
Complex Body Wall Closure Defects in Seven Dog Fetuses: An Anatomic and CT Scan Study

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
- Initial examination and fixation: each fetus was examined macroscopically and then fixed in 5% formaldehyde for preservation.
- Anthropometric measurements: body weight was recorded using a digital precision scale, and body length was measured using a flexible tape.
- Gross anatomical evaluation: standard dissection techniques were employed to assess and document structural malformations across multiple anatomical systems.
- Radiographic imaging: radiographs were taken using a Sedecal (Algete, Madrid, Spain) 20 kW three-phase high-frequency X-ray unit equipped with a Toshiba Rotanode E7239X F 2 mm–1 mm X-ray tube (Otawara, Japan) and an Optimus DR direct digital detector panel.
- Computed tomography (CT) analysis: high-resolution CT scans were performed using a Toshiba Aquilion 64 multislice CT scanner (350 mA, 120 kV, 512 × 512 matrix, 5.5 cm field of view). Images were acquired with a bone reconstruction algorithm (window level 550, window width 2550, slice thickness 0.5 mm, pitch 0.4 mm).
- 3D visualization: CT data were reconstructed in three dimensions to enhance identification and analysis of complex congenital anomalies.
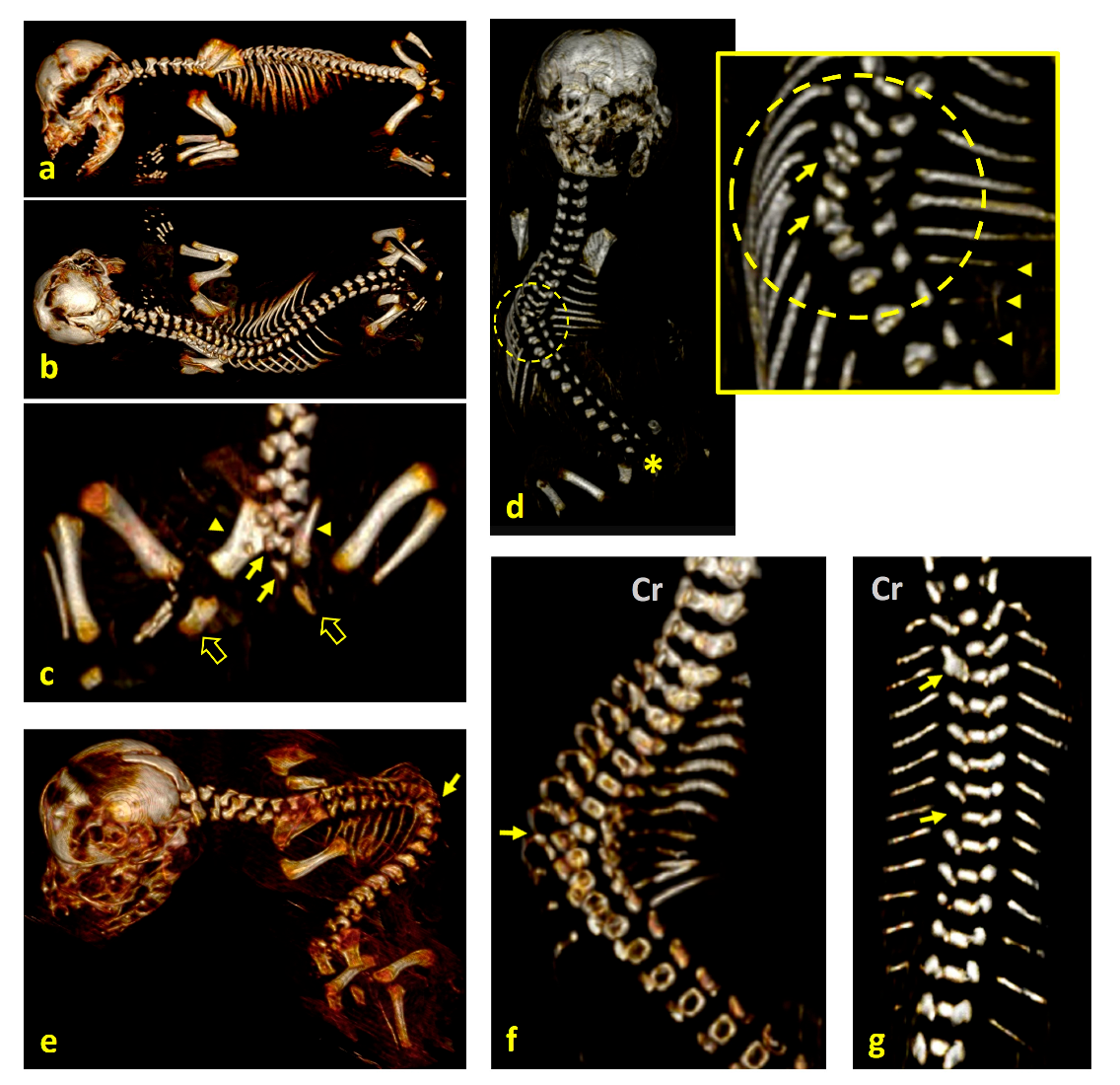
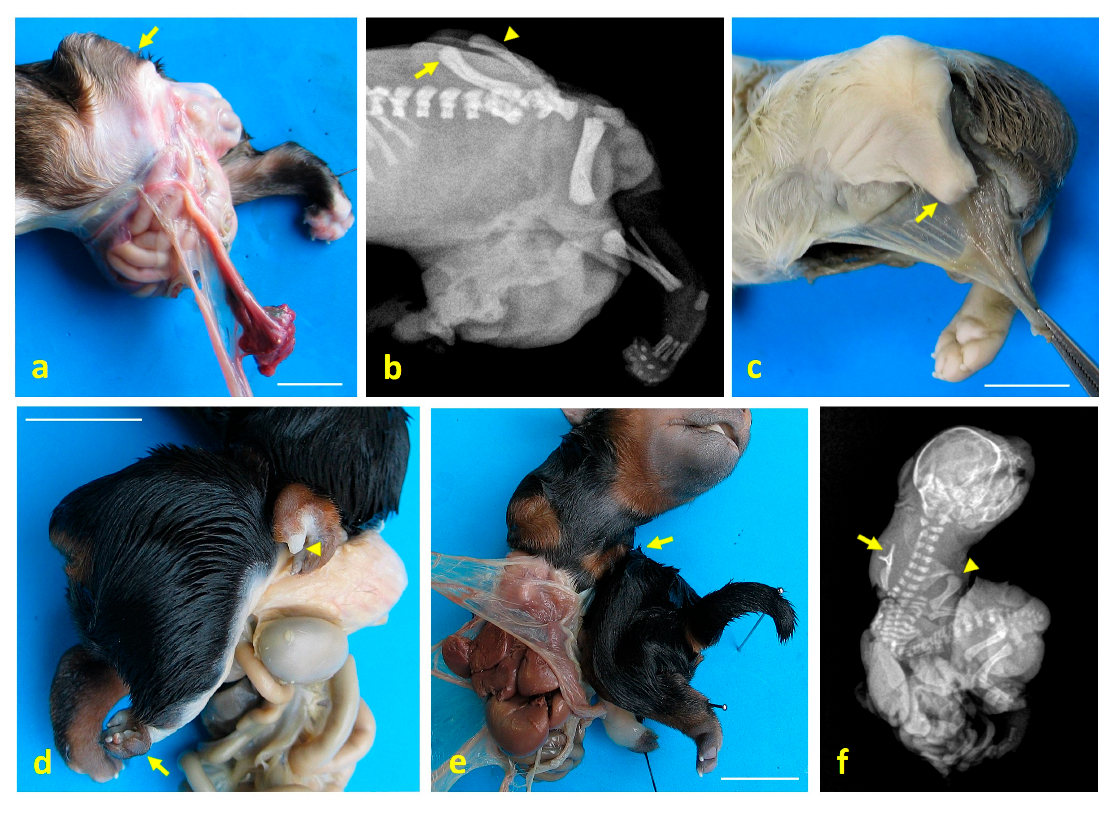
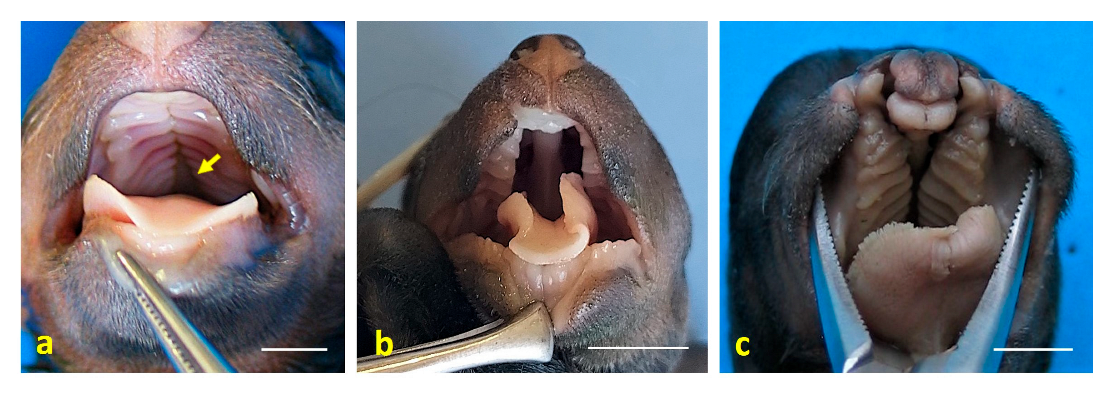
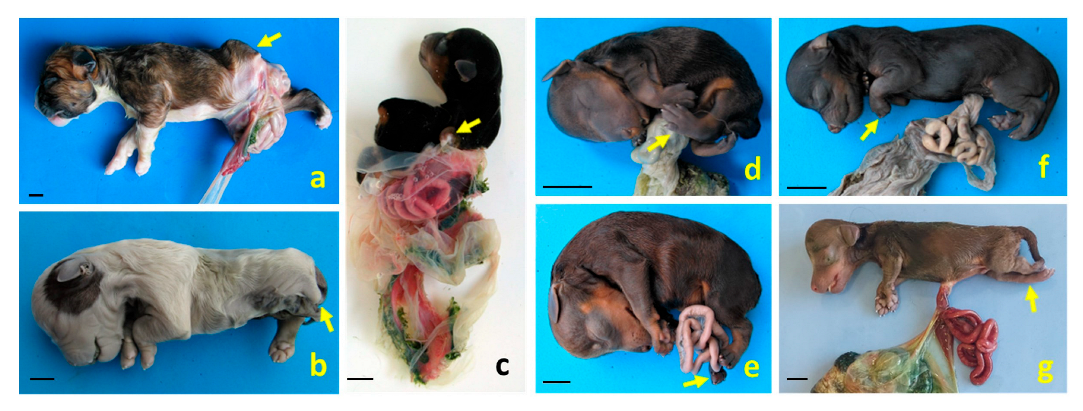
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Anal atresia |
| AB | Amniotic band |
| Ab | Abdominoschisis |
| ABDC | Amniotic band disruption complex |
| ArCTV | Arthrogryposis in the cervical and thoracic vertebrae |
| AI | Artificial insemination |
| ArPL | Arthrogryposis in the pelvic limb |
| ArLTL | Arthrogryposis in the left thoracic limb |
| ArRPL | Arthrogryposis in the right pelvic limb |
| ARTL | Amelia of the right thoracic limb |
| ASA | Axial skeletal anomaly |
| BCH | Bilateral cheiloschisis |
| BSA | Body stalk anomaly |
| BWD | Body wall defect |
| CBWA | Complex body wall anomaly |
| CFA | Craniofacial anomalies |
| ChPa | Cheilognathopalatoschisis |
| CP | Complete palatoschisis |
| CRL | Crown-rump length |
| DUV | Dispersed umbilical vessels |
| ES | Extreme scoliosis |
| HC | Hydrocephalus |
| HLUA | Hypoplastic left umbilical artery |
| HV | Hemivertebrae |
| JLE | Jejunal loop eventration |
| K | Kyphosis |
| LA | Limb anomaly |
| l-CDLAB | Left caudolateral abdominoschisis |
| l-CLO | Left caudolateral omphalocele |
| LLRF | Left lateral retroflexion |
| MLPL | Meromelia of the left pelvic limb |
| MLTL | Meromelia of the left thoracic limb |
| MS | Mild scoliosis |
