

Feeding Frequency Affects the Growth Performance and Intestinal Health of Juvenile Red-Tail Catfish (Hemibagrus wyckioides) with the Same Amount of Daily Feed

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Design and Culture Conditions
2.2. Sample Collection
2.3. Growth Performance and Composition Analysis
2.4. Serum Biochemical Parameter Determination
2.5. Intestinal Digestive Enzyme Activities and Hepatic Antioxidant Parameters
2.6. Morphology of Foregut Tissue
2.7. Reverse Transcription—Quantitative Real-Time PCR (RT-qPCR)
2.8. Profiling Hindgut Microbiome
2.9. Intestinal Transcriptome Analysis
2.10. Intestinal Metabolism Analysis
2.11. Statistical Analysis
3. Results
3.1. Growth Performance, Feed Utilization, and Body Composition
3.2. Serum Biochemistry
3.3. Liver Antioxidant Indices
3.4. Digestive Enzyme Activity and Morphological Characteristics in Foregut
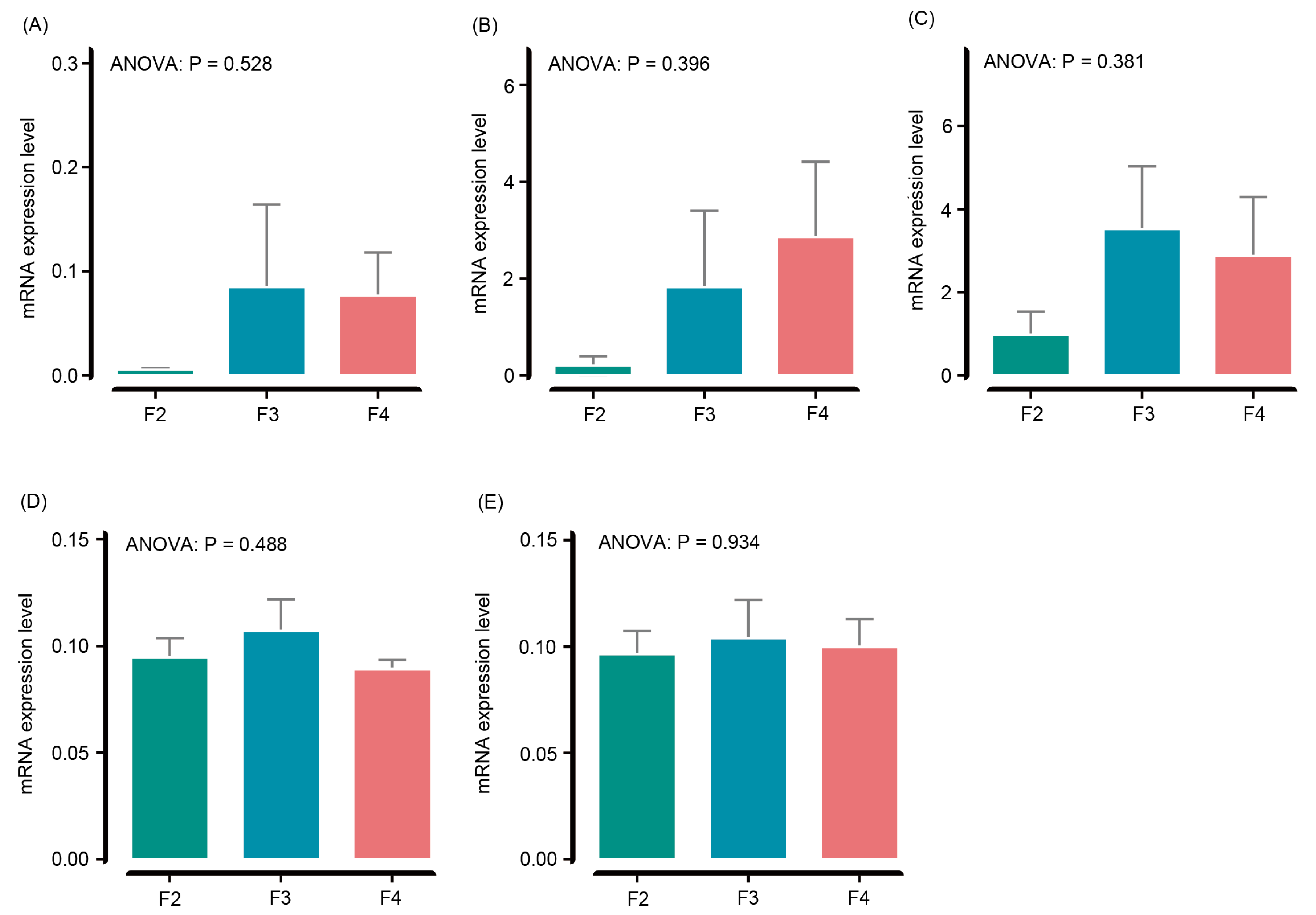
3.5. Liver Antioxidant Genes
3.6. Hindgut Microbiota Analysis
3.7. Differential Analysis and Mining Biomarkers Related to Specific Phenotypes
3.8. Molecular Ecological Network Analysis
3.9. Intestinal Transcriptome
3.10. Intestinal Metabolism
4. Discussion
4.1. Effects of the Feeding Frequency on the Physiological and Biochemical Indices of H. wyckioides
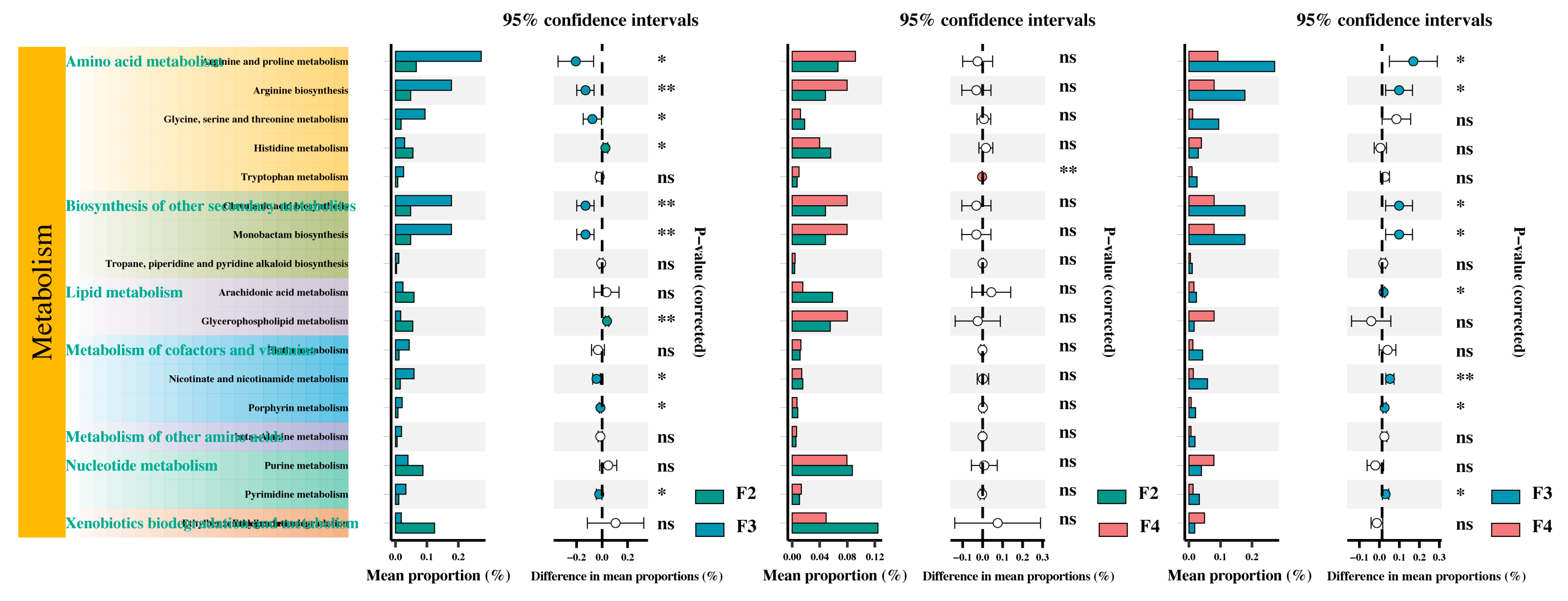
4.2. Effects of the Feeding Frequency on the Intestinal Microbiota of H. wyckioides
4.3. Effects of the Feeding Frequency on the Intestinal Transcriptome of H. wyckioides
4.4. Effects of the Feeding Frequency on the Intestinal Metabolome of H. wyckioides
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| FCR | Feed Conversion Ratio |
| SGR | Specific Growth Rate |
| WGR | Weight Gain Ratio |
| SR | Survival Rate |
| HSI | Hepatosomatic Index |
| VSI | Viscerosomatic Index |
| CF | Condition Factor |
| TC | Total Cholesterol |
| TG | Triglyceride |
| HDL-C | High-Density Lipoprotein Cholesterol |
| LDL-C | Low-Density Lipoprotein Cholesterol |
| GLU | Glucose |
| TP | Total Protein |
| ALB | Albumin |
| T-AOC | Total Antioxidant Capacity |
| MDA | Malondialdehyde |
| SOD | Superoxide Dismutase |
| GSH-PX | Glutathione Peroxidase |
| ALT | Alanine Transferase |
| ALP | Alkaline Phosphatase |
| AST | Aspartate Transferase |
| H&E | Hematoxylin and Eosin |
| RT-qPCR | Reverse Transcription—Quantitative Real-Time PCR |
| PCoA | Principal Coordinates Analysis |
| NMDS | Non-Metric Multidimensional Scaling |
| ASV | Amplicon Sequence Variant |
| QC | Quality Control |
| LC-MS | Liquid Chromatography-Mass Spectrometry |
References
