Simple Summary
The overpopulation of stray cats and dogs is a public health issue worldwide. Surgery is the most common method of contraception; however, it may be beneficial to have a non-invasive technique for controlling the reproduction of cats and dogs. Therefore, this review offers an updated and comprehensive analysis of non-surgical contraceptive methods tested in controlled in vivo studies involving female felines and canines, focusing on the effects of reproductive variables and the duration of sterility achieved.
Abstract
The issue of stray cats and dogs is a global concern with considerable implications for animal welfare and public health. This review aims to provide an updated and comprehensive analysis of non-surgical contraceptive methods tested in studies controlled in vivo in feline and canine females. Immunocontraception via vaccination against gonadotropin-releasing hormone (GnRH), the luteinizing hormone receptor, zona pellucida proteins, and sperm, or use of viral-vectored delivery, is yet developing. Hormonal treatment (progestins, androgens, or GnRH) analogs act directly to block the reproductive axis. However, it produced essential side effects. Analogs of kisspeptin, non-steroid anti-inflammatory drugs such as firocoxib, and delivery of cytotoxins to the pituitary have shown non-conclusive results. Additional methods have also been tested, such as intraovarian injection of necrosing compounds or intravaginal and intrauterine devices. At present, neither of these methods offers permanent sterility that can replace surgical sterilization techniques. To our knowledge, none are currently authorized by the Food and Drug Administration (FDA) or the European Medicines Agency (EMA) for contraceptive methods or sterilization of cats or dogs. Therefore, it is necessary to continue the development of a compound that warrants the sterility of cats and dogs.
1. Introduction
There is an urgent need to control the rapidly growing populations of feral cats and dogs worldwide [1]. These species were domesticated 9000 to 10,000 years ago and 40,000 years ago [2]. The original purposes of domestication for these species have shifted to “companion pets” or “domestic animals,” leading to significant overpopulation issues globally and creating a serious public health concern [3]. Approximately 600 million cats and between 700 and 1000 million dogs inhabit our planet, with around 300 million stray dogs and 70 million stray cats [2,3,4,5,6]. In Mexico, the pet dog population was estimated to be around 42 million, while there were about 7 million cats [7].
The number of stray cats and dogs poses a global concern for animal welfare and public health. Some dog breeds have lost their ability to survive independently of humans, increasing the risk of disease transmission and reports of aggression. Feral cats can have an ecological impact that harms native wildlife, threatening 8% of endangered birds, mammals, and reptiles [6]. Therefore, controlling the population of cats and dogs relates to improving various aspects of society.
As a result, the World Organization for Animal Health (WOAH) has stated that controlling the population of these species is necessary [8]. It is estimated that sterilizing 76–88% of female dogs would be required [9] to halt the growth of the canine and feline populations. Canine and feline population control methods are categorized into surgical and non-surgical types. Surgical methods, such as TNR (Trap–Neuter–Return), are the most widely practiced and accepted globally. However, they have several disadvantages that significantly complicate their implementation.
Surgical contraception is a costly method that may lead to complications such as hemorrhage, bloody vulvar discharge, the onset of pseudocyesis, infection at the incision site, and peritonitis [10]. Additionally, pediatric sterilization is a controversial topic, and its effects are still being studied. Authors like Oliveira et al. (2023) and McKenzie (2010) note that this procedure can cause behavioral changes such as increased fear and shyness, and it may also predispose individuals to conditions like cranial cruciate ligament rupture or a higher incidence of cystitis [11,12,13]. Furthermore, the risks may be heightened if the female is in heat during surgery, as well as due to her age, health status, and sensitivity to drugs or anesthetics [14,15,16].
Therefore, surgical contraception is not the better method for controlling stray populations of felines and canines. Non-surgical options include immunocontraception targeting gonadotropin-releasing hormone (GnRH) or the zona pellucida (ZP), hormone analog implants like deslorelin acetate, suppression of genes associated with GnRH release, and intrauterine devices. These methods avoid the adverse effects of surgery, do not require sedation, and eliminate the need for post-operative care [8,15,17].
Currently, there are no effective non-surgical contraception methods without side effects that provide long-lasting results. Consequently, this field is continuously evolving. This review offers a thorough and updated analysis of non-surgical contraceptive methods evaluated in controlled in vivo studies on female felines and canines. It highlights the impact of reproductive variables, the duration of sterility achieved in females, and the evaluated protocols and doses as the side effects while addressing current challenges and future research opportunities.
2. Materials and Methods
As a tool for searching for contraceptive methods used in female canines and felines, we used the PRISMA flow diagram (Figure 1). The search was performed in PubMed and Google Scholar from February 2024 until April 2024. To obtain broad information about contraceptive methods, we consider all the dates on which the articles were published. The topic keywords were established using the PECO principle from the PRISMA statement [18]. The study population [P] consisted of female cats or dogs subjected to in vivo exposure [E] to contraceptive drugs. The comparator [C] group consisted of animals that received a vehicle-only treatment (the control group). The primary outcome [O] was the suppression of fertility, which was evaluated at specific doses and time points (days post-administration) to assess the efficacy of contraceptive drugs.
