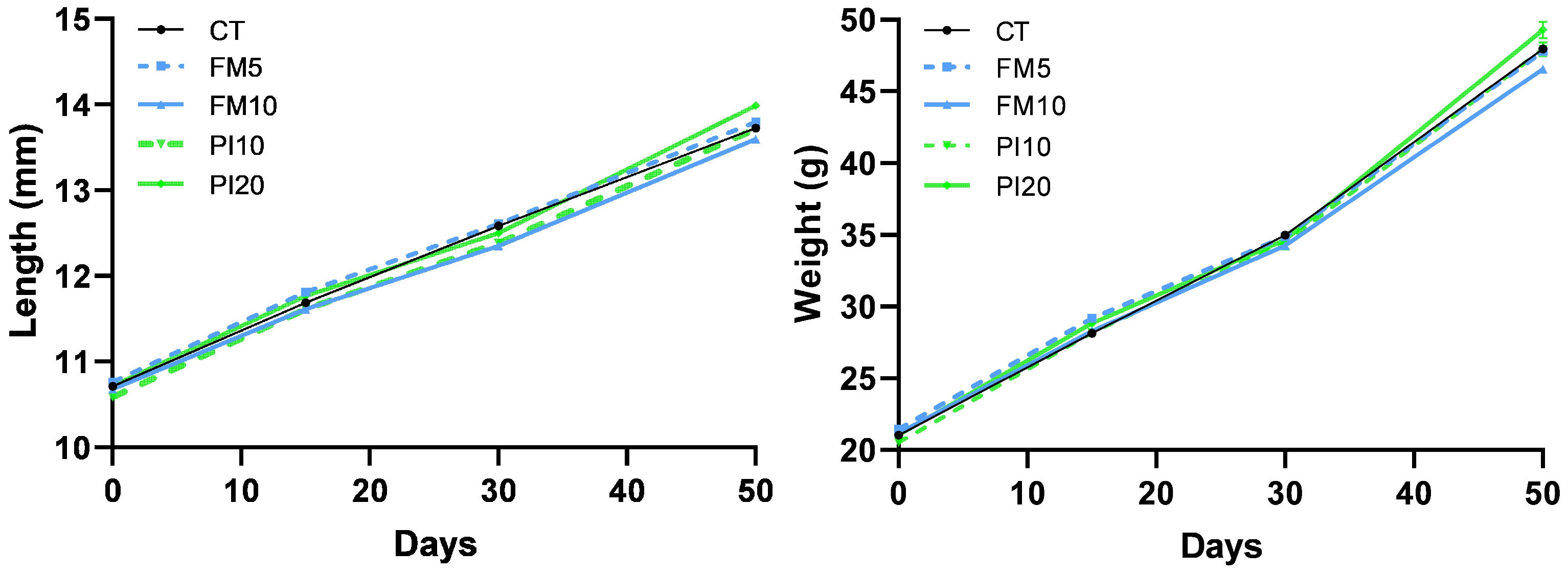
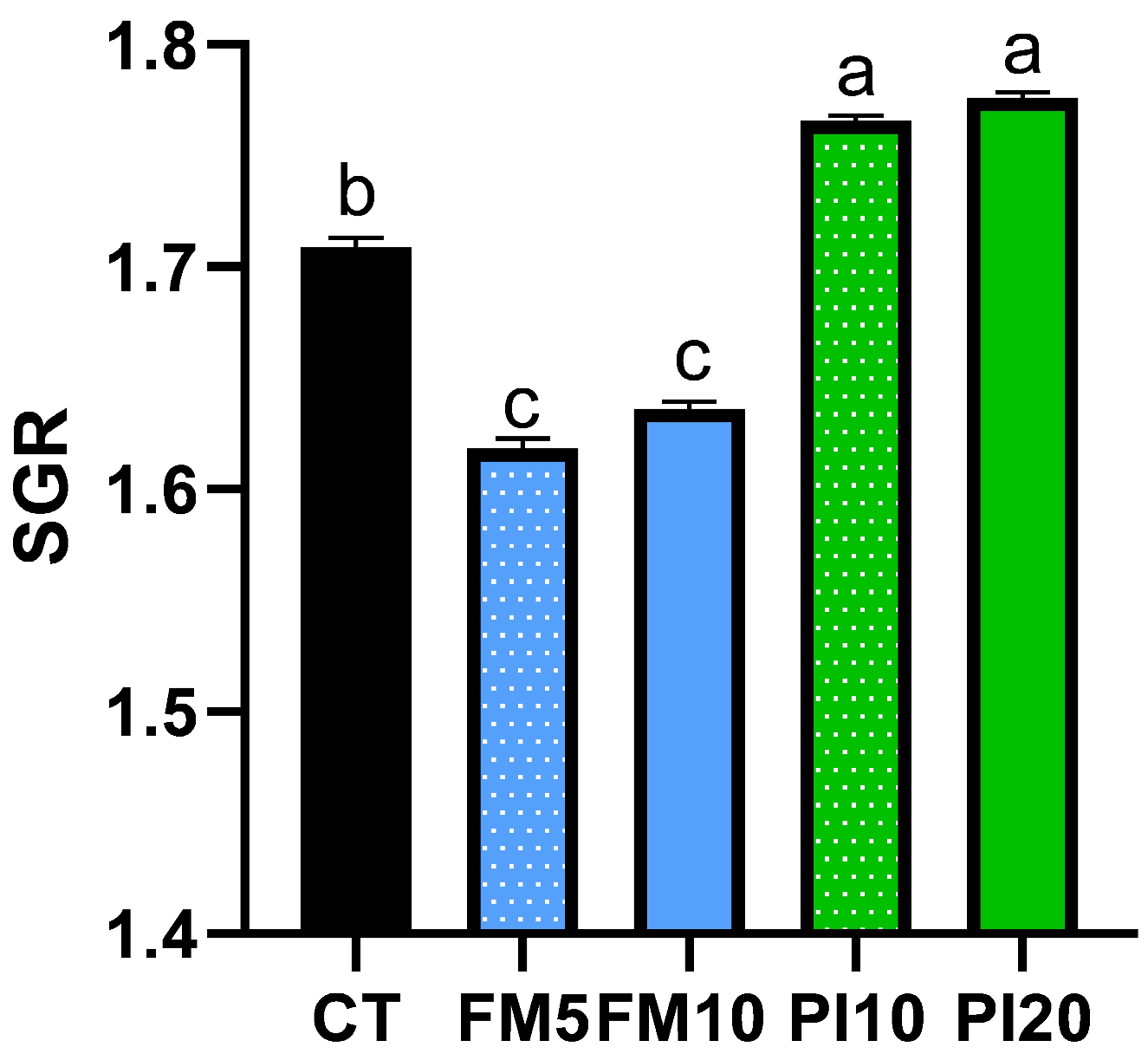
3.1. Growth Performance
3.2. Lipid Profile
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. El estado mundial de la pesca y la acuicultura 2022; Hacia la Transformación Azul; FAO: Roma, Italy, 2022. [Google Scholar]
- Hemre, G.-I.; Amlund, H.; Aursand, M.; Bakke, A.M.; Olsen, R.E.; Ringø, E.; Svihus, B.; Bernhoft, A.; Jenssen, B.M.; Møretrø, T.; et al. Criteria for safe use of plant ingredients in diets for aquacultured fish. Eur. J. Nutr. Food Saf. 2018, 8, 240–242. [Google Scholar] [CrossRef]
- Nizza, A.; Piccolo, G.; Loponte, R.; Marono, S.; Bovera, F.; Parisi, G.; Gasco, L. Use of Tenebrio molitor larvae meal as protein source in broiler diet: Effect on growth performance, nutrient digestibility, and carcass and meat traits. J. Anim. Sci. 2016, 94, 639–647. [Google Scholar] [CrossRef]
- Medrano Vega, L.C. Larvas de Gusano de Harina (Tenebrio molitor) Como Alternativa Proteica en la Alimentación Animal; Universidad Nacional Abierta ya Distancia UNAD: Bogotá, Colombia, 2019. [Google Scholar]
- Finke, M.D. Complete nutrient composition of commercially raised invertebrates used as food for insectivores. Zoo Biol. 2002, 21, 269–285. [Google Scholar] [CrossRef]
- de Haro Domínguez, C. Evaluación de la Harina de Insectos Como Fuente Alternativa a la Harina de Pescado en Piensos Para Acuicultura. Doctoral Dissertation, Universidad de Almería, La Cañada de San Urbano, Spain, 2015. [Google Scholar]
- Quang Tran, H.; Van Doan, H.; Stejskal, V. Environmental consequences of using insect meal as an ingredient in aquafeeds: A systematic view. Rev. Aquac. 2022, 14, 237–251. [Google Scholar] [CrossRef]
- Hachero-Cruzado, I.; Betancor, M.B.; Coronel-Dominguez, A.J.; Manchado, M.; Alarcón-López, F.J. Assessment of Full-Fat Tenebrio molitor as Feed Ingredient for Solea senegalensis: Effects on Growth Performance and Lipid Profile. Animals 2024, 14, 595. [Google Scholar] [CrossRef] [PubMed]
- De Marco, M.; Martínez, S.; Hernandez, F.; Madrid, J.; Gai, F.; Rotolo, L.; Belforti, M.; Bergero, D.; Katz, H.; Dabbou, S.; et al. Nutritional value of two insect larval meals (Tenebrio molitor and Hermetia illucens) for broiler chickens: Apparent nutrient digestibility, apparent ileal amino acid digestibility and apparent metabolizable energy. Anim. Feed Sci. Technol. 2015, 209, 211–218. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schlüter, O.K. Potential and challenges of insects as an innovative source for food and feed production. Innov. Food Sci. Emerg. Technol. 2013, 17, 1–11. [Google Scholar] [CrossRef]
- Mariod, A.A.; Saeed Mirghani, M.E.; Hussein, I. Chapter 50—Tenebrio molitor Mealworm. In Unconventional Oilseeds and Oil Sources 2017; Mariod, A.A., Mirghani, M.E.S., Hussein, I., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 331–336. [Google Scholar]