- FAO. The State of World Fisheries and Aquaculture 2024; FAO: Rome, Italy, 2024. [Google Scholar] [CrossRef]
- Chan, H.L.; Cai, J.; Leung, P. Aquaculture production and diversification: What causes what? Aquaculture 2024, 583, 740626. [Google Scholar] [CrossRef]
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- See, K.F.; Ibrahim, R.A.; Goh, K.H. Aquaculture efficiency and productivity: A comprehensive review and bibliometric analysis. Aquaculture 2021, 544, 736881. [Google Scholar] [CrossRef]
- Wu, B.; Huang, L.; Chen, J.; Zhang, Y.; Chen, X.; Wu, C.; Deng, X.; Gao, J.; He, J. Effects of feeding frequency on growth performance, feed intake, metabolism and expression of fgf21 in grass carp (Ctenopharyngodon idellus). Aquaculture 2021, 545, 737196. [Google Scholar] [CrossRef]
- Yúfera, M.; Romero, M.J.; Pujante, I.M.; Astola, A.; Mancera, J.M.; Sánchez-Vázquez, F.J.; Moyano, F.J.; Martínez-Rodríguez, G. Effect of feeding frequency on the daily rhythms of acidic digestion in a teleost fish (Gilthead seabream). Chronobiol. Int. 2014, 31, 1024–1033. [Google Scholar] [CrossRef]
- Costa-Bomfim, C.N.; Pessoa, W.V.N.; Oliveira, R.L.M.; Farias, J.L.; Domingues, E.C.; Hamilton, S.; Cavalli, R.O. The effect of feeding frequency on growth performance of juvenile cobia, Rachycentron canadum (Linnaeus, 1766). J. Appl. Ichthyol. 2014, 30, 135–139. [Google Scholar] [CrossRef]
- Calixto da Silva, E.; Sterzelecki, F.C.; Alves Musialak, L.; Sugai, J.K.; de Jesus Pinto Castro, J.; Pedrotti, F.S.; Magnotti, C.; dos Santos Cipriano, F.; Cerqueira, V.R. Effect of feeding frequency on growth performance, blood metabolites, proximate composition and digestive enzymes of Lebranche mullet (Mugil liza) juveniles. Aquac. Res. 2020, 51, 1162–1169. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, X.; Lai, J.; Liu, Y.; Song, M.; Li, F.; Gong, Q. Comprehensive transcriptome analysis reveals the effect of feeding rhythm on the immunity and metabolism of Acipenser dabryanus. Fish Shellfish Immunol. 2022, 122, 276–287. [Google Scholar] [CrossRef]
- Gao, X.; Wang, X.; Xinyi, W.; Hongxu, L.; Liang, X.; Huang, B.; Meng, X.-S.; Zhang, T.; Chen, H.-B.; Xing, R.; et al. Effects of different feeding frequencies on the growth, plasma biochemical parameters, stress status, and gastric evacuation of juvenile tiger puffer fish (Takifugu rubripes). Aquaculture 2022, 548, 737718. [Google Scholar] [CrossRef]
- Gilannejad, N.; Moyano, F.J.; Martínez-Rodríguez, G.; Yúfera, M. The digestive function of gilthead seabream juveniles in relation to feeding frequency. Aquaculture 2021, 531, 735867. [Google Scholar] [CrossRef]
