
Gastrointestinal Parasitic Infections in Macaca fascicularis in Northeast Thailand: A One Health Perspective on Zoonotic Risks
 ,
,  ,
,
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Study Design and Location
2.2. Sample Size Calculation
2.3. Sample Collection
2.4. Parasitological Examination
2.5. Statistical Analysis
3. Results
3.1. Overall Prevalence of GI Infections
3.2. Distribution of Protozoan Species
3.3. Distribution of Helminth Species
3.4. Diagnostic Comparison Between APC and FECT Methods for Strongyloidiasis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rahman, M.T.; Sobur, M.A.; Islam, M.S.; Ievy, S.; Hossain, M.J.; El Zowalaty, M.E.; Rahman, A.T.; Ashour, H.M. Zoonotic Diseases: Etiology, Impact, and Control. Microorganisms 2020, 8, 1405. [Google Scholar] [CrossRef] [PubMed]
- Pitt, S.J.; Gunn, A. The One Health Concept. Br. J. Biomed. Sci. 2024, 81, 12366. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Gong, Y.; Chen, Q.; Li, S.; Zhou, Y. Global burden of soil-transmitted helminth infections, 1990–2021. Infect. Dis. Poverty. 2024, 13, 77. [Google Scholar] [CrossRef] [PubMed]
- Khattak, I.; Akhtar, A.; Shams, S.; Usman, T.; Haider, J.; Nasreen, N.; Khan, A.; Ben Said, M. Diversity, prevalence and risk factors associated to gastrointestinal tract parasites in wild and domestic animals from Pakistan. Parasitol. Int. 2023, 97, 102777. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, M.M.; Fouad, E.A.; Kamel, N.O.; Auda, H.M.; El-Bahy, M.M.; Ramadan, R.M. Dogs as a source for the spreading of enteric parasites including zoonotic ones in Giza Province, Egypt. Res. Vet. Sci. 2023, 161, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Kladkempetch, D.; Tangtrongsup, S.; Tiwananthagorn, S. Ancylostoma ceylanicum: The Neglected Zoonotic Parasite of Community Dogs in Thailand and Its Genetic Diversity among Asian Countries. Animals 2020, 10, 2154. [Google Scholar] [CrossRef] [PubMed]
- Pumipuntu, N. Detection for potentially zoonotic gastrointestinal parasites in long-tailed macaques, dogs and cattle at Kosamphi forest park, Maha Sarakham. Vet. Integr. Sci. 2018, 16, 69–77. [Google Scholar]
- Vonfeld, I.; Prenant, T.; Polack, B.; Guillot, J.; Quintard, B. Gastrointestinal parasites in non-human primates in zoological institutions in France. Parasite 2022, 29, 43. [Google Scholar] [CrossRef] [PubMed]