- Koutsos, L.; McComb, A.; Finke, M. Insect Composition and Uses in Animal Feeding Applications: A Brief Review. Ann. Entomol. Soc. Am. 2019, 112, 544–551. [Google Scholar] [CrossRef]
- Valdez Pantoja, C.; Untiveros Bermúdez, G. Extracción y caracterización del aceite de las larvas del Tenebrio molitor. Rev. Soc. Química Perú 2010, 76, 407–414. [Google Scholar]
- Alfijo, Y.; Xie, D.; Astuti, R.T.; Wong, J.; Wang, L. Insects as a feed ingredient for fish culture: Status and trends. Aquac. Fish. 2022, 7, 166–178. [Google Scholar] [CrossRef]
- Sánchez-Muros, M.; De Haro, C.; Sanz, A.; Trenzado, C.E.; Villareces, S.; Barroso, F.G. Nutritional evaluation of Tenebrio molitor meal as fishmeal substitute for tilapia (Oreochromis niloticus) diet. Aquac. Nutr. 2016, 22, 943–955. [Google Scholar] [CrossRef]
- Chemello, G.; Renna, M.; Caimi, C.; Guerreiro, I.; Oliva-Teles, A.; Enes, P.; Biasato, I.; Schiavone, A.; Gai, F.; Gasco, L. Partially defatted Tenebrio molitor larva meal in diets for grow-out rainbow trout, Oncorhynchus mykiss (Walbaum): Effects on growth performance, diet digestibility and metabolic responses. Animals 2020, 10, 229. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.M.; Khosravi, S.; Mauliasari, I.R.; Lee, S.M. Dietary inclusion of mealworm (Tenebrio molitor) meal as an alternative protein source in practical diets for rainbow trout (Oncorhynchus mykiss) fry. Fish. Aquat. Sci. 2020, 23. [Google Scholar] [CrossRef]
- Mastoraki, M.; Ferrándiz, P.M.; Vardali, S.C.; Kontodimas, D.C.; Kotzamanis, Y.P.; Gasco, L.; Chatzifotis, S.; Antonopoulou, E. A comparative study on the effect of fish meal substitution with three different insect meals on growth, body composition and metabolism of European sea bass (Dicentrarchus labrax L.). Aquaculture 2020, 528, 735511. [Google Scholar] [CrossRef]
- Basto, A.; Matos, E.; Valente, L.M. Nutritional value of different insect larvae meals as protein sources for European sea bass (Dicentrarchus labrax) juveniles. Aquaculture 2020, 521, 735085. [Google Scholar] [CrossRef]
- Piccolo, G.; Iaconisi, V.; Marono, S.; Gasco, L.; Loponte, R.; Nizza, S.; Bovera, F.; Parisi, G. Effect of Tenebrio molitor larvae meal on growth performance, in vivo nutrients digestibility, somatic and marketable indexes of gilthead sea bream (Sparus aurata). Anim. Feed Sci. Technol. 2017, 226, 12–20. [Google Scholar] [CrossRef]
- Biancarosa, I.; Sele, V.; Belghit, I.; Ørnsrud, R.; Lock, E.J.; Amlund, H. Replacing fish meal with insect meal in the diet of Atlantic salmon (Salmo salar) does not impact the amount of contaminants in the feed and it lowers accumulation of arsenic in the fillet. Food Addit. Contam. Part A 2019, 36, 1191–1205. [Google Scholar] [CrossRef]