- Zhou, Y.-L.; Wang, Z.-W.; Guo, X.-F.; Wu, J.-J.; Lu, W.-J.; Zhou, L.; Gui, J.-F. Construction of a high-density genetic linkage map and fine mapping of QTLs for growth and sex-related traits in red-tail catfish (Hemibagrus wyckioides). Aquaculture 2021, 531, 735892. [Google Scholar] [CrossRef]
- Prangthip, P.; Nongyao, M.; Sungvien, N. Culture of red-tail mystus, Hemibagrus wyckioides, in earthen ponds with different stocking densities. In Proceedings of the 7th Technical Symposium on Mekong Fisheries, Ubon Ratchathani, Thailand, 15–17 November 2005; pp. 15–17. [Google Scholar]
- Deng, J.; Zhang, X.; Sun, Y.; Zhang, L.; Mi, H. Optimal dietary lipid requirement for juvenile Asian red-tailed catfish (Hemibagrus wyckioides). Aquac. Rep. 2021, 20, 100666. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, H.; Zhang, J.; Lin, B.; Chen, L.; Wang, Q.; Li, G.; Deng, J. Assessment of rapeseed meal as fish meal alternative in diets for juvenile Asian red-tailed catfish (Hemibagrus wyckioides). Aquac. Rep. 2020, 18, 100497. [Google Scholar] [CrossRef]
- Zhou, Y.-L.; Wu, J.-J.; Wang, Z.-W.; Li, G.-H.; Zhou, L.; Gui, J.-F. Microsatellite polymorphism and genetic differentiation of different populations screened from genome survey sequencing in red-tail catfish (Hemibagrus wyckioides). Aquac. Rep. 2021, 19, 100614. [Google Scholar] [CrossRef]
- Zhu, B.-P.; Zhou, J.; Zhang, J.; Xu, S.; Fu, G.; Dai, J.; Cai, M.; Hu, Y. Dietary enzymatic rice protein and enzymatic fish paste affect the growth, muscle development and quality traits of juvenile channel catfish (Ictalurus punctatus). Aquaculture 2022, 559, 738425. [Google Scholar] [CrossRef]
- Cunniff, P.; Washington, D. Official Methods of Analysis of AOAC International; AOAC International: Rockville, MD, USA, 1997. [Google Scholar]
- Fu, G.; Wang, J.; Cai, M.; Zhang, J.; Hu, H.; Dai, C.; Zheng, X.; Hu, Y.; Chen, K. Beneficial effects of oleanolic acid on hepatopancreas oxidative stress and intestinal microbiota structure of red swamp crayfish (Procambarus clarkia). Aquac. Rep. 2023, 33, 101797. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Dai, T.; Wen, D.; Bates, C.T.; Wu, L.; Guo, X.; Liu, S.; Su, Y.; Lei, J.; Zhou, J.; Yang, Y. Nutrient supply controls the linkage between species abundance and ecological interactions in marine bacterial communities. Nat. Commun. 2022, 13, 175. [Google Scholar] [CrossRef]
- Zheng, S.; Qin, W.; Chen, T.; Ouyang, R.; Wang, X.; Li, Q.; Zhao, Y.; Liu, X.; Wang, D.; Zhou, L.; et al. Strategy for comprehensive detection and annotation of gut microbiota-related metabolites based on liquid chromatography-high-resolution mass spectrometry. Anal. Chem. 2024, 96, 2206–2216. [Google Scholar] [CrossRef]
