
Locomotion, Postures, and Substrate Use in Captive Southern Pygmy Slow Lorises (Strepsirrhini, Primates): Implications for Conservation

 , , and
, , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Animals and Experimental Setting
2.2. Data Collection
2.3. Data Analysis
3. Results
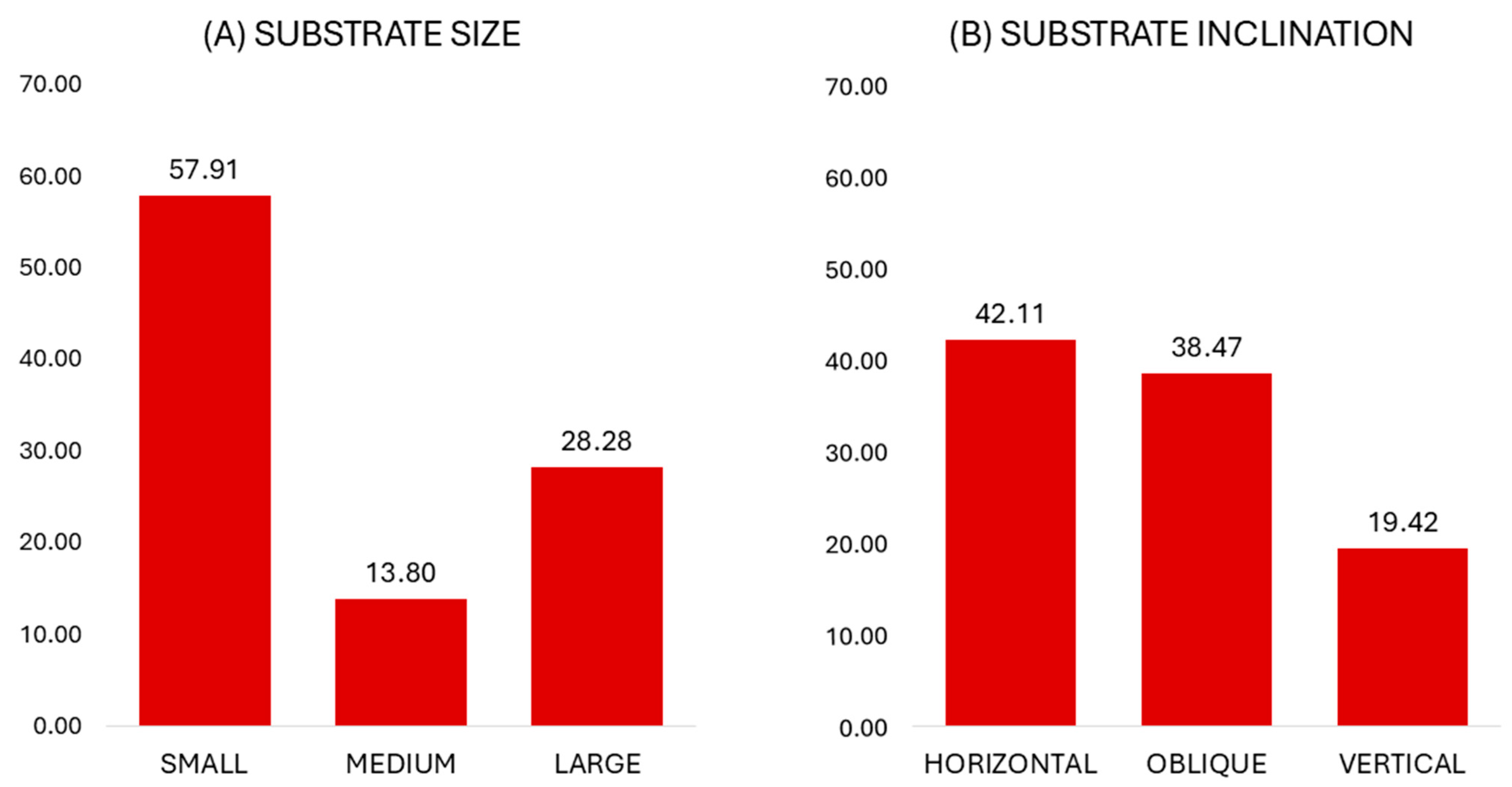
3.1. Substrate Use
3.2. General Behavior
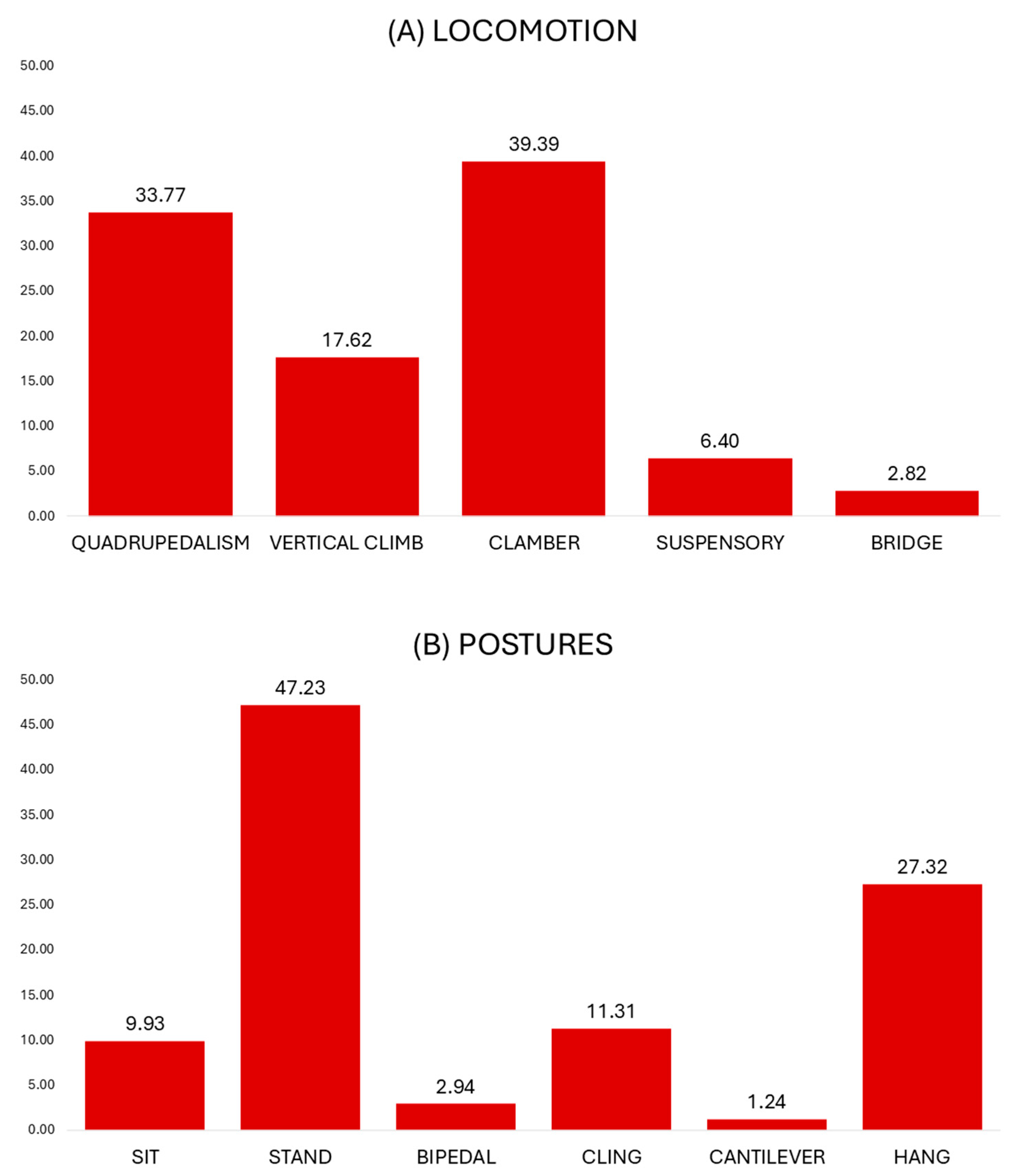
3.3. Locomotor Behavior
3.4. Postural Behavior
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prost, J.H. A Definitional System for the Classification of Primate Locomotion. Am. Anthropol. 1965, 67, 1198–1214. [Google Scholar] [CrossRef]
- Saunders, E.L.R.; Roberts, A.M.; Thorpe, S.K.S. Positional Behavior. In The International Encyclopedia of Primatology; Fuentes, A., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017. [Google Scholar] [CrossRef]
- Mekonnen, A.; Fashing, P.J.; Sargis, E.J.; Venkataraman, V.V.; Bekele, A.; Hernandez-Aguilar, R.A.; Rueness, E.K.; Stenseth, N.C. Flexibility in Positional Behavior, Strata Use, and Substrate Utilization among Bale Monkeys (Chlorocebus djamdjamensis) in Response to Habitat Fragmentation and Degradation. Am. J. Primatol. 2018, 80, e22760. [Google Scholar] [CrossRef]
- Ma, C.; Fan, P. Effect of Substrate Type on Langur Positional Repertoire. Glob. Ecol. Conserv. 2020, 22, e00956. [Google Scholar] [CrossRef]
- Clemente, C.J.; Dick, T.J.M.; Wheatley, R.; Gaschk, J.; Nasir, A.F.A.A.; Cameron, S.F.; Wilson, R.S. Moving in Complex Environments: A Biomechanical Analysis of Locomotion on Inclined and Narrow Substrates. J. Exp. Biol. 2019, 222, jeb189654. [Google Scholar] [CrossRef]
- Aronsen, G.P. Conservation Applications of Positional Behavior, Support Use, and Forest Structure Data. Am. J. Phys. Anthropol. 2005, 126, 66. [Google Scholar]
- Nekaris, K.A.-I.; Nijman, V. A New Genus Name for Pygmy Lorises, Xanthonycticebus Gen. Nov. (Mammalia, Primates). Zoosystematics Evol. 2022, 98, 87–92. [Google Scholar] [CrossRef]
- Blair, M.E.; Cao, G.T.H.; López-Nandam, E.H.; Veronese-Paniagua, D.A.; Birchette, M.G.; Kenyon, M.; Md-Zain, B.M.; Munds, R.A.; Nekaris, K.A.I.; Nijman, V.; et al. Molecular Phylogenetic Relationships and Unveiling Novel Genetic Diversity among Slow and Pygmy Lorises, Including Resurrection of Xanthonycticebus intermedius. Genes 2023, 14, 643. [Google Scholar] [CrossRef]
- Blair, M.; Nadler, T.; Ni, O.; Samun, E.; Streicher, U.; Nekaris, K. Nycticebus pygmaeus (Amended Version of 2020 Assessment). The IUCN Red List of Threatened Species: E. T14941A198267330. 2021. Available online: https://doi.org/10.2305/IUCN.UK.2021-2.RLTS.T14941A198267330.en (accessed on 30 April 2025).
