
Environmental Factors Modulate Feeding Behavior of Penaeus vannamei: Insights from Passive Acoustic Monitoring


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Collection and Maintenance
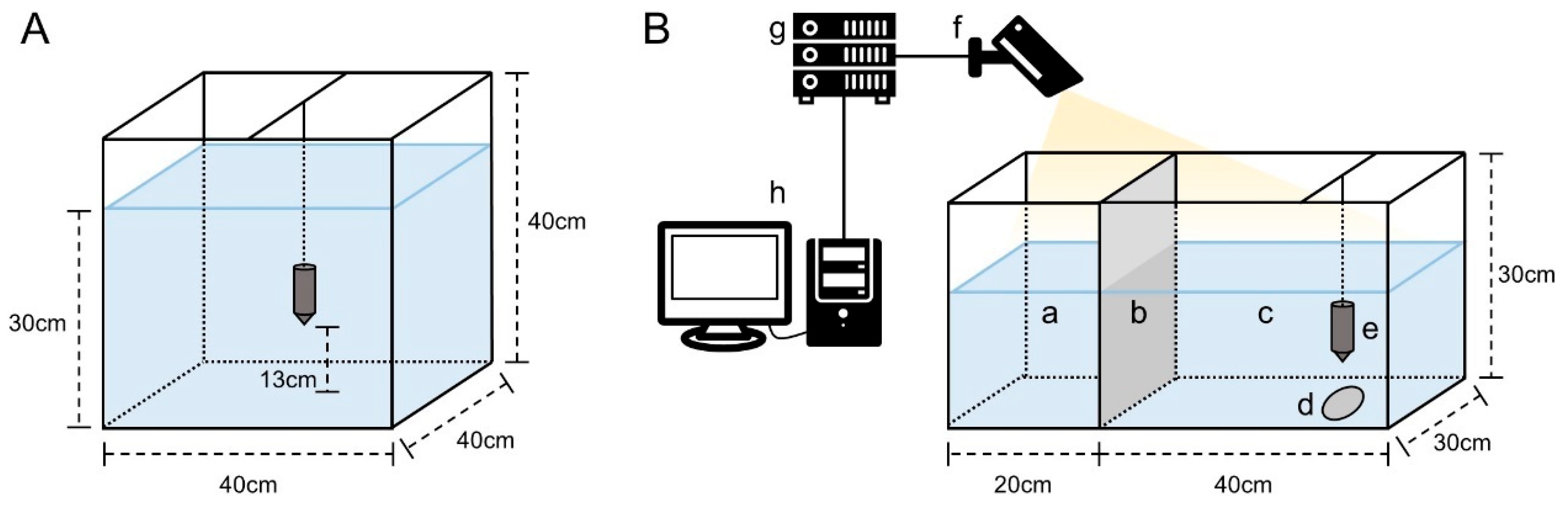
2.2. Experimental Design
2.2.1. Gradient Settings for Temperature, Ammonia Nitrogen, and Nitrite Nitrogen
2.2.2. Acquisition of Acoustic Feeding Signals and Calculation of Feed Consumption