- Biswas, G.; Thirunavukkarasu, A.R.; Sundaray, J.K.; Kailasam, M. Optimization of feeding frequency of Asian seabass (Lates calcarifer) fry reared in net cages under brackishwater environment. Aquaculture 2010, 305, 26–31. [Google Scholar] [CrossRef]
- Guo, Z.; Cui, J.; Li, M.; Liu, H.; Zhang, M.; Meng, F.; Shi, G.; Wang, R.; He, X.; Zhao, Y. Effect of feeding frequency on growth performance, antioxidant status, immune response and resistance to hypoxia stress challenge on juvenile dolly varden char Salvelinus malma. Aquaculture 2018, 486, 197–201. [Google Scholar] [CrossRef]
- Xie, F.; Ai, Q.; Mai, K.; Xu, W.; Ma, H. The optimal feeding frequency of large yellow croaker (Pseudosciaena crocea, Richardson) larvae. Aquaculture 2011, 311, 162–167. [Google Scholar] [CrossRef]
- Sveier, H.; Lied, E. The effect of feeding regime on growth, feed utilisation and weight dispersion in large Atlantic salmon (Salmo salar) reared in seawater. Aquaculture 1998, 165, 333–345. [Google Scholar] [CrossRef]
- Busti, S.; Bonaldo, A.; Dondi, F.; Cavallini, D.; Yúfera, M.; Gilannejad, N.; Moyano, F.J.; Gatta, P.P.; Parma, L. Effects of different feeding frequencies on growth, feed utilisation, digestive enzyme activities and plasma biochemistry of gilthead sea bream (Sparus aurata) fed with different fishmeal and fish oil dietary levels. Aquaculture 2020, 529, 735616. [Google Scholar] [CrossRef]
- Jafari, N.; Falahatkar, B.; Sajjadi, M.M. The effect of feeding strategies and body weight on growth performance and hematological parameters of Siberian sturgeon (Acipenser baerii, Brandt 1869): Preliminary results. J. Appl. Ichthyol. 2019, 35, 289–295. [Google Scholar] [CrossRef]
- Perswani, P.; Ismail, S.M.; Mumtaz, H.; Uddin, N.; Asfand, M.; Khalil, A.B.B.; Ijlal, A.; Khan, S.E.; Usman, M.; Younas, H.; et al. Rethinking HDL-C: An in-depth narrative review of its role in cardiovascular health. Curr. Probl. Cardiol. 2024, 49, 102152. [Google Scholar] [CrossRef] [PubMed]
- Bihari, S.; Bannard-Smith, J.; Bellomo, R. Albumin as a drug: Its biological effects beyond volume expansion. Crit. Care Resusc. 2020, 22, 257–265. [Google Scholar] [CrossRef]
- Yang, C.; Liao, A.-M.; Cui, Y.; Yu, G.; Hou, Y.; Pan, L.; Chen, W.; Zheng, S.; Li, X.; Ma, J.; et al. Wheat embryo globulin protects against acute alcohol-induced liver injury in mice. Food Chem. Toxicol. 2021, 153, 112240. [Google Scholar] [CrossRef]
- Zhu, J.; Zhao, Y.; Yang, J.; Li, Y.; Chen, P.; Qu, H. Feeding frequency affects liver health in largemouth bronze gudgeon Coreius guichenoti: Implications for lipid metabolism, oxidative stress, and inflammation response. Aquac. Rep. 2024, 35, 101941. [Google Scholar] [CrossRef]