- Streicher, U. Aspects of Ecology and Conservation of the Pygmy Loris Nycticebus pygmaeus in Vietnam. Ph.D. Dissertation, Ludwig-Maximilians-Universität München, München, Germany, 2004. [Google Scholar]
- Starr, C.; Nekaris, K.; Streicher, U.; Leung, L. Traditional Use of Slow Lorises Nycticebus bengalensis and N. pygmaeus in Cambodia: An Impediment to Their Conservation. Endanger. Species Res. 2010, 12, 17–23. [Google Scholar] [CrossRef]
- Streicher, U. Morphological Data of Pygmy Lorises (Nycticebus pygmaeus). Vietnam. J. Primatol. 2007, 1, 67–74. [Google Scholar]
- Nekaris, K.; Bearder, S. The Strepsirrhine Primates of Asia and Mainland Africa: Diversity Shrouded in Darkness. In Primates in Perspective, 2nd ed.; Campbell, C., Fuentes, A., MacKinnon, K., Bearder, S., Stumpf, R., Eds.; Oxford University Press: Oxford, UK, 2011; pp. 34–54. [Google Scholar]
- Starr, C.; Nekaris, K.A.I. Obligate Exudativory Characterizes the Diet of the Pygmy Slow Loris Nycticebus pygmaeus. Am. J. Primatol. 2013, 75, 1054–1061. [Google Scholar] [CrossRef]
- Starr, C.; Nekaris, K.A.I. Ranging Patterns of the Pygmy Slow Loris (Nycticebus pygmaeus) in a Mixed Deciduous Forest in Eastern Cambodia. In Evolution, Ecology and Conservation of Lorises and Pottos; Nekaris, K.A.I., Burrows, A.M., Eds.; Cambridge Studies in Biological and Evolutionary Anthropology; Cambridge University Press: Cambridge, UK, 2020; pp. 228–234. [Google Scholar]
- Yamanashi, Y.; Nemoto, K.; Alejandro, J. Social Relationships among Captive Male Pygmy Slow Lorises (Nycticebus pygmaeus): Is Forming Male Same-Sex Pairs a Feasible Management Strategy? Am. J. Primatol. 2021, 83, e23233. [Google Scholar] [CrossRef] [PubMed]
- Poindexter, S.A.; Nekaris, K.A.I. The Evolution of Social Organisation in Lorisiformes. In Evolution, Ecology and Conservation of Lorises and Pottos; Nekaris, K.A.I., Burrows, A.M., Eds.; Cambridge Studies in Biological and Evolutionary Anthropology; Cambridge University Press: Cambridge, UK, 2020; pp. 129–137. [Google Scholar]
- Nekaris, K.A.-I.; Moore, R.S.; Rode, E.J.; Fry, B.G. Mad, Bad and Dangerous to Know: The Biochemistry, Ecology and Evolution of Slow Loris Venom. J. Venom. Anim. Toxins Trop. Dis. 2013, 19, 21. [Google Scholar] [CrossRef]
- Nekaris, K.A.I.; Campera, M.; Nijman, V.; Birot, H.; Rode-Margono, E.J.; Fry, B.G.; Weldon, A.; Wirdateti, W.; Imron, M.A. Slow Lorises Use Venom as a Weapon in Intraspecific Competition. Curr. Biol. 2020, 30, R1252–R1253. [Google Scholar] [CrossRef] [PubMed]
