
Mitochondrial Regulation of Spermatozoa Function: Metabolism, Oxidative Stress and Therapeutic Insights
Simple Summary
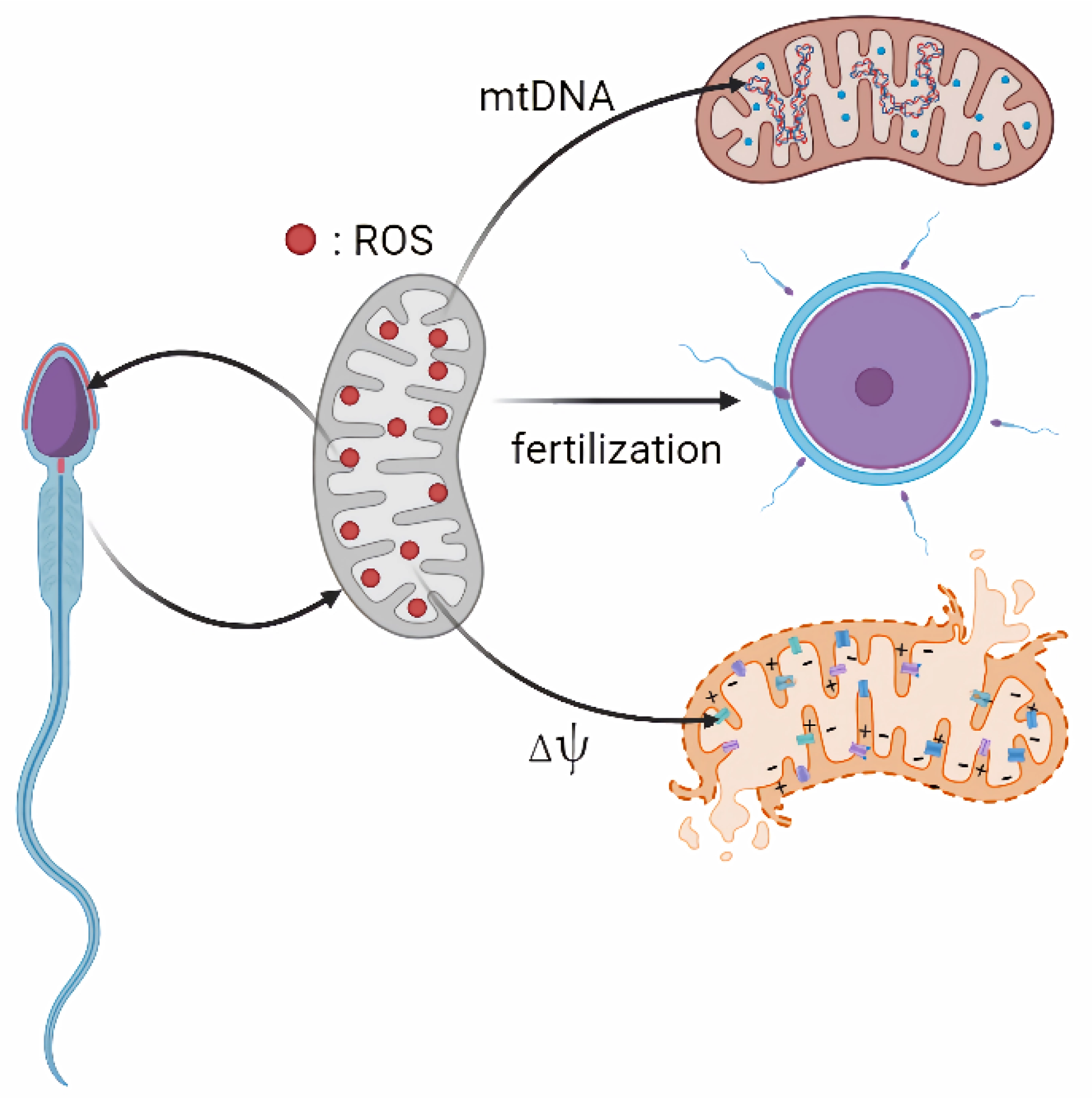
Abstract

1. Introduction
2. Relevance of Mitochondrial Energy Metabolism in Spermatozoa
2.1. The Mitochondrial Energy Metabolism in Spermatozoa
2.2. Proteins Affecting Mitochondrial Energy Metabolism in Spermatozoa
3. Effects of Mitochondrial ROS on Spermatozoa
3.1. Sources of Mitochondrial ROS in Spermatozoa
3.2. Consequences of Mitochondrial ROS Overproduction and Redox Imbalance on Sperm Quality
4. The Importance of mtDNA Stability in Mitochondrial Function Maintenance
4.1. The Characteristics of Sperm mtDNA
4.2. Effects of mtDNA Mutation on Sperm Function
5. Mitochondria-Targeted Antioxidant Therapeutic Approaches That Improve Sperm Quality
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eisenberg, M.L.; Esteves, S.C.; Lamb, D.J.; Hotaling, J.M.; Giwercman, A.; Hwang, K.; Cheng, Y.; Faculty, O.M.; Medicinska, F.; Strategiska, F.S.; et al. Male Infertility. Nature reviews. Dis. Primers 2023, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Michos, I.; Tsantarliotou, M.; Boscos, C.M.; Tsousis, G.; Basioura, A.; Tzika, E.D.; Tassis, P.D.; Lymberopoulos, A.G.; Tsakmakidis, I.A. Effect of Boar Sperm Proteins and Quality Changes on Field Fertility. Animals 2021, 11, 1813. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, M.J.; Minton, A.M.; Schwab, C.; Herickhoff, L.A. Effects of Boar Sperm Antioxidant Supplementation on Fertility. Anim. Reprod. Sci. 2022, 237, 106923. [Google Scholar] [CrossRef]
- Narud, B.; Klinkenberg, G.; Khezri, A.; Zeremichael, T.T.; Stenseth, E.B.; Nordborg, A.; Haukaas, T.H.; Morrell, J.M.; Heringstad, B.; Myromslien, F.D.; et al. Differences in Sperm Functionality and Intracellular Metabolites in Norwegian Red Bulls of Contrasting Fertility. Theriogenology 2020, 157, 24–32. [Google Scholar] [CrossRef]
- Simonik, O.; Bryndova, B.; Sur, V.P.; Ded, L.; Cockova, Z.; Benda, A.; Qasemi, M.; Pecina, P.; Pecinova, A.; Spevakova, D.; et al. Bioenergetics of Human Spermatozoa in Patients with Testicular Germ Cell Tumours. Mol. Hum. Reprod. 2025, 31, gaaf005. [Google Scholar] [CrossRef]
- Mai, Z.; Yang, D.; Wang, D.; Zhang, J.; Zhou, Q.; Han, B.; Sun, Z. A Narrative Review of Mitochondrial Dysfunction and Male Infertility. Transl. Androl. Urol. 2024, 13, 2134–2145. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, H.; Yan, H.; Han, P.; Zhang, J.; Liu, Y. Deciphering the Role of Oxidative Stress in Male Infertility: Insights from Reactive Oxygen Species to Antioxidant Therapeutics. Front. Biosci. 2025, 30, 27046. [Google Scholar] [CrossRef]
- Dhillon, V.S.; Shahid, M.; Husain, S.A. Associations of Mthfr Dnmt3B 4977 Bp Deletion in Mtdna and Gstm1 Deletion, and Aberrant Cpg Island Hypermethylation of Gstm1 in Non-Obstructive Infertility in Indian Men. Mol. Hum. Reprod. 2007, 13, 213–222. [Google Scholar] [CrossRef]
- Ferramosca, A.; Zara, V. Bioenergetics of Mammalian Sperm Capacitation. Biomed Res. Int. 2014, 2014, 902953. [Google Scholar] [CrossRef]
- Shi, L.Z.; Nascimento, J.; Botvinick, E.; Durrant, B.; Berns, M.W. An Interdisciplinary Systems Approach to Study Sperm Physiology and Evolution. Wiley Interdiscip. Rev. Syst. Biol. Med. 2011, 3, 36–47. [Google Scholar] [CrossRef] [PubMed]
- du Plessis, S.S.; Agarwal, A.; Mohanty, G.; van der Linde, M. Oxidative Phosphorylation Versus Glycolysis: What Fuel Do Spermatozoa Use? Asian J. Androl. 2015, 17, 230–235. [Google Scholar] [CrossRef]
- Hereng, T.H.; Elgstoen, K.B.; Cederkvist, F.H.; Eide, L.; Jahnsen, T.; Skalhegg, B.S.; Rosendal, K.R. Exogenous Pyruvate Accelerates Glycolysis and Promotes Capacitation in Human Spermatozoa. Hum. Reprod. 2011, 26, 3249–3263. [Google Scholar] [CrossRef]
- Sanchez-Guevara, Y.; Oliver, E.I.; Nishigaki, T. Ca2+ Concentrations in Mouse Sperm Mitochondria Fluctuate According to the Cytosol. Reproduction 2023, 167, REP-23-0237. [Google Scholar] [CrossRef]
- Mukai, C.; Okuno, M. Glycolysis Plays a Major Role for Adenosine Triphosphate Supplementation in Mouse Sperm Flagellar Movement. Biol. Reprod. 2004, 71, 540–547. [Google Scholar] [CrossRef]
- Takei, G.L.; Miyashiro, D.; Mukai, C.; Okuno, M. Glycolysis Plays an Important Role in Energy Transfer from the Base to the Distal End of the Flagellum in Mouse Sperm. J. Exp. Biol. 2014, 217 Pt 11, 1876–1886. [Google Scholar] [CrossRef]
- Piomboni, P.; Focarelli, R.; Stendardi, A.; Ferramosca, A.; Zara, V. The Role of Mitochondria in Energy Production for Human Sperm Motility. Int. J. Androl. 2012, 35, 109–124. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, J.M.; Shi, L.Z.; Tam, J.; Chandsawangbhuwana, C.; Durrant, B.; Botvinick, E.L.; Berns, M.W. Comparison of Glycolysis and Oxidative Phosphorylation as Energy Sources for Mammalian Sperm Motility, Using the Combination of Fluorescence Imaging, Laser Tweezers, and Real-Time Automated Tracking and Trapping. J. Cell. Physiol. 2008, 217, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.C.; Ford, W.C. The Role of Glucose in Supporting Motility and Capacitation in Human Spermatozoa. J. Androl. 2001, 22, 680–695. [Google Scholar] [CrossRef] [PubMed]
- Losano, J.; Angrimani, D.; Dalmazzo, A.; Rui, B.R.; Brito, M.M.; Mendes, C.M.; Kawai, G.; Vannucchi, C.I.; Assumpcao, M.; Barnabe, V.H.; et al. Effect of Mitochondrial Uncoupling and Glycolysis Inhibition on Ram Sperm Functionality. Reprod. Domest. Anim. 2017, 52, 289–297. [Google Scholar] [CrossRef]
- Wang, J.J.; Wang, S.X.; Tehmina; Feng, Y.; Zhang, R.F.; Li, X.Y.; Sun, Q.; Ding, J. Age-Related Decline of Male Fertility: Mitochondrial Dysfunction and the Antioxidant Interventions. Pharmaceuticals 2022, 15, 519. [Google Scholar] [CrossRef]
- Davila, M.P.; Muñoz, P.M.; Bolaños, J.M.G.; Stout, T.A.E.; Gadella, B.M.; Tapia, J.A.; Da Silva, C.B.; Ferrusola, C.O.; Peña, F.J. Mitochondrial Atp is Required for the Maintenance of Membrane Integrity in Stallion Spermatozoa, Whereas Motility Requires Both Glycolysis and Oxidative Phosphorylation. Reproduction 2016, 152, 683–694. [Google Scholar] [CrossRef]
- Ortiz-Rodriguez, J.M.; Bucci, D.; Tovar-Pascual, L.; Granata, S.; Spinaci, M.; Nesci, S. Analysis of Stallion Spermatozoa Metabolism Using Agilent Seahorse Xfp Technology. Anim. Reprod. Sci. 2024, 271, 107633. [Google Scholar] [CrossRef]
- Pena, F.J.; Martin-Cano, F.E.; Becerro-Rey, L.; Da, S.E.; Gaitskell-Phillips, G.; Aparicio, I.M.; Gil, M.C.; Ortega-Ferrusola, C. Redox Regulation and Glucose Metabolism in the Stallion Spermatozoa. Antioxidants 2025, 14, 225. [Google Scholar] [CrossRef]
- Plaza, D.M.; Martin, M.P.; Tapia, J.A.; Ortega, F.C.; Balao, D.S.C.C.; Pena, F.J. Inhibition of Mitochondrial Complex I Leads to Decreased Motility and Membrane Integrity Related to Increased Hydrogen Peroxide and Reduced Atp Production, while the Inhibition of Glycolysis Has Less Impact on Sperm Motility. PLoS ONE 2015, 10, e0138777. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Prieto, O.; Mislei, B.; Martinez-Pastor, F.; Spinaci, M.; Mari, G.; Bucci, D. Study of Mitochondrial Function in Thawed Bull Spermatozoa Using Selective Electron Transfer Chain Inhibitors. Theriogenology 2023, 208, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Bulkeley, E.A.; Foutouhi, A.; Wigney, K.; Santistevan, A.C.; Collins, C.; McNabb, B.; Meyers, S. Effects from Disruption of Mitochondrial Electron Transport Chain Function on Bull Sperm Motility. Theriogenology 2021, 176, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Hutson, S.M.; Van Dop, C.; Lardy, H.A. Mitochondrial Metabolism of Pyruvate in Bovine Spermatozoa. J. Biol. Chem. 1977, 252, 1309–1315. [Google Scholar] [CrossRef]
- Henning, H.; Nguyen, Q.T.; Luther, A.M.; Wallner, U.; Beyerbach, M.; Waberski, D. In Vitro Storage of Boar Spermatozoa Increases the Demand of Adenosine Triphosphate for Reactivation of Motility. Andrology 2022, 10, 1426–1440. [Google Scholar] [CrossRef]
