
Glycine Supplementation Enhances the Growth of Sow-Reared Piglets with Intrauterine Growth Restriction

 ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment 1
2.1.1. Oral Administration of Glycine
2.1.2. Determination of Milk Consumption by Piglets
2.1.3. Collection of Blood Samples
2.1.4. Collection of Skeletal Muscle and Other Tissues from Piglets
2.1.5. Analysis of Amino Acids in Plasma and Tissues
2.1.6. Analysis of Glucose, Urea, Ammonia, and Hormones in Plasma or Serum
2.1.7. Analysis of Creatine, Phosphocreatine, Creatinine, and Guanidinoacetate
2.1.8. Analysis of Reduced Glutathione (GSH) and Oxidized Glutathione (GSSG)
2.1.9. Western Blot Analysis
2.2. Experiment 2
2.3. Statistical Analysis
3. Results
3.1. Experiment 1
3.1.1. Milk Consumption of Piglets
3.1.2. Body Weights and Weight Gains of Piglets
3.1.3. Tissue Weights of Piglets
3.1.4. Concentrations of Amino Acids in Plasma
3.1.5. Concentrations of Amino Acids in Tissues
3.1.6. Concentrations of Glucose, Nitrogenous Metabolites, Lipids, TBARS, and Hormones in Plasma or Serum
3.1.7. Concentrations of Creatine, Phosphocreatine, Creatinine, and Guanidinoacetate in Plasma and Tissues
3.1.8. Concentrations of GSH and GSSG in Plasma and Tissues
3.2. Experiment 2
3.2.1. Rates of Protein Synthesis in Tissues
3.2.2. Proteins in the MTOR Cell Signaling Pathway in Skeletal Muscle
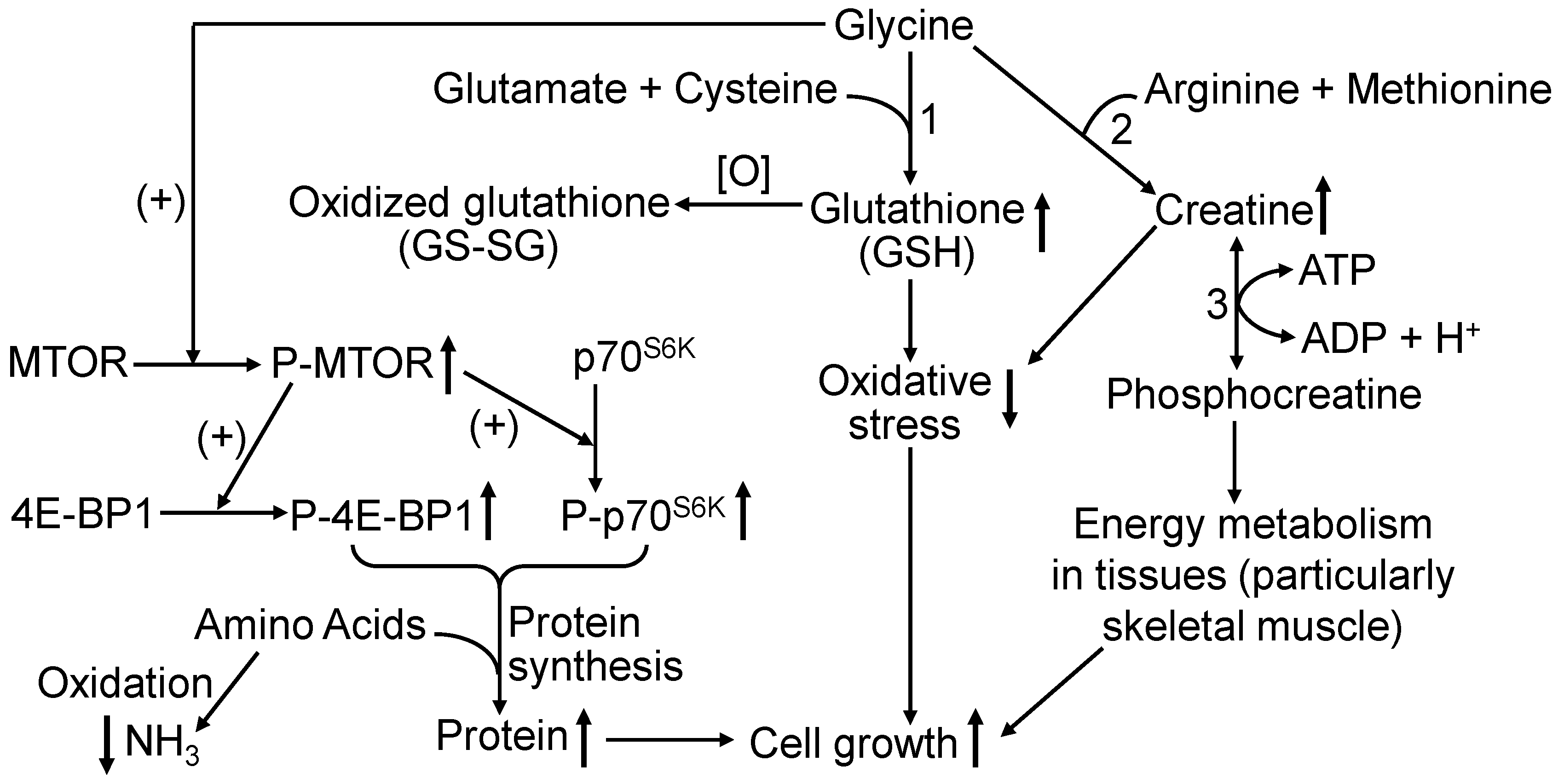
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, G.; Bazer, F.W.; Wallace, J.M.; Spencer, T.E. Intrauterine growth retardation: Implications for the animal sciences. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.D.; He, W.L.; Wu, G. Hydroxyproline in animal metabolism, nutrition, and cell signaling. Amino Acids 2022, 54, 513–528. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.A. The glycine story. Eur. J. Clin. Nutr. 1991, 45, 59–65. [Google Scholar]
- Mudd, S.H.; Brosnan, J.T.; Brosnan, M.E.; Jacobs, R.L.; Stabler, S.P.; Allen, R.H.; Vance, D.E.; Wagner, C. Methyl balance and transmethylation fluxes in humans. Am. J. Clin. Nutr. 2007, 85, 19–25. [Google Scholar] [CrossRef]
- Wang, W.; Wu, Z.; Dai, Z.; Yang, Y.; Wang, J.; Wu, G. Glycine metabolism in animals and humans: Implications for nutrition and health. Amino Acids 2013, 45, 463–477. [Google Scholar] [CrossRef]
- Walker, J.B. Creatine: Biosynthesis, regulation, and function. Adv. Enzymol. 1979, 50, 177–242. [Google Scholar]
- Rose, W.C. The sequence of events leading to the establishment of the amino acid needs of man. Am. J. Public Health 1968, 58, 2020–2027. [Google Scholar] [CrossRef]
- Baker, D.H. Advances in amino acid nutrition and metabolism of swine and poultry. In Nutrient Management of Food Animals to Enhance and Protect the Environment; Kornegay, E.T., Ed.; CRC Press: Boca Raton, FL, USA, 1996; pp. 41–53. [Google Scholar]
- Baker, D.H. Recent advances in use of the ideal protein concept for swine feed formulation. Asian-Aust. J. Anim. Sci. 2000, 13, 294–301. [Google Scholar]
- Reeds, P.J. Dispensable and indispensable amino acids for humans. J. Nutr. 2000, 130, 1835S–1840s. [Google Scholar] [CrossRef]
- Shemin, D. The biological conversion of l-serine to glycine. J. Biol. Chem. 1946, 162, 297–307. [Google Scholar] [CrossRef]
- Chao, F.C.; Delwiche, C.C.; Greenberg, D.M. Biological precursors of glycine. Biochim. Biophys. Acta 1953, 10, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Soloway, S.; Stetten, D., Jr. The metabolism of choline and its conversion to glycine in the rat. J. Biol. Chem. 1953, 204, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Lowry, M.; Hall, D.E.; Brosnan, J.T. Hydroxyproline metabolism by the rat kidney: Distribution of renal enzymes of hydroxyproline catabolism and renal conversion of hydroxyproline to glycine and serine. Metabolism 1985, 34, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Knight, J.; Jiang, J.; Assimos, D.G.; Holmes, R.P. Hydroxyproline ingestion and urinary oxalate and glycolate excretion. Kidney Int. 2006, 70, 1929–1934. [Google Scholar] [CrossRef]
- Hu, S.D.; He, W.L.; Bazer, F.W.; Johnson, G.A.; Wu, G. Synthesis of glycine from 4-hydroxyproline in tissues of neonatal pigs with intrauterine growth restriction. Exp. Biol. Med. 2023, 248, 1446–1458. [Google Scholar] [CrossRef]
- Wang, W.; Dai, Z.; Wu, Z.; Lin, G.; Jia, S.; Hu, S.; Dahanayaka, S.; Wu, G. Glycine is a nutritionally essential amino acid for maximal growth of milk-fed young pigs. Amino Acids 2014, 46, 2037–2045. [Google Scholar] [CrossRef]
- Ashworth, C.J.; Finch, A.M.; Page, K.R.; Nwagwu, M.O.; McArdle, H.J. Causes and consequences of fetal growth retardation in pigs. Reprod. Suppl. 2001, 58, 233–246. [Google Scholar]
- Long, D.W.; Long, B.D.; Nawaratna, G.I.; Wu, G. Oral Administration of L-arginine improves the growth and survival of sow-reared intrauterine growth-restricted piglets. Animals 2025, 15, 550. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Burghardt, R.C.; Johnson, G.A.; Kim, S.W.; Li, X.L.; Satterfield, M.C.; Spencer, T.E. Impacts of amino acid nutrition on pregnancy outcome in pigs: Mechanisms and implications for swine production. J. Anim. Sci. 2010, 88, E195–E204. [Google Scholar] [CrossRef]
- Quiniou, N.; Dagorn, J.; Gaudré, D. Variation of piglets’ birth weight and consequences on subsequent performance. Livest. Prod. Sci. 2002, 78, 63–70. [Google Scholar] [CrossRef]