| PRPL | Phocomelia of the right pelvic limb |
| O | Omphalocele |
| r-Lab | Right lateral abodominoschisis |
| S | Scoliosis |
| SPBWC | Spinal–body wall complex |
| SPLBWC | Spinal–limb–body wall complex |
| SSBWC | Sternal–spinal–body wall complex |
| STBWC | Sternal–body wall complex |
| SUA | Single umbilical artery |
| UC | Umbilical cord |
| UCA | Umbilical cord anomaly |
| UGA | Urogenital anomaly |
References
- Martín-Alguacil, N.; Avedillo, L. Body wall defects and amniotic band syndrome in pig (Sus scrofa domesticus). Anat. Histol. Embryol. 2020, 49, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, I.; Winn, H.N.; Hobbins, J.C. Prenatal diagnostic criteria for body stalk anomaly. Am. J. Perinatol. 1989, 6, 84–85. [Google Scholar] [CrossRef] [PubMed]
- Martín-Alguacil, N. Anatomy-based diagnostic criteria for complex body wall anomalies (CBWA). Mol. Genet. Genom. Med. 2020, 8, e1465. [Google Scholar] [CrossRef] [PubMed]
- Lockwood, C.J.; Scioscia, A.L.; Hobbins, J.C. Congenital absence of the umbilical cord resulting from maldevelopment of embryonic body folding. Am. J. Obstet. Gynecol. 1986, 155, 1049–1051. [Google Scholar] [CrossRef]
- Paul, C.; Zosmer, N.; Jurkovic, D.; Nicolaides, K. A case of body stalk anomaly at 10 weeks of gestation. Ultrasound Obstet. Gynecol. 2001, 17, 157–159. [Google Scholar] [CrossRef]
- Zeidler, S.; Oudesluijs, G.G.; Schoonderwaldt, E.M.; Van Bever, Y. Early prenatal disruption; a foetus with features of severe limb body wall sequence, body stalk anomaly and amniotic bands. J. Genet. Couns. 2014, 25, 315–320. [Google Scholar] [PubMed]
- Martín-Alguacil, N.; Avedillo, L. Body stalk anomalies in pig—Definition and classification. Mol. Genet. Genom. Med. 2020, 13, e1227. [Google Scholar] [CrossRef]
- Martín-Alguacil, N.; Cozar, J.M.; Avedillo, L. Body Stalk Anomalies in Pigs: Current Trends and Future Directions in Classification. Animals 2025, 15, 460. [Google Scholar] [CrossRef]
- Martín-Alguacil, N.; Cozar, J.M.; Avedillo, L. Body Stalk Anomalies and their Relationship to Amniotic Band Disruption Complex in Six Cats. JFMS 2025, 27, 1098612X251341068. [Google Scholar] [CrossRef]
- Noakes, D.E.; Parkinson, T.J.; England, G.C. (Eds.) Arthur’s Veterinary Reproduction and Obstetrics; Elsevier Health Sciences: Amsterdam, The Netherlands, 2018; pp. 168–171. [Google Scholar]
- Ginsberg, N.E.; Cadkin, A.; Strom, C. Prenatal diagnosis of body stalk anomaly in the first trimester of pregnancy. Ultrasound Obst. Gynecol. 1997, 10, 419–421. [Google Scholar] [CrossRef]
- Stein, W.; Haller, F.; Hawighorst, T.; Emons, G. Pentalogy of Cantrell vs. Limb Body Wall Complex: Differential Diagnosis of a Severe Malformation in Early Pregnancy. Ultraschall Med. 2009, 30, 598–601. [Google Scholar] [CrossRef]