2.2.3. Video Recording of Feeding Behavior
2.3. Data Processing
2.3.1. Audio Processing
2.3.2. Video Processing
2.4. Data Analysis
3. Results
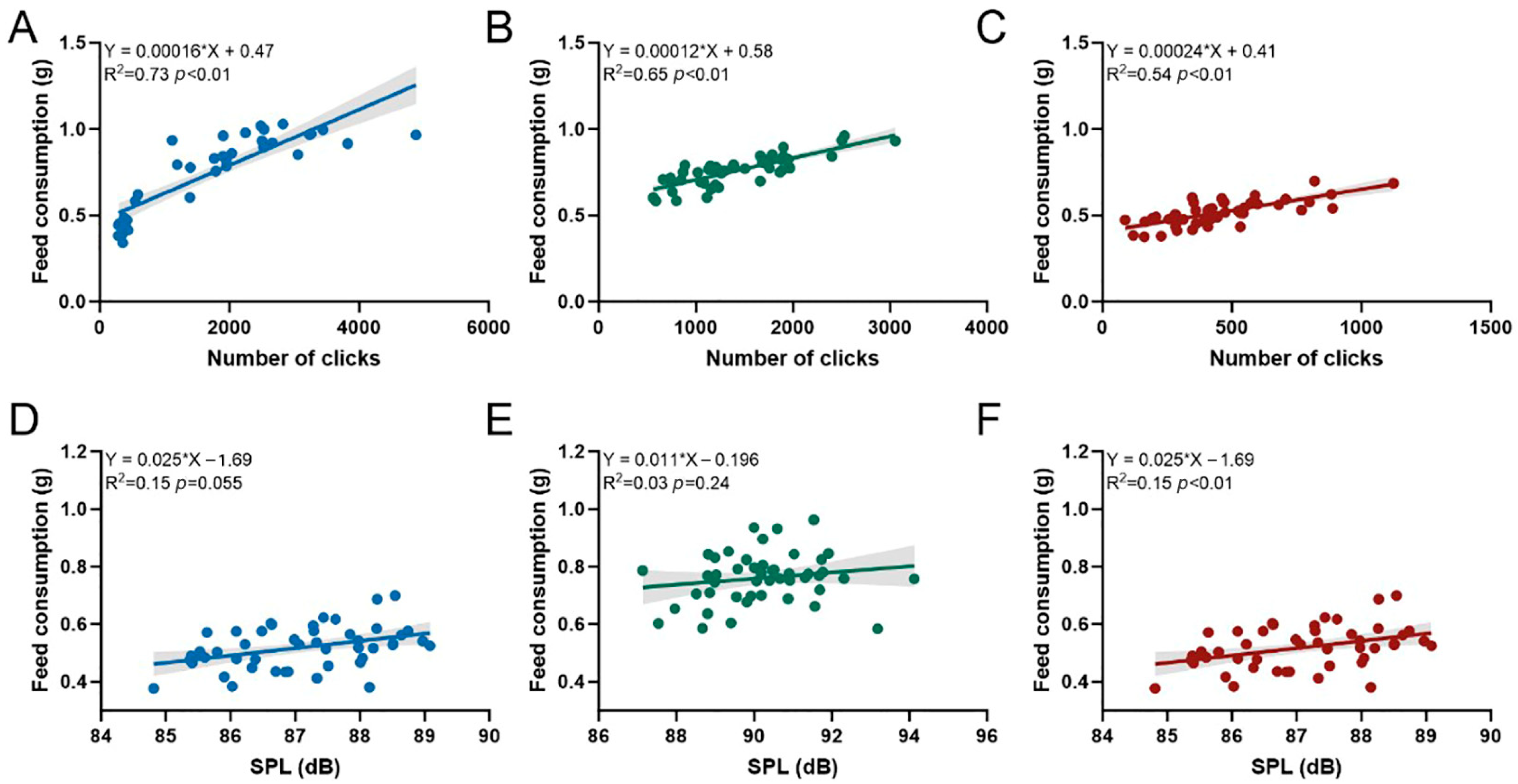
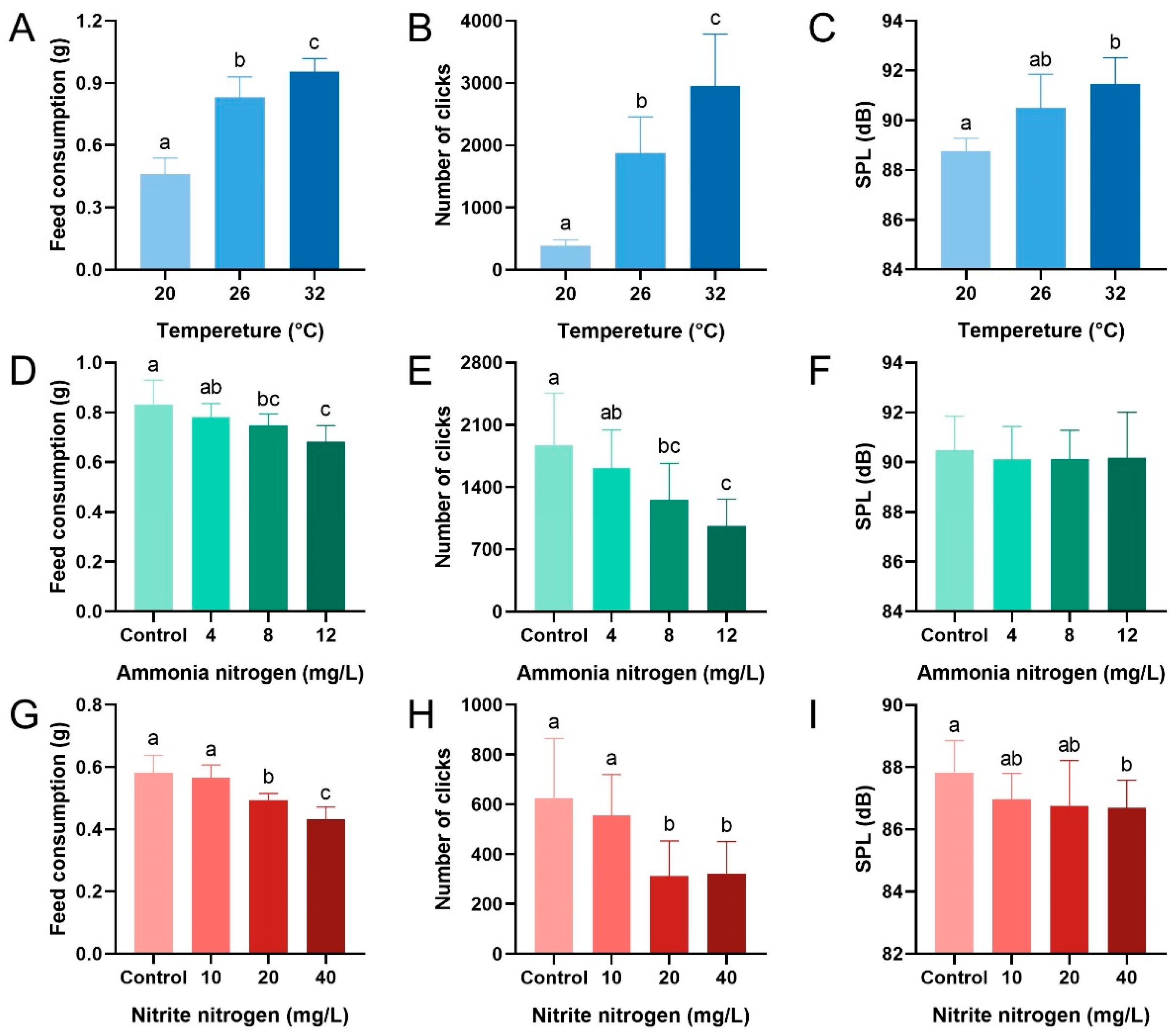
3.1. Effects of Environmental Factors on Feed Consumption and Acoustic Feeding Signals of P. vannamei
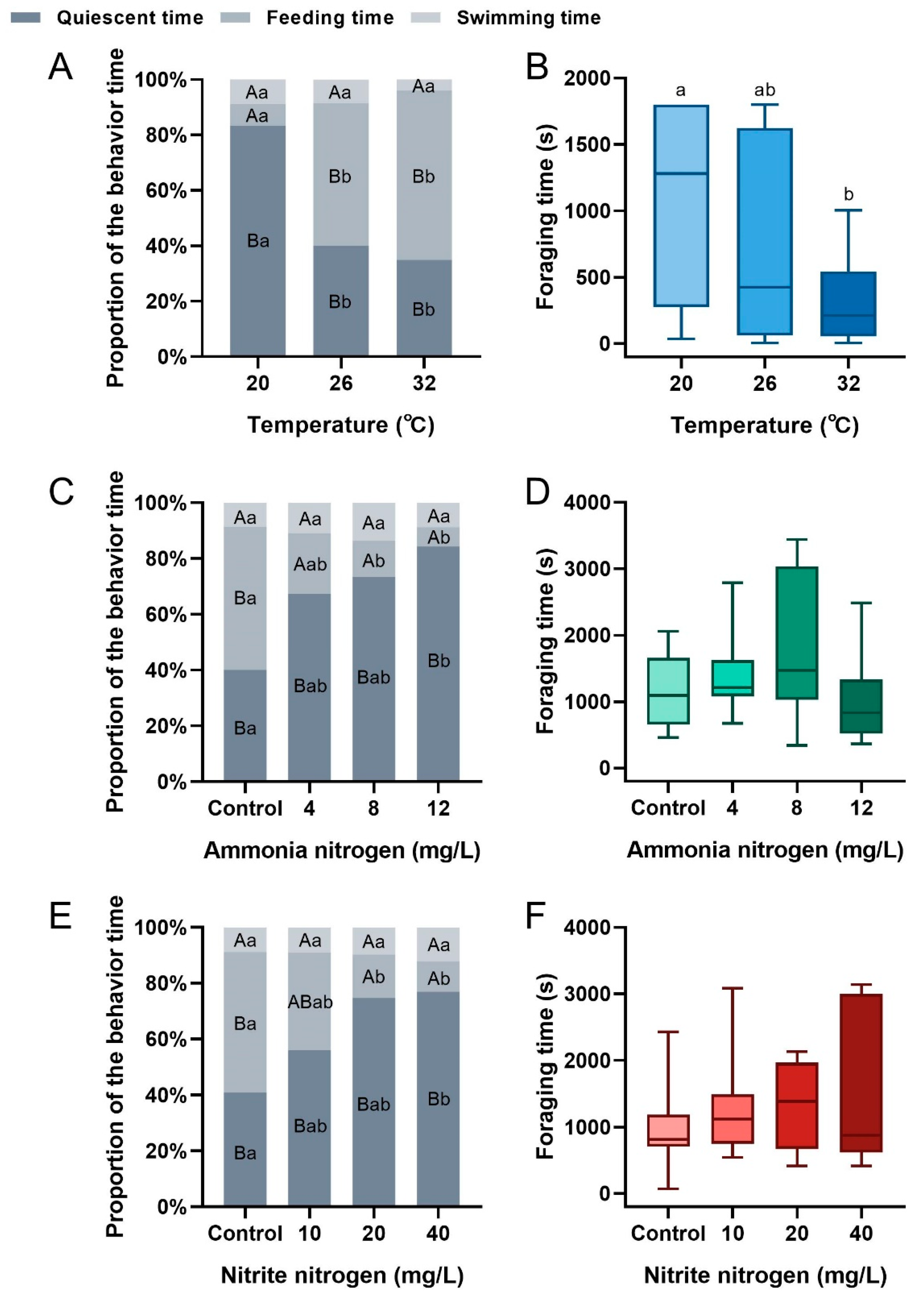
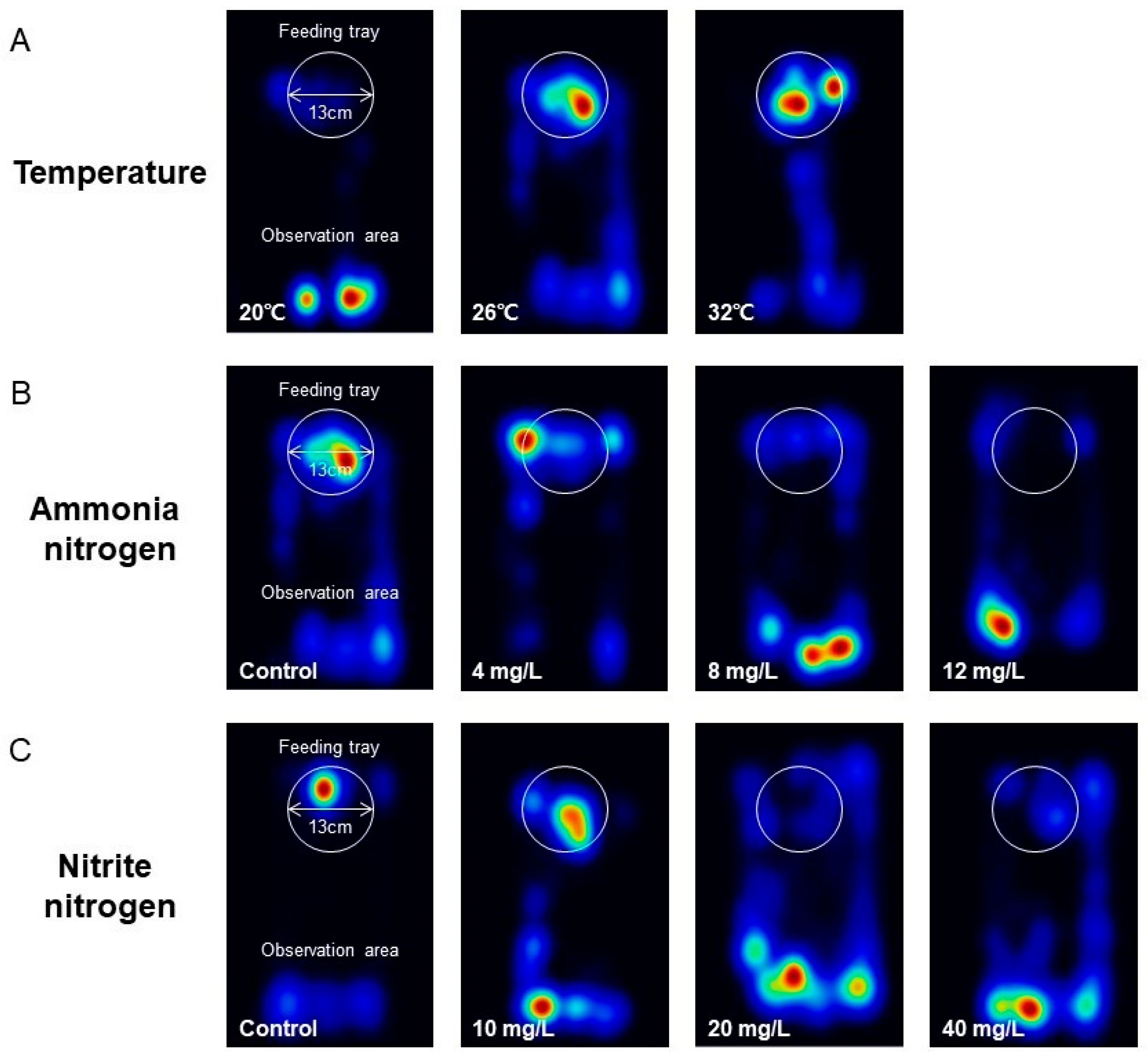