- Ido, A.; Hashizume, A.; Ohta, T.; Takahashi, T.; Miura, C.; Miura, T. Replacement of fish meal by defatted yellow mealworm (Tenebrio molitor) larvae in diet improves growth performance and disease resistance in red seabream (Pargus Major). Anim. 2019, 9, 100. [Google Scholar] [CrossRef] [PubMed]
- Basto, A.; Valente, L.M.; Conde-Sieira, M.; Soengas, J.L. Central regulation of food intake is not affected by inclusion of defatted Tenebrio molitor larvae meal in diets for European sea bass (Dicentrarchus labrax). Aquaculture 2021, 544, 737088. [Google Scholar] [CrossRef]
- Ennayer, I.; Chadli, H.; Chebbaki, K.; Akharbach, H.; Zim, J.; Nhhala, H.; Bouhdid, S.; Chiaar, A.; Sarehane, M.; Chairi, H. The effect of the substitution of fishmeal with mealworm (Tenebrio molitor) on the survival, growth and quality of the European seabass (Dicentrarchus labrax) reared in Morocco. Egypt. J. Aquat. Biol. Fish. 2022, 26, 727–744. [Google Scholar] [CrossRef]
- Basto, A.; Marques, A.; Silva, A.; Sá, T.; Sousa, V.; Oliveira, M.B.P.; Aires, T.; Valente, L.M. Nutritional, organoleptic and sensory quality of market-sized European sea bass (Dicentrarchus labrax) fed defatted Tenebrio molitor larvae meal as main protein source. Aquaculture 2023, 566, 739210. [Google Scholar] [CrossRef]
- Reyes Ruiz, M. Efecto en el Crecimiento e Índices Nutritivos de la Sustitución de Harina de Pescado por Harina de Insecto en Lubina (Dicentrarchus Labrax); Universidad de Almería: La Cañada de San Urbano, Spain, 2020. [Google Scholar]
- Gasco, L.; Gai, F.; Piccolo, G.; Rotolo, L.; Lussiana, C.; Molla, P.; Chatzifotis, S. Substitution of fishmeal by Tenebrio molitor meal in the diet of Dicentrarchus labrax juveniles. In 1st International conference “Insects to Feed the World”; Universitá di Torino: Via Pietro Giuria, Italy, 2014; p. 70. [Google Scholar]
- Gasco, L.; Henry, M.; Piccolo, G.; Marono, S.; Gai, F.; Renna, M.; Lussiana, C.; Antonopoulou, E.; Mola, P.; Chatzifotis, S. Tenebrio molitor meal in diets for European sea bass (Dicentrarchus labrax L.) juveniles: Growth performance, whole body composition and in vivo apparent digestibility. Anim. Feed Sci. Technol. 2016, 220, 34–45. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International, 17th ed.; Association of Official Analytical Communities: Gaithersburg, MD, USA, 2002. [Google Scholar]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.E.; Henderson, R.J. The rapid analysis of neutral and polar marine lipids using double-development HPTLC and scanning densitometry. J. Exp. Mar. Biol. Ecol. 1989, 129, 189–197. [Google Scholar] [CrossRef]
- Christie, W.W. Lipid Analysis: Isolation, Separation, Identification and Structural Analysis of Lipids; Amer Oil Chemists Society: Bridgwater, UK, 2003. [Google Scholar]