- Routhu, M.; Thakkallapelli, S.; Mohan, P.; Ahmed, N. Role of Ultrasound in Body Stalk Anomaly and Amniotic Band Syndrome. Int. J. Reprod. Med. 2016, 2016, 3974139. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Singh, J.; Gupta, K. Body stalk anomaly: Antenatal sonographic diagnosis of this rare entity with review of literature. J. Ultrason. 2017, 17, 133–135. [Google Scholar] [CrossRef]
- Patten, R.M.; Van Allen, M.; Mack, L.A.; Wilson, D.; Nyberg, D.; Hirsch, J.; Viamont, T. Limb-Body Wall Complex: In Utero Sonographic Diagnosis of a Complicated Fetal Malformation. Am. J. Roentgenol. 1986, 146, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Moerman, P.; Fryns, J.-P.; Vandenberghe, K.; Lauweryns, J.M. Constrictive Amniotic Bands, Amniotic Adhesions, and Limb-Body Wall Complex: Discrete Disruption Sequences with Pathogenetic Overlap. Am. J. Med. Genet. 1992, 42, 470–479. [Google Scholar] [CrossRef]
- Pumberger, W.; Schaller, A.; Bernaschek, G. Limb-body wall complex: A compound anomaly pattern in body-wall defects. Pediatr. Surg. Int. 2001, 17, 486–490. [Google Scholar] [CrossRef]
- Das, S.K.; Maharana, S.S.; Subudhi, M.; Rao, P.V. The Limb-Abdominal Wall Complex Defects, a form of Amniotic Band Syndrome: A Rare Case Report. J. Clin. Diagn. Res. 2013, 7, 543–544. [Google Scholar] [PubMed]
- Hartwig, N.G.; Vermeij-Keers, C.; de Vries, H.E.; Kagie, M.; Kragt, H. Limb Body Wall Malformation Complex: An Embryologic Etiology? Hum. Pathol. 1989, 20, 1071–1077. [Google Scholar] [CrossRef]
- Palacios, J.; Rodriguez, J.I. Limb Body Wall Malformation Complex Associated with Vascular Steal. Hum. Pathol. 1990, 21, 875–876. [Google Scholar] [CrossRef]
- Gulczyński, J.; Świątkowska-Freund, M.; Paluchowski, P.; Hermann-Okoniewska, B.; Iżycka-Świeszewska, E. Limb body wall complex—The history of the entity and presentation of our series of cases. Pol. J. Pathol. 2019, 70, 33–41. [Google Scholar] [CrossRef]
- Van Allen, M.I.; Curry, C.; Walden, C.E.; Gallagher, L.; Patten, R.M. Limb-Body Wall Complex: II. Limb and Spine Defects. Am. J. Med. Genet. 1987, 28, 549–565. [Google Scholar] [CrossRef]
- Craven, C.M.; Carey, J.C.; Ward, K. Umbilical Cord Agenesis in Limb Body Wall Defect. Am. J. Med. Genet. 1997, 71, 97–105. [Google Scholar] [CrossRef]
- Sahinoglu, Z.; Uludogan, M.; Arik, H.; Aydin, A.; Kucukbas, M.; Bilgic, R.; Toksoy, G. Prenatal Ultrasonographical Features of Limb Body Wall Complex: A Review of Etiopathogenesis and a New Classification. Fetal Pediatr. Pathol. 2007, 26, 135–151. [Google Scholar] [CrossRef]