- Shi, Y.; Hu, Y.; Wang, Z.; Zhou, J.; Zhang, J.; Zhong, H.; Fu, G.; Zhong, L. The protective effect of taurine on oxidized fish-oil-induced liver oxidative stress and intestinal barrier-function impairment in juvenile Ictalurus punctatus. Antioxidants 2021, 10, 1690. [Google Scholar] [CrossRef]
- Fabrikov, D.; Sánchez-Muros, M.J.; Barroso, F.G.; Tomás-Almenar, C.; Melenchón, F.; Hidalgo, M.C.; Morales, A.E.; Rodriguez-Rodriguez, M.; Montes-Lopez, J. Comparative study of growth performance and amino acid catabolism in Oncorhynchus mykiss, Tinca tinca and Sparus aurata and the catabolic changes in response to insect meal inclusion in the diet. Aquaculture 2020, 529, 735731. [Google Scholar] [CrossRef]
- Ding, Z.; Kong, Y.; Zhang, Y.; Li, J.; Cao, F.; Zhou, J.; Ye, J. Effect of feeding frequency on growth, body composition, antioxidant status and mRNA expression of immunodependent genes before or after ammonia-N stress in juvenile oriental river prawn, Macrobrachium nipponense. Fish Shellfish Immunol. 2017, 68, 428–434. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, S.; Nie, Z.; Li, Q.; Sun, Y.; Shao, N.; Gao, J.; Hu, J.; Xu, P.; Xu, G. Optimum feeding frequency of juvenile largemouth bass (Micropterus salmoides) reared in in-pond raceway recirculating culture system. Fish Physiol. Biochem. 2020, 46, 2197–2212. [Google Scholar] [CrossRef]
- Tian, H.-Y.; Zhang, D.-D.; Li, X.-F.; Zhang, C.-N.; Qian, Y.; Liu, W.-B. Optimum feeding frequency of juvenile blunt snout bream Megalobrama amblycephala. Aquaculture 2015, 437, 60–66. [Google Scholar] [CrossRef]
- Su, X.; Ji, D.; Yao, J.; Zou, Y.; Yan, M. Comparative analysis of intestinal characteristics of largemouth bass (Micropterus salmoides) and intestinal flora with different growth rates. Fishes 2022, 7, 65. [Google Scholar] [CrossRef]
- Holt, C.C.; Bass, D.; Stentiford, G.D.; van der Giezen, M. Understanding the role of the shrimp gut microbiome in health and disease. J. Invertebr. Pathol. 2021, 186, 107387. [Google Scholar] [CrossRef]
- Wang, P.; Wang, S.; Zhu, C.; Sun, Y.; Yan, Q.; Yi, G. Monascus purpureus M-32 fermented soybean meal improves the growth, immunity parameters, intestinal morphology, disease resistance, intestinal microbiota and metabolome in Pacific white shrimp (Litopenaeus vannamei). Anim. Nutr. 2024, 17, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, P.; Hou, D.; Li, M.; Chen, B.; Peng, K.; Huang, W.; Cao, J.; Zhao, H. Probiotic breakthrough: Clostridium butyricum improved the intestinal SCFAs content, digestive enzymes and antioxidant activities, gut morphology and microbiota composition of largemouth bass. Aquac. Rep. 2024, 36, 102090. [Google Scholar] [CrossRef]