- Randazzo, B.; Di Marco, P.; Zarantoniello, M.; Daniso, E.; Cerri, R.; Finoia, M.G.; Capoccioni, F.; Tibaldi, E.; Olivotto, I.; Cardinaletti, G. Effects of supplementing a plant protein-rich diet with insect, crayfish or microalgae meals on gilthead sea bream (Sparus aurata) and European seabass (Dicentrarchus labrax) growth, physiological status and gut health. Aquaculture 2023, 575, 739811. [Google Scholar] [CrossRef]
- Magnoni, L.J.; Gonçalves, O.; Cardoso, P.G.; Silva-Brito, F.; Ozório, R.O.A.; Palma, M.; Viegas, I. Growth performance and quality indicators of European seabass (Dicentrarchus labrax) fed diets including refined glycerol. Aquaculture 2023, 569, 739377. [Google Scholar] [CrossRef]
- Jeong, S.M.; Khosravi, S.; Yoon, K.Y.; Kim, K.W.; Lee, B.J.; Hur, S.W.; Lee, S.M. Mealworm, Tenebrio molitor, as a feed ingredient for juvenile olive flounder, Paralichthys Olivaceus. Aquac. Rep. 2021, 20, 100747. [Google Scholar] [CrossRef]
- Iaconisi, V.; Marono, S.; Parisi, G.; Gasco, L.; Genovese, L.; Maricchiolo, G.; Bovera, F.; Piccolo, G. Dietary inclusion of Tenebrio molitor larvae meal: Effects on growth performance and final quality treats of blackspot sea bream (Pagellus bogaraveo). Aquaculture 2017, 476, 49–58. [Google Scholar] [CrossRef]
- Fabrikov, D.; Barroso, F.G.; Sánchez-Muros, M.J.; Hidalgo, M.C.; Cardenete, G.; Tomás-Almenar, C.; Melenchón, F.; Guil-Guerrero, J.L. Effect of feeding with insect meal diet on the fatty acid compositions of sea bream (Sparus aurata), tench (Tinca tinca) and rainbow trout (Oncorhynchus mykiss) fillets. Aquaculture 2021, 545, 737170. [Google Scholar] [CrossRef]
- Borges, P.; Reis, B.; Fernandes, T.J.R.; Palmas, Â.; Castro-Cunha, M.; Médale, F.; Oliveira, M.B.P.P.; Valente, L.M.P. Senegalese sole juveniles can cope with diets devoid of supplemental fish oil while preserving flesh nutritional value. Aquaculture 2014, 418–419, 116–125. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Ingredients (% dw) | TM | CT | FM5 | FM10 | PI10 | PI20 |
|---|---|---|---|---|---|---|
| Fishmeal LT94 1 | 20.0 | 16.5 | 13.1 | 20.0 | 20.0 | |
| Squid meal 2 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | |
| CPSP90 3 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | |
| Krill meal 4 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | |
| Wheat gluten 5 | 11.0 | 11.0 | 11.0 | 9.3 | 7.4 | |
| Soybean protein concentrate 6 | 23.0 | 23.0 | 23.0 | 20.2 | 18.2 | |
| Pea protein concentrate 7 | 6.0 | 6.0 | 6.0 | 4.3 | 2.3 | |
| Full-fat Tenebrio molitor 8 | 5.0 | 10.0 | 10.0 | 20.0 | ||
| Fish oil 9 | 8.3 | 8.3 | 8.3 | 8.3 | 8.3 | |
| Soybean oil 10 | 7.0 | 5.7 | 4.3 | 3.5 | 0.1 | |
| Soybean lecithin 11 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | |
| Wheat flour 12 | 12.5 | 12.3 | 12.1 | 12.2 | 11.5 | |
| Choline chloride 13 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | |
| Betaine 14 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | |
| Vitamin and Mineral premix15 | 2.0 | 2.0 | 2.0 | 2.0 | 2.0 | |
| Vitamin C 16 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | |
| Lysine 17 | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 | |
| Methionine 18 | 0.6 | 0.6 | 0.6 | 0.6 | 0.6 | |
| Monoammonium phosphate 19 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | |
| Guar gum 20 | 2.0 | 2.0 | 2.0 | 2.0 | 2.0 | |
| Crude protein (% dw) | 43.77 | 46.7 | 47.0 | 47.2 | 47.3 | 47.8 |
| Crude lipid (% dw) | 35.05 | 21.8 | 21.1 | 21.5 | 21.0 | 21.8 |
| Ash (% dw) | 2.92 | 5.52 | 5.98 | 6.16 | 5.80 | 5.16 |
| Moisture (% dw) | 7.78 | 8.35 | 9.07 | 9.49 | 8.21 | 7.84 |
| Diets | ||||||
|---|---|---|---|---|---|---|
| TM | CT | FM5 | FM10 | PI10 | PI20 | |
| Lipid classes (%TL) | ||||||
| Total phospholipids | 5.9 | 8.24 | 9.9 | 9.6 | 9.2 | 8.7 |
| CHO | 7.4 | 10.8 | 12.1 | 12.0 | 11.0 | 12.5 |
| FFA | 34.6 | 9.9 | 13.0 | 12.6 | 13.1 | 12.5 |
| TAG | 43.4 | 49.4 | 41.3 | 48.6 | 47.4 | 44.3 |
| SE | 4.1 | 1.5 | 3.2 | 1.4 | 2.9 | 3.0 |
| Fatty acids % (TFA) | ||||||
| 14:0 | 4.2 | 1.2 | 1.3 | 1.4 | 1.7 | 2.1 |
| 16:0, PA | 18.0 | 16.0 | 16.1 | 16.5 | 17.4 | 18.1 |
| 18:0 | 2.5 | 4.9 | 4.6 | 4.7 | 4.6 | 4.3 |
| Total saturated FA | 24.7 | 23.6 | 23.6 | 24.0 | 25.2 | 25.9 |
| 16:1n-7 | 2.8 | 2.4 | 2.4 | 2.5 | 2.9 | 3.3 |
| 18:1n-9, OA | 51.7 | 19.0 | 22.1 | 25.5 | 25.2 | 30.9 |
| 18:1n-7 | 0.1 | 2.2 | 2.0 | 2.0 | 2.3 | 2.0 |
| 20:1n-9 | 0.1 | 1.0 | 0.9 | 0.9 | 1.0 | 0.9 |
| 22:1n-11 | nd | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 |
| Total MUFA | 55.9 | 25.8 | 28.9 | 32.5 | 33.1 | 39.0 |
| 18:2n-6, LA | 18.0 | 29.9 | 27.3 | 23.8 | 21.7 | 15.8 |
| 20:4n-6, ARA | nd | 0.8 | 0.8 | 0.9 | 0.8 | 1.0 |
| 22:5n-6 | nd | 0.6 | 0.6 | 0.6 | 0.6 | 0.6 |
| Total n-6 PUFA | 18.0 | 31.8 | 29.1 | 25.7 | 23.5 | 17.8 |
| 18:3n-3, LNA | 0.3 | 3.4 | 2.9 | 2.2 | 2.1 | 1.2 |
| 18:4n-3 | nd | 0.4 | 0.4 | 0.3 | 0.4 | 0.4 |
| 20:5n-3, EPA | nd | 3.4 | 3.2 | 3.1 | 3.4 | 3.3 |
| 22:6n-3, DHA | nd | 9.0 | 9.7 | 9.5 | 10.0 | 9.6 |
| Total n-3 PUFA | 0.3 | 17.3 | 17.2 | 16.2 | 17.0 | 15.5 |
| n-3PUFA/n-6PUFA | 0.02 | 0.54 | 0.59 | 0.63 | 0.72 | 0.87 |
| EPA/DHA | 0.00 | 0.39 | 0.33 | 0.32 | 0.34 | 0.34 |
| UFA/SFA | 3.00 | 3.21 | 3.23 | 3.14 | 2.95 | 2.83 |
| CT | FM5 | FM10 | PI10 | PI20 | |
|---|---|---|---|---|---|
| Initial body weight (g) | 21.16 ± 4.89 | 21.61 ± 4.64 | 21.23 ± 4.90 | 20.5 ± 4.92 | 21.1 ± 4.85 |
| Final body weight (g) | 48.45 ± 8.39 ab | 47.89 ± 8.61 ab | 46.61 ± 8.35 b | 48.01 ± 8.12 ab | 49.57 ± 7.85 a |