- Nesci, S.; Spinaci, M.; Galeati, G.; Nerozzi, C.; Pagliarani, A.; Algieri, C.; Tamanini, C.; Bucci, D. Sperm Function and Mitochondrial Activity: An Insight on Boar Sperm Metabolism. Theriogenology 2020, 144, 82–88. [Google Scholar] [CrossRef]
- Prieto, O.B.; Algieri, C.; Spinaci, M.; Trombetti, F.; Nesci, S.; Bucci, D. Cell Bioenergetics and Atp Production of Boar Spermatozoa. Theriogenology 2023, 210, 162–168. [Google Scholar] [CrossRef]
- Haran, M.; Gross, A. Balancing Glycolysis and Mitochondrial Oxphos: Lessons From the Hematopoietic System and Exercising Muscles. Mitochondrion 2014, 19 Pt A, 3–7. [Google Scholar] [CrossRef]
- Castello-Ruiz, M.; Gacem, S.; Sanchez, D.P.M.; Hidalgo, C.O.; Tamargo, C.; Alvarez-Rodriguez, M.; Yaniz, J.L.; Silvestre, M.A. Effect of Capacitation on Proteomic Profile and Mitochondrial Parameters of Spermatozoa in Bulls. J. Proteome Res. 2025, 24, 1817–1831. [Google Scholar] [CrossRef]
- Nolfi-Donegan, D.; Braganza, A.; Shiva, S. Mitochondrial Electron Transport Chain: Oxidative Phosphorylation, Oxidant Production, and Methods of Measurement. Redox Biol. 2020, 37, 101674. [Google Scholar] [CrossRef]
- Vercellino, I.; Sazanov, L.A. The Assembly, Regulation and Function of the Mitochondrial Respiratory Chain. Nat. Rev. Mol. Cell Biol. 2022, 23, 141–161. [Google Scholar] [CrossRef]
- Hirst, J. Mitochondrial Complex I. Annu. Rev. Biochem. 2013, 82, 551–575. [Google Scholar] [CrossRef]
- Braun, H.P.; Klusch, N. Promotion of Oxidative Phosphorylation by Complex I-Anchored Carbonic Anhydrases? Trends Plant Sci. 2024, 29, 64–71. [Google Scholar] [CrossRef]
- Bezawork-Geleta, A.; Rohlena, J.; Dong, L.; Pacak, K.; Neuzil, J. Mitochondrial Complex II: At the Crossroads. Trends Biochem. Sci. 2017, 42, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Cecchini, G. Function and Structure of Complex II of the Respiratory Chain. Annu. Rev. Biochem. 2003, 72, 77–109. [Google Scholar] [CrossRef] [PubMed]
- Miyadera, H.; Shiomi, K.; Ui, H.; Yamaguchi, Y.; Masuma, R.; Tomoda, H.; Miyoshi, H.; Osanai, A.; Kita, K.; Omura, S. Atpenins, Potent and Specific Inhibitors of Mitochondrial Complex II (Succinate-Ubiquinone Oxidoreductase). Proc. Natl. Acad. Sci. USA 2003, 100, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.; Purhonen, J.; Kallijärvi, J. The Mitochondrial Coenzyme Q Junction and Complex III: Biochemistry and Pathophysiology. FEBS J. 2022, 289, 6936–6958. [Google Scholar] [CrossRef]
- Kadenbach, B. Complex IV—the Regulatory Center of Mitochondrial Oxidative Phosphorylation. Mitochondrion 2021, 58, 296–302. [Google Scholar] [CrossRef]
- Juhaszova, M.; Kobrinsky, E.; Zorov, D.B.; Nuss, H.B.; Yaniv, Y.; Fishbein, K.W.; de Cabo, R.; Montoliu, L.; Gabelli, S.B.; Aon, M.A.; et al. Atp Synthase K(+)- And H(+)-Fluxes Drive Atp Synthesis and Enable Mitochondrial K(+)-“Uniporter” Function: I. Characterization of Ion Fluxes. Function 2022, 3, zqab065. [Google Scholar] [CrossRef] [PubMed]
- Bucak, M.N.; Ataman, M.B.; Baspinar, N.; Uysal, O.; Taspinar, M.; Bilgili, A.; Ozturk, C.; Gungor, S.; Inanc, M.E.; Akal, E. Lycopene and Resveratrol Improve Post-Thaw Bull Sperm Parameters: Sperm Motility, Mitochondrial Activity and Dna Integrity. Andrologia 2015, 47, 545–552. [Google Scholar] [CrossRef]
- Mughal, I.A.; Irfan, A.; Hameed, A.; Jahan, S. Sperm Mitochondrial Dna 15Bp Deletion of Cytochrome C Oxidase Subunit III is Significantly Associated with Human Male Infertility in Pakistan. J. Pak. Med. Assoc. 2016, 66, 3–7. [Google Scholar]
- Capaldi, R.A.; Aggeler, R.; Turina, P.; Wilkens, S. Coupling Between Catalytic Sites and the Proton Channel in F1F0-Type Atpases. Trends Biochem. Sci. 1994, 19, 284–289. [Google Scholar] [CrossRef]
- Nijtmans, L.G.; Klement, P.; Houstek, J.; van den Bogert, C. Assembly of Mitochondrial Atp Synthase in Cultured Human Cells: Implications for Mitochondrial Diseases. Biochim. Biophys. Acta 1995, 1272, 190–198. [Google Scholar] [CrossRef]
- Ramio-Lluch, L.; Yeste, M.; Fernandez-Novell, J.M.; Estrada, E.; Rocha, L.; Cebrian-Perez, J.A.; Muino-Blanco, T.; Concha, I.I.; Ramirez, A.; Rodriguez-Gil, J.E. Oligomycin a-Induced Inhibition of Mitochondrial Atp-Synthase Activity Suppresses Boar Sperm Motility and in Vitro Capacitation Achievement without Modifying Overall Sperm Energy Levels. Reprod. Fertil. Dev. 2014, 26, 883–897. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, I.; Heaton, R.A.; Mantle, D. Disorders of Human Coenzyme Q10 Metabolism: An Overview. Int. J. Mol. Sci. 2020, 21, 6695. [Google Scholar] [CrossRef]
- Lewin, A.; Lavon, H. The Effect of Coenzyme Q10 on Sperm Motility and Function. Mol. Aspects Med. 1997, 18, 213–219. [Google Scholar] [CrossRef]
- Littarru, G.P.; Tiano, L. Bioenergetic and Antioxidant Properties of Coenzyme Q10: Recent Developments. Mol. Biotechnol. 2007, 37, 31–37. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Fang, Y.; Zhang, F.; Liu, Y.; Cheng, X.; Wang, J.; Li, D.; Chen, D.; Wu, F. Adenine Nucleotide Translocase 2 (Ant2) is Required for Individualization of Spermatogenesis of Drosophila Melanogaster. Insect Sci. 2024, 31, 1055–1072. [Google Scholar] [CrossRef]
- Liu, Y.; Li, T.; Shi, M.; Wan, Y.; Li, H.; Zhang, M.; Wang, Z.; Wang, S.; Lv, Y.; Lu, G.; et al. Morn2 Regulates the Morphology and Energy Metabolism of Mitochondria and is Required for Male Fertility in Mice. J. Transl. Med. 2024, 22, 240. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Xiao, Y.; Gu, J.; Zhao, P.; Hu, Z.; Zheng, J.; Hua, R.; Hai, Z.; Su, J.; Zhang, J.V.; et al. Clpp/Clpx Deficiency Impairs Mitochondrial Functions and Mtorc1 Signaling During Spermatogenesis. Commun. Biol. 2023, 6, 1012. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, C.; Wang, Y.; Zhang, H.; Xu, C.; Cheng, Y.; Yuan, Y.; Sha, J.; Guo, X.; Cui, Y. A Novel Protein Encoded by Circrsrc1 Regulates Mitochondrial Ribosome Assembly and Translation During Spermatogenesis. BMC Biol. 2023, 21, 94. [Google Scholar] [CrossRef] [PubMed]
- Kuang, W.; Zhang, J.; Lan, Z.; Deepak, R.; Liu, C.; Ma, Z.; Cheng, L.; Zhao, X.; Meng, X.; Wang, W.; et al. Slc22a14 is a Mitochondrial Riboflavin Transporter Required for Sperm Oxidative Phosphorylation and Male Fertility. Cell Rep. 2021, 35, 109025. [Google Scholar] [CrossRef]
- Zhu, F.; Li, W.; Zhou, X.; Chen, X.; Zheng, M.; Cui, Y.; Liu, X.; Guo, X.; Zhu, H. Prss55 Plays an Important Role in the Structural Differentiation and Energy Metabolism of Sperm and is Required for Male Fertility in Mice. J. Cell Mol. Med. 2021, 25, 2040–2051. [Google Scholar] [CrossRef]
- Ortiz-Rodriguez, J.M.; Martin-Cano, F.E.; Gaitskell-Phillips, G.; Silva, A.; Tapia, J.A.; Gil, M.C.; Redondo, E.; Masot, J.; Ortega-Ferrusola, C.; Pena, F.J. The Slc7a11: Sperm Mitochondrial Function and Non-Canonical Glutamate Metabolism. Reproduction 2020, 160, 803–818. [Google Scholar] [CrossRef]
- Sun, Y.; Sun, X.; Zhao, L.; Zhang, Z.; Wang, Y.; Dai, Z.; Zhao, X.; Pu, X. Dj-1 Deficiency Causes Metabolic Abnormality in Ornidazole-Induced Asthenozoospermia. Reproduction 2020, 160, 931–941. [Google Scholar] [CrossRef]
- Zhu, Z.; Li, R.; Wang, L.; Zheng, Y.; Hoque, S.; Lv, Y.; Zeng, W. Glycogen Synthase Kinase-3 Regulates Sperm Motility and Acrosome Reaction Via Affecting Energy Metabolism in Goats. Front. Physiol. 2019, 10, 968. [Google Scholar] [CrossRef]
- Zhu, F.; Yan, P.; Zhang, J.; Cui, Y.; Zheng, M.; Cheng, Y.; Guo, Y.; Yang, X.; Guo, X.; Zhu, H. Deficiency of Tppp2, a Factor Linked to Oligoasthenozoospermia, Causes Subfertility in Male Mice. J. Cell Mol. Med. 2019, 23, 2583–2594. [Google Scholar] [CrossRef]
- Tartarin, P.; Guibert, E.; Toure, A.; Ouiste, C.; Leclerc, J.; Sanz, N.; Briere, S.; Dacheux, J.L.; Delaleu, B.; McNeilly, J.R.; et al. Inactivation of Ampkalpha1 Induces Asthenozoospermia and Alters Spermatozoa Morphology. Endocrinology 2012, 153, 3468–3481. [Google Scholar] [CrossRef]