- Grand, T.I. The Functional Anatomy of the Ankle and Foot of the Slow Loris (Nycticebus coucang). Am. J. Phys. Anthropol. 1967, 26, 207–218. [Google Scholar] [CrossRef]
- Cartmill, M.; Milton, K. The Lorisiform Wrist Joint and the Evolution of “Brachiating” Adaptations in the Hominoidea. Am. J. Phys. Anthropol. 1977, 47, 249–272. [Google Scholar] [CrossRef]
- McArdle, J.E. Functional Morphology of the Hip and Thigh of the Lorisiformes. Contrib. Primatol. 1981, 17, 1–132. [Google Scholar]
- Gebo, D.L. Postcranial Adaptation and Evolution in Lorisidae. Primates 1989, 30, 347–367. [Google Scholar] [CrossRef]
- Curtis, D.J. Functional Anatomy of the Trunk Musculature in the Slow Loris (Nycticebus coucang). Am. J. Phys. Anthropol. 1995, 97, 367–379. [Google Scholar] [CrossRef]
- Runestad, J.A. Postcranial Adaptations for Climbing in Loridae (Primates). J. Zool. 1997, 242, 261–290. [Google Scholar] [CrossRef]
- Shapiro, L.J.; Demes, B.; Cooper, J. Lateral Bending of the Lumbar Spine during Quadrupedalism in Strepsirhines. J. Hum. Evol. 2001, 40, 231–259. [Google Scholar] [CrossRef]
- Goto, R.; Kumakura, H. The Estimated Mechanical Advantage of the Prosimian Ankle Joint Musculature, and Implications for Locomotor Adaptation. J. Anat. 2013, 222, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Gyambibi, A.; Lemelin, P. Comparative and Quantitative Myology of the Forearm and Hand of Prosimian Primates. Anat. Rec. 2013, 296, 1196–1206. [Google Scholar] [CrossRef]
- Nekaris, K.A.I. Extreme Primates: Ecology and Evolution of Asian Lorises. Evol. Anthropol. 2014, 23, 177–187. [Google Scholar] [CrossRef]
- Yapuncich, G.S.; Granatosky, M.C. Footloose: Articular Surface Morphology and Joint Movement Potential in the Ankles of Lorisids and Cheirogaleids. Am. J. Phys. Anthropol. 2021, 175, 876–894. [Google Scholar] [CrossRef]
- Dykyj, D. Locomotion of the Slow Loris in a Designed Substrate Context. Am. J. Phys. Anthropol. 1980, 52, 577–586. [Google Scholar] [CrossRef]
- Glassman, D.M.; Wells, J.P. Positional and Activity Behavior in a Captive Slow Loris: A Quantitative Assessment. Am. J. Primatol. 1984, 7, 121–132. [Google Scholar] [CrossRef]
- Demes, B.; Jungers, W.L.; Nieschalk, U. Size-and Speed-Related Aspects of Quadrupedal Walking in Slender and Slow Lorises. In Gravity, Posture and Locomotion in Primates; Jouffroy, F.K., Stack, M.H., Niemiz, C., Eds.; Il Sedicesimo: Firenze, Italy, 1990; pp. 175–197. [Google Scholar]
- Ishida, H.; Hirasaki, E.; Matano, S. Locomotion of the Slow Loris between Discontinuous Substrates. Top. Primatol. 1992, 3, 139–152. [Google Scholar]