| Initial body length (cm) | 10.69 ± 0.74 | 10.76 ± 0.71 | 10.70 ± 0.79 | 10.59 ± 0.76 | 10.72 ± 0.73 |
| Final body length (cm) | 13.71 ± 0.85 ab | 13.80 ± 0.88 ab | 13.62 ± 0.88 b | 13.72 ± 0.86 ab | 13.99 ± 0.82 a |
| Fulton’s condition factor | 1.87 ± 0.13 | 1.81 ± 0.14 | 1.83 ± 0.13 | 1.85 ± 0.14 | 1.80 ± 0.11 |
| Feed conversion ratio (FCR) | 1.11± 0.21 | 1.08 ± 0.11 | 1.11 ± 0.13 | 1.06 ± 0.19 | 1.02 ± 0.11 |
| Hepatosomatic index (HSI, %) | 1.39 ± 0.26 | 1.20 ± 0.40 | 1.21 ± 0.23 | 1.12 ± 0.24 | 1.23 ± 0.28 |
| Perivisceral fat index (VFI, %) | 3.90 ± 0.61 ab | 3.30 ± 1.24 b | 4.85 ± 0.74 a | 3.71 ± 0.94 ab | 3.41 ± 0.75 b |
| Dietary Treatments | |||||
|---|---|---|---|---|---|
| CT | FM5 | FM10 | PI10 | PI20 | |
| Total lipids % (dw) | 54.94 ± 0.36 a | 45.28 ± 0.46 c | 52.03 ± 0.47 b | 36.32 ± 0.35 d | 47.07 ± 0.57 c |
| Lipid class (%TL) | |||||
| Total phospholipids | 27.74 ± 1.47 | 25.79 ± 2.01 | 25.53 ± 1.13 | 27.59 ± 0.92 | 25.87 ± 2.48 |
| FFA | 9.44 ± 0.18 ab | 10.05 ± 0.92 a | 8.79 ± 0.51 b | 9.18 ± 0.85 ab | 9.81 ± 0.18 ab |
| DAG | 5.24 ± 0.32 | 4.77 ± 0.77 | 4.78 ± 0.56 | 4.90 ± 0.51 | 5.04 ± 0.54 |
| CHO | 7.00 ± 0.60 ab | 7.40 ± 0.67 a | 6.28 ± 0.47 b | 6.81 ± 0.57 ab | 6.62 ± 0.39 ab |
| TAG | 42.43 ± 1.84 b | 46.86 ± 2.03 a | 46.61 ± 1.12 a | 43.34 ± 0.80 b | 44.65 ± 1.46 ab |
| SE | 1.61 ± 0.41 b | 1.87 ± 0.11 b | 3.30 ± 0.66 ab | 3.47 ± 0.65 a | 3.98 ± 0.14 a |
| Fatty acids % (TFA) | |||||
| 14:0 | 1.18 ± 0.12 c | 1.26 ± 0.16 bc | 1.32 ± 0.20 ab | 1.46 ± 0.18 ab | 1.55 ± 0.07 a |
| 16:0. PA | 18.62 ± 1.44 b | 18.44 ± 1.31 b | 20.80 ± 2.05 a | 20.15 ± 1.61 a | 18.31 ± 1.89 b |
| 18:0 | 6.44 ± 0.53 a | 5.98 ± 0.65 ab | 6.99 ± 0.67 a | 6.06 ± 1.11 a | 5.02 ± 0.40 b |
| Total saturated FA | 29.21 ± 1.40 b | 28.63 ± 1.14 b | 32.39 ± 2.05 a | 30.46 ± 2.39 a | 27.87 ± 1.92 b |
| 16:1n-9 | 0.55 ± 0.06 b | 0.62 ± 0.06 b | 0.74 ± 0.14 b | 0.80 ± 0.08 a | 0.90 ± 0.07 a |
| 16:1n-7 | 3.01 ± 0.29 | 3.15 ± 0.33 | 3.46 ± 0.59 | 3.5 ± 0.23 | 3.70 ± 0.28 |
| 18:1n-9. OA | 32.49 ± 3.42 | 33.18 ± 5.71 | 35.43 ± 3.13 | 37.46 ± 4.20 | 38.21 ± 1.96 |
| 18:1n-7 | 0.07 ± 0.03 | 0.07 ± 0.03 | 0.04 ± 0.04 | 0.07 ± 0.00 | 0.06 ± 0.03 |
| 22:1n-11 | 0.08 ± 0.04 a | 0.09 ± 0.05 a | 0.05 ± 0.05 b | 0.09 ± 0.01a | 0.11 ± 0.02 a |
| Total monounsaturated FA | 36.89 ± 3.84 b | 37.76 ± 6.87 b | 40.52 ± 3.57 a | 42.80 ± 5.65 a | 43.77 ± 3.98 a |
| 18:2n-6. LA | 15.47 ± 2.12 a | 15.26 ± 3.04 a | 10.17 ± 1.48 b | 11.16 ± 2.66 b | 10.18 ± 1.78 b |
| 18:3n-6 | 0.77 ± 0.19 b | 0.73 ± 0.24 b | 1.33 ± 0.66 a | 0.85 ± 0.08 b | 0.65 ± 0.11 b |
| 20:2n-6 | 0.51 ± 0.06 a | 0.49 ± 0.09 a | 0.33 ± 0.03 b | 0.40 ± 0.06 b | 0.38 ± 0.06 b |
| 20:3n-6 | 0.97 ± 0.20 b | 0.84 ± 0.30 b | 1.40 ± 0.89 a | 0.78 ± 0.10 b | 0.88 ± 0.12 b |