- Zhu, Z.; Li, R.; Ma, G.; Bai, W.; Fan, X.; Lv, Y.; Luo, J.; Zeng, W. 5′-Amp-Activated Protein Kinase Regulates Goat Sperm Functions Via Energy Metabolism in Vitro. Cell Physiol. Biochem. 2018, 47, 2420–2431. [Google Scholar] [CrossRef]
- Odet, F.; Gabel, S.A.; Williams, J.; London, R.E.; Goldberg, E.; Eddy, E.M. Lactate Dehydrogenase C and Energy Metabolism in Mouse Sperm. Biol. Reprod. 2011, 85, 556–564. [Google Scholar] [CrossRef]
- Gawlik, V.; Schmidt, S.; Scheepers, A.; Wennemuth, G.; Augustin, R.; Aumuller, G.; Moser, M.; Al-Hasani, H.; Kluge, R.; Joost, H.G.; et al. Targeted Disruption of Slc2a8 (Glut8) Reduces Motility and Mitochondrial Potential of Spermatozoa. Mol. Membr. Biol. 2008, 25, 224–235. [Google Scholar] [CrossRef]
- Koppers, A.J.; De Iuliis, G.N.; Finnie, J.M.; McLaughlin, E.A.; Aitken, R.J. Significance of Mitochondrial Reactive Oxygen Species in the Generation of Oxidative Stress in Spermatozoa. J. Clin. Endocrinol. Metab. 2008, 93, 3199–3207. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T. Signal Transduction by Reactive Oxygen Species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Cadenas, E.; Davies, K.J. Mitochondrial Free Radical Generation, Oxidative Stress, and Aging. Free Radic. Biol. Med. 2000, 29, 222–230. [Google Scholar] [CrossRef]
- Hussain, T.; Kandeel, M.; Metwally, E.; Murtaza, G.; Kalhoro, D.H.; Yin, Y.; Tan, B.; Chughtai, M.I.; Yaseen, A.; Afzal, A.; et al. Unraveling the Harmful Effect of Oxidative Stress on Male Fertility: A Mechanistic Insight. Front. Endocrinol. 2023, 14, 1070692. [Google Scholar] [CrossRef]
- Chianese, R.; Pierantoni, R. Mitochondrial Reactive Oxygen Species (Ros) Production Alters Sperm Quality. Antioxidants 2021, 10, 92. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Trocchia, S.; Abdel-Gawad, F.K.; Ciarcia, G. Roles of Reactive Oxygen Species in the Spermatogenesis Regulation. Front. Endocrinol. 2014, 5, 56. [Google Scholar] [CrossRef]
- Sikka, S.C. Relative Impact of Oxidative Stress on Male Reproductive Function. Curr. Med. Chem. 2001, 8, 851–862. [Google Scholar] [CrossRef]
- Sanocka, D.; Kurpisz, M. Reactive Oxygen Species and Sperm Cells. Reprod. Biol. Endocrinol. 2004, 2, 12. [Google Scholar] [CrossRef] [PubMed]
- Foutouhi, A.; Meyers, S. Comparative Oxidative Metabolism in Mammalian Sperm. Anim. Reprod. Sci. 2022, 247, 107095. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, C.L.; Perevoshchikova, I.V.; Hey-Mogensen, M.; Orr, A.L.; Brand, M.D. Sites of Reactive Oxygen Species Generation by Mitochondria Oxidizing Different Substrates. Redox Biol. 2013, 1, 304–312. [Google Scholar] [CrossRef]
- Kamali Sangani, A.; Masoudi, A.A.; Vaez Torshizi, R. Association of Mitochondrial Function and Sperm Progressivity in Slow- And Fast-Growing Roosters. Poult. Sci. 2017, 96, 211–219. [Google Scholar] [CrossRef]
- Zhu, Z.; Kawai, T.; Umehara, T.; Hoque, S.; Zeng, W.; Shimada, M. Negative Effects of Ros Generated During Linear Sperm Motility on Gene Expression and Atp Generation in Boar Sperm Mitochondria. Free Radic. Biol. Med. 2019, 141, 159–171. [Google Scholar] [CrossRef]
- Suen, D.F.; Norris, K.L.; Youle, R.J. Mitochondrial Dynamics and Apoptosis. Genes. Dev. 2008, 22, 1577–1590. [Google Scholar] [CrossRef]
- Ferramosca, A.; Zara, V. Diet and Male Fertility: The Impact of Nutrients and Antioxidants on Sperm Energetic Metabolism. Int. J. Mol. Sci. 2022, 23, 2542. [Google Scholar] [CrossRef] [PubMed]
- Lone, S.A.; Mohanty, T.K.; Baithalu, R.K.; Yadav, H.P. Sperm Protein Carbonylation. Andrologia 2019, 51, e13233. [Google Scholar] [CrossRef]
- Tirmarche, S.; Kimura, S.; Dubruille, R.; Horard, B.; Loppin, B. Unlocking Sperm Chromatin at Fertilization Requires a Dedicated Egg Thioredoxin in Drosophila. Nat. Commun. 2016, 7, 13539. [Google Scholar] [CrossRef]
- Montjean, D.; Ravel, C.; Benkhalifa, M.; Cohen-Bacrie, P.; Berthaut, I.; Bashamboo, A.; McElreavey, K. Methylation Changes in Mature Sperm Deoxyribonucleic Acid from Oligozoospermic Men: Assessment of Genetic Variants and Assisted Reproductive Technology Outcome. Fertil. Steril. 2013, 100, 1241–1247. [Google Scholar] [CrossRef]
- O’Flaherty, C.; Matsushita-Fournier, D. Reactive Oxygen Species and Protein Modifications in Spermatozoa. Biol. Reprod. 2017, 97, 577–585. [Google Scholar] [CrossRef]
- Fujii, J.; Tsunoda, S. Redox Regulation of Fertilisation and the Spermatogenic Process. Asian J. Androl. 2011, 13, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J. Reactive Oxygen Species as Mediators of Sperm Capacitation and Pathological Damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef]
- Aitken, R.J.; Baker, M.A.; Nixon, B. Are Sperm Capacit. Apoptosis Opposite Ends A Contin. Driven By Oxidative Stress? Asian J. Androl. 2015, 17, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Majzoub, A.; Agarwal, A. Oxidative Stress and Sperm Function: A Systematic Review on Evaluation and Management. Arab. J. Urol. 2019, 17, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Asadi, A.; Ghahremani, R.; Abdolmaleki, A.; Rajaei, F. Role of Sperm Apoptosis and Oxidative Stress in Male Infertility: A Narrative Review. Int. J. Reprod. Biomed. 2021, 19, 493–504. [Google Scholar] [CrossRef]
- Bui, A.D.; Sharma, R.; Henkel, R.; Agarwal, A. Reactive Oxygen Species Impact on Sperm Dna and its Role in Male Infertility. Andrologia 2018, 50, e13012. [Google Scholar] [CrossRef]
- Liu, Y.; Su, Z.; Tavana, O.; Gu, W. Understanding the Complexity of P53 in a New Era of Tumor Suppression. Cancer Cell 2024, 42, 946–967. [Google Scholar] [CrossRef]
- Amaral, A.; Lourenco, B.; Marques, M.; Ramalho-Santos, J. Mitochondria Functionality and Sperm Quality. Reproduction 2013, 146, R163–R174. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-Hydroxy-2′ -Deoxyguanosine (8-Ohdg): A Critical Biomarker of Oxidative Stress and Carcinogenesis. J. Envron. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef]
- Moor, N.A.; Vasil’Eva, I.A.; Kuznetsov, N.A.; Lavrik, O.I. Human Apurinic/Apyrimidinic Endonuclease 1 is Modified in Vitro by Poly(Adp-Ribose) Polymerase 1 Under Control of the Structure of Damaged Dna. Biochimie 2020, 168, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Moazamian, A.; Gharagozloo, P.; Aitken, R.J.; Drevet, J.R. Oxidative Stress and Reproductive Function: Sperm Telomeres, Oxidative Stress, and Infertility. Reproduction 2022, 164, F125–F133. [Google Scholar] [CrossRef]
- Stuppia, L.; Franzago, M.; Ballerini, P.; Gatta, V.; Antonucci, I. Epigenetics and Male Reproduction: The Consequences of Paternal Lifestyle on Fertility, Embryo Development, and Children Lifetime Health. Clin. Epigenetics 2015, 7, 120. [Google Scholar] [CrossRef]
- Bonetti, D.; Martina, M.; Falcettoni, M.; Longhese, M.P. Telomere-End Processing: Mechanisms and Regulation. Chromosoma 2014, 123, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Kumar, R.; Malhotra, N.; Singh, N.; Dada, R. Mild Oxidative Stress is Beneficial for Sperm Telomere Length Maintenance. World J. Methodol. 2016, 6, 163–170. [Google Scholar] [CrossRef]
- Aitken, R.J. Human Spermatozoa: Revelations on the Road to Conception. F1000Prime Rep. 2013, 5, 39. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.B.; Chawla, B.; Bisht, S.; Yadav, R.K.; Dada, R. Tobacco Use Increases Oxidative Dna Damage in Sperm—Possible Etiology of Childhood Cancer. Asian Pac. J. Cancer Prev. 2015, 16, 6967–6972. [Google Scholar] [CrossRef]
- Lee, W.; Zamudio-Ochoa, A.; Buchel, G.; Podlesniy, P.; Marti, G.N.; Puigros, M.; Calderon, A.; Tang, H.Y.; Li, L.; Mikhalchenko, A.; et al. Molecular Basis for Maternal Inheritance of Human Mitochondrial Dna. Nat. Genet. 2023, 55, 1632–1639. [Google Scholar] [CrossRef]