- Cruz, F.L.; Mendes, M.F.D.S.A.; Silva, T.O.; Filho, M.B.G.; de Abreu, M.L.T. L-Arginine supplementation for pregnant and lactating sows may improve the performance of piglets: A systematic review. J. Anim. Physiol. Anim. Nutr. 2024, 109, 76–95. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Murani, E.; Hadlich, F.; Liu, X.; Wimmers, K.; Ponsuksili, S. Prenatal skeletal muscle transcriptome analysis reveals novel microRNA-mRNA networks associated with intrauterine growth restriction in pigs. Cells 2021, 10, 1007. [Google Scholar] [CrossRef] [PubMed]
- Hales, J.; Moustsen, V.A.; Nielsen, M.B.F.; Hansen, C.F. Individual physical characteristics of neonatal piglets affect preweaning survival of piglets born in a noncrated system. J. Anim. Sci. 2012, 91, 4991–5003. [Google Scholar] [CrossRef] [PubMed]
- Lynegaard, J.C.; Hansen, C.F.; Kristensen, A.R.; Amdi, C. Body composition and organ development of intra-uterine growth restricted pigs at weaning. Animal 2020, 14, 322–329. [Google Scholar] [CrossRef]
- Harper, J.; Bunter, K.L. Improving pig survival with a focus on birthweight: A practical breeding perspective. Animal 2024, 18 (Suppl. 1), 100914. [Google Scholar] [CrossRef]
- Xiong, L.; You, J.; Zhang, W.; Zhu, Q.; Blachier, F.; Yin, Y.L.; Kong, X.F. Intrauterine growth restriction alters growth performance, plasma hormones, and small intestinal microbial communities in growing-finishing pigs. J. Anim. Sci. Biotechnol. 2020, 11, 86. [Google Scholar] [CrossRef]
- Wellington, M.O.; Rodrigues, L.A.; Li, Q.; Dong, B.; Panisson, J.C.; Yang, C.; Columbus, D.A. Birth weight and nutrient restriction affect jejunal enzyme activity and gene markers for nutrient transport and intestinal function in piglets. Animals 2021, 11, 2672. [Google Scholar] [CrossRef]
- Handel, S.E.; Stickland, N.C. The growth and differentiation of porcine skeletal muscle fibre types and the influence of birthweight. J. Anat. 1987, 152, 107–119. [Google Scholar]
- Tang, X.P.; Xiong, K.N. Intrauterine growth retardation affects intestinal health of suckling piglets via altering intestinal antioxidant capacity, glucose uptake, tight junction, and immune responses. Oxid. Med. Cell. Longev. 2022, 2022, 2644205. [Google Scholar] [CrossRef]
- Su, W.; Zhang, H.; Ying, Z.; Li, Y.; Zhou, L.; Wang, F.; Zhang, L.; Wang, T. Effects of dietary L-methionine supplementation on intestinal integrity and oxidative status in intrauterine growth-retarded weanling piglets. Eur. J. Nutr. 2018, 57, 2735–2745. [Google Scholar] [CrossRef]
- Santos, T.G.; Fernandes, S.D.; de Oliveira, S.B. Araújo. Intrauterine growth restriction and its impact on intestinal morphophysiology throughout postnatal development in pigs. Sci. Rep. 2022, 12, 11810. [Google Scholar] [CrossRef] [PubMed]
- Randunu, R.; Alawaini, K.; Huber, L.A.; Randell, E.; Brunton, J.; Bertolo, R. Intrauterine growth-restricted piglets are predisposed to develop metabolic disorders in adulthood when fed with parenteral nutrition in the neonatal period. Curr. Dev. Nutr. 2022, 6 (Suppl 1), 703. [Google Scholar] [CrossRef]
- D’Inca, R.; Kloareg, M.; Gras-Le Guen, C.; Le Huerou-Luron, I. Intrauterine growth restriction modifies the developmental pattern of intestinal structure, transcriptomic profile, and bacterial colonization in neonatal pigs. J. Nutr. 2010, 140, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Hegarty, P.V.; Allen, C.E. Effect of pre-natal runting on the post-natal development of skeletal muscles in swine and rats. J. Anim. Sci. 1978, 46, 1634–1640. [Google Scholar] [CrossRef]
- Nissen, P.M.; Oksbjerg, N. Birth weight and postnatal dietary protein level affect performance, muscle metabolism and meat quality in pigs. Animal 2011, 5, 1382–1389. [Google Scholar] [CrossRef]
- Ji, Y.; Fan, X.; Zhang, Y.; Li, J.; Dai, Z.L.; Wu, Z.L. Glycine regulates mucosal immunity and the intestinal microbial composition in weaned piglets. Amino Acids 2022, 54, 385–398. [Google Scholar] [CrossRef]
- Geiping, L.; Hartmann, M.; Kreienbrock, L.; Beilage, E.G. Killing underweighted low viable newborn piglets: Which health parameters are appropriate to make a decision? Porc. Health Manag. 2022, 8, 25. [Google Scholar] [CrossRef]
- Dalla Costa, F.A.; Gibson, T.J.; Oliveira, S.E.O.; Gregory, N.G.; Faucitano, L.; Dalla Costa, O.A. On-farm culling methods used for pigs. Anim. Welfare 2021, 30, 507–522. [Google Scholar] [CrossRef]
- He, W.L.; Posey, E.A.; Steele, C.C.; Savell, J.W.; Bazer, F.W.; Wu, G. Dietary glycine supplementation enhances post-weaning growth and meat quality of pigs with intrauterine growth restriction. J. Anim. Sci. 2023, 101, skad354. [Google Scholar] [CrossRef]
- Sun, Y.L.; Wu, Z.L.; Li, W.; Zhang, C.; Sun, K.J.; Ji, Y.; Wang, B.; Jiao, N.; He, B.B.; Wang, W.W.; et al. Dietary L-leucine supplementation enhances intestinal development in suckling piglets. Amino Acids 2015, 47, 1517–1525. [Google Scholar] [CrossRef]
- Ji, Y.; Sun, Y.; Liu, N.; Jia, H.; Dai, Z.; Yang, Y.; Wu, Z. L-Leucine supplementation reduces growth performance accompanied by changed profiles of plasma amino acids and expression of jejunal amino acid transporters in breast-fed intra-uterine growth-retarded piglets. Br. J. Nutr. 2023, 129, 2025–2035. [Google Scholar] [CrossRef] [PubMed]
- Jamin, A.; D’Inca, R.; Le Floc’h, N.; Kuster, A.; Orsonneau, J.L.; Darmaun, D.; Boudry, G.; Le Huërou-Luron, I.; Sève, B.; Gras-Le Guen, C. Fatal effects of a neonatal high-protein diet in low-birth-weight piglets used as a model of intrauterine growth restriction. Neonatology 2010, 97, 321–328. [Google Scholar] [CrossRef]