3.2. The Influence of Environmental Factors on the Feeding Behavior of P. vannamei
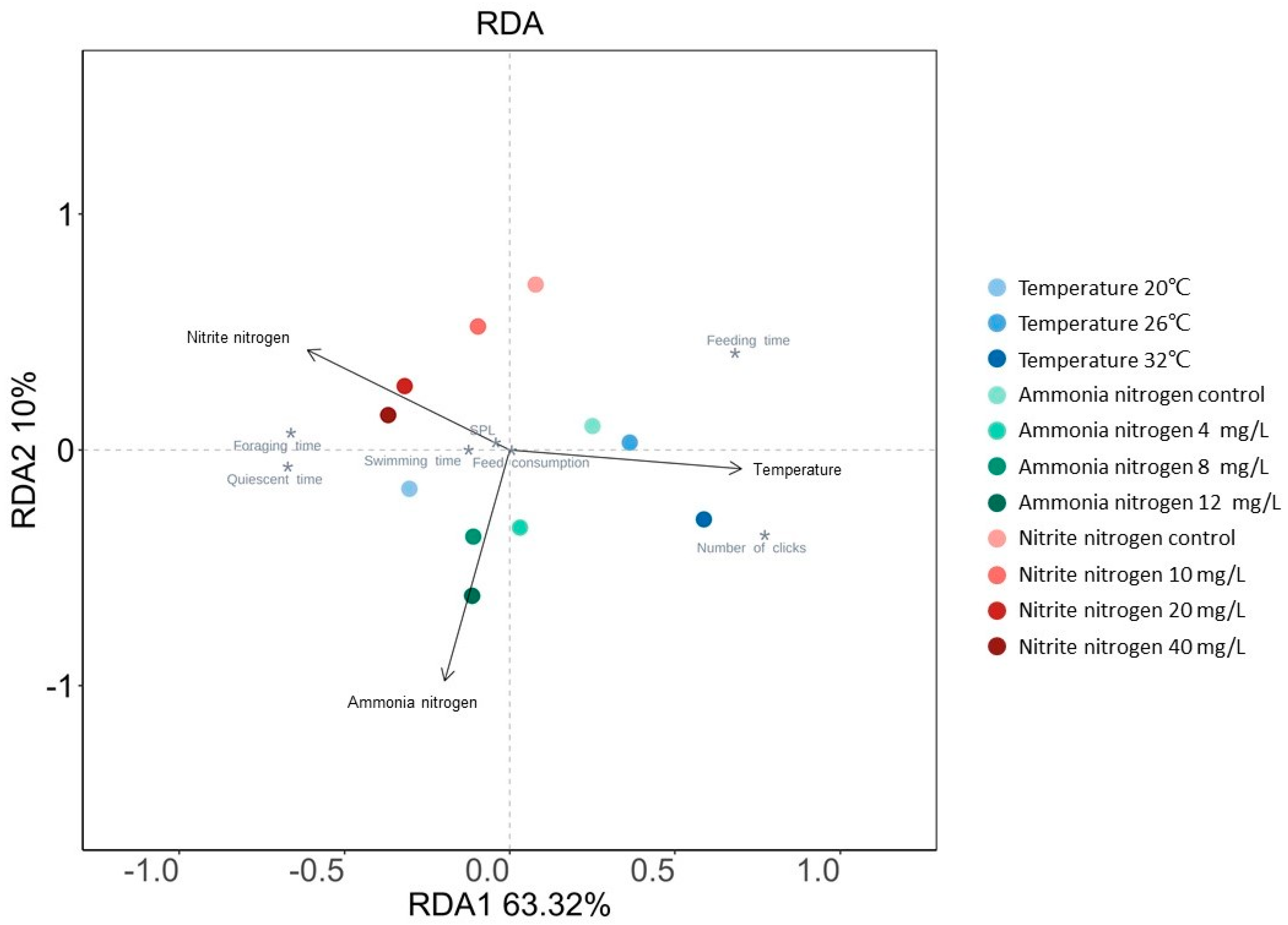
3.3. Comprehensive Analysis of Environmental Factors and Their Biological Response Variables on the Feeding Behavior of P. vannamei
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2024; Food and Agriculture Organization of the United Nations: Rome, Italy, 2024. [Google Scholar]
- Emerenciano, M.G.C.; Rombenso, A.N.; Vieira, F.d.N.; Martins, M.A.; Coman, G.J.; Truong, H.H.; Noble, T.H.; Simon, C.J. Intensification of Penaeid Shrimp Culture: An Applied Review of Advances in Production Systems, Nutrition and Breeding. Animals 2022, 12, 236. [Google Scholar] [CrossRef] [PubMed]
- Biao, X.; Kaijin, Y. Shrimp Farming in China: Operating Characteristics, Environmental Impact and Perspectives. Ocean Coast. Manag. 2007, 50, 538–550. [Google Scholar] [CrossRef]
- Davis, D.; Ullman, C.; Rhodes, M.; Novriadi, R.; Swanepoel, A. Automated Feeding Systems in Pond Production of Pacific White Shrimp. Global Aquaculture Advocate. 2018. Available online: https://www.globalseafood.org/advocate/automated-feeding-systems-in-pond-production-of-pacific-white-shrimp/ (accessed on 1 May 2025).