- Nishimura, Y.; Yoshinari, T.; Naruse, K.; Yamada, T.; Sumi, K.; Mitani, H.; Higashiyama, T.; Kuroiwa, T. Active Digestion of Sperm Mitochondrial Dna in Single Living Sperm Revealed by Optical Tweezers. Proc. Natl. Acad. Sci. USA 2006, 103, 1382–1387. [Google Scholar] [CrossRef]
- Yan, C.; Duanmu, X.; Zeng, L.; Liu, B.; Song, Z. Mitochondrial Dna: Distribution, Mutations, and Elimination. Cells 2019, 8, 379. [Google Scholar] [CrossRef] [PubMed]
- Durairajanayagam, D.; Singh, D.; Agarwal, A.; Henkel, R. Causes and Consequences of Sperm Mitochondrial Dysfunction. Andrologia 2021, 53, e13666. [Google Scholar] [CrossRef]
- Vertika, S.; Singh, K.K.; Rajender, S. Mitochondria, Spermatogenesis, and Male Infertility—An Update. Mitochondrion 2020, 54, 26–40. [Google Scholar] [CrossRef]
- Shamsi, M.B.; Kumar, R.; Bhatt, A.; Bamezai, R.N.; Kumar, R.; Gupta, N.P.; Das, T.K.; Dada, R. Mitochondrial Dna Mutations in Etiopathogenesis of Male Infertility. Indian. J. Urol. 2008, 24, 150–154. [Google Scholar] [CrossRef]
- Amor, H.; Hammadeh, M.E. A Systematic Review of the Impact of Mitochondrial Variations on Male Infertility. Genes 2022, 13, 1182. [Google Scholar] [CrossRef] [PubMed]
- Copeland, W.C. Defects of Mitochondrial Dna Replication. J. Child. Neurol. 2014, 29, 1216–1224. [Google Scholar] [CrossRef]
- Holyoake, A.J.; McHugh, P.; Wu, M.; O’Carroll, S.; Benny, P.; Sin, I.L.; Sin, F.Y.T. High Incidence of Single Nucleotide Substitutions in the Mitochondrial Genome is Associated with Poor Semen Parameters in Men. Int. J. Androl. 2001, 24, 175–182. [Google Scholar] [CrossRef]
- Spiropoulos, J.; Turnbull, D.M.; Chinnery, P.F. Can Mitochondrial Dna Mutations Cause Sperm Dysfunction? Mol. Hum. Reprod. 2002, 8, 719–721. [Google Scholar] [CrossRef] [PubMed]
- Saleh Jaweesh, M.; Hammadeh, M.E.; Dahadhah, F.W.; Al Zoubi, M.S.; Amor, H. Association Between the Single Nucleotide Variants of the Mitochondrial Cytochrome B Gene (Mt-Cyb) and the Male Infertility. Mol. Biol. Rep. 2022, 49, 3609–3616. [Google Scholar] [CrossRef] [PubMed]
- Kao, S.; Chao, H.T.; Wei, Y.H. Mitochondrial Deoxyribonucleic Acid 4977-Bp Deletion is Associated with Diminished Fertility and Motility of Human Sperm. Biol. Reprod. 1995, 52, 729–736. [Google Scholar] [CrossRef]
- Bahrehmand, N.I.; Vaziri, H. Sperm Mitochondrial Dna Deletion in Iranian Infertiles with Asthenozoospermia. Andrologia 2017, 49, e12627. [Google Scholar] [CrossRef]
- Karimian, M.; Babaei, F. Large-Scale Mtdna Deletions as Genetic Biomarkers for Susceptibility to Male Infertility: A Systematic Review and Meta-Analysis. Int. J. Biol. Macromol. 2020, 158, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Talebi, E.; Karimian, M.; Nikzad, H. Association of Sperm Mitochondrial Dna Deletions with Male Infertility in an Iranian Population. Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2018, 29, 615–623. [Google Scholar] [CrossRef]
- Ieremiadou, F.; Rodakis, G.C. Correlation of the 4977 Bp Mitochondrial Dna Deletion with Human Sperm Dysfunction. BMC Res. Notes 2009, 2, 18. [Google Scholar] [CrossRef]
- Mirabutalebi, S.H.; Karami, N.; Ashrafzadeh, H.R.; Akhvansales, Z.; Tavakoli, M.; Ghasemi, N. Detection of 4977-Bp Deletion of Mitochondrial Dna in in Vitro Fertilization Failure Women: A Case-Control Study. Int. J. Reprod. Biomed. 2018, 16, 571–576. [Google Scholar] [CrossRef]
- Gashti, N.G.; Salehi, Z.; Madani, A.H.; Dalivandan, S.T. 4977-Bp Mitochondrial Dna Deletion in Infertile Patients with Varicocele. Andrologia 2014, 46, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Schapira, A.H. Primary and Secondary Defects of the Mitochondrial Respiratory Chain. J. Inherit. Metab. Dis. 2002, 25, 207–214. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Do, S.Q.; Wakai, T.; Funahashi, H. Mitochondrial Content and Mtdna Copy Number in Spermatozoa and Penetrability into Oocytes. Theriogenology 2025, 234, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Huffman, A.M.; Whitcomb, B.W.; Josyula, S.; Labrie, S.; Tougias, E.; Rahil, T.; Sites, C.K.; Pilsner, J.R. Sperm Mitochondrial Dna Measures and Semen Parameters Among Men Undergoing Fertility Treatment. Reprod. Biomed. Online 2019, 38, 66–75. [Google Scholar] [CrossRef]
- Teixeira, J.; Deus, C.M.; Borges, F.; Oliveira, P.J. Mitochondria: Targeting Mitochondrial Reactive Oxygen Species with Mitochondriotropic Polyphenolic-Based Antioxidants. Int. J. Biochem. Cell Biol. 2018, 97, 98–103. [Google Scholar] [CrossRef]
- Wang, J.Y.; Li, J.Q.; Xiao, Y.M.; Fu, B.; Qin, Z.H. Triphenylphosphonium (Tpp)-Based Antioxidants: A New Perspective on Antioxidant Design. ChemMedChem 2020, 15, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Escada-Rebelo, S.; Cristo, M.I.; Ramalho-Santos, J.; Amaral, S. Mitochondria-Targeted Compounds to Assess and Improve Human Sperm Function. Antioxid. Redox Signal. 2022, 37, 451–480. [Google Scholar] [CrossRef]
- Xiao, L.; Xu, X.; Zhang, F.; Wang, M.; Xu, Y.; Tang, D.; Wang, J.; Qin, Y.; Liu, Y.; Tang, C.; et al. The Mitochondria-Targeted Antioxidant Mitoq Ameliorated Tubular Injury Mediated by Mitophagy in Diabetic Kidney Disease Via Nrf2/Pink1. Redox Biol. 2017, 11, 297–311. [Google Scholar] [CrossRef]
- Alevra, A.I.; Exadactylos, A.; Mente, E.; Papadopoulos, S. The Protective Role of Melatonin in Sperm Cryopreservation of Farm Animals and Human: Lessons for Male Fish Cryopreservation. Animals 2022, 12, 791. [Google Scholar] [CrossRef]
- Moradi Gardeshi, T.; Shahandeh, E.; Tavakolpoor Saleh, N.; Karami, S.; Mirzaei Azandaryani, Z.; Mazaheri, F.; Mohammadi, H. Evaluation of the Effect of Mitoquinone on Functional Parameters, Dna Structure, and Genes Expression Related to the Apoptotic and Antioxidants of Human Sperm After Freezing–Thawing. Mol. Biol. Rep. 2024, 51, 183. [Google Scholar] [CrossRef]
- Tiwari, S.; Mohanty, T.K.; Bhakat, M.; Kumar, N.; Baithalu, R.K.; Nath, S.; Yadav, H.P.; Dewry, R.K. Comparative Evidence Support Better Antioxidant Efficacy of Mitochondrial-Targeted (Mitoquinone) than Cytosolic (Resveratrol) Antioxidant in Improving in-Vitro Sperm Functions of Cryopreserved Buffalo (Bubalus bubalis) Semen. Cryobiology 2021, 101, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Bai, C.; Chen, Y.; Dai, J.; Xiang, Y.; Ji, X.; Huang, C.; Dong, Q. Inhibition of Ros Production through Mitochondria-Targeted Antioxidant and Mitochondrial Uncoupling Increases Post-Thaw Sperm Viability in Yellow Catfish. Cryobiology 2014, 69, 386–693. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.M.; Dong, X.; Xue, X.D.; Xu, S.; Zhang, X.; Xu, Y.L.; Wang, Z.S.; Wang, Y.; Gao, H.; Liang, Y.X.; et al. Melatonin Attenuates Diabetic Cardiomyopathy and Reduces Myocardial Vulnerability to Ischemia-Reperfusion Injury by Improving Mitochondrial Quality Control: Role of Sirt6. J. Pineal Res. 2021, 70, e12698. [Google Scholar] [CrossRef]
- Govender, J.; Loos, B.; Marais, E.; Engelbrecht, A.M. Mitochondrial Catastrophe During Doxorubicin-Induced Cardiotoxicity: A Review of the Protective Role of Melatonin. J. Pineal Res. 2014, 57, 367–680. [Google Scholar] [CrossRef]
- Acuna-Castroviejo, D.; Escames, G.; Rodriguez, M.I.; Lopez, L.C. Melatonin Role in the Mitochondrial Function. Front Biosci. 2007, 12, 947–963. [Google Scholar] [CrossRef]
- Fang, Y.; Zhao, C.; Xiang, H.; Zhao, X.; Zhong, R. Melatonin Inhibits Formation of Mitochondrial Permeability Transition Pores and Improves Oxidative Phosphorylation of Frozen-Thawed Ram Sperm. Front. Endocrinol. 2019, 10, 896. [Google Scholar] [CrossRef]
- Zhang, X.; Xiong, Y.; Tan, Y.; Wang, L.; Li, R.; Zhang, Y.; Liu, X.; Lin, X.; Jin, L.; Hu, Y.; et al. Melatonin Rescues Impaired Penetration Ability of Human Spermatozoa Induced by Mitochondrial Dysfunction. Reproduction 2019, 158, 465–475. [Google Scholar] [CrossRef]
- Niu, F.W.; Liu, M.D.; Yao, K.; Yang, R.; Gao, L.; Zhai, J.X.; Wang, C.; Zhang, S.H.; Xu, D.X.; Zhang, Z.H. Mitochondrial Ros-Associated Integrated Stress Response is Involved in Arsenic-Induced Blood-Testis Barrier Disruption and Protective Effect of Melatonin. Environ. Int. 2025, 197, 109346. [Google Scholar] [CrossRef]