- Posey, E.A.; Davis, T.A. Nutritional regulation of muscle growth in neonatal swine. Animal 2023, 17 (Suppl. 3), 100831. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, J.M.; Niu, X.; Odle, J.; Graves, L.M. Role of mTOR signaling in intestinal cell migration. Am. J. Physiol. 2006, 291, G510–G517. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liao, S.F. Physiological effects of dietary amino acids on gut health and functions of swine. Front. Vet. Sci. 2019, 6, 169. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements of Swine; National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Rezaei, R.; San Gabriel, A.; Wu, G. Dietary supplementation with monosodium glutamate enhances milk production by lactating sows and the growth of suckling piglets. Amino Acids 2022, 54, 1055–1068. [Google Scholar] [CrossRef]
- Draper, H.H.; Squires, E.J.; Mahmoodi, H.; Wu, J.; Agarwal, S.; Hadley, M. A comparative evaluation of thiobarbituric acid methods for the determination of malondialdehyde in biological materials. Free Radic. Biol. Med. 1993, 15, 353–363. [Google Scholar] [CrossRef]
- He, W.L.; Li, X.Y.; Wu, G. Dietary glycine supplementation enhances syntheses of creatine and glutathione by tissues of hybrid striped bass (Morone saxatilis ♀× Morone chrysops ♂) fed soybean meal-based diets. J. Anim. Sci. Biotechnol. 2024, 15, 67. [Google Scholar] [CrossRef]
- Rezaei, R.; Wu, G. Branched-chain amino acids regulate intracellular protein turnover in porcine mammary epithelial cells. Amino Acids 2022, 54, 1491–1504. [Google Scholar] [CrossRef]
- Frank, J.W.; Escobar, J.; Hguyen, H.V.; Jobgen, S.C.; Jobgen, W.S.; Davis, T.A.; Wu, G. Oral N-carbamylglutamate supplementation increases protein synthesis in skeletal muscle of piglets. J. Nutr. 2007, 137, 315–319. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics: A Biometrical Approach, 2nd ed.; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Kim, S.W.; Wu, G. Dietary arginine supplementation enhances the growth of milk-fed young pigs. J. Nutr. 2004, 134, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Mansilla, W.D.; Saraswathy, S.; Garcia-Ruiz, A.I. Dietary protein reduction with stepwise addition of crystalline amino acids and the effect of considering a minimum glycine-serine content in broiler diets. Poult. Sci. 2023, 102, 102684. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sun, K.; Ji, Y.; Wu, Z.; Wang, W.; Dai, Z.; Wu, G. Glycine regulates expression and distribution of claudin-7 and ZO-3 proteins in intestinal porcine epithelial cells. J. Nutr. 2016, 146, 964–969. [Google Scholar] [CrossRef]
- Sun, K.; Wu, Z.; Ji, Y.; Wu, G. Glycine regulates protein turnover by activating Akt/mTOR and by inhibiting MuRF1 and atrogin-1 gene expression in C2C12 myoblasts. J. Nutr. 2016, 146, 2461–2467. [Google Scholar] [CrossRef]
- Ham, D.J.; Caldow, M.K.; Chhen, V.; Chee, A.; Wang, X.; Proud, C.G.; Lynch, G.S.; Koopman, R. Glycine restores the anabolic response to leucine in a mouse model of acute inflammation. Am. J. Physiol. 2016, 310, E970–E981. [Google Scholar] [CrossRef]
- Koopman, R.; Caldow, M.K.; Ham, D.J.; Lynch, G.S. Glycine metabolism in skeletal muscle: Implications for metabolic homeostasis. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 237–242. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Wang, X.; Wu, H.; Chen, S.; Zhu, H.; Zhang, J.; Hou, Y.; Hu, C.A.; Zhang, G. Glycine enhances muscle protein mass associated with maintaining Akt-mTOR-FOXO1 signaling and suppressing TLR4 and NOD2 signaling in piglets challenged with LPS. Am. J. Physiol. 2016, 311, R365–R373. [Google Scholar] [CrossRef]
- Ost, M.; Keipert, S.; van Schothorst, E.M.; Donner, V.; van der Stelt, I.; Kipp, A.P.; Petzke, K.J.; Jove, M.; Pamplona, R.; Portero-Otin, M.; et al. Muscle mitohormesis promotes cellular survival via serine/glycine pathway flux. FASEB J. 2015, 29, 1314–1328. [Google Scholar] [CrossRef]
- Wang, W.; Wu, Z.; Lin, G.; Hu, S.; Wang, B.; Dai, Z.; Wu, G. Glycine stimulates protein synthesis and inhibits oxidative stress in pig small intestinal epithelial cells. J. Nutr. 2014, 144, 1540–1548. [Google Scholar] [CrossRef]
- Yang, Y.; Fan, X.; Ji, Y.; Li, J.; Dai, Z.; Wu, Z. Glycine represses endoplasmic reticulum stress-related apoptosis and improves intestinal barrier by activating mammalian target of rapamycin complex 1 signaling. Anim. Nutr. 2022, 8, 1–9. [Google Scholar] [CrossRef]
- Zhong, Y.; Yan, Z.; Song, B.; Zheng, C.; Duan, Y.; Kong, X.; Deng, J.; Li, F. Dietary supplementation with betaine or glycine improves the carcass trait, meat quality and lipid metabolism of finishing mini-pigs. Anim. Nutr. 2021, 7, 376–383. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.A.; Fiorotto, M.L.; Burrin, D.G.; Reeds, P.J.; Nguyen, H.V.; Beckett, P.R.; Vann, R.C.; O’Connor, P.M.J. Stimulation of protein synthesis by both insulin and amino acids is unique to skeletal muscle in neonatal pigs. Am. J. Physiol. 2002, 282, E880–E890. [Google Scholar] [CrossRef] [PubMed]
- Saxton, R.A.; Sabatini, D.M. mTOR signaling in growth, metabolism, and disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [PubMed]
- Tato, I.; Bartrons, R.; Ventura, F.; Rosa, J.L. Amino acids activate mammalian target of rapamycin complex 2 (mTORC2) via PI3K/Akt signaling. J. Biol. Chem. 2011, 286, 6128–6142. [Google Scholar] [CrossRef]