| 20:4n6. ARA | 0.85 ± 0.03 b | 0.80 ± 0.02 b | 0.93 ± 0.05 a | 0.80 ± 0.00 b | 0.97 ± 0.07 a |
| 22:4n-6 | 0.33 ± 0.05 | 0.47 ± 0.07 | 0.32 ± 0.05 | 0.33 ± 0.08 | 0.41 ± 0.06 |
| 22:5n-6 | 1.11 ± 0.24 | 1.01 ± 0.27 | 1.29 ± 0.66 | 0.85 ± 0.16 | 1.06 ± 0.15 |
| Total n-6 polyunsaturated FA | 19.15 ± 2.76 a | 18.65 ± 3.82 a | 14.64 ± 2.19 b | 14.75 ± 3.07 b | 13.68 ± 2.10 c |
| 18:3n-3. LNA | 1.39 ± 0.15 a | 1.33 ± 0.30 a | 0.73 ± 0.17 b | 0.85 ± 0.26 b | 0.65 ± 0.15 b |
| 20:3n-3 | 0.10 ± 0.01 ab | 0.11 ± 0.03 a | 0.04 ± 0.04 b | 0.09 ± 0.03 ab | 0.12 ± 0.03 a |
| 20:4n-3 | 1.91 ± 0.12 ab | 1.94 ± 0.37 ab | 1.44 ± 0.29 b | 1.73 ± 0.32 ab | 2.15 ± 0.32 a |
| 20:5n-3. EPA | 0.97 ± 0.30 b | 0.75 ± 0.34 b | 1.55 ± 1.10 a | 0.65 ± 0.11 c | 0.74 ± 0.12 bc |
| 22:5n-3 | 0.11 ± 0.01 | 0.08 ± 0.01 | 0.06 ± 0.02 | 0.10 ± 0.01 | 0.09 ± 0.01 |
| 22:6n-3. DHA | 6.19 ± 1.29 ab | 6.59 ± 1.69 a | 4.98 ± 1.28 b | 5.44 ± 1.11 ab | 6.94 ± 1.11 a |
| Total n-3 polyunsaturated FA | 10.17 ± 1.82 | 10.27 ± 2.51 | 9.33 ± 1.77 | 9.34 ± 1.94 | 10.92 ± 1.58 |
| n-3PUFA/n-6PUFA | 0.60 ± 0.04 c | 0.62 ± 0.04 bc | 0.69 ± 0.02 b | 0.66 ± 0.02 b | 0.85 ± 0.02 a |
| Dietary Treatment | |||||
|---|---|---|---|---|---|
| CT | FM5 | FM10 | PI10 | PI20 | |
| Total lipids % | 13.02 ± 0.50 | 12.08 ± 2.34 | 13 ± 2.44 | 12.76 ± 1.80 | 12.97 ± 1.67 |
| Lipid classes (%TL) | |||||
| Total phospholipids | 24.78 ± 1.71 b | 32.46 ± 0.36 a | 32.80 ± 0.99 a | 31.98 ± 4.34 a | 29.42 ± 3.89 ab |
| FFA | 11.12 ± 1.85 b | 12.58 ± 0.73 ab | 12.78 ± 1.19 ab | 14.06 ± 0.46 a | 14.41 ± 0.32 a |
| DAG | 3.39 ± 0.22 c | 3.64 ± 0.29 ab | 3.57 ± 0.25 b | 3.71 ± 0.32 a | 3.83 ± 0.32 a |
| CHO | 10.03 ± 0.05 | 9.78 ± 0.08 | 9.85 ± 0.01 | 9.88 ± 0.09 | 10.29 ± 0.12 |
| TAG | 43.04 ± 2.52 a | 36.46 ± 3.85 b | 36.53 ± 1.30 b | 34.63 ± 2.66 b | 35.88 ± 3.67 b |
| SE | 3.96 ± 0.86 a | 2.47 ± 1.01 b | 1.26 ± 0.45 c | 2.38 ± 0.37 b | 0.71 ± 0.32 c |
| Fatty acids % (TFA) | |||||
| 14:0 | 1.68 ± 0.06 b | 1.65 ± 0.07 b | 1.83 ± 0.09 b | 1.93 ± 0.06 a | 2.01 ± 0.26 a |
| 16:0. PA | 16.49 ± 0.41 b | 16.71 ± 0.47 b | 17.65 ± 0.54 ab | 17.17 ± 0.66 ab | 17.95 ± 1.17 a |
| 18:0 | 4.33 ± 0.20 | 4.36 ± 0.21 | 4.24 ± 0.21 | 4.79 ± 0.49 | 4.19 ± 0.23 |
| Total saturated FA | 23.85 ± 0.42 b | 24.16 ± 0.57 ab | 24.97 ± 0.74 ab | 24.41 ± 1.22 ab | 25.43 ± 1.21 a |
| 16:1n-7 | 3.70 ± 0.26 | 3.29 ± 0.17 | 3.61 ± 0.24 | 3.59 ± 0.29 | 3.81 ± 0.50 |
| 18:1n-9. OA | 23.58 ± 1.26 b | 24.54 ± 0.83 b | 28.92 ± 1.23 a | 27.39 ± 2.53 a | 27.98 ± 3.06 a |
| 18:1n-7 | 3.26 ± 0.38 | 3.16 ± 0.22 | 2.96 ± 0.58 | 3.59 ± 0.29 | 3.74 ± 0.52 |
| 22:1n-11 | 0.74 ± 0.11 a | 0.71 ± 0.09 a | 0.59 ± 0.05 b | 0.69 ± 0.07 a | 0.87 ± 0.08 a |
| Total monounsaturated FA | 33.89 ± 1.85 b | 34.27 ± 1.16 b | 38.79 ± 1.73 a | 37.92 ± 3.08 a | 39.23 ± 3.42 a |