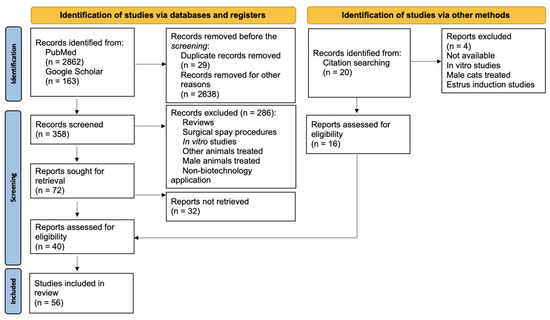
Figure 1.
PRISMA flow diagram.
The terms searched were generally the following: contraception AND non-surgical AND female cats AND/OR bitches; biotechnology; vaccines, hormone, gonadotropin-releasing hormone (GnRH), corpus luteum, kisspeptin, estrus cycle, exfoliative cytology, female. Furthermore, filters of species and sex were used in PubMed, and the combinations were used to obtain more specific results. Additionally, references from other reviews and original articles were checked manually. The articles were selected based on the following criteria:
- English articles about non-surgical spaying based on biotechnology;
- Studies in vivo involving female canines or felines.
Reviews and non-English articles, which include surgical spay and in vitro or situ trials, utilized models other than female felines or canines, which were excluded.
Two reviewers conducted the screening process. One reviewer read the titles and abstracts to pre-select articles, while another evaluated the retrieved articles individually to exclude duplicates or those not meeting the inclusion criteria. Finally, the screening was completed by reading the full article texts.
3. Results
Our research initially identified 2862 studies from PubMed and 163 from Google Scholar, with approximately 56 articles selected for inclusion based on their relevance to the research topic (Figure 1). As the initial search progressed, we discovered methods tested in bitches to induce the contraceptive state through non-surgical procedures, vaccines, hormone analogs, modulators of the corpus luteum (CL), kisspeptin (KP), ultrasonic exposure, and zinc gluconate. In cats, vaccines and hormone analogs have also been tested. In this section, we describe our search results (Table 1) and explain and discuss the findings of the contraceptive methods in the subsections.

Table 1.
Articles that accomplish the inclusion criteria in the search.
3.1. Immunocontraception
Vaccines aimed at reproductive processes represent a method of nonlethal control over abundant free-roaming species [18,23]. Immunocontraception stimulates the production of antibodies that neutralize and block hormone receptors, disrupting the estrus cycle by inhibiting critical proteins or hormones required for reproduction [20,24], such as GnRH (which prevents ovulation by blocking the release of gonadotropins) [23] and zona pellucida (ZP) (which inhibits fertilization and folliculogenesis) [21]. A booster is necessary to prolong the infertile period, ensuring a sustained effect for approximately one year [36,37]. These methods are safe and generate high titers of antibodies for at least one year in female cats and dogs (Figure 2).
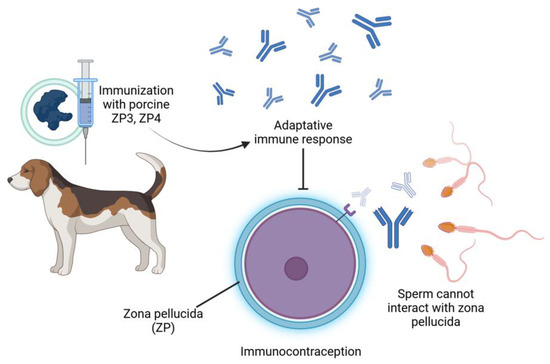
Figure 2.
Mechanism of immunocontraception in female dogs using porcine ZP3 and ZP4. Immunization with porcine zona pellucida proteins (ZP3 and ZP4) triggers an adaptive immune response in the female dog.
The most common antigen in vaccines is GnRH, conjugated to KLH (keyhole limpet hemocyanin) as a carrier in the commercial vaccine GonaCon® (National Wildlife Research Center NWRC, Fort Collins, CO, USA) or as a recombinant protein (e.g., in eight tandem repeats or coupled to a fraction of ZP) [25]. This is safe and to produce a high antibody titer for at least one year in female cats and dogs [23,25,34]. In bitches, the use of sperm as an antigen has led to the development of specific anti-sperm antibodies. However, three immunizations are necessary to achieve a higher titer of antibodies [35]. Another frequently used antigen is porcine zona pellucida (pZP), which induces a high titer of antibody production in bitches and cats. Eade et al. (2009) [19] evaluated the contraception in cats resulting from immunization with ZP polypeptide (55 kDa) and feline ZP A, B, and C subunits expressed by plasmid vectors. The authors concluded that the administration of feline ZPA and ZPB+C subunits are potential candidate antigens for immunocontraceptive vaccines in domestic cats [19]. Immunization of female dogs with bovine luteinizing hormone receptor (LH-R) immunomodulates ovarian function, resulting in a reversible state of infertility [2]. Given the results described, using vaccines provides a reliable method to promote reversible contraception; however, no vaccines guarantee infertility for an extended period (Table 2).
Table 2.
Summary of contraception vaccines tested in female cats and dogs.
As with cats and dogs, overcoming tolerance to self-antigens and inducing durable immunity without repeated booster vaccines is essential for improving the development of immunocontraceptive vaccines [23]. It is also crucial to consider the risk of autoimmune diseases arising from immunization [25] and to manage the duration of contraception, which can vary. In some studies, the vaccines were ineffective during existing pregnancies in cats [20,21,24], while others reported effective durations ranging from days and months to five years [21,23] (Table 2).