- Ribas, A.; Jollivet, C.; Morand, S.; Thongmalayvong, B.; Somphavong, S.; Siew, C.C.; Ting, P.J.; Suputtamongkol, S.; Saensombath, V.; Sanguankiat, S.; et al. Intestinal Parasitic Infections and Environmental Water Contamination in a Rural Village of Northern Lao PDR. Korean J. Parasitol. 2017, 55, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.J.d.A.; Rocha, A.C.d.S.; Assis-Silva, Z.M.d.; Maia, G.O.; Alves-Ribeiro, B.S.; Duarte, R.B.; Moraes, I.d.S.; Urzedo, N.F.d.; Fernandes-Silva, L.; Gomes, A.P.C.; et al. Risk of Environmental Contamination by Gastrointestinal Parasites in Public Areas of the Central Plateau Microregion of Brazil: A Public Health Concern. Pathogens 2025, 14, 211. [Google Scholar] [CrossRef] [PubMed]
- Crudo Blackburn, C.; Yan, S.M.; McCormick, D.; Herrera, L.N.; Iordanov, R.B.; Bailey, M.D.; Bottazzi, M.E.; Hotez, P.J.; Mejia, R. Parasitic Contamination of Soil in the Southern United States. Am. J. Trop. Med. Hyg. 2024, 111, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Jinatham, V.; Nonebudsri, C.; Wandee, T.; Popluechai, S.; Tsaousis, A.D.; Gentekaki, E. Blastocystis in tap water of a community in northern Thailand. Parasitol. Int. 2022, 91, 102624. [Google Scholar] [CrossRef] [PubMed]
- Wattanawong, O.; Iamsirithaworn, S.; Kophachon, T.; Nak-ai, W.; Wisetmora, A.; Wongsaroj, T.; Dekumyoy, P.; Nithikathkul, C.; Suwannatrai, A.T.; Sripa, B. Current status of helminthiases in Thailand: A cross-sectional, nationwide survey, 2019. Acta Trop. 2021, 223, 106082. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, A. Monkeys on the Edge: Ecology and Management of Long-Tailed Macaques and Their Interface with Humans; Cambridge University Press: Cambridge, UK, 2011; Volume 60. [Google Scholar]
- Malaivijitnond, S.; Hamada, Y. Current situation and status of long-tailed macaques (Macaca fascicularis) in Thailand. Trop. Nat. Hist. 2008, 8, 185–204. [Google Scholar] [CrossRef]
- Sricharern, W.; Inpankaew, T.; Kaewmongkol, S.; Jarudecha, T.; Inthong, N. Molecular identification of Trichuris trichiura and Hymenolepis diminuta in long-tailed macaques (Macaca fascicularis) in Lopburi, Thailand. Vet. World 2021, 14, 884–888. [Google Scholar] [CrossRef] [PubMed]
- Thanchomnang, T.; Intapan, P.M.; Sanpool, O.; Rodpai, R.; Sadaow, L.; Phosuk, I.; Somboonpatarakun, C.; Laymanivong, S.; Tourtip, S.; Maleewong, W. First molecular identification of Strongyloides fuelleborni in long-tailed macaques in Thailand and Lao People’s Democratic Republic reveals considerable genetic diversity. J. Helminthol. 