| 18:2n-6. LA | 21.74 ± 1.52 a | 20.10 ± 1.02 a | 17.13 ± 1.82 b | 17.18 ± 1.07 b | 15.29 ± 0.38 c |
| 18:3n-6 | 0.20 ± 0.03 a | 0.18 ± 0.02 ab | 0.16 ± 0.02 b | 0.17 ± 0.02 b | 0.13 ± 0.00 c |
| 20:2n-6 | 0.63 ± 0.05 a | 0.58 ± 0.06 ab | 0.53 ± 0.07 b | 0.58 ± 0.04 ab | 0.52 ± 0.04 b |
| 20:3n-6 | 0.10 ± 0.01 | 0.11 ± 0.03 | 0.09 ± 0.00 | 0.11 ± 0.01 | 0.10 ± 0.01 |
| 20:4n6. ARA | 0.71 ± 0.02 | 0.82 ± 0.09 | 0.76 ± 0.02 | 0.78 ± 0.18 | 0.79 ± 0.16 |
| 22:4n-6 | 0.47 ± 0.03 | 0.52 ± 0.03 | 0.48 ± 0.06 | 0.51 ± 0.08 | 0.49 ± 0.05 |
| Total n-6 polyunsaturated FA | 23.76 ± 1.39 a | 22.32 ± 0.95 a | 19.16 ± 1.69 b | 19.33 ± 0.97 b | 15.84 ± 0.45 c |
| 18:3n-3. LNA | 2.56 ± 0.14 a | 2.26 ± 0.13 a | 1.89 ± 0.35 b | 1.88 ± 0.13 b | 1.61 ± 0.17 c |
| 20:3n-3 | 0.10 ± 0.01 | 0.11 ± 0.00 | 0.10 ± 0.00 | 0.10 ± 0.00 | 0.10 ± 0.00 |
| 20:4n-3 | 0.27 ± 0.01 | 0.26 ± 0.01 | 0.25 ± 0.01 | 0.25 ± 0.01 | 0.26 ± 0.02 |
| 20:5n-3. EPA | 3.50 ± 0.22 | 3.35 ± 0.21 | 3.22 ± 0.25 | 3.27 ± 0.15 | 3.25 ± 0.10 |
| 22:5n-3 | 0.79 ± 0.02 | 0.81 ± 0.04 | 0.73 ± 0.05 | 0.69 ± 0.06 | 0.77 ± 0.03 |
| 22:6n-3. DHA | 8.69 ± 0.70 ab | 9.89 ± 0.95 a | 8.74 ± 0.95 ab | 9.50 ± 0.78 a | 8.55 ± 0.68 b |
| Total n-3 polyunsaturated FA | 16.14 ± 0.75 | 16.94 ± 1.08 | 15.13 ± 1.35 | 15.92 ± 1.99 | 14.93 ± 1.20 |
| n-3PUFA/n-6PUFA | 0.67 ± 0.04 c | 0.76 ± 0.04 b | 0.70 ± 0.08 c | 0.82 ± 0.04 b | 0.94 ± 0.05 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flores-Moreno, S.; Alarcón-López, F.J.; Coronel-Domínguez, A.J.; Zuasti, E.; Hachero-Cruzado, I. A Comparative Study of the Effect of Including Full-Fat Tenebrio molitor for Replacing Conventional Ingredients in Practical Diets for Dicentrarchus labrax Juveniles. Animals 2025, 15, 131. https://doi.org/10.3390/ani15020131
Flores-Moreno S, Alarcón-López FJ, Coronel-Domínguez AJ, Zuasti E, Hachero-Cruzado I. A Comparative Study of the Effect of Including Full-Fat Tenebrio molitor for Replacing Conventional Ingredients in Practical Diets for Dicentrarchus labrax Juveniles. Animals. 2025; 15(2):131. https://doi.org/10.3390/ani15020131
Chicago/Turabian StyleFlores-Moreno, Sara, Francisco Javier Alarcón-López, Antonio J. Coronel-Domínguez, Eugenia Zuasti, and Ismael Hachero-Cruzado. 2025. "A Comparative Study of the Effect of Including Full-Fat Tenebrio molitor for Replacing Conventional Ingredients in Practical Diets for Dicentrarchus labrax Juveniles" Animals 15, no. 2: 131. https://doi.org/10.3390/ani15020131
APA StyleFlores-Moreno, S., Alarcón-López, F. J., Coronel-Domínguez, A. J., Zuasti, E., & Hachero-Cruzado, I. (2025). A Comparative Study of the Effect of Including Full-Fat Tenebrio molitor for Replacing Conventional Ingredients in Practical Diets for Dicentrarchus labrax Juveniles. Animals, 15(2), 131. https://doi.org/10.3390/ani15020131