Despite this, vaccines that can maintain their long-term antigenic effect are still being developed, as well as devices that allow good antigenic absorption for a long time and with a single administered dose of the antigen without harming the animals [73,74]. Studies are also being carried out in other animals such as mice, where, although the total fertility of the animals has not been eliminated, the number of up to five specimens per litter has been reduced in mice from the experimental groups to which the antigen was inoculated, either in the quadriceps femoris muscle or intradermally [75,76] (Table 2).
The SPRASA protein has also suggested an advance in this method since it only develops in sperm and oocytes of fertile men and women, having an essential function in fertility, and creating a specific antibody would decrease the fertility of individuals [77].
In some other studies, contraceptive vaccines for animals have been explored, which can be divided into regulators of gamete production and regulators of gamete function. Within the first group are GnRH, FSH, and LH [78]. Cats were inoculated with vaccines containing phages made of chemical conjugates of multiple copies of GnRH causing reduced release of gonadotropin hormones and gonadal atrophy, observing a decrease in epididymal vacuolization and an increase in abnormal sperm [79]. However, the authors recommend its application at an early age (prepubertal), and it may apply to other species (Figure 3) (Table 2).
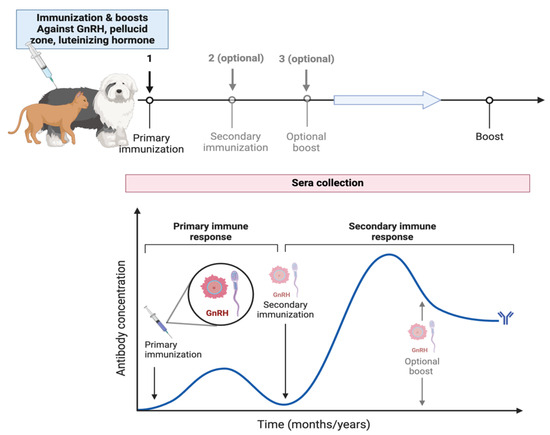
Figure 3.
Canine and feline contraception through immunization and booster vaccines targeting GnRH, the pellucid zone, and luteinizing hormone. It illustrates the timeline and process of primary vaccination followed by optional secondary immunizations and optional boosts, leading to a final boost. The graph below represents the antibody concentration over time, highlighting the primary immune response initiated by the first immunization and a secondary immune response after subsequent immunizations. The antibody concentration increases with each vaccine, peaking during the secondary immune response and declining until the next boost.
The practical application of immunizations as a strategy for birth control in large populations remains a subject of debate, mainly due to inherent limitations regarding their efficacy and safety. Firstly, although contraceptive vaccines represent a non-surgical alternative, their implementation should not be considered a primary option. This is mainly because uncertainties persist concerning the potential health risks associated with these immunizations. Such risks cannot be regarded as secondary to the goal of inducing infertility—especially when compared to surgical methods, which offer permanent cessation of reproductive capacity, along with possible additional benefits such as the prevention of diseases linked to hormone-driven sexual stimulation. Secondly, the outcomes associated with the use of contraceptive vaccines show considerable variability. To date, absolute infertility has not been guaranteed, even under protocols involving periodic revaccination. This lack of predictability significantly limits their utility in large-scale population control programs. For these reasons, the implementation of contraceptive immunizations must be accompanied by comprehensive longitudinal studies, including extended follow-up periods of individuals receiving these immunogens, to assess both long-term efficacy and safety.
3.2. Hormone Analogs
Steroid hormones like progestins, estrogens, and androgens have been utilized as reproductive inhibitors in cats and bitches, yielding variable results, and they represent the most evaluated non-surgical method (Table 3).
Table 3.
Summary of contraceptive hormone analogs tested in female cats and dogs.
Numerous studies have explored drugs that function as agonists, analogs, or antagonists to GnRH receptors to induce contraception in female animals (Figure 4). These drugs disrupt the signaling pathways responsible for hormone synthesis and release, delaying estrus or puberty and extending anestrus periods. Deslorelin acetate is a decapeptide that differs from GnRH by only two amino acids. It acts as an analog of GnRH, exhibiting greater potency at GnRH receptors. In cats, deslorelin acetate has been tested [26,30]. In all articles, reversible alterations in ovulation were observed. In cats, melatonin plays a critical role in regulating the estrus cycle. GnRH antagonists, such as acyline, competitively block the GnRH receptors in the pituitary, inhibiting the activity of the gonadal axis; they also exert an immediate effect compared to agonists [59]. The application of GnRH analog or agonist implants serves not only as a contraceptive method in adult animals but also can delay the onset of puberty in cats and dogs. In a study by Risso et al. (2012) [80], 30 prepubertal crossbreed female domestic cats were divided into two groups of 15. The animals were kept under a photoperiod of 14 h of light and 10 h of darkness. One group received a subcutaneous implant of 4.7 mg deslorelin acetate, while the control group remained untreated. The study aimed to compare the ages at puberty onset between the two groups. The results indicated that the deslorelin-treated group reached puberty between 180 and 428 days, whereas the control group reached puberty earlier, between 134 and 286 days. Deslorelin-treated ovaries appeared small, while control gonads were normal [80] (Table 3).