2019, 93, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Schurer, J.M.; Ramirez, V.; Kyes, P.; Tanee, T.; Patarapadungkit, N.; Thamsenanupap, P.; Trufan, S.; Grant, E.T.; Garland-Lewis, G.; Kelley, S.; et al. Long-Tailed Macaques (Macaca fascicularis) in Urban Landscapes: Gastrointestinal Parasitism and Barriers for Healthy Coexistence in Northeast Thailand. Am. J. Trop. Med. Hyg. 2019, 100, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Zanzani, S.A.; Gazzonis, A.L.; Epis, S.; Manfredi, M.T. Study of the gastrointestinal parasitic fauna of captive non-human primates (Macaca fascicularis). Parasitol. Res. 2016, 115, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Arizono, N.; Yamada, M.; Tegoshi, T.; Onishi, K. Molecular identification of Oesophagostomum and Trichuris eggs isolated from wild Japanese macaques. Korean J. Parasitol. 2012, 50, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Klaus, A.; Zimmermann, E.; Röper, K.M.; Radespiel, U.; Nathan, S.; Goossens, B.; Strube, C. Co-infection patterns of intestinal parasites in arboreal primates (proboscis monkeys, Nasalis larvatus) in Borneo. Int. J. Parasitol. Parasites Wildl. 2017, 6, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Rivero, J.; Cutillas, C.; Callejón, R. Trichuris trichiura (Linnaeus, 1771) From Human and Non-human Primates: Morphology, Biometry, Host Specificity, Molecular Characterization, and Phylogeny. Front. Vet. Sci. 2020, 7, 626120. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.H.E.; Kusolsuk, T.; Watthanakulpanich, D.; Pakdee, W.; Doanh, P.N.; Yasin, A.M.; Dekumyoy, P.; Thaenkham, U. Prevalence of Strongyloides in Southeast Asia: A systematic review and meta-analysis with implications for public health and sustainable control strategies. Infect. Dis. Poverty 2023, 12, 83. [Google Scholar] [CrossRef] [PubMed]
- Esposito, M.M.; Turku, S.; Lehrfield, L.; Shoman, A. The Impact of Human Activities on Zoonotic Infection Transmissions. Animals 2023, 13, 1646. [Google Scholar] [CrossRef] [PubMed]
- Phosuk, I.; Thanchomnang, T.; Banglua, J.; Laymanivong, S.; Puangpronpitag, D.; Jongthawin, J. Molecular identification of Trichuris species in long-tailed macaques from Dong Ling Don Chao Pu Park and Kumphawapi Monkey Garden, Northeast Thailand: First report suggesting possible Trichuris ovis infection in non-human primates. Int. J. Parasitol. Parasites Wildl. 2025, 27, 101063. [Google Scholar] [CrossRef] [PubMed]
- Phosuk, I.; Thanchomnang, T.; Puimat, N.; Banglua, J.; Jongthawin, J. Prevalence and diversity of gastrointestinal parasites in long-tailed macaques at Kosamphi Forest Park, Kumphawapi Monkey Garden, and Dong Ling Don Chao Pu Park, North-east Thailand. Vet. World. 2024, 17, 1391–1396. [Google Scholar] [CrossRef] [PubMed]
- Wayne, W.D. Biostatistics: A Foundation for Analysis in the Health Sciences, 6th ed.; Wiley & Sons: New York, NY, USA, 1995. [Google Scholar]