- Deng, S.L.; Sun, T.C.; Yu, K.; Wang, Z.P.; Zhang, B.L.; Zhang, Y.; Wang, X.X.; Lian, Z.X.; Liu, Y.X. Melatonin Reduces Oxidative Damage and Upregulates Heat Shock Protein 90 Expression in Cryopreserved Human Semen. Free Radic. Biol. Med. 2017, 113, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Karimfar, M.H.; Niazvand, F.; Haghani, K.; Ghafourian, S.; Shirazi, R.; Bakhtiyari, S. The Protective Effects of Melatonin Against Cryopreservation-Induced Oxidative Stress in Human Sperm. Int. J. Immunopathol. Pharmacol. 2015, 28, 69–76. [Google Scholar] [CrossRef]
- Najafi, A.; Adutwum, E.; Yari, A.; Salehi, E.; Mikaeili, S.; Dashtestani, F.; Abolhassani, F.; Rashki, L.; Shiasi, S.; Asadi, E. Melatonin Affects Membrane Integrity, Intracellular Reactive Oxygen Species, Caspase3 Activity and Akt Phosphorylation in Frozen Thawed Human Sperm. Cell Tissue Res. 2018, 372, 149–159. [Google Scholar] [CrossRef]
- Perumal, P.; Chang, S.; Baruah, K.K.; Srivastava, N. Administration of Slow Release Exogenous Melatonin Modulates Oxidative Stress Profiles and in Vitro Fertilizing Ability of the Cryopreserved Mithun (Bos Frontalis) Spermatozoa. Theriogenology 2018, 120, 79–90. [Google Scholar] [CrossRef]
- Pool, K.R.; Rickard, J.P.; Tumeth, E.; de Graaf, S.P. Treatment of Rams with Melatonin Implants in the Non-Breeding Season Improves Post-Thaw Sperm Progressive Motility and Dna Integrity. Anim. Reprod. Sci. 2020, 221, 106579. [Google Scholar] [CrossRef] [PubMed]
- Inyawilert, W.; Rungruangsak, J.; Liao, Y.J.; Tang, P.C.; Paungsukpaibool, V. Melatonin Supplementation Improved Cryopreserved Thai Swamp Buffalo Semen. Reprod. Domest. Anim. 2021, 56, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Martín-Hidalgo, D.; Barón, F.J.; Bragado, M.J.; Carmona, P.; Robina, A.; García-Marín, L.J.; Gil, M.C. The Effect of Melatonin on the Quality of Extended Boar Semen After Long-Term Storage at 17 °C. Theriogenology 2011, 75, 1550–1560. [Google Scholar] [CrossRef]
- Ofosu, J.; Qazi, I.H.; Fang, Y.; Zhou, G. Use of Melatonin in Sperm Cryopreservation of Farm Animals: A Brief Review. Anim. Reprod. Sci. 2021, 233, 106850. [Google Scholar] [CrossRef]
- Esplugues, J.V.; Rocha, M.; Nunez, C.; Bosca, I.; Ibiza, S.; Herance, J.R.; Ortega, A.; Serrador, J.M.; D’Ocon, P.; Victor, V.M. Complex I Dysfunction and Tolerance to Nitroglycerin: An Approach Based on Mitochondrial-Targeted Antioxidants. Circ. Res. 2006, 99, 1067–1075. [Google Scholar] [CrossRef]
- Veiko, A.G.; Sekowski, S.; Lapshina, E.A.; Wilczewska, A.Z.; Markiewicz, K.H.; Zamaraeva, M.; Zhao, H.C.; Zavodnik, I.B. Flavonoids Modulate Liposomal Membrane Structure, Regulate Mitochondrial Membrane Permeability and Prevent Erythrocyte Oxidative Damage. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183442. [Google Scholar] [CrossRef]
- de Oliveira, M.R.; Nabavi, S.M.; Braidy, N.; Setzer, W.N.; Ahmed, T.; Nabavi, S.F. Quercetin and the Mitochondria: A Mechanistic View. Biotechnol. Adv. 2016, 34, 532–549. [Google Scholar] [CrossRef]
- Ferramosca, A.; Lorenzetti, S.; Di Giacomo, M.; Lunetti, P.; Murrieri, F.; Capobianco, L.; Dolce, V.; Coppola, L.; Zara, V. Modulation of Human Sperm Mitochondrial Respiration Efficiency by Plant Polyphenols. Antioxidants 2021, 10, 217. [Google Scholar] [CrossRef]
- Nass-Arden, L.; Breitbart, H. Modulation of Mammalian Sperm Motility by Quercetin. Mol. Reprod. Dev. 1990, 25, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Abdelnour, S.A.; Sindi, R.A.; Abd, E.M.; Khalifa, N.E.; Khafaga, A.F.; Noreldin, A.E.; Samir, H.; Tufarelli, V.; Losacco, C.; Gamal, M.; et al. Quercetin: Putative Effects on the Function of Cryopreserved Sperms in Domestic Animals. Reprod. Domest. Anim. 2023, 58, 191–206. [Google Scholar] [CrossRef]
- Salehi, E.; Shadboorestan, A.; Mohammadi-Bardbori, A.; Mousavi, A.; Kargar-Abargouei, E.; Sarkoohi, P.; Omidi, M. Effect of Crocin and Quercetin Supplementation in Cryopreservation Medium on Post-Thaw Human Sperm Quality. Cell Tissue Bank. 2024, 25, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Bang, S.; Tanga, B.M.; Fang, X.; Seong, G.; Saadeldin, I.M.; Qamar, A.Y.; Lee, S.; Kim, K.J.; Park, Y.J.; Nabeel, A.; et al. Cryopreservation of Pig Semen Using a Quercetin-Supplemented Freezing Extender. Life 2022, 12, 1155. [Google Scholar] [CrossRef] [PubMed]
- Mao, T.; Han, C.; Wei, B.; Zhao, L.; Zhang, Q.; Deng, R.; Liu, J.; Luo, Y.; Zhang, Y. Protective Effects of Quercetin Against Cadmium Chloride-Induced Oxidative Injury in Goat Sperm and Zygotes. Biol. Trace Elem. Res. 2018, 185, 344–355. [Google Scholar] [CrossRef]
- Najafi, A.; Kia, H.D.; Mehdipour, M.; Hamishehkar, H.; Alvarez-Rodriguez, M. Effect of Quercetin Loaded Liposomes or Nanostructured Lipid Carrier (Nlc) on Post-Thawed Sperm Quality and Fertility of Rooster Sperm. Theriogenology 2020, 152, 122–128. [Google Scholar] [CrossRef]
- Bang, S.; Qamar, A.Y.; Tanga, B.M.; Fang, X.; Seong, G.; Nabeel, A.; Yu, I.J.; Saadeldin, I.M.; Cho, J. Quercetin Improves the Apoptotic Index and Oxidative Stress in Post-Thaw Dog Sperm. Environ. Sci. Pollut. Res. Int. 2022, 29, 21925–21934. [Google Scholar] [CrossRef]
- Bharati, S.; Shetty, S. Mitochondria-Targeted Antioxidants and Cancer; Springer: Singapore, 2022; pp. 1167–1188. [Google Scholar]
- Wang, P.F.; Xie, K.; Cao, Y.X.; Zhang, A. Hepatoprotective Effect of Mitochondria-Targeted Antioxidant Mito-Tempo Against Lipopolysaccharide-Induced Liver Injury in Mouse. Mediat. Inflamm. 2022, 2022, 6394199. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Zhang, Y.; Bai, H.; Liu, J.; Li, J.; Wu, B. Mitochondria-Targeted Antioxidant Mitotempo Improves the Post-Thaw Sperm Quality. Cryobiology 2018, 80, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Liu, J.; Zheng, Q.Y.; Liu, N.; Huang, X.L.; Wu, Y.Y.; Yao, X.F.; Tan, Q.Y.; Huang, Y.; Hu, C.H.; et al. The Effect of the Mitochondria-Targeted Antioxidant Mito-Tempo During Sperm Ultra-Rapid Freezing. Cryobiology 2024, 114, 104860. [Google Scholar] [CrossRef]
- Zhang, X.; Lu, X.; Li, J.; Xia, Q.; Gao, J.; Wu, B. Mito-Tempo Alleviates Cryodamage by Regulating Intracellular Oxidative Metabolism in Spermatozoa from Asthenozoospermic Patients. Cryobiology 2019, 91, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ghosh, S.K.; Katiyar, R.; Rautela, R.; Bisla, A.; Ngou, A.A.; Pande, M.; Srivastava, N.; Bhure, S.K. Effect of Mito-Tempo Incorporated Semen Extender on Physico-Morphological Attributes and Functional Membrane Integrity of Frozen Thawed Buffalo Spermatozoa. Cryo Lett. 2021, 42, 111–119. [Google Scholar]
- Kumar, A.; Kumar, G.S.; Katiyar, R.; Gemeda, A.E.; Rautela, R.; Bisla, A.; Srivastava, N.; Kumar, B.S.; Devi, H.L.; Chandra, V. Supplementation of Mito Tempo and Acetovanillone in Semen Extender Improves Freezability of Buffalo Spermatozoa. Andrology 2022, 10, 775–788. [Google Scholar] [CrossRef]
- Asadzadeh, N.; Abdollahi, Z.; Esmaeilkhanian, S.; Masoudi, R. Fertility and Flow Cytometry Evaluations of Ram Frozen Semen in Plant-Based Extender Supplemented with Mito-Tempo. Anim. Reprod. Sci. 2021, 233, 106836. [Google Scholar] [CrossRef]
- Shi, L.; Shi, J.; Feng, J.; Zhang, P.; Ren, Y. Proteomic Analysis Reveals the Potential Positive Effects of Mito-Tempo on Ram Sperm Motility and Fertility During Cryopreservation. Theriogenology 2023, 205, 27–39. [Google Scholar] [CrossRef]
- Zarei, F.; Daghigh-Kia, H.; Masoudi, R. Supplementation of Ram’s Semen Extender with Mito-Tempo II: Quality Evaluation and Flow Cytometry Study of Post-Thawed Spermatozoa. Andrologia 2022, 54, e14299. [Google Scholar] [CrossRef]
- Zarei, F.; Kia, H.D.; Masoudi, R.; Moghaddam, G.; Ebrahimi, M. Supplementation of Ram’s Semen Extender with Mito-Tempo I: Improvement in Quality Parameters and Reproductive Performance of Cooled-Stored Semen. Cryobiology 2021, 98, 215–218. [Google Scholar] [CrossRef]