- Jia, K.; Yang, N.; Zhang, X.; Cai, R.; Zhang, Y.; Tian, J.; Raza, S.H.A.; Kang, Y.; Qian, A.; Li, Y.; et al. Genomic, morphological and functional characterization of virulent bacteriophage IME-JL8 targeting Citrobacter freundii. Front. Microbiol. 2020, 11, 585261. [Google Scholar] [CrossRef]
- Deng, D.; Mu, Z.; Lv, X.; Jiang, X.; Zhou, J.; Guo, H.; Zhang, W.; Wang, B.; Lu, Y.; Wu, J.; et al. Pathogenicity of Plesiomonas shigelloides and Citrobacter freundii isolated from the endangered Chinese sturgeon (Acipenser sinensis). Microb. Pathog. 2022, 173, 105818. [Google Scholar] [CrossRef]
- Bandeira Junior, G.; Dos Santos, A.C.; Souza, C.F.; Baldissera, M.D.; Moreira, K.L.D.S.; da Veiga, M.L.; da Rocha, M.I.U.M.; de Vargas, A.P.C.; da Cunha, M.A.; Baldisserotto, B. Citrobacter freundii infection in silver catfish (Rhamdia quelen): Hematological and histological alterations. Microb. Pathog. 2018, 125, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Pan, Y.; Jin, S.; Zheng, Y.; Xu, J.; Fan, H.; Khalid, M.; Wang, Y.; Hu, M. Effects of Citrobacter freundii on sturgeon: Insights from skin mucosal immunology and microbiota. Fish Shellfish Immunol. 2024, 149, 109527. [Google Scholar] [CrossRef]
- Yilmaz, S. Effects of dietary blackberry syrup supplement on growth performance, antioxidant, and immunological responses, and resistance of Nile tilapia, Oreochromis niloticus to Plesiomonas shigelloides. Fish Shellfish Immunol. 2019, 84, 1125–1133. [Google Scholar] [CrossRef]
- Evans, J.J.; Klesius, P.H.; Shoemaker, C.A. First isolation and characterization of Lactococcus garvieae from Brazilian Nile tilapia, Oreochromis niloticus (L.), and pintado, Pseudoplathystoma corruscans (Spix & Agassiz). J. Fish Dis. 2009, 32, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Shahin, K.; Veek, T.; Heckman, T.I.; Littman, E.; Mukkatira, K.; Adkison, M.; Welch, T.J.; Imai, D.M.; Pastenkos, G.; Camus, A.; et al. Isolation and characterization of Lactococcus garvieae from rainbow trout, Onchorhyncus mykiss, from California, USA. Transbound. Emerg. Dis. 2022, 69, 2326–2343. [Google Scholar] [CrossRef] [PubMed]
- Egger, R.C.; Rosa, J.C.C.; Resende, L.F.L.; de Pádua, S.B.; de Oliveira Barbosa, F.; Zerbini, M.T.; Tavares, G.C.; Figueiredo, H.C.P. Emerging fish pathogens Lactococcus petauri and L. garvieae in Nile tilapia (Oreochromis niloticus) farmed in Brazil. Aquaculture 2023, 565, 739093. [Google Scholar] [CrossRef]