- Koga, K.; Kasuya, S.; Khamboonruang, C.; Sukhavat, K.; Ieda, M.; Takatsuka, N.; Kita, K.; Ohtomo, H. A modified agar plate method for detection of Strongyloides stercoralis. Am. J. Trop. Med. Hyg. 1991, 45, 518–521. [Google Scholar] [CrossRef] [PubMed]
- Kaewrat, W.; Sengthong, C.; Yingklang, M.; Intuyod, K.; Haonon, O.; Onsurathum, S.; Dangtakot, R.; Saisud, P.; Sangka, A.; Anutrakulchai, S.; et al. Improved agar plate culture conditions for diagnosis of Strongyloides stercoralis. Acta Trop. 2020, 203, 105291. [Google Scholar] [CrossRef] [PubMed]
- Intapan, P.M.; Maleewong, W.; Wongsaroj, T.; Singthong, S.; Morakote, N. Comparison of the quantitative formalin ethyl acetate concentration technique and agar plate culture for diagnosis of human strongyloidiasis. J. Clin. Microbiol. 2005, 43, 1932–1933. [Google Scholar] [CrossRef] [PubMed]
- Anamnart, W.; Intapan, P.M.; Maleewong, W. Modified formalin-ether concentration technique for diagnosis of human strongyloidiasis. Korean J. Parasitol. 2013, 51, 743–745. [Google Scholar] [CrossRef] [PubMed]
- Sengthong, C.; Yingklang, M.; Intuyod, K.; Hongsrichan, N.; Pinlaor, S. An optimized agar plate culture improves diagnostic efficiency for Strongyloides stercoralis infection in an endemic community. Parasitol. Res. 2020, 119, 1409–1413. [Google Scholar] [CrossRef] [PubMed]
- Fauziyah, N.N.; Lastuti, N.D.R.; Damayanti, R.; Koesdarto, S.; Suprihati, E.; Witaningrum, A.M. Identification and The Prevalence of Gastrointestinal Endoparasite in Long-Tailed Macaque (Macaca fascicularis) in Wonorejo and Gunung Anyar Mangrove Eco-Tourism Surabaya. Vet. Biomed. Clin. J. 2023, 5, 35–39. [Google Scholar] [CrossRef]
- Baloria, H.T.; Gamalinda, E.F.; Rosal, J.J.; Estaño, L.A. Determination of enteroparasites in long-tailed macaques (Macaca fascicularis) of Barangay Sumile, Butuan city, Philippines. Asian J. Biol. Sci. 2022, 11, 751–756. [Google Scholar] [CrossRef]
- Teo, S.Z.; Tuen, A.A.; Madinah, A.; Aban, S.; Chong, Y.L. Occurrence of gastrointestinal nematodes in captive nonhuman primates at Matang Wildlife Centre, Sarawak. Trop. Biomed. 2019, 36, 594–603. [Google Scholar] [PubMed]
- de Ree, V.; Nath, T.C.; Barua, P.; Harbecke, D.; Lee, D.; Rödelsperger, C.; Streit, A. Genomic analysis of Strongyloides stercoralis and Strongyloides fuelleborni in Bangladesh. PLoS Negl. Trop. Dis. 2024, 18, e0012440. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cui, Z.; Li, X.; Zhang, L. Review of zoonotic amebiasis: Epidemiology, clinical signs, diagnosis, treatment, prevention and control. Res. Vet. Sci. 2021, 136, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Braseth, A.L.; Elliott, D.E.; Ince, M.N. Parasitic Infections of the Gastrointestinal Track and Liver. Gastroenterol. Clin. N. Am. 2021, 50, 361–381. [Google Scholar] [CrossRef] [PubMed]