- Masoudi, R.; Asadzadeh, N.; Sharafi, M. Effects of Freezing Extender Supplementation with Mitochondria-Targeted Antioxidant Mito-Tempo on Frozen-Thawed Rooster Semen Quality and Reproductive Performance. Anim. Reprod. Sci. 2021, 225, 106671. [Google Scholar] [CrossRef]
- Masoudi, R.; Asadzadeh, N.; Sharafi, M. The Mitochondria-Targeted Antioxidant Mito-Tempo Conserves Rooster’s Cooled Semen Quality and Fertility Potential. Theriogenology 2020, 156, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.H.; Banchi, P.; Domain, G.; Vanderheyden, L.; Prochowska, S.; Nizanski, W.; Van Soom, A. Mito-Tempo Improves Acrosome Integrity of Frozen-Thawed Epididymal Spermatozoa in Tomcats. Front. Vet. Sci. 2023, 10, 1170347. [Google Scholar] [CrossRef]
- Alipour-Jenaghard, P.; Daghigh-Kia, H.; Masoudi, R.; Hatefi, A. Mitoq Preserves Epigenetic Modifications and Quality Parameters of Rooster Sperm During Cryopreservation Process. Reprod. Domest. Anim. 2025, 60, e70012. [Google Scholar] [CrossRef]
- Nateghi, R.; Masoudi, R.; Asadzadeh, N. Supplementing the Beltsville Extender with Mitoquinol Improves the Quality and Fertility Potential of the Rooster’s Cooled Sperm. Reprod. Domest. Anim. 2024, 59, e14740. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yu, J.; Yang, Q.; Xie, Y.; Li, X.; Chen, Z.; Xiong, Y.; Fu, W.; He, H.; Yin, S.; et al. Mitochondria-Targeted Antioxidant Mitoq Improves the Quality of Low Temperature-Preserved Yak Semen Via Alleviating Oxidative Stress. Anim. Reprod. Sci. 2025, 273, 107680. [Google Scholar] [CrossRef]
- Gonzalez, M.; Prashar, T.; Connaughton, H.; Barry, M.; Robker, R.; Rose, R. Restoring Sperm Quality Post-Cryopreservation Using Mitochondrial-Targeted Compounds. Antioxidants 2022, 11, 1808. [Google Scholar] [CrossRef]
- Elkhawagah, A.R.; Donato, G.G.; Poletto, M.; Martino, N.A.; Vincenti, L.; Conti, L.; Necchi, D.; Nervo, T. Effect of Mitoquinone on Sperm Quality of Cryopreserved Stallion Semen. J. Equine Vet. Sci. 2024, 141, 105168. [Google Scholar] [CrossRef]
- Hatami, M.; Masoudi, R.; Hatefi, A.; Alipour-Jenaghard, P.; Esmaeili, V. The Effects of Mitoq as a Mitochondrial-Targeted Antioxidant in a Plant-Based Extender on Buck Sperm Quality Parameters During Cryopreservation. Anim. Reprod. Sci. 2024, 266, 107517. [Google Scholar] [CrossRef]
- Camara, D.R.; Ibanescu, I.; Siuda, M.; Bollwein, H. Mitoquinone Does Not Improve Sperm Cryo-Resistance in Bulls. Reprod. Domest. Anim. 2022, 57, 10–18. [Google Scholar] [CrossRef]
- Tvrda, E.; Debacker, M.; Duracka, M.; Kovac, J.; Bucko, O. Quercetin and Naringenin Provide Functional and Antioxidant Protection to Stored Boar Semen. Animals 2020, 10, 1930. [Google Scholar] [CrossRef]
- Mehdipour, M.; Mohammadi, H.; Salih, S.A.; Rashidi, A. Mitochondrial Specific Antioxidant Mitopbn Mitigates Oxidative Stress and Improves Mitochondrial Function in Cryopreserved Ram Sperm. Sci. Rep. 2025, 15, 10526. [Google Scholar] [CrossRef]
- Boussabbeh, M.; Haddar, M.; Sallem, A.; Chaieb, A.; Khdhiri, R.; Abid Essefi, S.; Mehdi, M. Enhancing Male Fertility: The Role of Crocin in Boosting Sperm Motility through Antioxidant Activity and Mitochondrial Pathways. J. Biochem. Mol. Toxicol. 2025, 39, e70275. [Google Scholar] [CrossRef] [PubMed]
- Khan, G.S.; Tahir, M.Z.; Zahoor, M.Y.; Rahman, H.U.; Riaz, A. Ameliorative Effect of Crocin on Post-Thaw Quality, Fertility-Associated Gene Expression and Fertilization Potential of Buffalo (Bubalus bubalis) Bull Sperm. Reprod. Domest. Anim. 2024, 59, e14519. [Google Scholar] [CrossRef]
- Lv, Y.Q.; Ji, S.; Chen, X.; Xu, D.; Luo, X.T.; Cheng, M.M.; Zhang, Y.Y.; Qu, X.L.; Jin, Y. Effects of Crocin on Frozen-Thawed Sperm Apoptosis, Protamine Expression and Membrane Lipid Oxidation in Yanbian Yellow Cattle. Reprod. Domest. Anim. 2020, 55, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Salman, A.; Caamano, J.N.; Fernandez-Alegre, E.; Hidalgo, C.O.; Nadri, T.; Tamargo, C.; Fueyo, C.; Fernandez, A.; Merino, M.J.; Martinez-Pastor, F. Supplementation of the Bioxcell Extender with the Antioxidants Crocin, Curcumin and Gsh for Freezing Bull Semen. Res. Vet. Sci. 2021, 136, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Mehdipour, M.; Daghigh, K.H.; Najafi, A.; Mohammadi, H.; Alvarez-Rodriguez, M. Effect of Crocin and Naringenin Supplementation in Cryopreservation Medium on Post-Thaw Rooster Sperm Quality and Expression of Apoptosis Associated Genes. PLoS ONE 2020, 15, e0241105. [Google Scholar] [CrossRef] [PubMed]
- Khan, G.S.; Tahir, M.Z.; Zahoor, M.Y.; Hifz-Ul-Rahman; Riaz, A. Effect of Naringenin on Post-Thaw Quality, Fertility-Associated Gene Expression and Fertilization Potential of Buffalo (Bubalus bubalis) Bull Sperm. Cryobiology 2024, 116, 104953. [Google Scholar] [CrossRef]
- Usuga, A.; Vergara, A.K.; Tobon, M.C.; Vargas, S.; Rojano, B.; Restrepo, G. Metformin and Rosiglitazone Affect Motility, Lipid Peroxidation and Mitochondrial Activity of Thawed Equine Spermatozoa. J. Equine Vet. Sci. 2025, 149, 105570. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, S.M.; Zhu, D.W.; Li, Y.; Wen, F.; Xian, M.; Hu, Z.T.; Zou, Q.L.; Zhang, L.K.; Chen, Y.L.; et al. Metformin Improves Sheep Sperm Cryopreservation Via Vitalizing the Ampk Pathway. Theriogenology 2023, 208, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhu, C.; Ma, Y.; Wang, Y.; Zheng, L.; Jin, T.; He, S.; Yang, F.; Dong, W. Metformin Improves Fish Sperm Quality by Regulating Glucose Uptake Capacity During In Vitro Storage. J. Anim. Sci. 2023, 101, skad152. [Google Scholar] [CrossRef]
- Calle-Guisado, V.; Gonzalez-Fernandez, L.; Martin-Hidalgo, D.; Garcia-Marin, L.J.; Bragado, M.J. Metformin Inhibits Human Spermatozoa Motility and Signalling Pathways Mediated by Protein Kinase a and Tyrosine Phosphorylation without Affecting Mitochondrial Function. Reprod. Fertil. Dev. 2019, 31, 787–795. [Google Scholar] [CrossRef]
- Hurtado, D.L.A.; Martin-Hidalgo, D.; Garcia-Marin, L.J.; Bragado, M.J. Metformin Blocks Mitochondrial Membrane Potential and Inhibits Sperm Motility in Fresh and Refrigerated Boar Spermatozoa. Reprod. Domest. Anim. 2018, 53, 733–741. [Google Scholar] [CrossRef]
- Li, R.N.; Zhu, Z.D.; Zheng, Y.; Lv, Y.H.; Tian, X.E.; Wu, D.; Wang, Y.J.; Zeng, W.X. Metformin Improves Boar Sperm Quality Via 5′-Amp-Activated Protein Kinase-Mediated Energy Metabolism in Vitro. Zool. Res. 2020, 41, 527–538. [Google Scholar] [CrossRef]
- Grandhaye, J.; Partyka, A.; Ligocka, Z.; Dudek, A.; Nizanski, W.; Jeanpierre, E.; Estienne, A.; Froment, P. Metformin Improves Quality of Post-Thaw Canine Semen. Animals 2020, 10, 287. [Google Scholar] [CrossRef]
- Swegen, A.; Lambourne, S.R.; Aitken, R.J.; Gibb, Z. Rosiglitazone Improves Stallion Sperm Motility, Atp Content, and Mitochondrial Function. Biol. Reprod. 2016, 95, 107. [Google Scholar] [CrossRef] [PubMed]
- Losano, J.; Daigneault, B.W. Pharmacological Perturbation of Peroxisome-Proliferator-Activated Receptor Gamma Alters Motility and Mitochondrial Function of Bovine Sperm. Andrology 2023, 11, 155–166. [Google Scholar] [CrossRef]
- Mehdipour, M.; Daghigh-Kia, H.; Najafi, A.; Mehdipour, Z.; Mohammadi, H. Protective Effect of Rosiglitazone on Microscopic and Oxidative Stress Parameters of Ram Sperm After Freeze-Thawing. Sci. Rep. 2022, 12, 13981. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Yang, K.; Guo, H.T.; Wang, J.R.; Sun, H.H.; Wang, S.W.; Sun, M.; Sun, L.Z.; Yue, S.L.; Zhou, J.B. Protective Influence of Rosiglitazone Against Time-Dependent Deterioration of Boar Spermatozoa Preserved at 17 Degrees C. Reprod. Domest. Anim. 2019, 54, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, Q.; Min, L.; Cao, H.; Adetunji, A.O.; Zhou, K.; Zhu, Z. Pyrroloquinoline Quinone Improved Boar Sperm Quality Via Maintaining Mitochondrial Function During Cryopreservation. Antioxidants 2025, 14, 102. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Li, W.; Yang, Q.; Zhao, H.; Zhang, W.; Adetunji, A.O.; Hoque, S.; Kou, X.; Min, L. Pyrroloquinoline Quinone Improves Ram Sperm Quality through its Antioxidative Ability During Storage at 4 Degrees C. Antioxidants 2024, 13, 104. [Google Scholar] [CrossRef]