- Mathai, A.M.; Menezes, R.G.; Kumar, S.; Pai, M.R.; Bhandary, A.; Fitzhugh, V.A. A fetal autopsy case of body stalk anomaly. Leg. Med. 2009, 11, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Gajzer, D.C.; Hirzel, A.C.; Saigal, G.; Rojas, C.P.; Rodriguez, M.M. Possible Genetic Origin of Limb-Body Wall Complex. Fetal Pediatr. Pathol. 2015, 34, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Martín-Alguacil, N.; Avedillo, L. Body Wall Defects: Gastroschisis and Omphalocoele in Pigs (Sus scrofa domesticus). J. Comp. Pathol. 2020, 175, 69–74. [Google Scholar] [CrossRef]
- Hyttel, P.; Sinowatz, F.; Vejlsted, M. Essentials of Domestic Animals Embryology; Betteridge, K., Ed.; Saunders Ltd.: Southampton, UK, 2009. [Google Scholar]
- Hunter, A.G.W.; Seaver, L.H.; Stevenson, R.E. Limb–body wall defect. Is there a defensible hypothesis and can it explain all the associated anomalies? Am. J. Med. Genet. Part A 2011, 155, 2045–2059. [Google Scholar] [CrossRef] [PubMed]
- Laughton, K.W.; Fisher, K.R.; Halina, W.G.; Partlow, G.D. Schistosomus reflexus syndrome: A heritable defect in ruminants. AHE 2005, 34, 312–318. [Google Scholar] [CrossRef]
- Notter, C. Schistosome in pigs. Virchows Arch. Pathol. Anat. Physiol. Klin. Med. 1927, 264, 280–298. [Google Scholar] [CrossRef]
- Mateo, I.; Camón, J. Schistosoma reflexum in a cat: Insights into aetiopathogenesis. JFMS 2008, 10, 376–379. [Google Scholar] [CrossRef]
- Ozalp, G.R.; Celikler, S.; Simsek, G.; Ozyigit, M.O.; Inan, S.A. A case of schistosoma reflexum in a cat with chromosomal aberrations. Reprod. Domest. Anim. 2011, 46, 373–376. [Google Scholar] [CrossRef] [PubMed]
- Nalluri, H.; Leela, V.; Sirikonda, P. Origin, course and associated congenital anomalies of type 2 single umbilical artery: A fetal anatomic study. Int. J. Anat. Res. 2016, 4, 2041–2046. [Google Scholar] [CrossRef]
- Monie, I. Genesis of single umbilical artery. Am. J. Obstet. Gynecol. 1970, 108, 400–405. [Google Scholar] [CrossRef]
- Martín-Alguacil, N.; Avedillo, L. Cantrell Syndrome (Thoracoabdominal Ectopia Cordis; Anomalous Umbilical Cord; Diaphragmatic, Pericardial and Intracardiac Defects) in the Pig (Sus scrofa domesticus). J. Comp. Pathol. 2020, 174, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Gaikwad, S.M.; Gulavane, S.U.; Kumbhar, U.B.; Shelar, R.R.; Chaudhari, R.J.; Ribeiro, R.A. Doppler evaluation of maternal vessels in normal gestation and threatened abortion in canines. Ir. Vet. J. 2020, 73, 15. [Google Scholar] [CrossRef] [PubMed]
- Estevam, M.V.; Toniollo, G.H.; Apparicio, M. The most common congenital malformations in dogs: Literature review and practical guide. Res. Vet. Sci. 2024, 171, 105230. [Google Scholar] [CrossRef]