- Janwan, P.; Rodpai, R.; Intapan, P.M.; Sanpool, O.; Tourtip, S.; Maleewong, W.; Thanchomnang, T. Possible transmission of Strongyloides fuelleborni between working Southern pig-tailed macaques (Macaca nemestrina) and their owners in Southern Thailand: Molecular identification and diversity. Infect. Genet. Evol. 2020, 85, 104516. [Google Scholar] [CrossRef] [PubMed]
- Ko, P.P.; Haraguchi, M.; Hara, T.; Hieu, D.D.; Ito, A.; Tanaka, R.; Tanaka, M.; Suzumura, T.; Ueda, M.; Yoshida, A.; et al. Population genetics study of Strongyloides fuelleborni and phylogenetic considerations on primate-infecting species of Strongyloides based on their mitochondrial genome sequences. Parasitol. Int. 2023, 92, 102663. [Google Scholar] [CrossRef] [PubMed]
- Thanchomnang, T.; Intapan, P.M.; Sanpool, O.; Rodpai, R.; Tourtip, S.; Yahom, S.; Kullawat, J.; Radomyos, P.; Thammasiri, C.; Maleewong, W. First molecular identification and genetic diversity of Strongyloides stercoralis and Strongyloides fuelleborni in human communities having contact with long-tailed macaques in Thailand. Parasitol. Res. 2017, 116, 1917–1923. [Google Scholar] [CrossRef] [PubMed]
- Shams, M.; Shamsi, L.; Yousefi, A.; Sadrebazzaz, A.; Asghari, A.; Mohammadi-Ghalehbin, B.; Shahabi, S.; Hatam, G. Current global status, subtype distribution and zoonotic significance of Blastocystis in dogs and cats: A systematic review and meta-analysis. Parasit. Vectors 2022, 15, 225. [Google Scholar] [CrossRef] [PubMed]
- Yowang, A.; Tsaousis, A.D.; Chumphonsuk, T.; Thongsin, N.; Kullawong, N.; Popluechai, S.; Gentekaki, E. High diversity of Blastocystis subtypes isolated from asymptomatic adults living in Chiang Rai, Thailand. Infect. Genet. Evol. 2018, 65, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Udonsom, R.; Prasertbun, R.; Mahittikorn, A.; Mori, H.; Changbunjong, T.; Komalamisra, C.; Pintong, A.-r.; Sukthana, Y.; Popruk, S. Blastocystis infection and subtype distribution in humans, cattle, goats, and pigs in central and western Thailand. Infect. Genet. Evol. 2018, 65, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Chistyakova, L.V.; Kostygov, A.Y.; Kornilova, O.A.; Yurchenko, V. Reisolation and redescription of Balantidium duodeni Stein, 1867 (Litostomatea, Trichostomatia). Parasitol. Res. 2014, 113, 4207–4215. [Google Scholar] [CrossRef] [PubMed]