- Cai, K.; Yang, Z.; Gao, T.; Liang, M.; Liu, P.; Zhou, S.; Pang, H.; Liu, Y. Efficient Recognition of Fish Feeding Behavior: A Novel Two-stage Framework Pioneering Intelligent Aquaculture Strategies. Comput. Electron. Agric. 2024, 224, 109129. [Google Scholar] [CrossRef]
- Zhou, C.; Xu, D.; Lin, K.; Sun, C.; Yang, X. Intelligent Feeding Control Methods in Aquaculture with an Emphasis on Fish: A Review. Rev. Aquac. 2017, 10, 975–993. [Google Scholar] [CrossRef]
- Li, D.; Du, Z.; Wang, Q.; Wang, J.; Du, L. Recent Advances in Acoustic Technology for Aquaculture: A Review. Rev. Aquac. 2023, 16, 357–381. [Google Scholar] [CrossRef]
- Peixoto, S.; Soares, R. Recent Advances and Applications of Passive Acoustic Monitoring in Assessing Shrimp Feeding Behaviour Under Laboratory and Farm Conditions. Rev. Aquac. 2024, 17, e12978. [Google Scholar] [CrossRef]
- Silva, J.F.; Hamilton, S.; Rocha, J.V.; Borie, A.; Travassos, P.; Soares, R.; Peixoto, S. Acoustic Characterization of Feeding Activity of Litopenaeus vannamei in Captivity. Aquaculture 2019, 501, 76–81. [Google Scholar] [CrossRef]
- Zhu, B.; Wang, Z.; Li, Y.; Shan, H.; Liu, D.; Dong, S.; Han, X.; Wang, F. Morphological and Structural Analysis of Penaeus vannamei Mandibles and an Attempt at Real-time Cannibalism Monitoring Based on Passive Acoustics. Aquac. Rep. 2024, 37, 102199. [Google Scholar] [CrossRef]
- Darodes de Tailly, J.B.; Keitel, J.; Owen, M.A.G.; Alcaraz-Calero, J.M.; Alexander, M.E.; Sloman, K.A. Monitoring Methods of Feeding Behaviour to Answer Key Questions in Penaeid Shrimp Feeding. Rev. Aquac. 2021, 13, 1828–1843. [Google Scholar] [CrossRef]
- Ullman, C.; Rhodes, M.; Hanson, T.; Cline, D.; Davis, D.A. A New Paradigm for Managing Shrimp Feeding. World Aquac. 2017, 48, 30–34. [Google Scholar]
- Ullman, C.; Rhodes, M.A.; Allen Davis, D. Feed Management and the Use of Automatic Feeders in the Pond Production of Pacific white shrimp Litopenaeus vannamei. Aquaculture 2019, 498, 44–49. [Google Scholar] [CrossRef]
- Reis, J.; Novriadi, R.; Swanepoel, A.; Jingping, G.; Rhodes, M.; Davis, D.A. Optimizing Feed Automation: Improving Timer-feeders and on Demand Systems in Semi-intensive Pond Culture of Shrimp Litopenaeus vannamei. Aquaculture 2020, 519, 734759. [Google Scholar] [CrossRef]
- Bardera, G.; de Tailly, J.-B.D.; de Fátima Arruda, M.; Pontes, C.S. Shrimp Feeding Behaviour. In The Shrimp Book II; CABI GB: New York, NY, USA, 2021; pp. 336–356. [Google Scholar]
- Hou, D.; Li, H.; Wang, S.; Weng, S.; He, J. Ammonia Nitrogen Stress Induces Dysbiosis of the Intestinal Bacterial Community and Facilitates the Enrichment of Pathogenic Bacteria in Intestines of Shrimp. Aquaculture 2025, 595, 741510. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, J.; Wang, Y.; Liu, Q.; Xiong, D. Nitrite Stress Disrupts the Structural Integrity and Induces Oxidative Stress Response in the Intestines of Pacific White Shrimp Litopenaeus vannamei. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2018, 329, 43–50. [Google Scholar] [CrossRef]
- Huang, W.; Ren, C.; Li, H.; Huo, D.; Wang, Y.; Jiang, X.; Tian, Y.; Luo, P.; Chen, T.; Hu, C. Transcriptomic Analyses on Muscle Tissues of Litopenaeus vannamei Provide the First Profile Insight into the Response to Low Temperature Stress. PLoS ONE 2017, 12, e0178604. [Google Scholar] [CrossRef] [PubMed]
- Wyban, J.; Walsh, W.A.; Godin, D.M. Temperature Effects on Growth, Feeding Rate and Feed Conversion of the Pacific White Shrimp (Penaeus vannamei). Aquaculture 1995, 138, 267–279. [Google Scholar] [CrossRef]
- Zhu, J.; Shi, W.; Zhao, R.; Gu, C.; Li, H.; Wang, L.; Wan, X. Effects of Cold Stress on the Hemolymph of the Pacific White Shrimp Penaeus vannamei. Fishes 2024, 9, 36. [Google Scholar] [CrossRef]
- Manoj, M.; Dhilip Kumar, V.; Arif, M.; Bulai, E.-R.; Bulai, P.; Geman, O. State of the Art Techniques for Water Quality Monitoring Systems for Fish Ponds Using IoT and Underwater Sensors: A Review. Sensors 2022, 22, 2088. [Google Scholar] [CrossRef]
- Bardera, G.; Usman, N.; Owen, M.; Pountney, D.; Sloman, K.A.; Alexander, M.E. The Importance of Behaviour in Improving the Production of Shrimp in Aquaculture. Rev. Aquac. 2018, 11, 1104–1132. [Google Scholar] [CrossRef]
- Shen, G.; Shi, B. Marine Ecology; Xiamen University Press: Xiamen, China, 2002. (In Chinese) [Google Scholar]
- Bardera, G.; Owen, M.A.G.; Façanha, F.N.; Alcaraz-Calero, J.M.; Alexander, M.E.; Sloman, K.A. The Influence of Density and Dominance on Pacific White Shrimp (Litopenaeus vannamei) Feeding Behaviour. Aquaculture 2021, 531, 735949. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. Available online: https://www.canoco.com/ (accessed on 5 October 2023).