- Pishesha, N.; Harmand, T.J.; Ploegh, H.L. A guide to antigen processing and presentation. Nat. Rev. Immunol. 2022, 22, 751–764. [Google Scholar] [CrossRef]
- Wishart, D.S. Metabolomics for investigating physiological and pathophysiological processes. Physiol. Rev. 2019, 99, 1819–1875. [Google Scholar] [CrossRef]
- Iside, C.; Scafuro, M.; Nebbioso, A.; Altucci, L. SIRT1 activation by natural phytochemicals: An overview. Front. Pharmacol. 2020, 11, 1225. [Google Scholar] [CrossRef]
- Aşkın, Ö.; Uzunçakmak, T.K.Ü.; Altunkalem, N.; Tüzün, Y. Vitamin deficiencies/hypervitaminosis and the skin. Clin. Dermatol. 2021, 39, 847–857. [Google Scholar] [CrossRef]
- Yoshii, K.; Hosomi, K.; Sawane, K.; Kunisawa, J. Metabolism of dietary and microbial vitamin B family in the regulation of host immunity. Front. Nutr. 2019, 6, 48. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zheng, S.; Wu, G. Nutrition and functions of amino acids in fish. Adv. Exp. Med. Biol. 2021, 1285, 133–168. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wang, X.; Wang, X.; Li, H.; Xu, L.; Fang, Y.; Cao, S.; Huang, B.; Chen, H.; Xing, R.; et al. Effect of winter feeding frequency on growth performance, biochemical blood parameters, oxidative stress, and appetite-related genes in Takifugu rubripes. Fish Physiol. Biochem. 2022, 48, 1167–1181. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter 1 | F2 | F3 | F4 | p Value |
|---|---|---|---|---|
| FCR | 1.73 ± 0.142 ab | 1.48 ± 0.111 b | 1.85 ± 0.181 a | 0.048 |
| SGR (%/d) | 1.47 ± 0.002 ab | 1.67 ± 0.002 a | 1.37 ± 0.003 b | 0.044 |
| WGR (%) | 114.17 ± 0.191 ab | 136.35 ± 0.093 a | 102.22 ± 0.071 b | 0.043 |
| SR (%) | 99.33 ± 0.011 | 99.33 ± 0.013 | 99.33 ± 0.014 | 0.999 |
| HSI (%) | 1.81 ± 0.002 | 1.96 ± 0.001 | 1.91 ± 0.003 | 0.709 |
| VSI (%) | 11.82 ± 0.021 | 11.46 ± 0.013 | 11.69 ± 0.021 | 0.878 |
| CF (g/cm3) | 0.01 ± 0.001 | 0.01 ± 0.002 | 0.01 ± 0.001 | 0.531 |
| Moisture (%) | 69.78 ± 0.001 | 69.90 ± 0.012 | 69.89 ± 0.002 | 0.942 |
| Crude protein (%) | 17.82 ± 0.013 | 17.67 ± 0.001 | 17.81 ± 0.012 | 0.901 |
| Crude lipid (%) | 8.08 ± 0.003 | 8.53 ± 0.009 | 8.15 ± 0.002 | 0.548 |
| Crude ash (%) | 2.64 ± 0.003 | 2.79 ± 0.002 | 2.72 ± 0.003 | 0.713 |
| Parameter 1 | F2 | F3 | F4 | p Value |
|---|---|---|---|---|
| TC (mmol/L) | 2.84 ± 1.034 b | 3.30 ± 0.791 b | 4.54 ± 0.971 a | 0.023 |
| TG (mmol/L) | 12.20 ± 1.634 | 14.05 ± 2.821 | 14.42 ± 0.472 | 0.371 |
| HDL-C (mmol/L) | 3.09 ± 0.574 b | 4.37 ± 0.252 a | 3.41 ± 0.662 ab | 0.049 |
| LDL-C (mmol/L) | 5.26 ± 0.431 | 5.06 ± 0.594 | 5.61 ± 0.374 | 0.406 |
| GLU (mmol/L) | 4.64 ± 0.205 | 5.04 ± 0.835 | 4.20 ± 0.342 | 0.253 |