- Macrì, F.; Marino, F.; Rapisarda, G.; Lanteri, G.; Mazzullo, G. A case of unilateral pelvic limb Adactyly in a puppy dog. AHE. 2010, 40, 104–106. [Google Scholar] [CrossRef]
- Macri, F.; Rapisarda, G.; Lanteri, G.; Di Pietro, S.; Auriemma, E.; Marino, F. Congenital absence of humerus with preaxial terminal longitudinal hemimelia and hypoplasia of the scapula in a dog: A case report. Vet. Med. 2014, 59, 506–510. [Google Scholar] [CrossRef]
- Macri, F.; Di Pietro, S.; Piccionello, A.P.; Rapisarda, G.; Lanteri, G.; Angileri, V.; Marino, F. A rare case of partial paraxial radial hemimelia in a puppy. Vet. Med. 2017, 12, 681–684. [Google Scholar] [CrossRef]
- Barrios, N.; Gómez, M.; Mieres, M.; Vera, F.; Alvial, G. Spinal dermoid sinus in a dachshund with vertebral and thoracic limb malformations. BMC Vet. Res. 2014, 10, 54–60. [Google Scholar] [CrossRef]
- Ortega-Pacheco, A.; Lezama-García, M.A.; Colín-Flores, R.; Jiménez-Coello, M.; Acevedo-Arcique, C.; Gutiérrez-Blanco, E. Presence of congenital anomalies in three dog litters. Reprod. Domest. Anim. 2020, 55, 652–655. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Frías, M. Epidemiological characteristics of amniotic band sequence (ABS) and body wall complex (BWC): Are they two different entities? Am. J. Med. Genet. 1997, 73, 176–179. [Google Scholar] [CrossRef]
- Sundaram, V.; Mohammed, T.; Rampersad, S.N.; Williams, G. A rare report on 18-month survival of a dog born with multiple anomalies including atresia ani. Morphologie 2021, 105, 252–258. [Google Scholar] [CrossRef]
- Switonski, M.; Dzimira, S.; Aleksiewicz, R.; Szczerbal, I.; Nowacka-Woszuk, J.; Krzeminska, P.; Deska, T.; Nizanski, W. Hypospadias is not rare in dogs: Five new cases, a retrospective study, and a review of the literature. Sex. Dev. 2018, 12, 244–250. [Google Scholar] [CrossRef]
- Bae, S.W.; Chung, D.J.; Chung, W.H.; Park, N.W.; Lim, C.Y.; Kim, H.Y. A rare case of a complex of multiple congenital anomalies diagnosed using computed tomography in a male puppy. Vet. Med. 2018, 63, 50–53. [Google Scholar] [CrossRef]
- Bouty, A.; Ayers, K.L.; Pask, A.; Heloury, Y.; Sinclair, A.H. The genetic and environmental factors underlying hypospadias. Sex. Dev. 2015, 9, 239–259. [Google Scholar] [CrossRef]
- Roman, N.; Carney, P.C.; Fiani, N.; Peralta, S. Incidence patterns of orofacial clefts in purebred dogs. PLoS ONE 2019, 14, e0224574. [Google Scholar] [CrossRef]
- Peralta, S.; Fiani, N.; Kan-Rohrer, K.H.; Verstraete, F.J.M. Morphological evaluation of clefts of the lip, palate, or both in dogs. Am. J. Vet. Res. 2017, 78, 926–933. [Google Scholar] [CrossRef]