- Suryawan, A.; Davis, T.A. Regulation of protein synthesis by amino acids in muscle of neonates. Front. Biosci. 2011, 16, 1445–1460. [Google Scholar] [CrossRef]
- Suryawan, A.; Rudar, M.; Fiorotto, M.L.; Davis, T.A. Differential regulation of mTORC1 activation by leucine and β-hydroxy-β-methylbutyrate in skeletal muscle of neonatal pigs. J. Appl. Physiol. 2020, 128, 286–295. [Google Scholar] [CrossRef]
- Brosnan, J.T.; Brosnan, M.E. Creatine: Endogenous metabolite, dietary, and therapeutic supplement. Annu. Rev. Nutr. 2007, 27, 241–261. [Google Scholar] [CrossRef]
- Brosnan, J.T.; Wijekoon, E.P.; Warford-Woolgar, L.; Trottier, N.L.; Brosnan, M.E.; Brunton, J.A.; Bertolo, R.F.P. Creatine synthesis is a major metabolic process in neonatal piglets and has important implications for amino acid metabolism and methyl balance. J. Nutr. 2009, 139, 1292–1297. [Google Scholar] [CrossRef]
- da Silva, R.P.; Nissim, I.; Brosnan, M.E.; Brosnan, J.T. Creatine synthesis: Hepatic metabolism of guanidinoacetate and creatine in the rat in vitro and in vivo. Am. J. Physiol. 2009, 296, E256–E261. [Google Scholar] [CrossRef]
- da Silva, R.P.; Clow, K.; Brosnan, J.T.; Brosnan, M.E. Synthesis of guanidinoacetate and creatine from amino acids by rat pancreas. Br. J. Nutr. 2014, 111, 571–577. [Google Scholar] [CrossRef]
- Dinesh, O.C.; Kankayaliyan, T.; Rademacher, M.; Tomlinson, C.; Bertolo, R.F.; Brunton, J.A. Neonatal piglets can synthesize adequate creatine, but only with sufficient dietary arginine and methionine, or with guanidinoacetate and excess methionine. J. Nutr. 2021, 151, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Daly, M.M. Guanidinoacetate methyltransferase activity in tissues and cultured cells. Arch. Biochem. Biophys. 1985, 236, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Edison, E.E.; Brosnan, M.E.; Meyer, C.; Brosnan, J.T. Creatine synthesis: Production of guanidinoacetate by the rat and human kidney in vivo. Am. J. Physiol. 2007, 293, F1799–F1804. [Google Scholar] [CrossRef] [PubMed]
- Guthmiller, P.; Van Pilsum, J.F.; Boen, J.R.; McGuire, D.M. Cloning and sequencing of rat kidney L-arginine:glycine amidinotransferase. Studies on the mechanism of regulation by growth hormone and creatine. J. Biol. Chem. 1994, 269, 17556–17560. [Google Scholar] [CrossRef]
- Griffith, O.W.; Meister, A. Glutathione: Interorgan translocation, turnover, and metabolism. Proc. Natl. Acad. Sci. USA 1979, 76, 5606–5610. [Google Scholar] [CrossRef]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and creatinine metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef]
- Kazak, L.; Cohen, P. Creatine metabolism: Energy homeostasis, immunity and cancer biology. Nat. Rev. Endocrinol. 2020, 16, 421–436. [Google Scholar] [CrossRef]
- Lu, S.C. Glutathione synthesis. Biochim. Biophys. Acta 2013, 1830, 3143–5313. [Google Scholar] [CrossRef]
- Meister, A.; Anderson, M.E. Glutathione. Annu. Rev. Biochem. 1983, 52, 711–760. [Google Scholar] [CrossRef]
- Sen, C.K. Redox signaling and the emerging potential of thiol antioxidants. Biochem. Pharmacol. 1998, 55, 1747–1758. [Google Scholar] [CrossRef]
- Wank, V.; Fischer, M.S.; Walter, B.; Bauer, R. Muscle growth and fiber type composition in hind limb muscles during postnatal development in pigs. Cells Tissues Organs 2006, 182, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Posey, E.A.; He, W.L.; Steele, C.C.; Savell, J.W.; Bazer, F.W.; Wu, G. Dietary glycine supplementation enhances creatine availability in tissues of pigs with intrauterine growth restriction. J. Anim. Sci. 2024, 102, skae344. [Google Scholar] [CrossRef] [PubMed]
- Wallimann, T.; Tokarska-Schlattner, M.; Schlattner, U. The creatine kinase system and pleiotropic effects of creatine. Amino Acids 2011, 40, 1271–1296. [Google Scholar] [CrossRef] [PubMed]
- Maddock, R.J.; Bidner, B.S.; Carr, S.N.; McKeith, F.K.; Berg, E.P.; Savell, J.W. Creatine monohydrate supplementation and the quality of fresh pork in normal and halothane carrier pigs. J. Anim. Sci. 2002, 80, 997–1004. [Google Scholar] [CrossRef]
- He, W.L.; Posey, E.A.; Steele, C.C.; Savell, J.W.; Bazer, F.W.; Wu, G. Dietary glycine supplementation enhances glutathione availability in tissues of pigs with intrauterine growth restriction. J. Anim. Sci. 2024, 102, skae025. [Google Scholar] [CrossRef]
- Ringseis, R.; Peter, L.; Gessner, D.K.; Meyer, S.; Most, E.; Eder, K. Effect of Tenebrio molitor larvae meal on the antioxidant status and stress response pathways in tissues of growing pigs. Arch. Anim. Nutr. 2021, 75, 237–250. [Google Scholar] [CrossRef]
- Nolan, M.R.; Kennedy, D.G.; Blanchflower, W.J.; Kennedy, S. Feeding corn oil to vitamin E-deficient pigs increases lipid peroxidation and decreases tissue glutathione concentrations. Int. J. Vitam. Nutr. Res. 1995, 65, 181–186. [Google Scholar]
- Wu, G. Principles of Animal Nutrition; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Hahn, R.G.; Stalberg, H.P.; Gustafsson, S.A. Vasopressin and cortisol levels in response to glycine infusion. Scand. J. Urol. Nephrol. 1991, 25, 121–123. [Google Scholar] [CrossRef]
- Muller, W.A.; Aoki, T.T.; Cahill, G.F.J. Effect of alanine and glycine on glucagon secretion in postabsorptive and fasting obese man. J. Clin. Endocrinol. Metab. 1975, 40, 418–425. [Google Scholar] [CrossRef]
- Alves, A.; Lamarche, F.; Lefebvre, R.; Drevet Mulard, E.; Bassot, A.; Chanon, S.; Loizon, E.; Pinteur, C.; Bloise, A.M.N.d.L.G.; Godet, M.; et al. Glycine supplementation in obesity worsens glucose intolerance through enhanced liver gluconeogenesis. Nutrients 2023, 15, 96. [Google Scholar] [CrossRef]