- Peixoto, S.; Strebel, L.; Soares, R.; Davis, D.A. Acoustic Feeding Responses Using Marine Chemoattractants in Plant-based Diets for Naive and Non-naive Litopenaeus vannamei. Appl. Anim. Behav. Sci. 2022, 257, 105792. [Google Scholar] [CrossRef]
- Reis, J.; Hussain, A.S.; Weldon, A.; Walsh, S.; Stites, W.; Rhodes, M.; Davis, D.A. Passive Acoustic Feeders as a Tool to Assess Feed Response and Growth in Shrimp Pond Production. Aquac. Int. 2023, 31, 1643–1657. [Google Scholar] [CrossRef]
- Tabbara, M.; Strebel, L.; Peixoto, S.; Soares, R.; Morais, S.; Davis, D.A. Use of Passive Acoustic Monitoring to Evaluate the Effects of a Feed Effector on Feeding Behavior, Growth Performance, and Salinity Stress Tolerance of Litopenaeus vannamei. Aquaculture 2024, 582, 740499. [Google Scholar] [CrossRef]
- Peixoto, S.; Soares, R.; Silva, J.F.; Hamilton, S.; Morey, A.; Davis, D.A. Acoustic Activity of Litopenaeus vannamei Fed Pelleted and Extruded Diets. Aquaculture 2020, 525, 735307. [Google Scholar] [CrossRef]
- Soares, R.; Peixoto, S.; Galkanda-Arachchige, H.S.C.; Davis, D.A. Growth Performance and Acoustic Feeding Behavior of Two Size Classes of Litopenaeus vannamei Fed Pelleted and Extruded Diets. Aquac. Int. 2021, 29, 399–415. [Google Scholar] [CrossRef]
- Hamilton, S.; Filho, F.C.; Silva, J.F.; Duarte-Neto, P.J.; Soares, R.; Peixoto, S. The Loud Crowd: Interactions Between Stocking Density and Acoustic Feeding Activity of Different Size Classes of Litopenaeus vannamei. Aquaculture 2023, 563, 738904. [Google Scholar] [CrossRef]
- Ponce-Palafox, J.; Martinez-Palacios, C.A.; Ross, L.G. The Effects of Salinity and Temperature on the Growth and Survival Rates of Juvenile White Shrimp, Penaeus vannamei, Boone, 1931. Aquaculture 1997, 157, 107–115. [Google Scholar] [CrossRef]
- Walker, S.J.; Neill, W.H.; Lawrence, A.L.; Gatlin, D.M. Effects of Temperature and Starvation on Ecophysiological Performance of the Pacific White Shrimp (Litopenaeus vannamei). Aquaculture 2011, 319, 439–445. [Google Scholar] [CrossRef]
- Xu, H.; Liu, H.; Lin, Y. Effect of Temperature and Salinity on Respiration of Mantis Shrimp (Oratosquilla oratoria). Fish. Sci. 2008, 9, 443–446. (In Chinese) [Google Scholar]
- Ching, C.; Limsuwan, C. Temperature Affects Feeding Behaviour of Pacific White Shrimp. Global Aquaculture Advocate, 2012. Available online: https://www.aquaculturealliance.org/advocate/temperature-affects-feeding-behavior-pacific-white-shrimp/?headlessPrint=AAAAAPIA9c8r7gs82oW (accessed on 1 May 2025).