- Wolf, Z.T.; Brand, H.A.; Shaffer, J.R.; Leslie, E.J.; Arzi, B.; Willet, C.E.; Cox, T.C.; McHenry, T.; Narayan, N.; Feingold, E.; et al. Genome-wide association studies in dogs and humans identify ADAMTS20 as a risk variant for cleft lip and palate. PLoS Genet. 2015, 11, e1005059. [Google Scholar] [CrossRef]
- Gică, N.; Apostol, L.M.; Huluță, I.; Panaitescu, A.M.; Vayna, A.M.; Peltecu, G.; Gana, N. Body Stalk Anomaly. Diagnostics 2024, 14, 518. [Google Scholar] [CrossRef]
- Rowsell, A. The amniotic band disruption complex. The pathogenesis of oblique facial clefts; an experimental study in the foetal rat. Br. J. Plast. Surg. 1989, 42, 291–295. [Google Scholar] [CrossRef]
- Stelnicki, E.; Hoffman, W.; Vanderwall, K.; Harrison, M.; Foster, R.; Longaker, M. A New In Utero Model for Lateral Facial Clefts. J. Craniofac. Surg. 1997, 8, 460–465. [Google Scholar] [CrossRef]
- Eppley, B.; David, L.; Li, M.; Moore, C.; Sadove, A. Amniotic Band Facies. J. Craniofac. Surg. 1998, 9, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Morovic, C.; Berwart, F.; Varas, J. Craniofacial Anomalies of the Amniotic Band Syndrome in Serial Clinical Cases. Plast. Reconstr. Surg. 2004, 113, 1556–1562. [Google Scholar] [CrossRef]
- Weinstein, B.; Hassouba, M.; Flores, R.L.; Staffenberg, D.A.; Gordon, C.B.; Runyan, C.M. Digital-Facial Translocation in Amniotic Band Sequence: Evidence of the Intrinsic Theory. J. Craniofac. Surg. 2018, 29, 1890–1892. [Google Scholar] [CrossRef] [PubMed]
- Miyajima, K.; Natsume, N.; Kawai, T.; Iizuka, T. Oblique facial cleft, cleft palate, and supernumerary teeth secondary to amniotic bands. Cleft Palate Craniofac. J. 1994, 31, 483–486. [Google Scholar] [CrossRef]
- Rasul, M.; Fujiwara, K.; Ruslin, M.; Astuti, I.; Takaichi, M.; Noguchi, M. Amniotic Band Syndrome with Severe Facial Cleft: A Case Report. Cleft Palate Craniofac. J. 2023, 62, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Taub, P.; Bradley, J.; Setoguchi, Y.; Schimmenti, L.; Kawamoto, H. Typical facial clefting and constriction band anomalies: An unusual association in three unrelated patients. Am. J. Med. Genet. Part A 2003, 120A, 256–260. [Google Scholar] [CrossRef]
- Chen, H.; Chen, C.; Hsu, C.; Wang, W. Typical body wall defect associated with craniofacial anomalies and amniotic bands diagnosed in early pregnancy. Taiwan. J. Obstet. Gynecol. 2007, 46, 286–287. [Google Scholar] [CrossRef]
- Rita, D.; Pushpalatha, V.S. A rare case report of amniotic band syndrome associated with gastroschisis and limb body wall complex. Int. J. Reprod. Contracept. Obstet. Gynecol. 2023, 12, 1944–1946. [Google Scholar] [CrossRef]