- Nakashima, K.; Yakabe, Y.; Ishida, A.; Katsumata, M. Effects of orally administered glycine on myofibrillar proteolysis and expression of proteolytic-related genes of skeletal muscle in chicks. Amino Acids 2008, 35, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xu, J.; Wang, T.; Wan, X.; Zhang, F.; Wang, L.; Zhu, X.; Gao, P.; Shu, G.; Jiang, Q.; et al. The dipeptide Pro-Gly promotes IGF-1 expression and secretion in HepG2 and female mice via PepT1-JAK2/STAT5 pathway. Front. Endocrinol. 2018, 9, 424. [Google Scholar] [CrossRef] [PubMed]
- Kasai, K.; Suzuki, H.; Nakamura, T.; Shiina, H.; Shimoda, S.I. Glycine stimulated growth hormone release in man. Acta Endocrinol. 1980, 93, 283–286. [Google Scholar]
- Estienne, M.J.; Harter-Dennis, J.M.; Barb, C.R. Role of neuropeptides and amino acids in controlling secretion of hormones from the anterior pituitary gland in pigs. J. Reprod. Fertil. Suppl. 1997, 52, 3–17. [Google Scholar] [CrossRef]
- Flynn, N.E.; Shaw, M.H.; Becker, J.T. Amino acids in health and endocrine function. Adv. Exp. Med. Biol. 2020, 1265, 97–109. [Google Scholar]
- Dennis, M.D.; Baum, J.I.; Kimball, S.R.; Jefferson, L.S. Mechanisms involved in the coordinate regulation of mTORC1 by insulin and amino acids. J. Biol. Chem. 2011, 286, 8287–8296. [Google Scholar] [CrossRef]
- Kim, J.; Guan, K. Amino acid signaling in TOR activation. Annu. Rev. Biochem. 2011, 80, 1001–1032. [Google Scholar] [CrossRef]
- Suryawan, A.; Jeyapalan, A.S.; Orellana, R.A.; Wilson, F.A.; Nguyen, H.V.; Davis, T.A. Leucine stimulates protein synthesis in skeletal muscle of neonatal pigs by enhancing mTORC1 activation. Am. J. Physiol. 2008, 295, E868–E875. [Google Scholar] [CrossRef]
- Jewell, J.L.; Russell, R.C.; Guan, K.L. Amino acid signalling upstream of mTOR. Nat. Rev. Mol. Cell. Biol. 2013, 14, 133–139. [Google Scholar] [CrossRef]
- Caldow, M.K.; Ham, D.J.; Trieu, J.; Chung, J.D.; Lynch, G.S.; Koopman, R. Glycine protects muscle cells from wasting in vitro via mTORC1 signaling. Front. Nutr. 2019, 6, 172. [Google Scholar] [CrossRef]
- Lin, C.; Han, G.; Ning, H.; Song, J.; Ran, N.; Yi, X.; Seow, Y.; Yin, H. Glycine enhances satellite cell proliferation, cell transplantation, and oligonucleotide efficacy in dystrophic muscle. Mol. Ther. 2020, 28, 1339–1358. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Amino Acids: Biochemistry and Nutrition, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2022. [Google Scholar]
- Atherton, P.J.; Smith, K.; Etheridge, T.; Rankin, D.; Rennie, M.J. Distinct anabolic signalling responses to amino acids in C2C12 skeletal muscle cells. Amino Acids 2010, 38, 1533–1539. [Google Scholar] [CrossRef] [PubMed]
- Nakajo, T.; Yamatsuji, T.; Ban, H.; Shigemitsu, K.; Haisa, M.; Motoki, T.; Noma, K.; Nobuhisa, T.; Matsuoka, J.; Gunduz, M.; et al. Glutamine is a key regulator for amino acid-controlled cell growth through the mTOR signaling pathway in rat intestinal epithelial cells. Biochem. Biophys. Res. Commun. 2005, 326, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.; Reynolds, N.; Downie, S.; Patel, A.; Rennie, M.J. Effects of flooding amino acids on incorporation of labeled amino acids into human muscle protein. Am. J. Physiol. 1998, 275, E73–E78. [Google Scholar] [CrossRef]
- Zeng, J.D.; Wu, W.K.K.; Wang, H.-Y.; Li, X.-X. Serine and one-carbon metabolism, a bridge that links mTOR signaling and DNA methylation in cancer. Pharmacol. Res. 2019, 149, 104352. [Google Scholar] [CrossRef]
- Yevenes, G.E.; Zeilhofer, H.U. Allosteric modulation of glycine receptors. Br. J. Pharmacol. 2011, 164, 224–236. [Google Scholar] [CrossRef]
- Armengaud, J.B.; Yzydorczyk, C.; Siddeek, B.; Peyter, A.C.; Simeoni, U. Intrauterine growth restriction: Clinical consequences on health and disease at adulthood. Reprod. Toxicol. 2021, 99, 168–176. [Google Scholar] [CrossRef]
- Selak, M.A.; Storey, B.T.; Peterside, I.; Simmons, R.A. Impaired oxidative phosphorylation in skeletal muscle of intrauterine growth-retarded rats. Am. J. Physiol. 2003, 285, E130–E137. [Google Scholar] [CrossRef]
- Matyba, P.; Florowski, T.; Dasiewicz, K.; Ferenc, K.; Olszewski, J.; Trela, M.; Galemba, G.; Słowiński, M.; Sady, M.; Domańska, D.; et al. Performance and meat quality of intrauterine growth restricted pigs. Animals 2021, 11, 254. [Google Scholar] [CrossRef]
- Sukhwani, M.; Antolín, E.; Herrero, B.; Rodríguez, R.; de la Calle, M.; López, F.; Bartha, J.L. Management and perinatal outcome of selective intrauterine growth restriction in monochorionic pregnancies. J. Matern. Fetal Neonatal Med. 2021, 34, 3838–3843. [Google Scholar] [CrossRef]
- Zhang, H.; Su, W.; Ying, Z.; Chen, Y.; Zhou, L.; Li, Y.; Zhang, J.; Zhang, L.; Wang, T. N-acetylcysteine attenuates intrauterine growth retardation-induced hepatic damage in suckling piglets by improving glutathione synthesis and cellular homeostasis. Eur. J. Nutr. 2018, 57, 327–338. [Google Scholar] [CrossRef]
- Shah, D.K.; Pereira, S.; Lodygensky, G.A. Long-term neurologic consequences following fetal growth restriction: The impact on brain reserve. Dev. Neurosci. 2025, 47, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.X.; Li, S.; Wu, Z.L.; Dai, Z.L.; Li, J.; Wang, X.L.; Wu, G. Glycine supplementation to breast-fed piglets attenuates postweaning jejunal epithelial apoptosis: A functional role of CHOP signaling. Amino Acids 2019, 51, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Silva, K.E.; Mansilla, W.D.; Shoveller, A.K.; Htoo, J.K.; Cant, J.P.; de Lange, C.F.M.; Huber, L.A. The effect of supplementing glycine and serine to a low crude protein diet on growth and skin collagen abundance of nursery pigs. J. Anim. Sci. 2020, 98, skaa023. [Google Scholar] [CrossRef]