- da Silva Barbosa, A.; Ponce-Gordo, F.; Dib, L.V.; Antunes Uchôa, C.M.; Bastos, O.M.P.; Pissinatti, A.; Amendoeira, M.R.R. First molecular characterization of Balantioides coli (Malmsten, 1857) isolates maintained in vitro culture and from feces of captive animals, Rio de Janeiro, Brazil. Vet. Parasitol. Reg. Stud. Rep. 2017, 10, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Pomajbíková, K.; Oborník, M.; Horák, A.; Petrželková, K.J.; Grim, J.N.; Levecke, B.; Todd, A.; Mulama, M.; Kiyang, J.; Modrý, D. Novel insights into the genetic diversity of Balantidium and Balantidium-like cyst-forming ciliates. PLoS Negl. Trop. Dis. 2013, 7, e2140. [Google Scholar] [CrossRef] [PubMed]
- Ghai, R.R.; Wallace, R.M.; Kile, J.C.; Shoemaker, T.R.; Vieira, A.R.; Negron, M.E.; Shadomy, S.V.; Sinclair, J.R.; Goryoka, G.W.; Salyer, S.J.; et al. A generalizable one health framework for the control of zoonotic diseases. Sci. Rep. 2022, 12, 8588. [Google Scholar] [CrossRef] [PubMed]



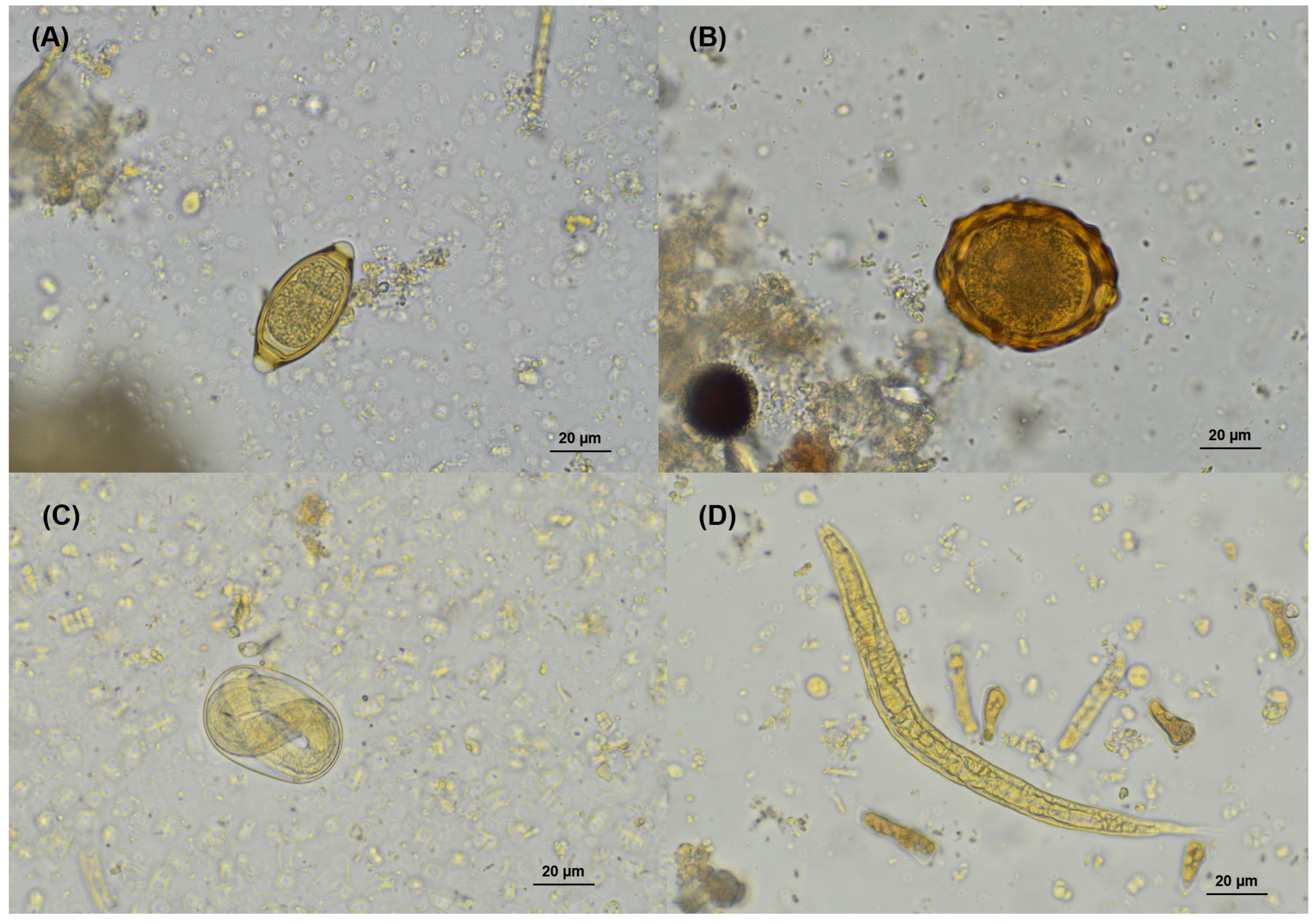

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Infection | Loei (n = 121) | Khon Kaen (n = 125) | Bueng Kan (n = 105) | Sisaket (n = 94) | Total (n = 445) | χ2 | p-Value * |
|---|---|---|---|---|---|---|---|
| n (%) | n (%) | n (%) | n (%) | n (%) | |||
| No. Positive (%) | 110 (90.91) | 114 (91.20) | 95 (90.48) | 66 (70.21) | 385 (86.52) | 26.31 | <0.001 |
| 95% CI | 84.45–94.85 | 84.93–95.02 | 83.35–94.74 | 60.32–78.52 | 83.00–89.40 | ||
| Protozoa | 95 (78.51) | 74 (59.20) | 38 (36.19) | 44 (46.81) | 251 (56.40) | ||
| Helminth | 81 (66.94) | 91 (72.80) | 86 (81.90) | 41 (43.62) | 299 (67.19) | ||
| Mono Protozoa | 29 (23.97) | 23 (18.40) | 9 (8.57) | 25 (26.60) | 86 (19.33) | ||
| Mono Helminth | 15 (12.40) | 40 (32.00) | 56 (61.90) | 22 (23.40) | 133 (29.89) | ||
| Mixed Infection | 66 (54.55) | 51 (40.80) | 30 (28.57) | 19 (20.21) | 166 (37.30) |
| Province | No. Examined | Species of Protozoa Identified | |||||
|---|---|---|---|---|---|---|---|
| Number of Positive Samples (%) | |||||||
| Balantioides coli-like | Iodamoeba bütschlii | Enthamoeba coli | Enthamoeba histiolytica like | Blastocytis sp. | Chilomastix mesnili | ||
| Loei | 121 | 71 (58.68) | 41 (33.88) | 12 (9.92) | 43 (35.54) | 0 (0.00) | 8 (6.61) |
| Khon Kaen | 125 | 33 (26.40) | 3 (2.40) | 20 (16.00) | 51 (40.80) | 1 (0.80) | 4 (3.20) |
| Bueng Kan | 105 | 13 (12.38) | 7 (6.67) | 1 (0.95) | 17 (16.19) | 0 (0.00) | 0 (0.00) |
| Sisaket | 94 | 21 (22.34) | 24 (25.53) | 22 (23.40) | 17 (18.09) | 0 (0.00) | 0 (0.00) |
| Total | 445 | 138 (31.01) | 75 (16.85) | 55 (12.36) | 128 (28.76) | 1 (0.22) | 12 (2.70) |