- Nekaris, K.A.I. Activity Budget and Positional Behavior of the Mysore Slender Loris (Loris tardigradus lydekkerianus): Implications for Slow Climbing Locomotion. Folia Primatol. 2001, 72, 228–241. [Google Scholar] [CrossRef]
- Nekaris, K.A.I.; Jayewardene, J. Pilot Study and Conservation Status of the Slender Loris (Loris tardigradus and L. lydekkerianus) in Sri Lanka. Primate Conserv. 2003, 19, 83–90. [Google Scholar]
- Schmitt, D.; Lemelin, P. Locomotor Mechanics of the Slender Loris (Loris tardigradus). J. Hum. Evol. 2004, 47, 85–94. [Google Scholar] [CrossRef]
- Hanna, J.B. Kinematics of Vertical Climbing in Lorises and Cheirogaleus Medius. J. Hum. Evol. 2006, 50, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Nekaris, K.A.I.; Stevens, N.J. Not All Lorises Are Slow: Rapid Arboreal Locomotion in Loris tardigradus of Southwestern Sri Lanka. Am. J. Primatol. 2007, 69, 112–120. [Google Scholar] [CrossRef]
- Poindexter, S.A.; Nekaris, K.A.I. Vertical Clingers and Gougers: Rapid Acquisition of Adult Limb Proportions Facilitates Feeding Behaviours in Young Javan Slow Lorises (Nycticebus javanicus). Mamm. Biol. 2017, 87, 40–49. [Google Scholar] [CrossRef]
- Das, N.; Nekaris, K.A.I. Positional Behaviour and Substrate Preference of Slow Lorises, with a Case Study of Nycticebus bengalensis in Northeast India. In Evolution, Ecology and Conservation of Lorises and Pottos; Nekaris, K.A.I., Burrows, A.M., Eds.; Cambridge Studies in Biological and Evolutionary Anthropology; Cambridge University Press: Cambridge, UK, 2020; pp. 210–218. [Google Scholar]
- Alejandro, J.; Yamanashi, Y.; Nemoto, K.; Bercovitch, F.B.; Huffman, M.A. Behavioral Changes of Solitary Housed Female Pygmy Slow Lorises (Nycticebus pygmeaus) after Introduction into Group Enclosures. Animals 2021, 11, 2751. [Google Scholar] [CrossRef]
- IUCN. IUCN SSC Position Statement on Research Involving Species at Risk of Extinction; IUCN Species Survival Commission (SSC): Abu Dhabi, United Arab Emirates, 1998. [Google Scholar]
- IPS/ASP. Code of Best Practices for Field Primatology; International Primatological Society: Washington, DC, USA, 2014. [Google Scholar]
- ASAB/ABS. Guidelines for the Treatment of Animals in Behavioural Research and Teaching. Anim. Behav. 2012, 83, 301–309. [Google Scholar] [CrossRef]
- Daschbach, N.J.; Schein, M.W.; Haines, D.E. Cage-Size Effects on Locomotor, Grooming and Agonistic Behaviours of the Slow Loris, Nycticebus coucang (Primates, Lorisidae). Appl. Anim. Ethol. 1983, 9, 317–330. [Google Scholar] [CrossRef]