- Powell, S.; Bidner, T.D.; Payne, R.L.; Southern, L.L. Growth performance of 20- to 50-kilogram pigs fed low-crude-protein diets supplemented with histidine, cystine, glycine, glutamic acid, or arginine. J. Anim. Sci. 2011, 89, 3643–3650. [Google Scholar] [CrossRef]
- Jiang, S.; Quan, W.; Luo, J.; Lou, A.; Zhou, X.; Li, F.; Shen, Q.W. Low-protein diets supplemented with glycine improve pig growth performance and meat quality: An untargeted metabolomic analysis. Front. Vet. Sci. 2023, 10, 1170573. [Google Scholar] [CrossRef]
- Wahid, S.T.; Lee, S.S.; Kim, I.H. The impact of glycine and glutamate, as components of glutathione precursors, on the productivity, digestive performance and blood profile of weaning pigs. J. Anim. Physiol. Anim. Nutr. 2024, 108, 1704–1711. [Google Scholar] [CrossRef]
- Zhou, X.H.; Liu, Y.H.; Zhang, L.Y.; Kong, X.F.; Li, F.N. Serine-to-glycine ratios in low-protein diets regulate intramuscular fat by affecting lipid metabolism and myofiber type transition in the skeletal muscle of growing-finishing pigs. Anim. Nutr. 2021, 7, 384–392. [Google Scholar] [CrossRef]
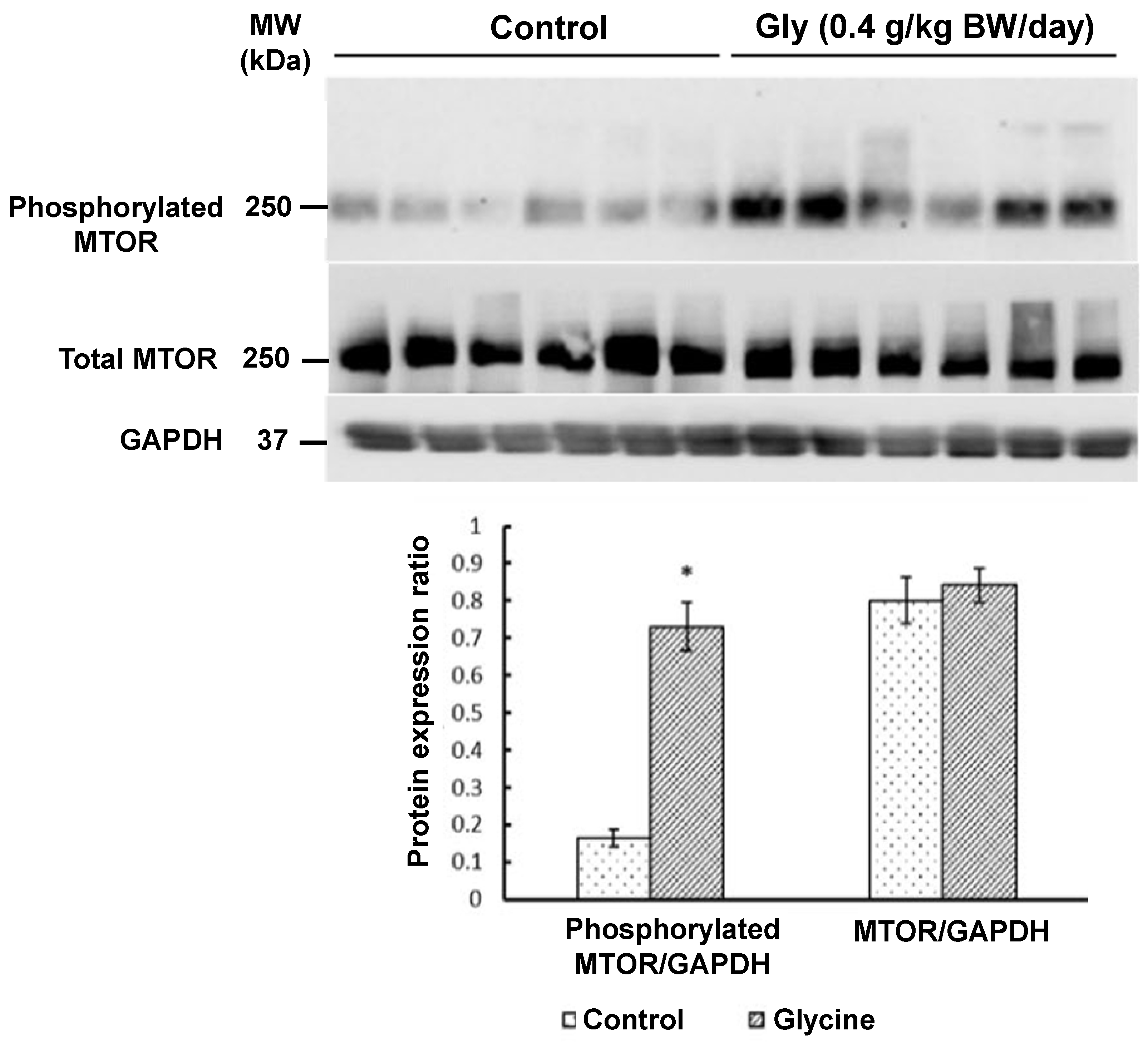
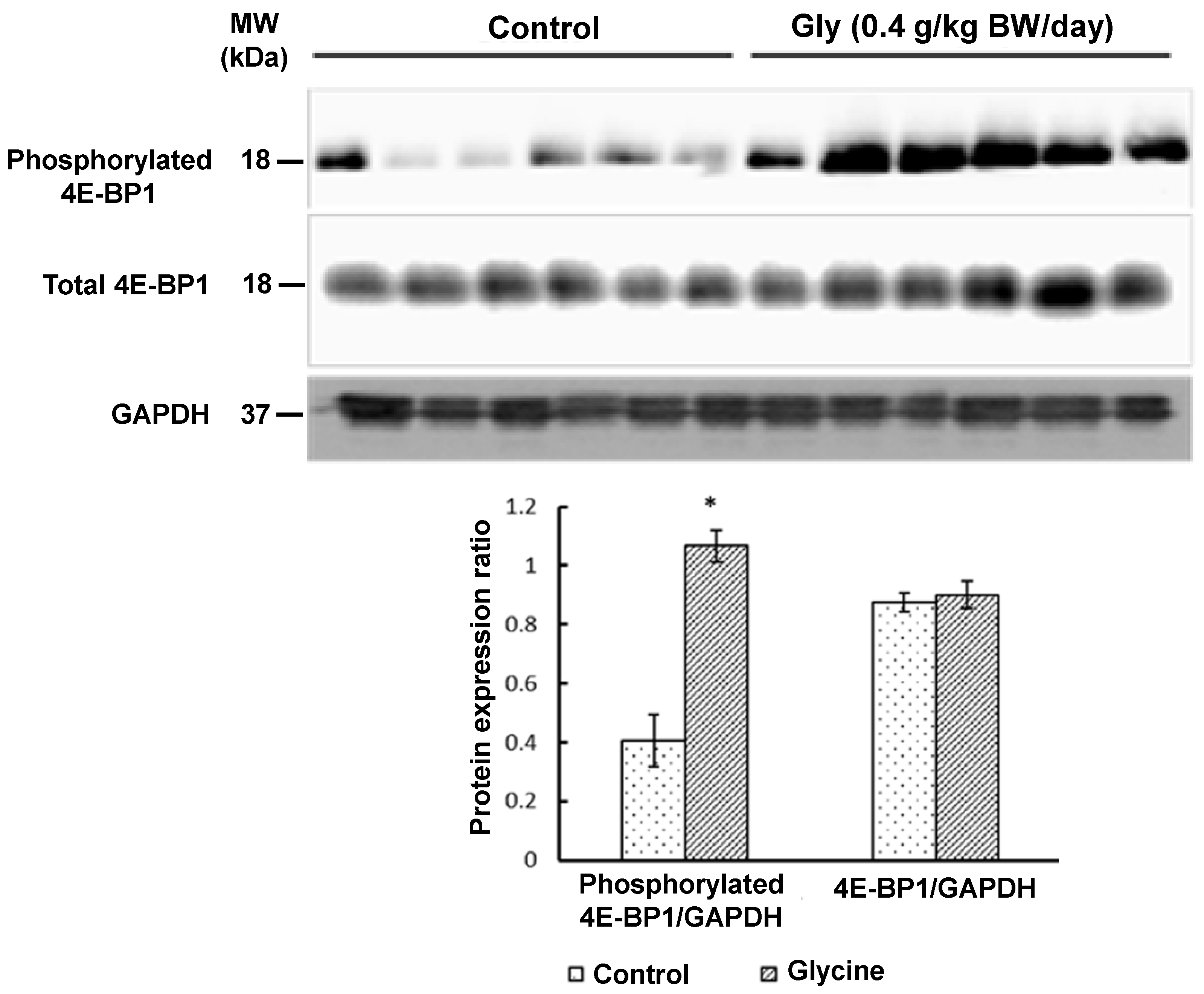
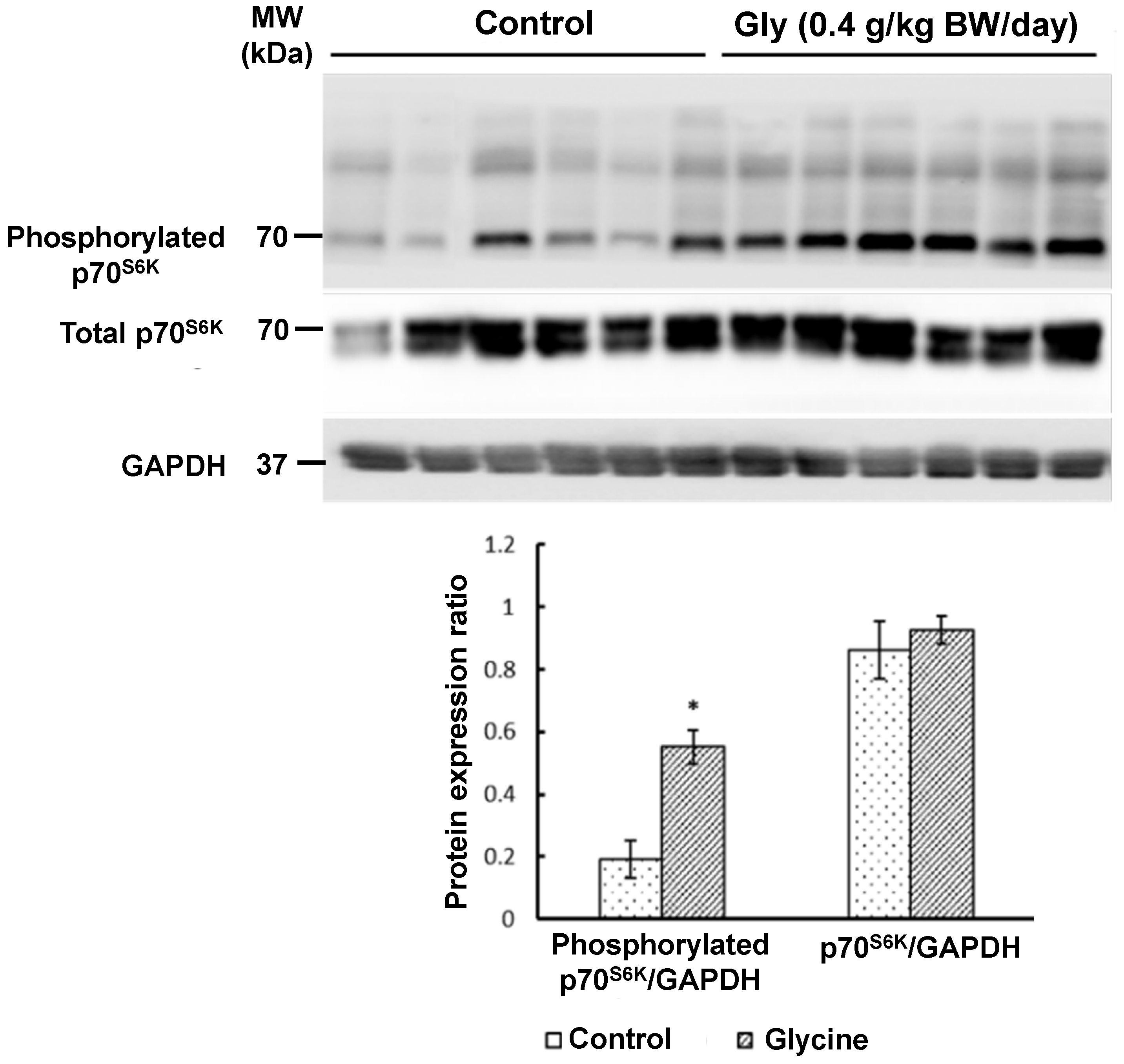

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | % (as-Fed Basis) 1 |
|---|---|
| Ingredient | |
| Corn grain | 57.50 |
| Soybean meal, 44.5% crude protein | 27.00 |
| Cornstarch | 2.0 |
| Sugarcane molasses | 3.85 |
| Potassium chloride | 0.10 |
| Salt | 0.35 |
| Vitamin–mineral premix 2 | 3.00 |
| Vegetable oil | 3.00 |
| Dicalcium phosphate | 2.50 |
| Limestone | 0.70 |
| Chemical composition | |
| Dry matter, % | 90.0 |
| Metabolizable energy, Mcal/kg | 3.32 |
| Crude protein, % | 17.5 |
| Calcium, % | 1.04 |
| Available phosphorus, % | 0.54 |
| Total phosphorus, % | 0.79 |
| Age | Oral Administration of Glycine (g/kg Body Weight/day) | |||
|---|---|---|---|---|
| (days) | 0 | 0.2 | 0.4 | 0.8 |
| Body weight (kg) | ||||
| 0 | 0.86 ± 0.07 | 0.88 ± 0.02 | 0.85 ± 0.04 | 0.88 ± 0.04 |
| 7 | 1.63 ± 0.08 b | 1.88 ± 0.06 a | 2.07 ± 0.05 a | 2.01 ± 0.08 a |
| 14 | 2.30 ± 0.12 c | 2.76 ± 0.08 b | 3.16 ± 0.14 a | 3.07 ± 0.07 a |
| Weight gain (kg) | ||||
| Days 7–0 | 0.77 ± 0.04 c | 1.00 ± 0.05 b | 1.22 ± 0.02 a | 1.14 ± 0.05 a,b |
| Days 14–7 | 0.67 ± 0.04 c | 0.88 ± 0.04 b | 1.10 ± 0.09 a | 1.06 ± 0.06 a |
| Days 14–0 | 1.44 ± 0.08 c | 1.88 ± 0.06 b | 2.31 ± 0.10 a | 2.20 ± 0.04 a |
| p-values | ||||
| Glycine | Age | Glycine × Age | ||
| Body weight | <0.001 | <0.001 | 0.862 | |
| Weight gain | <0.001 | <0.001 | 0.735 | |
| Tissue | Oral Administration of Glycine (g/kg Body Weight/day) | ||||
|---|---|---|---|---|---|
| 0 | 0.2 | 0.4 | 0.8 | p-Value | |
| Absolute Tissue Weight (g) | |||||
| LLM 2 | 7.36 ± 0.22 c | 9.02 ± 0.19 b | 10.5 ± 0.28 a | 10.1 ± 0.34 a | <0.001 |
| GM | 32.9 ± 0.73 c | 40.1 ± 1.5 b | 46.9 ± 0.76 a | 45.7 ± 1.2 a | <0.001 |
| SI | 84.0 ± 1.8 c | 102.6 ± 2.5 b | 116.4 ± 3.7 a | 112.9 ± 4.3 a | <0.001 |
| Liver | 70.6 ± 1.9 c | 85.1 ± 1.5 b | 97.9 ± 2.8 a | 94.6 ± 4.0 a | <0.001 |
| Kidneys | 16.6 ± 0.40 c | 20.1 ± 0.56 b | 23.2 ± 0.69 a | 22.6 ± 0.63 a | <0.001 |
| Pancreas | 3.20 ± 0.12 c | 3.91 ± 0.09 b | 4.67 ± 0.29 a | 4.42 ± 0.15 a | <0.001 |
| Stomach | 10.7 ± 0.43 c | 13.2 ± 0.35 b | 15.3 ± 0.40 a | 14.6 ± 0.21 a | <0.001 |
| Heart | 13.5 ± 0.39 c | 16.0 ± 0.61 b | 18.5 ± 0.57 a | 18.1 ± 0.45 a | <0.001 |
| Percentage of Body Weight (%) | |||||
| LLM | 0.33 ± 0.02 | 0.32 ± 0.01 | 0.33 ± 0.01 | 0.33 ± 0.02 | 0.996 |
| GM | 1.47 ± 0.08 | 1.44 ± 0.02 | 1.48 ± 0.04 | 1.49 ± 0.06 | 0.925 |
| SI | 3.76 ± 0.20 | 3.70 ± 0.08 | 3.65 ± 0.05 | 3.66 ± 0.13 | 0.928 |
| Liver | 3.16 ± 0.15 | 3.07 ± 0.06 | 3.08 ± 0.05 | 3.06 ± 0.15 | 0.946 |
| Kidneys | 0.74 ± 0.04 | 0.72 ± 0.02 | 0.73 ± 0.02 | 0.73 ± 0.02 | 0.962 |
| Pancreas | 0.14 ± 0.01 | 0.14 ± 0.01 | 0.15 ± 0.01 | 0.14 ± 0.01 | 0.861 |
| Stomach | 0.48 ± 0.03 | 0.47 ± 0.01 | 0.48 ± 0.02 | 0.47 ± 0.01 | 0.965 |
| Heart | 0.60 ± 0.03 | 0.57 ± 0.02 | 0.58 ± 0.03 | 0.59 ± 0.02 | 0.856 |
| Amino Acid | Oral Administration of Glycine (g/kg Body Weight/day) | ||||
|---|---|---|---|---|---|
| 0 | 0.2 | 0.4 | 0.8 | p-Value | |
| nmol/mL | |||||
| Alanine | 903 ± 9.6 a | 627 ± 6.1 b | 613 ± 8.5 b | 615 ± 7.6 b | <0.001 |
| β-Alanine | 7.3 ± 1.1 | 7.1 ± 0.8 | 6.9 ± 0.5 | 7.2 ± 0.7 | 0.987 |
| Arginine | 120 ± 9.2 | 122 ± 7.2 | 127 ± 5.7 | 125 ± 8.9 | 0.925 |
| Aspartate | 14.1 ± 1.6 | 15.3 ± 1.7 | 14.7 ± 1.7 | 16.0 ± 2.5 | 0.908 |
| Asparagine | 82.2 ± 6.1 | 83.4 ± 6.3 | 79.5 ± 9.9 | 75.2 ± 5.7 | 0.857 |
| Citrulline | 52.8 ± 1.6 | 54.4 ± 2.8 | 51.6 ± 3.1 | 53.6 ± 4.6 | 0.936 |
| Cysteine | 182 ± 10 | 177 ± 12 | 180 ± 13 | 186 ± 11 | 0.955 |
| Glutamate | 189 ± 12 a | 161 ± 11 a,b | 144 ± 8.9 b | 137 ± 9.6 b | 0.010 |
| Glutamine | 654 ± 18 a | 547 ± 19 b | 478 ± 21 c | 455 ± 4.9 c | <0.001 |
| Glycine | 586 ± 10 a | 890 ± 17 b | 1134 ± 19 c | 1382 ± 22 d | <0.001 |
| Histidine | 67.2 ± 4.3 | 65.4 ± 3.4 | 64.6 ± 5.3 | 67.9 ± 7.2 | 0.967 |
| 4-Hydroxyproline | 78.0 ± 2.7 | 75.7 ± 4.2 | 76.2 ± 3.2 | 77.3 ± 3.5 | 0.964 |