| Province | No. Examined | Species of Helminth Identified | ||
|---|---|---|---|---|
| Number of Positive Samples (%) | ||||
| Strongyloides sp. | Trichuris sp. | Ascaris sp. | ||
| Loei | 121 | 78 (64.46) | 1 (0.83) | 7 (5.79) |
| Khon Kaen | 125 | 91 (72.80) | 1 (0.80) | 0 (0.00) |
| Bueng Kan | 105 | 83 (79.05) | 12 (11.43) | 0 (0.00) |
| Sisaket | 94 | 38 (40.43) | 0 (0.00) | 0 (0.00) |
| Total | 445 | 290 (65.17) | 14 (3.15) | 7 (1.57) |
| Province | No. Examined | Prevalence of Strongyloides sp. Detected by APC and FECT (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| Total of Positive (APC+ FECT) | APC Positive | FECT Positive | Both APC and FECT Positive (Similar Results) | Only APC Positive but Negative by FECT | Only FECT Positive but Negative by APC | Ratio (APC: FECT) | ||
| Loei | 121 | 78 (64.46) | 75 (61.98) | 48 (39.67) | 45 (37.19) | 30 (24.79) | 3 (2.48) | 1.6 |
| Khon Kaen | 125 | 91 (72.80) | 87 (69.60) | 64 (51.20) | 60 (48.00) | 27 (21.60) | 4 (3.20) | 1.4 |
| Bueng Kan | 105 | 83 (79.05) | 81 (77.14) | 51 (48.57) | 49 (46.67) | 32 (30.48) | 2 (1.90) | 1.6 |
| Sisaket | 94 | 38 (40.43) | 33 (35.11) | 21 (22.34) | 16 (17.02) | 17 (18.09) | 5 (5.32) | 1.6 |
| Total | 445 | 290 (65.17) | 276 (62.02) | 184 (41.35) | 170 (38.20) | 106 (23.82) | 14 (3.15) | 1.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuasit, T.; Yingklang, M.; Janwan, P.; Maleewong, W.; Saijuntha, W.; Kuanamon, S.; Thanchomnang, T. Gastrointestinal Parasitic Infections in Macaca fascicularis in Northeast Thailand: A One Health Perspective on Zoonotic Risks. Animals 2025, 15, 2112. https://doi.org/10.3390/ani15142112
Kuasit T, Yingklang M, Janwan P, Maleewong W, Saijuntha W, Kuanamon S, Thanchomnang T. Gastrointestinal Parasitic Infections in Macaca fascicularis in Northeast Thailand: A One Health Perspective on Zoonotic Risks. Animals. 2025; 15(14):2112. https://doi.org/10.3390/ani15142112
Chicago/Turabian StyleKuasit, Teputid, Manachai Yingklang, Penchom Janwan, Wanchai Maleewong, Weerachai Saijuntha, Siriporn Kuanamon, and Tongjit Thanchomnang. 2025. "Gastrointestinal Parasitic Infections in Macaca fascicularis in Northeast Thailand: A One Health Perspective on Zoonotic Risks" Animals 15, no. 14: 2112. https://doi.org/10.3390/ani15142112
APA StyleKuasit, T., Yingklang, M., Janwan, P., Maleewong, W., Saijuntha, W., Kuanamon, S., & Thanchomnang, T. (2025). Gastrointestinal Parasitic Infections in Macaca fascicularis in Northeast Thailand: A One Health Perspective on Zoonotic Risks. Animals, 15(14), 2112. https://doi.org/10.3390/ani15142112