- Jacobs, J. Quantitative Measurement of Food Selection. Oecologia 1974, 14, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Bateson, P. Measuring Behaviour: An Introductory Guide, 2nd ed.; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Dagosto, M. Testing Positional Behavior of Malagasy Lemurs: A Randomization Approach. Am. J. Phys. Anthropol. 1994, 94, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Dawkins, M.S. Observing Animal Behaviour: Design and Analysis of Quantitative Data, 1st ed.; Oxford University Press: Oxford, UK, 2007; ISBN 0-19-856935-1. [Google Scholar]
- Huy Huynh, D. Ecology, Biology and Conservation Status of Prosimian Species in Vietnam. Folia Primatol. 1998, 69, 101–108. [Google Scholar] [CrossRef]
- Starr, C.; Nekaris, K.A.I.; Streicher, U.; Leung, L.K.-P. Field Surveys of the Vulnerable Pygmy Slow Loris Nycticebus pygmaeus Using Local Knowledge in Mondulkiri Province, Cambodia. Oryx 2011, 45, 135–142. [Google Scholar] [CrossRef]
- Nguyen, A.T.; Van Nguyen, T.; Timmins, R.; McGowan, P.; Van Hoang, T.; Le, M.D. Efficacy of Camera Traps in Detecting Primates in Hue Saola Nature Reserve. Primates J. Primatol. 2020, 61, 697–705. [Google Scholar] [CrossRef]
- Tenaza, R.; Ross, B.A.; Tanticharoenyos, P.; Berkson, G. Individual Behaviour and Activity Rhythms of Captive Slow Lorises (Nycticebus coucang). Anim. Behav. 1969, 17, 664–669. [Google Scholar] [CrossRef]
- Ruf, T.; Streicher, U.; Stalder, G.L.; Nadler, T.; Walzer, C. Hibernation in the Pygmy Slow Loris (Nycticebus pygmaeus): Multiday Torpor in Primates Is Not Restricted to Madagascar. Sci. Rep. 2015, 5, 17392. [Google Scholar] [CrossRef]
- Tan, C.L.; Drake, J.H. Evidence of Tree Gouging and Exudate Eating in Pygmy Slow Lorises (Nycticebus pygmaeus). Folia Primatol. 2001, 72, 37–39. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.D.; Nekaris, K.A.I. Behaviour and Habitat Use of the Bengal Slow Loris Nycticebus bengalensis in the Dry Dipterocarp Forests of Phnom Samkos Wildlife Sanctuary, Cambodia. Cambodian J. Nat. Hist. 2011, 2011, 104–133. [Google Scholar]
- Nekaris, K.A.I.; Blackham, G.V.; Nijman, V. Conservation Implications of Low Encounter Rates of Five Nocturnal Primate Species (Nycticebus spp.) in Asia. Biodivers. Conserv. 2008, 17, 733–747. [Google Scholar] [CrossRef]
- Birot, H.; Campera, M.; Imron, M.A.; Nekaris, K.A.I. Artificial Canopy Bridges Improve Connectivity in Fragmented Landscapes: The Case of Javan Slow Lorises in an Agroforest Environment. Am. J. Primatol. 2020, 82, e23076. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Substrate Type | |
|---|---|
| Branch | Network of branches of different diameters |
| Artificial | Wooden nest box, ledges, feeders |