| Isoleucine | 106 ± 5.8 | 112 ± 4.7 | 107 ± 9.4 | 110 ± 8.2 | 0.933 |
| Leucine | 151 ± 6.8 | 155 ± 4.0 | 153 ± 6.5 | 148 ± 8.4 | 0.893 |
| Lysine | 136 ± 11 | 141 ± 9.4 | 149 ± 11 | 153 ± 9.0 | 0.639 |
| Methionine | 60.1 ± 4.1 | 62.5 ± 5.3 | 67.5 ± 7.7 | 66.9 ± 4.9 | 0.759 |
| Ornithine | 57.0 ± 4.2 | 59.5 ± 3.1 | 57.5 ± 6.1 | 55.6 ± 3.2 | 0.935 |
| Phenylalanine | 71.5 ± 5.0 | 64.0 ± 3.9 | 71.0 ± 8.7 | 74.3 ± 5.8 | 0.678 |
| Proline | 287 ± 17 | 291 ± 15 | 283 ± 16 | 280 ± 15 | 0.964 |
| Serine | 278 ± 13 b | 312 ± 8.6 a,b | 328 ± 12 a | 347 ± 6.8 a | 0.001 |
| Taurine | 124 ± 2.7 | 126 ± 11 | 129 ± 5.4 | 127 ± 3.8 | 0.959 |
| Threonine | 154 ± 7.7 a | 171 ± 6.1 a,b | 189 ± 11 b | 194 ± 11 b | 0.024 |
| Tryptophan | 76.0 ± 5.1 | 73.2 ± 2.9 | 72.2 ± 5.6 | 76.1 ± 4.1 | 0.902 |
| Tyrosine | 154 ± 7.2 | 149 ± 4.7 | 156 ± 9.1 | 157 ± 8.2 | 0.877 |
| Valine | 210 ± 9.1 | 211 ± 5.1 | 216 ± 6.3 | 218 ± 4.2 | 0.783 |
| Amino Acid | Oral Administration of Glycine (g/kg Body Weight/day) | ||||
|---|---|---|---|---|---|
| 0 | 0.2 | 0.4 | 0.8 | p-Value | |
| Longissimus lumborum muscle (nmol/g of wet tissue) | |||||
| Glycine | 2108 ± 79 d | 2635 ± 64 c | 3221 ± 87 b | 3958 ± 96 a | <0.001 |
| Serine | 2564 ± 69 c | 2893 ± 66 b | 3124 ± 79 a | 3181 ± 75 a | <0.001 |
| Gastrocnemius muscle (nmol/g of wet tissue) | |||||
| Glycine | 2264 ± 88 d | 2986 ± 92 c | 3706 ± 108 b | 4483 ± 112 a | <0.001 |
| Serine | 2487 ± 74 c | 2793 ± 81 b | 3065 ± 91 a | 3124 ± 96 a | <0.001 |
| Liver (nmol/g of wet tissue) | |||||
| Glycine | 4413 ± 120 d | 5290 ± 109 c | 6193 ± 147 b | 7253 ± 178 a | <0.001 |
| Serine | 1060 ± 60 c | 1187 ± 55 b,c | 1341 ± 69 a,b | 1404 ± 75 a | 0.006 |
| Jejunum (nmol/g of wet tissue) | |||||
| Glycine | 2304 ± 98 d | 2806 ± 67 c | 3367 ± 80 b | 3975 ± 77 a | <0.001 |
| Serine | 1296 ± 65 b | 1420 ± 60 a,b | 1581 ± 55 a | 1635 ± 74 a | 0.005 |
| Kidney (nmol/g of wet tissue) | |||||
| Glycine | 7324 ± 112 d | 8430 ± 168 c | 9855 ± 367 b | 12,086 ± 574 a | <0.001 |
| Serine | 869 ± 32 c | 923 ± 36 b,c | 1038 ± 41 a,b | 1097 ± 48 a | 0.004 |
| Glucose or Hormone Concentrations | Oral Administration of Glycine (g/kg BW/day) | p-Value | |||
|---|---|---|---|---|---|
| 0 | 0.2 | 0.4 | 0.8 | ||
| Glucose in plasma, mM | 5.73 ± 0.17 | 5.64 ± 0.23 | 5.60 ± 0.20 | 5.84 ± 0.12 | 0.802 |
| Ammonia in plasma, μM | 141 ± 5.7 a | 124 ± 5.5 b | 106 ± 5.1 c | 104 ± 5.3 c | <0.001 |
| Urea in plasma, mM | 2.18 ± 0.09 a | 1.96 ± 0.07 b | 1.70 ± 0.06 c | 1.73 ± 0.06 c | <0.001 |
| TBARS in plasma, μM | 5.26 ± 0.23 a | 4.68 ± 0.21 b | 4.07 ± 0.16 c | 3.52 ± 0.11 d | <0.001 |
| Free fatty acids in plasma, μM | 253 ± 16 | 260 ± 20 | 248 ± 14 | 272 ± 23 | 0.815 |
| Triacylglycerols in plasma, μM | 793 ± 48 | 812 ± 65 | 772 ± 46 | 788 ± 51 | 0.962 |
| Total cholesterol in plasma, mM | 2.03 ± 0.10 | 1.94 ± 0.08 | 2.07 ± 0.15 | 1.87 ± 0.12 | 0.618 |
| Cortisol in plasma, nM | 64.7 ± 3.1 | 63.2 ± 2.8 | 61.9 ± 3.4 | 62.6 ± 3.9 | 0.942 |
| Insulin in serum, pM | 60.9 ± 3.4 | 62.8 ± 3.0 | 61.2 ± 3.7 | 63.4 ± 2.7 | 0.933 |
| Growth hormone in serum, pM | 359 ± 16 | 376 ± 18 | 371 ± 13 | 365 ± 21 | 0.907 |
| IGF-I in serum, µg/L | 31.5 ± 1.4 | 30.9 ± 2.2 | 32.1 ± 1.8 | 33.0 ± 2.4 | 0.893 |
| Variable | Oral Administration of Glycine (g/kg Body Weight/day) | ||||
|---|---|---|---|---|---|
| 0 | 0.2 | 0.4 | 0.8 | p-Value | |
| Plasma (nmol/mL) | |||||
| Guanidinoacetate | 39.4 ± 1.0 c | 45.0 ± 1.5 b | 50.9 ± 1.8 a | 52.4 ± 2.3 a | <0.001 |
| Creatine | 252 ± 8.3 c | 295 ± 8.8 b | 327 ± 11 a | 336 ± 13 a | <0.001 |
| Phosphocreatine (PCr) | ND | ND | ND | ND | --- |
| Creatine + PCr | 252 ± 8.3 c | 295 ± 8.8 b | 327 ± 11 a | 336 ± 13 a | <0.001 |
| Creatinine | 38.1 ± 1.5 | 39.0 ± 1.9 | 41.8 ± 2.3 | 42.5 ± 2.6 | 0.408 |
| Longissimus lumborum muscle (μmol/g of wet tissue) | |||||
| Guanidinoacetate | 0.17 ± 0.01 | 0.18 ± 0.02 | 0.18 ± 0.02 | 0.19 ± 0.02 | 0.892 |
| Creatine | 12.0 ± 0.45 c | 13.9 ± 0.57 b | 15.6 ± 0.51 a | 16.1 ± 0.63 a | <0.001 |
| Phosphocreatine (PCr) | 19.5 ± 0.62 c | 22.4 ± 0.71 b | 24.7 ± 0.89 a | 25.6 ± 0.75 a | 0.001 |
| Creatine + PCr | 31.5 ± 1.1 c | 36.3 ± 1.2 b | 40.3 ± 1.4 a | 41.7 ± 1.4 a | <0.001 |
| Creatinine | 1.10 ± 0.06 | 1.15 ± 0.08 | 1.24 ± 0.07 | 1.28 ± 0.06 | 0.255 |
| Gastrocnemius muscle (μmol/g of wet tissue) | |||||
| Guanidinoacetate | 0.13 ± 0.01 | 0.14 ± 0.01 | 0.15 ± 0.01 | 0.16 ± 0.02 | 0.434 |
| Creatine | 11.2 ± 0.41 c | 13.0 ± 0.49 b | 14.8 ± 0.66 a | 15.3 ± 0.58 a | 0.001 |
| Phosphocreatine (PCr) | 18.3 ± 0.55 c | 20.8 ± 0.68 b | 23.9 ± 0.78 a | 24.4 ± 0.92 a | <0.001 |
| Creatine + PCr | 29.5 ± 0.70 c | 33.7 ± 1.1 b | 38.7 ± 1.2 a | 39.7 ± 1.2 a | <0.001 |
| Creatinine | 0.91 ± 0.03 | 0.93 ± 0.04 | 0.95 ± 0.05 | 0.97 ± 0.05 | 0.786 |
| Liver (μmol/g of wet tissue) | |||||
| Guanidinoacetate | 0.21 ± 0.02 | 0.23 ± 0.02 | 0.24 ± 0.02 | 0.25 ± 0.02 | 0.547 |
| Creatine | 1.93 ± 0.07 c | 2.24 ± 0.09 b | 2.52 ± 0.11 a | 2.60 ± 0.10 a | <0.001 |
| Phosphocreatine (PCr) | 0.34 ± 0.02 | 0.36 ± 0.02 | 0.39 ± 0.03 | 0.41 ± 0.02 | 0.260 |
| Creatine + PCr | 2.27 ± 0.06 c | 2.60 ± 0.11 b | 2.92 ± 0.12 a | 3.01 ± 0.10 a | 0.001 |
| Creatinine | 0.11 ± 0.01 | 0.12 ± 0.01 | 0.13 ± 0.02 | 0.13 ± 0.01 | 0.671 |
| Jejunum (μmol/g of wet tissue) | |||||
| Guanidinoacetate | 0.051 ± 0.003 | 0.053 ± 0.003 | 0.055 ± 0.004 | 0.058 ± 0.004 | 0.558 |
| Creatine | 3.43 ± 0.10 c | 3.88 ± 0.14 b | 4.50 ± 0.16 a | 4.61 ± 0.19 a | 0.001 |