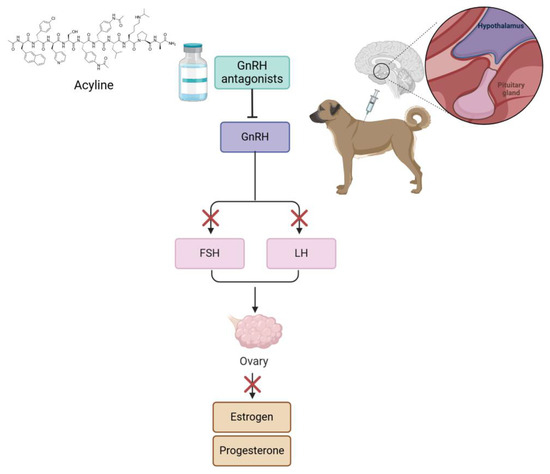
Figure 4.
The mechanism of action of GnRH antagonists in a dog model. The process begins with administering acyline, a GnRH antagonist, which blocks the GnRH receptor in the pituitary gland. This inhibits the secretion of FSH and LH, which are normally produced in response to GnRH stimulation. As a result, ovarian production of estrogen and progesterone is suppressed, thereby impacting the reproductive cycle.
As GnRH, synthetic steroids can also delay puberty. Medroxyprogesterone acetate (MPA) has been tested for this purpose in female cats. In a study by Lopez et al. (2016) [29], postnatal female kittens (n = 10) were reared freely in indoor catteries with 14 h of light per day and were weaned at 40 days. The kittens were randomly assigned within the first 24 h of birth to receive either 10 mg of MPA per animal (n = 6) or a placebo of 0.2 mL of corn oil (n = 4), administered subcutaneously. During puberty, a mating trial was conducted. Twenty-four days after the end of estrus or ovulation, the kittens were ovariohysterectomized. Ovulation occurred in four of the six MPA-treated kittens and three of the four placebo-treated kittens after estrus, leading to pregnancy. All pubertal kittens displayed normal sexual behavior and accepted repeated matings when exposed to males during estrus [29]. In mature bitches, administration of 4–12.5 mg/kg or 2–12.5 mg/kg of chlormadinone acetate (synthetic progesterone) was conducted for at least 1 year [56]. Additionally, melatonin administration has been used to suppress estrous cyclicity in queens temporarily. Melatonin inhibited ovulation and extended the duration of the interestrus interval (2 to 4 months) [28] (Table 3).
Around nineteen articles focused on the use of these drugs in female dogs. GnRH agonists such as nafarelin or deslorelin mimic their action when used in sustained applications, as they stimulate the production and release of gonadotropins s [26,41,81]. The application of these drugs in bitches during the anestrus phase induces a fertile estrous cycle (flare-up effect). Subsequently, it inhibits the gonadal axis, followed by ovarian quiescence due to the downregulation of GnRH receptors [82]. One of the most studied drugs in bitches is the Suprelorin® implant containing deslorelin acetate, which, with periodic administration, reversibly delays estrus in anestrus females and puberty in prepubertal females for an extended period, depending on the implant dose; for instance, the effect period of the 4.7 mg dose is shorter than that of the 9.4 mg implant. When the implant is removed, the estrous cycle returns to normal [45].
Combinations of Suprelorin® with other drugs have also been tested and have shown favorable responses. For instance, Suprelorin® combined with megestrol acetate (MA) results in a more extended interestrus period [44]; however, the combination with acyline (ACY) does not yield a significant difference compared to Suprelorin® treatment alone [59] (Table 3). Lacoste et al. conducted a study involving nine female beagle dogs that received daily subcutaneous injections of 100 μg [D-Trp6, des-Gly-NH210] GnRH ethylamide dissolved in 0.9% NaCl–1% gelatin for 23 months. This treatment inhibited sexual maturation, but normal pituitary–gonadal functions resumed after cessation, returning to normal within two months post-treatment. Secondary follicles developed following a 14-month recovery period [49] (Table 3).
The continuous administration of synthetic steroids, such as chlormadinone acetate (CMA), in bitches can prevent estrus and delay it for at least one year [56]. As compiled in the present paper, hormone analogs have demonstrated effective results in most cases. Despite their contraceptive efficacy, these hormones can induce side effects that may be life-threatening, including pyometra, mammary fibroadenomatosis, neoplasms, and insulin resistance [28]. In a study conducted by [83] Tamada et al. (2003), estrus was prevented with weekly oral administration of 2 mg chlormadinone acetate for 2.0 to 9.8 years in bitches and queens. Although effective, abnormalities such as mammary or uterine disorders, or both, were observed in seven out of 14 bitches and nine out of 24 queens during this long-term treatment [83] (Table 3).
Progestins are widely investigated due to their progesterone-like structure, which suppresses gonadotropin secretion. When applied, they can cause side effects such as an increased risk of diabetes, tumors, and breast hyperplasia, which is why their use is not yet permitted [77] (Table 3).
In vitro cell cultures have designed peptides with a high capacity and specificity to bind to LH, which administers auristatin to Leydig cells, and another peptide targeting FSH that administers menadione to Sertoli cells. These peptides induce apoptosis of germ cells and can achieve non-surgical sterilization in individuals with a single administration. However, the variation in responses from these experiments remains significant, indicating that further investigation is warranted [84] (Table 3).