| Ground | Enclosure floor |
| Substrate size (relative to foot span [12]) | |
| Small | Substrate fully grasped by a completely flexed foot (<20 mm in diameter) |
| Medium | Substrate partly grasped by a semi-extended foot (20–50 mm in diameter) |
| Large | Substrate cannot be grasped by a fully extended foot (>50 mm in diameter) |
| Substrate inclination | |
| Horizontal | Angle between 0° and 22.5° |
| Oblique | Angle between 22.5° and 67.5° |
| Vertical | Angle between 67.5° and 90° |
| Behavior | |
| Move | Body displacement through its surroundings |
| Feed | Search, acquisition, and processing of food items |
| Rest/pause | Inactivity for short or long periods |
| Groom | Care of external body surface on the same (auto-grooming) or different animal (allo-grooming) |
| Social | Social interactions between two or more individuals (e.g., spatial proximity, play, cooperation, agonistic encounters, vocal communication, etc.) |
| Locomotor modes | |
| Quadrupedal walking | Symmetrical slow/moderate (walk) and fast (run) progression along single horizontal and moderately inclined substrates |
| Vertical Climbing | Symmetrical quadrupedal ascent or descent along vertical or steeply inclined substrates |
| Clambering | Irregular pronograde or semi-pronograde quadrupedal locomotion across multiple substrates |
| Bridging | Gap crossing mode involving reaching action across distantly located substrates with irregular limb movements |
| Suspensory locomotion | Supinograde symmetrical slow/moderate or irregular asymmetrical below branch locomotion employing all four limbs |
| Postural modes | |
| Standing | Pronograde tri/quadrupedal posture with flexed, semi-extended, or fully extended limbs |
| Sitting | Orthograde or leaning seated posture with strongly flexed hind limbs |
| Bipedal | Above-branch standing on two moderately flexed limbs assisted by forelimbs |
| Clinging | Orthograde grasping posture with extremely flexed limbs and the head upwards or downwards on vertical or steeply inclined substrates |
| Cantilevering | Grasping feet secure the lower part of the body to a steeply inclined substrate while the trunk and forelimbs are extended horizontally |
| Hanging | Suspensory posture below a substrate, with the body in a flexible supinograde or semi-pronograde position, using all combinations of fore- and hindlimbs |
| Move | Rest | Feed | Groom | Social | |
|---|---|---|---|---|---|
| Substrate Size | (%) | (%) | (%) | (%) | (%) |
| Small | 45.24 | 32.66 | 42.65 | 24.96 | 19.25 |
| Medium | 24.52 | 24.43 | 27.49 | 11.29 | 16.26 |