| Phosphocreatine (PCr) | 3.91 ± 0.12 c | 4.46 ± 0.17 b | 5.09 ± 0.21 a | 5.17 ± 0.18 a | 0.001 |
| Creatine + PCr | 7.34 ± 0.21 c | 8.34 ± 0.31 b | 9.59 ± 0.34 a | 9.77 ± 0.37 a | 0.001 |
| Creatinine | 0.13 ± 0.01 | 0.14 ± 0.01 | 0.14 ± 0.01 | 0.16 ± 0.02 | 0.459 |
| Kidney (μmol/g of wet tissue) | |||||
| Guanidinoacetate | 0.68 ± 0.02 c | 0.78 ± 0.03 b | 0.89 ± 0.03 a | 0.91 ± 0.04 a | 0.001 |
| Creatine | 1.65 ± 0.06 c | 1.92 ± 0.07 b | 2.18 ± 0.09 a | 2.24 ± 0.11 a | <0.001 |
| Phosphocreatine (PCr) | 1.16 ± 0.05 c | 1.34 ± 0.05 b | 1.57 ± 0.07 a | 1.64 ± 0.06 a | <0.001 |
| Creatine + PCr | 2.81 ± 0.09 c | 3.26 ± 0.12 b | 3.75 ± 0.16 a | 3.88 ± 0.16 a | <0.001 |
| Creatinine | 0.34 ± 0.02 c | 0.41 ± 0.02 b | 0.49 ± 0.03 a | 0.51 ± 0.02 a | 0.001 |
| Variable | Oral Administration of Glycine (g/kg Body Weight/day) | |||||
|---|---|---|---|---|---|---|
| 0 | 0.2 | 0.4 | 0.8 | p-Value | ||
| Plasma | ||||||
| GSH, nmol/mL | 4.58 ± 0.13 d | 5.18 ± 0.16 c | 5.92 ± 0.20 b | 6.68 ± 0.23 a | <0.001 | |
| GSSG, nmol/mL | 0.768 ± 0.036 a | 0.723 ± 0.031 a,b | 0.685 ± 0.032 a,b | 0.637 ± 0.026 b | 0.045 | |
| GSSG/GSH, nmol/nmol | 0.169 ± 0.011 a | 0.140 ± 0.007 b | 0.116 ± 0.006 c | 0.096 ± 0.006 c | <0.001 | |
| Longissimus lumborum muscle | ||||||
| GSH, μmol/g of wet tissue | 0.679 ± 0.031 d | 0.773 ± 0.034 c,d | 0.895 ± 0.040 b | 1.02 ± 0.05 a | <0.001 | |
| GSSG, μmol/g of wet tissue | 0.118 ± 0.008 a | 0.106 ± 0.007 a,b | 0.095 ± 0.007 a,b | 0.089 ± 0.006 b | 0.041 | |
| GSSG/GSH, μmol/μmol | 0.173 ± 0.005 a | 0.138 ± 0.006 b | 0.105 ± 0.004 c | 0.088 ± 0.005 d | <0.001 | |
| Gastrocnemius muscle | ||||||
| GSH, μmol/g of wet tissue | 0.818 ± 0.033 d | 0.902 ± 0.037 c,d | 1.04 ± 0.05 b | 1.18 ± 0.06 a | <0.001 | |
| GSSG, μmol/g of wet tissue | 0.140 ± 0.009 a | 0.128 ± 0.008 a,b | 0.117 ± 0.007 a,b | 0.108 ± 0.006 b | 0.039 | |
| GSSG/GSH, μmol/μmol | 0.171 ± 0.007 a | 0.142 ± 0.006 b | 0.113 ± 0.009 c | 0.092 ± 0.003 d | <0.001 | |
| Liver | ||||||
| GSH, μmol/g of wet tissue | 4.19 ± 0.14 d | 4.78 ± 0.18 c | 5.40 ± 0.20 b | 6.15 ± 0.23 a | <0.001 | |
| GSSG, μmol/g of wet tissue | 0.282 ± 0.015 a | 0.252 ± 0.014 a,b | 0.237 ± 0.012 a,b | 0.227 ± 0.010 b | 0.032 | |
| GSSG/GSH, μmol/μmol | 0.067 ± 0.003 a | 0.053 ± 0.003 b | 0.044 ± 0.002 c | 0.037 ± 0.001 d | <0.001 | |
| Jejunum | ||||||
| GSH, μmol/g of wet tissue | 1.04 ± 0.08 d | 1.28 ± 0.07 c | 1.53 ± 0.06 b | 1.78 ± 0.09 a | <0.001 | |
| GSSG, μmol/g of wet tissue | 0.163 ± 0.008 a | 0.155 ± 0.006 a,b | 0.147 ± 0.008 a,b | 0.133 ± 0.006 b | 0.042 | |
| GSSG/GSH, μmol/μmol | 0.161 ± 0.011 a | 0.122 ± 0.006 b | 0.096 ± 0.004 c | 0.075 ± 0.003 d | <0.001 | |
| Kidney | ||||||
| GSH, μmol/g of wet tissue | 0.732 ± 0.020 c | 0.798 ± 0.026 b,c | 0.881 ± 0.037 a,b | 0.962 ± 0.043 a | <0.001 | |
| GSSG, μmol/g of wet tissue | 0.056 ± 0.003 a | 0.051 ± 0.003 a,b | 0.046 ± 0.003 a,b | 0.044 ± 0.003 b | 0.025 | |
| GSSG/GSH, μmol/μmol | 0.076 ± 0.003 a | 0.063 ± 0.002 b | 0.053 ± 0.002 c | 0.046 ± 0.001 d | <0.001 | |
| Variable | Oral Administration of Glycine (g/kg Body Weight/day) | ||
|---|---|---|---|
| 0 | 0.4 | p-Value | |
| Longissimus lumborum muscle 2 | |||
| Fractional rate of protein synthesis, %/day | 14.1 ± 0.62 | 16.3 ± 0.71 | 0.035 |
| Amount of protein per dissected tissue, g | 1.39 ± 0.05 | 1.82 ± 0.09 | <0.001 |
| Absolute rate of protein synthesis, g/day | 0.20 ± 0.013 | 0.30 ± 0.020 | 0.001 |
| Gastrocnemius muscle | |||
| Fractional rate of protein synthesis, %/day | 15.3 ± 0.59 | 17.6 ± 0.74 | 0.029 |
| Amount of protein per tissue, g | 6.28 ± 0.23 | 8.33 ± 0.44 | 0.001 |
| Absolute rate of protein synthesis, g/day | 0.96 ± 0.05 | 1.47 ± 0.09 | <0.001 |
| Liver | |||
| Fractional rate of protein synthesis, %/day | 80.6 ± 3.2 | 94.5 ± 4.1 | 0.018 |
| Amount of protein per tissue, g | 11.2 ± 0.43 | 14.6 ± 0.45 | <0.001 |
| Absolute rate of protein synthesis, g/day | 9.09 ± 0.61 | 13.8 ± 0.82 | <0.001 |
| Jejunum | |||
| Fractional rate of protein synthesis, %/day | 57.2 ± 2.4 | 67.5 ± 2.7 | 0.013 |
| Amount of protein per tissue, g | 9.71 ± 0.49 | 12.6 ± 0.79 | 0.008 |
| Absolute rate of protein synthesis, g/day | 5.54 ± 0.30 | 8.64 ± 0.83 | 0.003 |
| Kidneys | |||
| Fractional rate of protein synthesis, %/day | 36.2 ± 1.5 | 42.0 ± 2.1 | 0.041 |
| Amount of protein in two kidneys, g | 1.93 ± 0.11 | 2.59 ± 0.14 | 0.002 |
| Absolute rate of protein synthesis, g/day | 0.70 ± 0.05 | 1.08 ± 0.07 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, S.; Long, D.W.; Bazer, F.W.; Burghardt, R.C.; Johnson, G.A.; Wu, G. Glycine Supplementation Enhances the Growth of Sow-Reared Piglets with Intrauterine Growth Restriction. Animals 2025, 15, 1855. https://doi.org/10.3390/ani15131855
Hu S, Long DW, Bazer FW, Burghardt RC, Johnson GA, Wu G. Glycine Supplementation Enhances the Growth of Sow-Reared Piglets with Intrauterine Growth Restriction. Animals. 2025; 15(13):1855. https://doi.org/10.3390/ani15131855
Chicago/Turabian StyleHu, Shengdi, David W. Long, Fuller W. Bazer, Robert C. Burghardt, Gregory A. Johnson, and Guoyao Wu. 2025. "Glycine Supplementation Enhances the Growth of Sow-Reared Piglets with Intrauterine Growth Restriction" Animals 15, no. 13: 1855. https://doi.org/10.3390/ani15131855
APA StyleHu, S., Long, D. W., Bazer, F. W., Burghardt, R. C., Johnson, G. A., & Wu, G. (2025). Glycine Supplementation Enhances the Growth of Sow-Reared Piglets with Intrauterine Growth Restriction. Animals, 15(13), 1855. https://doi.org/10.3390/ani15131855