- Younus, A.M.; Yamanaka, T.; Shimada, M. The Protective Effects of Antioxidants Against Endogenous and Exogenous Oxidative Stress on Bull Sperm. Vitr. Cell Dev. Biol. Anim. 2024, 60, 969–982. [Google Scholar] [CrossRef]
- Gao, S.; Zhao, B.X.; Long, C.; Heng, N.; Guo, Y.; Sheng, X.H.; Wang, X.G.; Xing, K.; Xiao, L.F.; Ni, H.M.; et al. Natural Astaxanthin Improves Testosterone Synthesis and Sperm Mitochondrial Function in Aging Roosters. Antioxidants 2022, 11, 1684. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Y.; Kang, H.Y.; Guo, Y.; Sheng, X.H.; Wang, X.G.; Xing, K.; Xiao, L.F.; Lv, X.Z.; Long, C.; Qi, X.L. Effect of Natural Astaxanthin on Sperm Quality and Mitochondrial Function of Breeder Rooster Semen Cryopreservation. Cryobiology 2024, 117, 104979. [Google Scholar] [CrossRef]
- Sohail, T.; Zhang, L.; Wang, X.; Jiang, C.; Wang, J.; Sun, X.; Li, Y. Astaxanthin Improved the Quality of Hu Ram Semen by Increasing the Antioxidant Capacity and Mitochondrial Potential and Mitigating Free Radicals-Induced Oxidative Damage. Animals 2024, 14, 319. [Google Scholar] [CrossRef]
- Qamar, A.Y.; Fang, X.; Bang, S.; Shin, S.T.; Cho, J. The Effect of Astaxanthin Supplementation on the Post-Thaw Quality of Dog Semen. Reprod. Domest. Anim. 2020, 55, 1163–1171. [Google Scholar] [CrossRef]
- Guo, Q.; Liu, X.; Li, Y.; Cheng, Y.; Li, J. Ergothioneine Improves the Quality of Boar Sperm During In Vitro Liquid Preservation by Regulating Mitochondrial Respiratory Chain. Animals 2025, 15, 1450. [Google Scholar] [CrossRef]
- Usuga, A.; Tejera, I.; Gomez, J.; Restrepo, O.; Rojano, B.; Restrepo, G. Cryoprotective Effects of Ergothioneine and Isoespintanol on Canine Semen. Animals 2021, 11, 2757. [Google Scholar] [CrossRef]
- Abdalkarim, S.S.; Daghigh-Kia, H.; Mehdipour, M.; Najafi, A. Does Ergothioneine and Thawing Temperatures Improve Rooster Semen Post-Thawed Quality? Poult. Sci. 2021, 100, 101405. [Google Scholar] [CrossRef]
- Vandvali, F.; Kia, H.D.; Ebrahimi, M.; Moghaddam, G.; Najafi, A. Impact of Losartan-Supplemented Extender on Enhancing Rooster Sperm Quality Post-Freezing and Thawing. Poult. Sci. 2025, 104, 105179. [Google Scholar] [CrossRef]
- Sindi, R.A.; Abdelnour, S.A.; El-Haroun, E.; Alfattah, M.A.; Saber, Y.; Sheiha, A.M. Does Lagenaria Siceraria Seed Oil-Enriched Extender Regulate Sperm Quality, Oxidant/Antioxidant Markers, and Sperm Mitochondrial Enzymes in Chilled Diluted Rabbit Semen? BMC Vet. Res. 2025, 21, 345. [Google Scholar] [CrossRef] [PubMed]
- Jorge, M.; Ferreira, F.C.; Marques, C.C.; Batista, M.C.; Oliveira, P.J.; Lidon, F.; Duarte, S.C.; Teixeira, J.; Pereira, R. Effect of Urolithin a on Bovine Sperm Capacitation and in Vitro Fertilization. Animals 2024, 14, 2726. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.C.; Marques, C.C.; Baptista, M.C.; Pimenta, J.; Teixeira, J.; Montezinho, L.; Cagide, F.; Borges, F.; Oliveira, P.J.; Pereira, R. Effect of a Novel Hydroxybenzoic Acid Based Mitochondria Directed Antioxidant Molecule on Bovine Sperm Function and Embryo Production. Animals 2022, 12, 804. [Google Scholar] [CrossRef]
- Coyan, K.; Baspinar, N.; Bucak, M.N.; Akalin, P.P. Effects of Cysteine and Ergothioneine on Post-Thawed Merino Ram Sperm and Biochemical Parameters. Cryobiology 2011, 63, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xiong, M.; Zhang, J.; Li, S.; Ma, S.; Jiang, S.; Jiang, Y.; Li, X. Polydopamine-Based Nano-Protectant for Prolonged Boar Semen Preservation by Eliminating Ros and Regulating Protein Phosphorylation Via D2Dr-Mediated Camp/Pka Signaling Pathway. J. Nanobiotechnol. 2025, 23, 151. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Protein | Species | Function | Ref. |
|---|---|---|---|
| Ant2 | Drosophila | Disrupting mitochondrial morphogenesis and spermatid maturation | [51] |
| MORN2 | Mouse | MORN2 functions in mitochondrial sheath formation and regulates mitochondrial respiratory activity | [52] |
| ClpP/ClpX | Mouse | ClpP/ClpX deficiency disrupted energy supply during meiosis and impaired zygotene-pachytene transformation in male germ cells | [53] |
| Rsrc1-161aa | Mouse | Regulating mitochondrial ribosome assembly and translation, and then affecting male fertility. | [54] |
| SLC22A14 | Mouse | Shortage of SLC22A14 could decrease the energy metabolism of spermatozoa | [55] |
| PRSS55 | Mouse | Deficiency of PRSS55 damages the mitochondrial structure, which leads to lower ETC molecules and ATP contents | [56] |
| SLC7A11 antiporter | Stallion | Transporting cystine into the cell in exchange for glutamate | [57] |
| DJ-1 | Rat | Interacting mitochondrial complex I subunits NDUFS3 and NDUFA4 affect mitochondrial energy metabolism | [58] |
| GSK3 | Goat | Phosphorylation of GSK3 regulated sperm motility and acrosome reaction by glycolysis and OXPHOS | [59] |
| TPPP2 | Mouse | Deficiency of TPPP2 altered the structure and function of sperm mitochondria and caused decreased sperm count, motility and ATP content | [60] |
| AMPK | Mouse/Goat | Increasing AMPK activity maintains MMP and ATP levels in spermatozoa | [61,62] |
| LDHC | Mouse | Maintenance of energy metabolism in sperm | [63] |
| GLUT8 | Mouse | Lack of GLUT8 leads to impaired mitochondrial function and sperm motility in spermatozoa | [64] |
| Antioxidants | Species | Functions | Ref. |
|---|---|---|---|
| MitoQ | Rooster | Improve the sperm quality in cryopreserved and cooled rooster spermatozoa | [167,168] |
| Yak | Increase the sperm quality and antioxidant capacity in low-temperature yak semen preservation | [169] | |
| Human | Enhance functional sperm parameters, reduce DNA fragmentation, and upregulate the expression of antioxidant-related genes in human sperm cryopreservation medium | [125,170] | |
| Stallion | Improve the frozen-thawed stallion sperm motility and kinetics, while detrimental to the motility and viability at higher concentrations | [171] | |
| Buck | Improve the motility, viability, membrane functionality, mitochondrial activity, acrosome integrity, and reduce OS during the cryopreservation process of buck sperm | [172] | |
| Bull | Do not affect the Simmental bull sperm cryo-resistance | [173] | |
| Enhance post-thaw buffalo bull sperm function and cryo-tolerance | [126] | ||
| yellow catfish | Reduce ROS production and LP, and enhance post-thaw sperm viability of yellow catfish | [127] | |
| Melatonin | Mouse | Alleviate post-weaning NaAsO2 exposure-mediated decline of sperm counts in adult mice | [133] |
| Human | Improve human sperm quality and overall male fertility | [132] | |
| Counteract intracellular ROS and MDA, increase post-thaw viability, motility and membrane integrity of human spermatozoa | [134,135,136] | ||
| Ram | Improve OXPHOS of frozen-thawed ram sperm | [131] | |
| Improve progressive motility and reduce DNA fragmentation of ram spermatozoa following cryopreservation | [138] | ||
| Bull | Improve sperm function parameters in the fresh and post-thaw mithun bull semen | [137] | |
| Improve sperm viability and motility of swamp buffalo during cryopreservation | [139] | ||
| Boar | Increase the number of live sperms with intact acrosome, but did not affect the other spermatic quality of boar semen | [140] | |
| Quercetin | Human | Improve the human sperm progressive motility, MMP, and decrease ROS levels, DNA fragmentation | [148] |
| Stimulate the respiratory active state and enhance mitochondrial function in asthenozoospermic samples | [145] | ||
| Boar | Maintain sperm motility and decrease superoxide in the liquid preservation of boar semen, and improve the post-thaw sperm quality and reduce the OS of boar sperm | [149,174] | |