Another possibility that has been considered is GnRH hormone agonists produced through amino acid substitutions of the hormone, which grant greater potency and a longer half-life. These agonists negatively regulate the hypophysial GnRH receptors by inhibiting their production, thereby postponing puberty in individuals. For this purpose, postnatal deslorelin in dogs delays puberty in both females and males [85] (Table 3).
Reproductive endocrinology is a well-established and widely utilized field within animal health, holding significant potential to completely inhibit reproductive capacity in this species. However, such interventions must be approached with caution due to the inherent risks associated with manipulating the neuroendocrine axis. The administration of substances that interfere with the synthesis of sex hormones impacts the production of male and female gametes, as well as a range of physiological processes essential for the organism’s overall metabolic function.
Among the most notable consequences is an increased predisposition to insulin resistance and a reduced capacity for synthesizing key neurotransmitters, ultimately disrupting the neurochemical and functional balance of the nervous system. Although hormonal strategies for reproductive control in animals are promising, their implementation must be carefully considered in light of their potential impact on overall health. The sustained alteration of sex hormone levels may lead to systemic pathologies that compromise the organism’s development and well-being, underscoring the complex interconnection between reproductive function and broader physiological systems.
3.3. Use of Modulators of Corpus Luteum (CL) as Contraceptive Methods in Bitches
Disrupting the estrus cycle in bitches through CL regulation has been investigated for non-surgical contraception. Trilostane is a drug that disrupts CL function by competitively inhibiting the 3β-hydroxysteroid dehydrogenase/isomerase system (3β-HSD), which is expressed in luteal cells to synthesize progesterone. When administered during the luteal phase, trilostane resulted in a decrease in progesterone levels without affecting ACTH or prolactin levels [62] (Table 4).
Table 4.
Summary of compounds that regulate the CL function as contraceptive tested in bitches.
Bromocriptine and cabergoline administration affect the maintenance of the CL. Bromocriptine causes an abrupt decrease in progesterone levels [61], and cabergoline inhibits the production of prolactin, a hormone that helps maintain the function of progesterone released by the CL [65]. The CL plays a vital role in the estrus cycle by inducing progesterone production, and previous studies have shown that prostaglandins serve as essential regulators of the CL [63,64,66]. Daily administration of firocoxib, a selective cyclooxygenase-2 inhibitor, to females after ovulation (around the diestrus stage) has been demonstrated to cause, on average, a 10-day suppression in the expression of messenger RNA (mRNA) of prostaglandin E2 synthase, leading to decreased levels of prostaglandins E2 and 2α. The steroidogenic acute regulatory (STAR) protein and prolactin receptor (PR) are other proteins that are not expressed and regulate the CL [64]. Research indicates that if non-surgical sterilization is desired, further investigation can be conducted on the SLC2A1/GLUT1 receptors and the insulin receptor (INR), as these contribute energy to the CL and are related to vasculogenesis.
In bitches, luteotropic modulators offer a potentially safer therapeutic alternative for interrupting the estrous cycle. This method may help avoid adverse effects associated with the direct manipulation of primary neuroendocrine axes, as its action is limited to maintaining the corpus luteum. Although any form of reproductive cycle interruption or manipulation without a clinical indication may pose risks to the organism’s homeostasis, in this context, a strategy that halts the progression of the estrous cycle without causing significant systemic alterations or triggering a pathological condition is preferred.
3.4. Kisspeptin (KP)
In 2003, it was discovered that KP signaling serves as a gateway to reproductive function in both males and females. This discovery marked a milestone in unraveling the endocrinological regulation of reproduction [86,87] (Figure 5). The KISS1 gene encodes KP and its receptor (GPR54 or KiSS1R). When they bind, they trigger the release of GnRH, which travels through the hypothalamic–pituitary system to the anterior pituitary and stimulates the secretion of LH and FSH. This makes it vital in regulating the hypothalamic–pituitary–gonadal (HPG) axis. They also promote the release of LH and other hormones related to the estrus cycle and ovulation, such as E2 and FHS [88]. Therefore, various studies have assessed their role as crucial molecules in controlling the feline and canine estrous cycle [69] (Table 5).
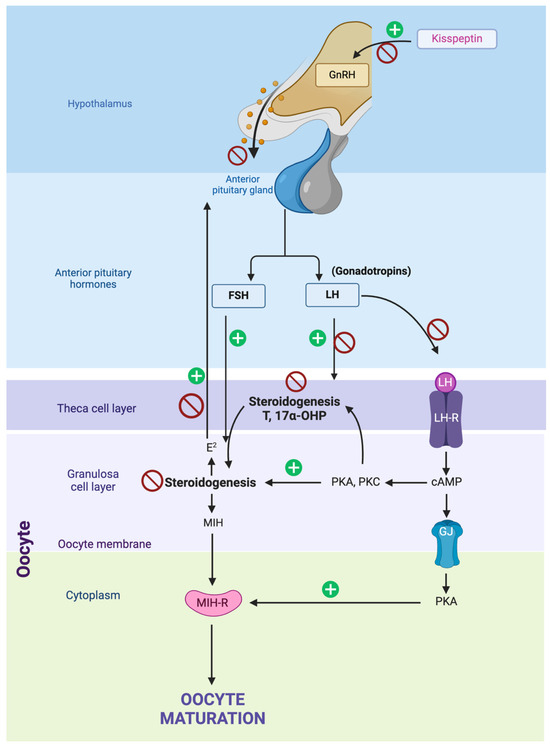
Figure 5.