| Large | 30.24 | 42.91 | 29.85 | 63.75 | 80.79 |
| Substrate inclination | (%) | (%) | (%) | (%) | (%) |
| Horizontal | 28.52 | 40.87 | 27.49 | 66.29 | 67.49 |
| Oblique | 43.87 | 39.84 | 42.42 | 25.60 | 20.20 |
| Vertical | 27.61 | 19.28 | 30.09 | 8.11 | 12.31 |
| N | 7351 | 7826 | 2818 | 4199 | 1357 |
| Quadrupedalism | Vertical Climbing | Clambering | Suspensory | Bridging | |
|---|---|---|---|---|---|
| Substrate Size | (%) | (%) | (%) | (%) | (%) |
| Small | 24.51 | 23.22 | 72.43 | 33.33 | 44.12 |
| Medium | 22.55 | 36.81 | 22.94 | 50.62 | 32.35 |
| Large | 52.94 | 39.97 | 4.63 | 16.05 | 23.53 |
| Substrate inclination | (%) | (%) | (%) | (%) | (%) |
| Horizontal | 80.64 | 0.00 | 17.47 | 65.43 | 17.65 |
| Oblique | 19.36 | 4.26 | 58.10 | 34.57 | 52.94 |
| Vertical | 0.00 | 95.74 | 24.43 | 0.00 | 29.41 |
| N | 7953 | 4150 | 9277 | 1507 | 664 |
| Sitting | Standing | Bipedal | Clinging | Cantilevering | Hanging | |
|---|---|---|---|---|---|---|
| Substrate Size | (%) | (%) | (%) | (%) | (%) | (%) |
| Small | 22.91 | 21.31 | 25.37 | 32.95 | 67.87 | 47.46 |
| Medium | 35.24 | 13.55 | 59.70 | 28.72 | 21.42 | 33.02 |
| Large | 41.85 | 65.13 | 14.92 | 38.33 | 10.71 | 19.52 |
| Substrate inclination | (%) | (%) | (%) | (%) | (%) | (%) |
| Horizontal | 44.05 | 67.57 | 20.90 | 0.00 | 0.00 | 40.63 |
| Oblique | 55.95 | 31.71 | 79.10 | 7.72 | 28.57 | 43.50 |
| Vertical | 0.00 | 0.72 | 0.00 | 92.28 | 71.43 | 15.87 |
| N | 2340 | 11,124 | 694 | 2665 | 292 | 6436 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youlatos, D.; Pylarinos, D.; Karantanis, N.E.; Rychlik, L. Locomotion, Postures, and Substrate Use in Captive Southern Pygmy Slow Lorises (Strepsirrhini, Primates): Implications for Conservation. Animals 2025, 15, 1576. https://doi.org/10.3390/ani15111576
Youlatos D, Pylarinos D, Karantanis NE, Rychlik L. Locomotion, Postures, and Substrate Use in Captive Southern Pygmy Slow Lorises (Strepsirrhini, Primates): Implications for Conservation. Animals. 2025; 15(11):1576. https://doi.org/10.3390/ani15111576
Chicago/Turabian StyleYoulatos, Dionisios, Dimitris Pylarinos, Nikolaos Evangelos Karantanis, and Leszek Rychlik. 2025. "Locomotion, Postures, and Substrate Use in Captive Southern Pygmy Slow Lorises (Strepsirrhini, Primates): Implications for Conservation" Animals 15, no. 11: 1576. https://doi.org/10.3390/ani15111576
APA StyleYoulatos, D., Pylarinos, D., Karantanis, N. E., & Rychlik, L. (2025). Locomotion, Postures, and Substrate Use in Captive Southern Pygmy Slow Lorises (Strepsirrhini, Primates): Implications for Conservation. Animals, 15(11), 1576. https://doi.org/10.3390/ani15111576