| Canine | Improve the post-thaw sperm quality and reduce the oxidative damage in the dog | [152] | |
| Goat | Decrease the goat sperm MDA and ROS levels, enhance motility, viability, membrane integrity, and mitochondria activity | [150] | |
| Rooster | Improve post-thawed sperm quality and fertility of roosters | [151] | |
| Ram | Inhibit the ram spermatozoa motility during the first 2 h of incubation, but subsequently stimulate motility over the following 3–4 h by sustaining mitochondrial respiration | [146] | |
| MitoTEMPO | Human | Improve post-thaw human sperm motility, viability, membrane integrity, MMP and antioxidant enzymes activities decrease DNA fragmentation index, ROS and MDA level | [155,156,157] |
| Bull | Improve post-thaw buffalo sperm progressive motility, viability, acrosomal and membrane integrity, cholesterol to phospholipids ratio, MMP, antioxidant enzymes activities and fertility | [158,159] | |
| Ram | Improve the post-thaw ram sperm motility, kinematics, viability, membrane functionality, MMP, antioxidant capacity, glucose transporter and fertility, decreasing MDA | [160,161,162,163] | |
| Rooster | Improve the post-thaw rooster sperm motility kinematics, membrane functionality, MMP, viability and fertility, decrease lipid peroxidation, late apoptotic-like changes, DNA fragmentation and hydrogen peroxide level | [164,165] | |
| Tomcat | Improve the acrosome integrity of frozen-thawed epididymal spermatozoa in tomcats | [166] | |
| MitoPBN | Ram | Mitigate OS and improve mitochondrial function in cryopreserved ram sperm | [175] |
| Crocin | Human | Improve human sperm progressive motility, MMP, and decrease ROS levels, DNA fragmentation, enhance the asthenozoospermic sperm motility and reduce OS | [148,176] |
| Bull | Improve the sperm post-thaw quality and fertility of buffalo bull and Yanbian yellow cattle sperm | [177,178,179] | |
| Rooster | Improve the post-thawing rooster sperm viability, membrane functionality, MMP and fertility, reduce sperm LP and apoptosis | [180] | |
| Naringenin | Bull | Improve buffalo bull post-thaw quality, fertility-associated gene expression and fertilization potential | [181] |
| Boar | Maintenance of sperm motility and decrease of superoxide in the liquid preservation of boar semen | [174] | |
| Rooster | Improve the post-thawing rooster sperm viability, membrane functionality, MMP and fertility, reduce sperm LP and apoptosis | [180] | |
| Metformin | Equine | Improve total motility and reduce LP of thawed equine spermatozoa | [182] |
| Sheep | Improve the post-thawing sheep sperm quality and reduce sperm OS | [183] | |
| S. prenanti | Improve the quality and fertility of S. prenanti sperm by increasing the ATP content | [184] | |
| Human | Inhibit human sperm motility, PKA and protein tyrosine phosphorylation pathways, do not affect sperm viability, MMP, mitochondrial superoxide anion generation | [185] | |
| Boar | The effects on boar sperm quality remain controversial | [186,187] | |
| Canine | Improve the frozen-thawed canine sperm motility, mitochondrial activity, NAD+ content, and reduce the OS level | [188] | |
| Rosiglitazone | Equine | Increase the ROS level of thawed equine spermatozoa. Increase motility, MMP, ATP content, glucose uptake capacity, and decrease ROS level of equine spermatozoa in vitro. | [182,189] |
| Bull | Maintain MMP, total and progressive motility of bovine sperm | [190] | |
| Ram | Enhance motility, membrane and acrosome integrity, MMP, and decrease ROS level in frozen-thawed ram spermatozoa | [191] | |
| Boar | Enhance boar sperm motility, membrane and acrosome integrity, MMP, ATP production and reduce ROS during storage at 17 °C | [192] | |
| Pyrroloquinoline quinone | Boar | Enhance post-thaw boar sperm motility, viability, acrosome integrity, MMP, ATP levels, mtDNA stability, and decrease MDA, ROS levels and DNA damage | [193] |
| Ram | Improve sperm motility, MMP, membrane and acrosome integrity, ATP levels and fertility, decrease sperm MDA and ROS levels of ram sperm at 4 °C | [194] | |
| Bull | Reduce mitochondrial ROS in fresh bull semen, improve sperm motility and reduce ROS in frozen-thawed bull sperm | [195] | |
| Astaxanthin | Rooster | Improve rooster freeze-thaw sperm viability, motility, plasma membrane and acrosome integrity, mitochondrial activities, ATP level and antioxidant ability | [196,197] |
| Ram | Increase sperm motility, viability, plasma membrane and acrosome integrity, MMP and antioxidant ability of Hu ram spermatozoa at 4 °C | [198] | |
| Canine | Increase sperm motility, viability, plasma membrane and acrosome integrity, and mitochondria activity of post-thaw dog spermatozoa | [199] | |
| Ergothioneine | Boar | Improve sperm motility parameters, MMP, ATP, antioxidant capacity, plasma membrane and acrosome integrity in in vitro liquid preservation of boar spermatozoa | [200] |
| Bull | Improve sperm motility and reduce ROS level in frozen-thawed bull sperm | [195] | |
| Canine | Increase the total motility, acrosomal integrity and reduce the abnormal morphology and ROS production of cryopreserved canine spermatozoa | [201] | |
| Rooster | Increase the total motility, membrane integrity and mitochondria activity, reduce the apoptotic and dead sperm of post-thawed rooster spermatozoa | [202] | |
| Losartan | Rooster | Improve the total and progressive motility, mitochondrial activity, membrane integrity, antioxidant levels, and reduce LP and apoptosis of post-thawed rooster spermatozoa | [203] |
| Lagenaria siceraria seed oil | Rabbit | Improve sperm motility, viability, membrane integrity and antioxidant capacity during 72-h chilled storage of rabbit spermatozoa | [204] |
| Urolithin A | Bull | Improve bovine sperm motility quality and reduce OS | [205] |
| AntiOxBEN2 | Bull | Enhance the bovine sperm quality, fertility, and reduce ROS | [206] |
| BGP-15 | Human | Improve sperm motility, mucous penetration and MMP, reduce sperm DNA oxidative damage of post-thawed human spermatozoa | [170] |
| L-Carnitine | Human | Improve sperm motility, viability, and reduce sperm DNA oxidative damage of post-thawed human spermatozoa | [170] |
| Cysteine | Ram | Improve the rates of ALH, membrane integrity, and mitochondrial activity of the post-thawed ram sperm | [207] |
| Isoespintanol | Canine | Increase the acrosomal integrity and reduce the abnormal morphology and ROS production of cryopreserved canine spermatozoa | [201] |
| Vitamin C | Bull | Reduce mitochondrial ROS in fresh bull semen, improve sperm motility and reduce ROS in frozen-thawed bull sperm | [195] |
| EGCG@PDA NPs | Boar | Maintain sperm motility, acrosome integrity, MMP, extending semen storage time from 3 days to 10 days, reducing ROS and sperm apoptosis of boar sperm during storage at 4 °C | [208] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Z.; Yan, Q.; Zhang, K.; Lei, Y.; Zhou, C.; Ren, T.; Gao, N.; Wen, F.; Li, X. Mitochondrial Regulation of Spermatozoa Function: Metabolism, Oxidative Stress and Therapeutic Insights. Animals 2025, 15, 2246. https://doi.org/10.3390/ani15152246
Xu Z, Yan Q, Zhang K, Lei Y, Zhou C, Ren T, Gao N, Wen F, Li X. Mitochondrial Regulation of Spermatozoa Function: Metabolism, Oxidative Stress and Therapeutic Insights. Animals. 2025; 15(15):2246. https://doi.org/10.3390/ani15152246
Chicago/Turabian StyleXu, Zhiqian, Qi Yan, Ke Zhang, Ying Lei, Chen Zhou, Tuanhui Ren, Ning Gao, Fengyun Wen, and Xiaoxia Li. 2025. "Mitochondrial Regulation of Spermatozoa Function: Metabolism, Oxidative Stress and Therapeutic Insights" Animals 15, no. 15: 2246. https://doi.org/10.3390/ani15152246
APA StyleXu, Z., Yan, Q., Zhang, K., Lei, Y., Zhou, C., Ren, T., Gao, N., Wen, F., & Li, X. (2025). Mitochondrial Regulation of Spermatozoa Function: Metabolism, Oxidative Stress and Therapeutic Insights. Animals, 15(15), 2246. https://doi.org/10.3390/ani15152246