- Chatvijitkul, S.; Boyd, C.E.; Davis, D.A. Nitrogen, Phosphorus, and Carbon Concentrations in Some Common Aquaculture Feeds. J. World Aquac. Soc. 2017, 49, 477–483. [Google Scholar] [CrossRef]
- Ortega, V.A.; Renner, K.J.; Bernier, N.J. Appetite-suppressing Effects of Ammonia Exposure in Rainbow Trout Associated with Regional and Temporal Activation of Brain Monoaminergic and CRF Systems. J. Exp. Biol. 2005, 208, 1855–1866. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liang, X.; He, S.; Li, L. Effects of Long-term Low-concentration Nitrite Exposure and Detoxification on Growth Performance, Antioxidant Capacities, and Immune Responses in Chinese Perch (Siniperca chuatsi). Aquaculture 2021, 533, 736123. [Google Scholar] [CrossRef]
- Fregoso-López, M.G.; Morales-Covarrubias, M.S.; Franco-Nava, M.A.; Ramírez-Rochín, J.; Fierro-Sañudo, J.F.; Ponce-Palafox, J.T.; Páez-Osuna, F. Histological Alterations in Gills of Shrimp Litopenaeus vannamei in Low-salinity Waters under Different Stocking Densities: Potential Relationship with Nitrogen Compounds. Aquac. Res. 2017, 48, 5854–5863. [Google Scholar] [CrossRef]
- Li, Z.; Ma, S.; Shan, H.; Wang, T.; Xiao, W. Responses of Hemocyanin and Energy Metabolism to Acute Nitrite Stress in Juveniles of the Shrimp Litopenaeus vannamei. Ecotoxicol. Environ. Saf. 2019, 186, 109753. [Google Scholar] [CrossRef]
- Zhao, M.; Aweya, J.J.; Feng, Q.; Zheng, Z.; Yao, D.; Zhao, Y.; Chen, X.; Zhang, Y. Ammonia Stress Affects the Structure and Function of Hemocyanin in Penaeus vannamei. Ecotoxicol. Environ. Saf. 2022, 241, 113827. [Google Scholar] [CrossRef]
- da Costa, F.P.; Gomes, B.S.F.d.F.; Pereira, S.D.d.N.A.; de Fátima Arruda, M. Influence of Stocking Density on the Behaviour of Juvenile Litopenaeus vannamei (Boone, 1931). Aquac. Res. 2016, 47, 912–924. [Google Scholar] [CrossRef]
- Kramer, D.L. Dissolved oxygen and fish behavior. Environ. Biol. Fishes 1987, 18, 81–92. [Google Scholar] [CrossRef]
- Israeli-Weinstein, D.; Kimmel, E.J.A. Behavioral Response of Carp (Cyprinus carpio) to Ammonia Stress. Aquaculture 1998, 165, 81–93. [Google Scholar] [CrossRef]
- Carrillo-Farnes, O.; Forrellat-Barrios, A.; Guerrero-Galvan, S.; Vega-Villasante, F.J.C. A Review of Digestive Enzyme Activity in Penaeid Shrimps. Crustaceana 2007, 80, 257–275. [Google Scholar] [CrossRef]
- Kır, M.; Sunar, M.C.; Topuz, M.; Sarıipek, M. Thermal Acclimation Capacity and Standard Metabolism of the Pacific White Shrimp Litopenaeus vannamei (Boone, 1931) at Different Temperature and Salinity Combinations. J. Therm. Biol. 2023, 112, 103429. [Google Scholar] [CrossRef]
- Camargo, J.A.; Alonso, A.; Salamanca, A. Nitrate Toxicity to Aquatic Animals: A Review with New Data for Freshwater Invertebrates. Chemosphere 2005, 58, 1255–1267. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Liu, Q.; Du, J.; Zhu, W.; Li, Q.; Chen, X.; Chen, X.; Liu, H.; Zhou, X.; Zhao, Y.; et al. Integrated Analysis of Physiological, Transcriptomic and Metabolomic Responses and Tolerance Mechanism of Nitrite Exposure in Litopenaeus vannamei. Sci. Total Environ. 2020, 711, 134416. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, L.; Dai, X. Combined Effects of Ammonia Nitrogen, Nitrite, Salinity, and Temperature Negatively Impact the Growth, Survival, Physiological, and Biochemical Parameters, and Hepatopancreatic Structure of Litopenaeus vannamei. Aquaculture 2025, 596, 741845. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavioral Component | Description |
|---|---|
| Feeding time | The duration (in seconds) that P. vannamei spends feeding within the feeding tray. In EthoVision, this is defined as “the time spent within the observation area (i.e., the feeding tray).” |
| Quiescent time | The duration (in seconds) during which P. vannamei remains stationary during the experiment. |
| Swimming time | The duration (in seconds) that P. vannamei continuously swims during the experiment. |
| Foraging time | The duration (in seconds) taken by P. vannamei to first enter the feeding tray. In EthoVision, this is defined as “the time of first entry into the observation area.” |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Yang, C.; Li, Y.; Ma, B.; Zhu, B. Environmental Factors Modulate Feeding Behavior of Penaeus vannamei: Insights from Passive Acoustic Monitoring. Animals 2025, 15, 2113. https://doi.org/10.3390/ani15142113
Zhang H, Yang C, Li Y, Ma B, Zhu B. Environmental Factors Modulate Feeding Behavior of Penaeus vannamei: Insights from Passive Acoustic Monitoring. Animals. 2025; 15(14):2113. https://doi.org/10.3390/ani15142113
Chicago/Turabian StyleZhang, Hanzun, Chao Yang, Yesen Li, Bin Ma, and Boshan Zhu. 2025. "Environmental Factors Modulate Feeding Behavior of Penaeus vannamei: Insights from Passive Acoustic Monitoring" Animals 15, no. 14: 2113. https://doi.org/10.3390/ani15142113
APA StyleZhang, H., Yang, C., Li, Y., Ma, B., & Zhu, B. (2025). Environmental Factors Modulate Feeding Behavior of Penaeus vannamei: Insights from Passive Acoustic Monitoring. Animals, 15(14), 2113. https://doi.org/10.3390/ani15142113