- Halder, A. Amniotic band syndrome and/or limb body wall complex: Split or lump. Appl. Clin. Genet. 2010, 3, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, H.; Dhungel, P.; Harper, T.; Lampley, C. Amniotic Band Syndrome: A Case Series. J. Preg. Child Health 2014, 1, 1. [Google Scholar] [CrossRef]
- Miller, M.; Graham, J.; Higginbottom, M.; Smith, D. Compression-related defects from early amnion rupture: Evidence for mechanical teratogenesis. J. Pediat. 1981, 98, 292–297. [Google Scholar] [CrossRef]
- Robin, N.; Franklin, J.; Prucka, S.; Ryan, A.B.; Grant, J.H. Clefting, amniotic bands, and polydactyly: A distinct phenotype that supports an intrinsic mechanism for amniotic band sequence. Am. J. Med. Genet. Part A 2005, 137A, 298–301. [Google Scholar] [CrossRef]
- Guion-Almeida, M.L.; Richieri-Costa, A. Amniotic band sequence versus the autosomal recessive microcephaly, facial clefting, and preaxial polydactyly syndrome. Clin. Dysmorphol. 2000, 9, 297–299. [Google Scholar] [CrossRef]
- Saritha, S.; Gouri, G.; Sumangala, S. Limb body wall complex or body stalk complex or cyllosomas: A case report. Int. J. Res. Med. Sci. 2013, 1, 132–137. [Google Scholar] [CrossRef]
- Di Pietro, S.; Rapisarda, G.S.; Cicero, L.; Angileri, V.; Morabito, S.; Cassata, G.; Macrì, F. Four Unusual Cases of Congenital Forelimb Malformations in Dogs. Animals 2021, 11, 813. [Google Scholar] [CrossRef] [PubMed]
- Cornillie, P.; Van Lancker, S.; Simoens, P. Two cases of brachymelia in cats. AHE 2004, 33, 115–118. [Google Scholar] [CrossRef]
- Di Dona, F.; Valle, G.D.; Meomartino, L.; Lamagna, F.; Fatone, G. Congenital deformity of the distal extremities in three dogs. Open Vet. J. 2016, 6, 228–233. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Weight (g) | CRL (cm) | BWD (cm) | Parturition | Breed | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Type | Mother’s Age (years) | Litter Size | Gestation Time | Maternal Parity | Mating | |||||
| C 1 ♀ | 280 | 13.5 | 2.9 × 2.9 | C-section | 3 | 6 | 59 | P | Natural | Pitbull |
| C 2 ♂ | 125 | 9.8 | 1.1 ×2.9 | Natural delivery | 2 | 7 | 60 | M | Natural | Medium-sized mongrel dog |
| C 3 ♂ | 105 | 8.2 | 2.9 × 3 | C-section | 2 | 3 | 59 | P | Natural | Yorkshire terrier |
| C 4 ♂ | 85 | 7.5 | 0.6 × 0.6 | C-section | 3 | 6 | 60 | M | Natural | Teckel |
| C 5 ♂ | 62 | 11.4 | 1.5 × 0.9 | Dystocia | 4 | 3 | 60 | P | Natural | Yorkshire terrier |
| C 6 ♂ | 60 | 9.5 | 0.4 × 0.4 | C-section | 2.5 | 4 | 61 | UNK | AI | Pincher |
| C 7 ♂ | 105 | 10.1 | 0.5 × 0.5 | Natural delivery | 6 | 5 | 60 | P | Natural | Mongrel dog (teckel-like) |
| Case | BWD | ASA | UCA | LA | AA | CFA | UGA | AB | Diagnosis |
|---|---|---|---|---|---|---|---|---|---|