The hypothalamic–pituitary–gonadal axis and the inhibition points tested for canine reproduction inhibition are represented by red stop symbols. Inhibition of kisspeptin, which positively regulates GnRH (gonadotropin-releasing hormone) secretion in the hypothalamus, blocks the initial signal necessary for gonadotropic hormone production. Direct inhibition of GnRH in the hypothalamus prevents the release of FSH (follicle-stimulating hormone) and LH (luteinizing hormone) from the anterior pituitary gland. Inhibiting LH at the pituitary gland level prevents the stimulation of theca and granulosa cells in the ovaries, reducing the production of essential steroids like testosterone (T) and 17α-OHP (17α-hydroxyprogesterone) required for oocyte maturation. Inhibiting FSH in the pituitary gland disrupts the stimulation of granulosa cell activity, affecting estradiol (E2) production and other factors needed for oocyte maturation. Finally, inhibition of steroidogenesis in the theca cell layer reduces the production of testosterone and 17α-OHP, crucial precursors for estradiol synthesis in granulosa cells, thus impacting oocyte maturation.
Table 5.
Summary of studies administered kisspeptin as a contraceptive method in bitches.
The KISS1 gene encodes a 145-amino-acid peptide that can be cleaved into four peptides, each containing a common C-terminal RF-amide decapeptide (KP-54, KP-14, KP-13, and KP-10) [89,90]. According to the data compiled in this review, KP-10 is the most tested peptide in female cats and dogs as a contraceptive, yielding variable results (Table 5).
The administration of KP-10 to anestrous bitches rapidly induced significant serum concentrations (Tmax ≤ 5 min, CPmax ≤ 30 min) of hormones associated with the estrous cycle (LH, FSH, E2), with effects lasting up to fourteen days [69]. Terse et al. (2021) observed in dogs that administration of 1000 μg/kg of KP-10 intravenously for 14 days did not induce changes in clinical signs, body weight, hematological values, urine, body temperature, food or water consumption, or sinus rhythms, considering it a safe compound. Therefore, bitches remained in the anestrus phase throughout the entire dosing period [70] (Table 5).
The specificity between ligand and receptor has allowed for investigating antagonist substances with varying responses across the species under study. The continuous intravenous infusion of KP antagonists (p271, p354, and p356) did not result in significant alterations in basal plasma LH concentrations in bitches, into which canine KP-10 was administered two hours after the infusion began [68] (Table 5).
The KP-10 molecule has shown the most significant inhibitory effect during the estrous cycle of female cats and dogs compared to hormone treatments. In other species, its administration has also demonstrated alterations in reproductive processes. The KP-10 analog peptide 234 (p234) exhibited a strong inhibitory effect. Intracerebroventricular administration in rats resulted in delayed vaginal opening (an indicator of puberty) and did not permit serum LH levels to rise when co-administered with KP-10. In intact rats and mice, it required combination with penetratin (p271) to prevent the increase in circulating LH in castrated rodents and sheep [70,91,92] (Table 5).
The administration of agonists and antagonists presents intriguing therapeutic possibilities for addressing endocrinological needs in mammals, such as controlling canine and feline overpopulation, by developing minimally invasive pharmaceutical forms to suppress reproductive capacity temporarily or permanently. However, it remains essential to standardize the dosage and the timing of administration during the estral cycle (Table 5).
It is essential to mention that using vaccines with the KISS1 gene in sheep, conjugated with the HBsAG-S gene, has already reduced sexual behavior and spermatogenesis. Likewise, utilizing the kisspeptin-54 gene could produce humoral antibodies that neutralize endogenous kisspeptin, thereby reducing testicular growth, scrotal circumference, and spermatogenesis, making it a potential option for non-surgical sterilizations [93] (Table 5).
Alteration of the hypothalamic–pituitary–gonadal axis affects not only reproductive physiology but also a wide range of systemic functions regulated by sex steroid hormones, whose disruption can compromise overall animal health. In this context, modulation of the kisspeptin/GPR54 system represents a promising pathway for reproductive suppression; however, its influence extends beyond reproduction and may lead to broader metabolic and endocrine disturbances. Despite its potential as a contraceptive tool, there remain significant gaps in knowledge regarding the appropriate dosage, route of administration, and safety profile of kisspeptin-based interventions in domestic animals. Therefore, until standardized protocols are established to ensure both efficacy and safety, it is advisable to prioritize contraceptive strategies that do not involve direct suppression of key neuroendocrine axes.
3.5. Other Contraception Methods
The effect of therapeutic ultrasound exposure on the ovaries was evaluated, revealing changes indicative of potential fertility loss. Ultrasound treatment (1 MHz frequency, 1.5 W/cm2) applied to dog ovaries for 5 or 10 min heightened the circulating inflammatory response and oxidative stress, leading to a reduction in the number of preserved follicles and oocytes, as well as tissue alterations and changes in ovarian dimensions. However, the results remain inconclusive, necessitating further exploration of the effects of direct exposure of the ovaries to therapeutic ultrasound utilizing different frequencies and power levels [72] (Table 6).