| TP (g/L) | 29.82 ± 4.306 | 32.88 ± 1.338 | 28.93 ± 2.471 | 0.301 |
| ALB (g/L) | 13.42 ± 1.711 | 14.82 ± 3.213 | 13.05 ± 2.312 | 0.673 |
| Parameter 1 | F2 | F3 | F4 | p Value |
|---|---|---|---|---|
| T-AOC (mmol/g prot) | 0.15 ± 0.023 b | 0.17 ± 0.021 b | 0.23 ± 0.042 a | 0.026 |
| MDA (nmol/mg prot) | 0.75 ± 0.092 b | 0.60 ± 0.101 b | 0.97 ± 0.151 a | 0.001 |
| SOD (U/mg prot) | 7.78 ± 1.661 b | 6.60 ± 0.713 b | 10.15 ± 0.883 a | 0.025 |
| GSH-PX (U/mg prot) | 34.74 ± 4.892 b | 35.29 ± 7.421 b | 55.62 ± 12.412 a | 0.044 |
| ALT (U/g prot) | 3.46 ± 0.511 b | 9.36 ± 0.472 a | 3.77 ± 1.031 b | 0.001 |
| AST (U/g prot) | 42.31 ± 5.361 b | 71.17 ± 24.592 ab | 77.83 ± 3.492 a | 0.053 |
| ALP (King unit/g prot) | 1.62 ± 0.734 a | 0.46 ± 0.291 b | 2.22 ± 0.221 a | 0.004 |
| Parameter | F2 | F3 | F4 | p Value |
|---|---|---|---|---|
| trypsin (U/mg prot) | 2.24 ± 0.991 a | 3.30 ± 0.382 b | 4.27 ± 0.611 c | 0.004 |
| amylase (U/mg prot) | 1.09 ± 0.972 | 1.19 ± 0.774 | 1.00 ± 0.621 | 0.937 |
| lipase (U/g prot) | 38.51 ± 8.592 b | 84.31 ± 14.819 a | 49.50 ± 1.032 b | 0.002 |
| Parameter 1 | F2 | F3 | F4 | p Value |
|---|---|---|---|---|
| GCs (number) | 31.80 ± 12.261 | 25.67 ± 9.363 | 24.57 ± 8.048 | 0.359 |
| MT (mm) | 0.13 ± 0.028 | 0.16 ± 0.022 | 0.14 ± 0.032 | 0.275 |
| VL (mm) | 0.39 ± 0.071 | 0.44 ± 0.072 | 0.35 ± 0.082 | 0.122 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, B.; Wen, Z.; Zheng, C.; Tang, S.; Xiao, T.; Qiu, Y.; Liu, Q. Feeding Frequency Affects the Growth Performance and Intestinal Health of Juvenile Red-Tail Catfish (Hemibagrus wyckioides) with the Same Amount of Daily Feed. Animals 2025, 15, 1621. https://doi.org/10.3390/ani15111621
Xu B, Wen Z, Zheng C, Tang S, Xiao T, Qiu Y, Liu Q. Feeding Frequency Affects the Growth Performance and Intestinal Health of Juvenile Red-Tail Catfish (Hemibagrus wyckioides) with the Same Amount of Daily Feed. Animals. 2025; 15(11):1621. https://doi.org/10.3390/ani15111621
Chicago/Turabian StyleXu, Baohong, Zheyu Wen, Chen Zheng, Shengguo Tang, Tiaoyi Xiao, Yaqun Qiu, and Qiaolin Liu. 2025. "Feeding Frequency Affects the Growth Performance and Intestinal Health of Juvenile Red-Tail Catfish (Hemibagrus wyckioides) with the Same Amount of Daily Feed" Animals 15, no. 11: 1621. https://doi.org/10.3390/ani15111621
APA StyleXu, B., Wen, Z., Zheng, C., Tang, S., Xiao, T., Qiu, Y., & Liu, Q. (2025). Feeding Frequency Affects the Growth Performance and Intestinal Health of Juvenile Red-Tail Catfish (Hemibagrus wyckioides) with the Same Amount of Daily Feed. Animals, 15(11), 1621. https://doi.org/10.3390/ani15111621