| C 1 ♀ | l-CLO Exposure to the exterior of the spleen, liver along with the gallbladder, pancreas, part of the stomach, and all the intestine except the descending colon and the rectum | S Hypoplasia of S1 and S2, agenesis of S3 and caudal vertebrae. Hypoplasia of both the ilium and the ischium and agenesis of both pubes | DUV. SUA | MLPL | + | − | Persistent urogenital sinus. Genital hypoplasia. Left renal agenesis | − | BSA III SPLBWC |
| C 2 ♂ | l-CLAb JLE | S HV at the T5–T7 level, hypoplasia of left ribs 6, 7, and 8 and agenesis of left ribs 10 to 13. Agenesis of sacral and caudal vertebrae | DUV HLUA | MLPL | − | − | Hypoplasia of the genital tract and the genital organs | − | BSA III SPLBWC |
| C 3 ♂ | r-LAb External exposure of the kidneys, spleen, liver along with the gallbladder, pancreas, part of the stomach, and all the intestine except the descending colon and the rectum | LLRF ES Vertebral block at the point of maximum flexion of the spine and presence of HV at the thoracolumbar junction. | DUV HLUA | ARTL MRTL PRPL ArRPL | SP | Bilateral renal hipoplasia and genital hypoplasia | − | BSA III SPLBWC | |
| C 4 ♂ | O JLE | K Presence of HV at level T7–T9 corresponding with the point of maximum flexion of the spine. | DUV | ArPL | + | SP Mouse snout | Hypoplasia of the genital tract and the genital organs | Amniotic adhesion in the right ventrolateral portion of the frontal region. Remnants of the amnion attached to the nostrils | BSA IV SPBWC |
| C 5 ♂ | O JLE | ArCTV Agenesis of the sacrum and the caudal vertebrae. | DUV | ArPL The plantar surface of both extremities is directed medially | + | − | Complete hypospadias | − | BSA IV SPBWC |
| C 6 ♂ | O JLE | MS Presence of a vertebral block T2–T3 and HV at T8 level | DUV | ArLTL | − | ChPa | − | BSA VIII SPBWC | |
| C 7 ♂ | O JLE | MS | DUV | ArPL The plantar surface of both extremities is directed dorsally | − | HC | − | BSA VIII SPBWC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-Alguacil, N.; Cozar, J.M.; Avedillo, L. Complex Body Wall Closure Defects in Seven Dog Fetuses: An Anatomic and CT Scan Study. Animals 2025, 15, 2030. https://doi.org/10.3390/ani15142030
Martín-Alguacil N, Cozar JM, Avedillo L. Complex Body Wall Closure Defects in Seven Dog Fetuses: An Anatomic and CT Scan Study. Animals. 2025; 15(14):2030. https://doi.org/10.3390/ani15142030
Chicago/Turabian StyleMartín-Alguacil, Nieves, José M. Cozar, and Luis Avedillo. 2025. "Complex Body Wall Closure Defects in Seven Dog Fetuses: An Anatomic and CT Scan Study" Animals 15, no. 14: 2030. https://doi.org/10.3390/ani15142030
APA StyleMartín-Alguacil, N., Cozar, J. M., & Avedillo, L. (2025). Complex Body Wall Closure Defects in Seven Dog Fetuses: An Anatomic and CT Scan Study. Animals, 15(14), 2030. https://doi.org/10.3390/ani15142030