Table 6.
Summary of other contraceptive methods tested in female cats and dogs.
There are commercial spermicides and intrauterine devices for dogs; however, they are not recommended. Reports indicate that these barrier systems are impractical due to the challenges associated with the transcervical cannulation necessary for their application. Other contraceptive methods being tested include those that act with Müllerian inhibitory substances (MISs), also known as anti-Müllerian hormones (AMHs). In one study, AAV9-MIS treatments were placed in female mice that later mated with breeding males and obtained litters very similar to those of the group that did not receive MISs. However, after 6 weeks, the females treated with high AAV9-MISs became infertile, which suggests that this contraceptive method can be beneficial in dogs and cats [94].
Another method used is the administration of zinc gluconate in the ovaries, an invasive treatment that yields favorable results. A study conducted by [71] Mogheiseh et al. (2017) involved healthy, fertile female dogs aged 1–2 years in anestrus or diestrus (n = 5) and evaluated the effects of zinc gluconate injection on the ovaries. A sterile, neutralizing zinc gluconate solution (13.1 mg/mL) was injected into the ovaries, with the volume determined by ultrasound measurements of ovarian diameters. Under general anesthesia, a laparotomy was performed for the injection. One month post-injection, the diameter of the ovaries was significantly reduced, and leukocytes in vaginal cytology decreased from the second to the third day. Histopathological examination revealed hyperemia, fibrosis, and hemosiderin pigment in the ovaries, indicating that not all ovarian structures were destroyed after one month [95]. The application of zinc gluconate administration in females is limited. It requires surgical procedures and the development of protocols to evaluate the side effects of intraovarian administration. It still does not seem to be affordable to the population at this moment because it requires direct contact or administration in ovaries through a laparotomy, which involves the same risks as an ovariohysterectomy. Therefore, it is necessary to try more accessible and non-invasive ways to carry out these treatments.
In recent years, targeted gene silencing has been utilized to induce permanent sterility to control feral populations [96]. Current research on contraceptive vaccines focuses on using adeno-associated virus (AAV). This non-enveloped virus can be engineered to deliver DNA to target cells [97]. Vansandt et al. (2023) [1] utilized AAV9 to provide a female cat with an anti-Müllerian hormone (AMH) transgene (designated fcMISv2). Researchers have demonstrated that the vectored contraceptive prevents breeding-induced ovulation, leading to complete infertility [1] (Table 6).
The implementation of innovative tools in contraceptive methods, encompassing both hormonal and non-hormonal options, has the potential to create new strategies that expand the current perspective on contraception. In this context, the integration of technologies like ultrasound-based contraception signifies a promising area of research, offering a non-invasive alternative that avoids the risk of causing systemic hormonal dysregulation. Furthermore, the development of new hormonal options that do not directly interfere with the endocrine system may encourage the generation of applied knowledge with significant clinical implications. However, these emerging strategies still require a solid experimental foundation and more substantial scientific evidence to uphold their efficacy and safety, allowing them to compete with established methods in the field of contraception.
4. Final Considerations
The challenge of suppressing fertility in cats and dogs has existed for over 50 years. In recent years, the interference of the estral cycle through the block of its regulating molecules has been tested (Figure 5), since the regulation of the production of KP [69], until AAV9 was used to prevent breeding-induced ovulation [1]. The primary factor that disrupts the regulation of the estral cycle in female cats and dogs is the involvement of numerous molecules in its control, making it challenging to manage the positive or negative feedback mechanisms that inhibit a particular molecule or signaling pathway.
Technological advances have allowed us to develop more contraceptive-specific methods with fewer side effects. According to the World Small Animal Veterinary Association, an ideal contraceptive product would have the following characteristics: (i) rapidly induce permanent sterilization, (ii) eliminate breeding behavior and fertility, (iii) require only a single dose, (iv) be effective for male and female dogs and cats of all ages, and (v) be safe and easy to administer. The global need for a contraceptive method for feral populations has led to the establishment of prizes and grants, such as The Michelson Prize and Grants. This program aims to prevent the euthanasia of healthy, adoptable companion animals in shelters and reduce the populations of feral and free-roaming cats and dogs. It offers a USD 25 million prize for developing a non-surgical sterilant that can effectively sterilize dogs and cats in a single treatment.
Currently, no product meets all of these criteria, and government institutions authorize none for contraceptive methods or the sterilization of cats or dogs. The tested non-surgical contraceptive methods in female cats and dogs have shown variable results, including immunocontraception, hormonal treatments, kisspeptin analogs, non-steroidal anti-inflammatory drugs, and cytotoxins. However, some promising techniques exist, such as gene therapy, small interfering RNA to inhibit reproductive targets, and the delivery of cytotoxins to pituitary gonadotrophs or GnRH-producing neurons in the hypothalamus. Continued research and innovation in non-surgical population control procedures are necessary, as they are safer for females, less costly, and quicker, allowing for administration to more females in a shorter time frame.
Surgical sterilizations are usually performed at five to seven months of age in bitches, as they have reached sexual or hormonal maturity; it is not suggested that they be performed earlier because bitches may present infantilism; however, it is not suggested that they be performed after the first estrus, as the possibility of mammary gland tumor incidences increases up to seven times [98].
Among the different surgical sterilization techniques, ovariohysterectomy (OVH) midline is the technique most used in the daily practice of veterinary medicine; in this technique, an incision is made in the skin of the ventral midline 4–6 cm below the navel and extending caudally. All these procedures are expected to use a 2–0 polyglycolic acid suture, which will depend on the tissue’s friability, the patient’s size, and the stage of the estrous cycle of the female. With this technique, the hormonal action of the ovaries and uterus is eliminated totally and permanently [98]. In the ovariectomy, only the ovaries are surgically removed, keeping the body of the uterus. It can be performed through two types of approaches. The first and most used is through the midline or lateral ovariectomy, which is a variant of the previous technique [15,99,100,101]. The third most common technique is tubal ligation. One of the advantages of this technique is that it is not very invasive and quick. Still, it does not prevent secondary hormonal reproductive problems, and the females continue their normal sexual cycle [15].
It is essential to mention that wound-healing complications are among the side effects of these types of surgeries. This issue is related to the duration of the surgery, with a greater amount of post-surgical inflammation and infections occurring in procedures lasting more than 90 min. Another common side effect is hemorrhage, which is the most frequent complication in ovariohysterectomies for females over 25 kg, potentially leading to hypovolemic shock and the animal’s death. Stump pyometra is another issue that can arise from these surgical procedures if the ovary is not entirely removed, which may cause the bitch to experience hormonal peaks of endogenous progesterone. Ureteral ligation is also a possible complication from accidentally ligating a ureter during the ligation of the uterine corpus or ovarian arteriovenous complex. Additionally, vaginal bleeding cannot be disregarded as a surgical side effect, and if it becomes severe, celiotomy may be indicated [99,100].
A primary factor in the varied results of non-surgical contraceptives included in this review is the diversity of breeds and sizes of cats and dogs, along with the influence of keeping bitches together, as they may have synchronized estrous cycles [95,102]. Long-haired breeds of cats seem more sensitive to daylight than short-haired breeds [102]. There are distinctions between long- and short-haired cat breeds; for example, Persian cats do not have regular estrous cycles, even during extended daylight. In contrast, short-haired Siamese and related breeds have estrous cycles year-round, regardless of daylight length [102].
Regarding dogs, factors intrinsic to species, such as seasonality, have influenced the results of studies focused on reproduction [103]. The study populations include free-roaming, mixed-breed, purebred, and laboratory dogs. Thus, the diversity of animals examined in studies assessing contraceptive methods may be a crucial element contributing to bias in the results.
5. Conclusions
There is an urgent need for effective non-surgical contraceptive methods to manage the increasing populations of stray cats and dogs, which raise significant concerns for animal welfare and public health. While surgical sterilization is the most common form of contraception, researchers are actively exploring non-invasive alternatives, including immunocontraception, hormonal treatments, and targeted gene silencing. These non-surgical methods circumvent surgery-related complications, such as the need for anesthesia and postoperative care. However, none of the current techniques offer permanent sterility or a universally accepted solution, and most have demonstrated limited efficacy with varying outcomes. It is crucial to develop treatments with long-lasting effects, minimal side effects, and the potential for widespread application. Therefore, further research is necessary to enhance these techniques and make them suitable for controlling the overpopulation of stray animals.
Author Contributions
S.I.P.-C., M.A.V.-G., J.C.-G., A.E.I.-R., A.S.-R., J.J.P.-R., J.I.C.-C., S.R.-M., P.A.V.-D., D.Q.-G., G.L.-G. and D.V.-E. significantly contributed to the reported work, encompassing conception, study design, execution, data acquisition, analysis, interpretation, as well as revising or critically reviewing the article. They also provided final approval for the published version, agreed on the journal of submission, and confirmed accountability for every aspect of the work. All authors have read and agreed to the published version of the manuscript.
Funding
PAPIIT IN204722 and PAPIME PE205524 provided by DGAPA-UNAM awarded to Gerardo Leyva-Gómez.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding authors.
Acknowledgments
Sheila Peña acknowledges the Postdoctoral Program Scholarship from the Secretaría de Ciencia, Humanidades, Tecnología e Innovación, titled “Estancias Posdoctorales por México 2022; Estancia Posdoctoral Académica Inicial 2022”, awarded to Sheila Irais Peña-Corona (CVU:495850) and Claudia Fernandez Diez for their assistance in preparing the draft. Juan Isaac Chávez Corona acknowledges the Programa de Doctorado en Ciencias de la Producción y de la Salud Animal at the Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AAV | adeno-associated virus |
| ALHS | anti-LH serum |
| Al(OH)3 | aluminum hydroxide |
| b.w. | body weight |
| CL | corpus luteum |
| cZP | crude porcine zona pellucida |
| CP | complete Freund’s adjuvant |
| CP-20 | CP-20 adjuvant |
| DA | deslorelin acetate |
| DMSO | dimethyl sulfoxide |
| E2 | estradiol |
| eCG | equine chorionic gonadotropin |
| FSH | follicle-stimulating hormone |
| GnRH | gonadotropin-releasing hormone |
| hCG | human chorionic gonadotropin |
| IgG | immunoglobulin G |
| i.m. | intramuscular |
| i.v. | intravenous |
| KLH | keyhole limpet hemocyanin |
| KP-10 | kisspeptin-10 |
| LH | luteinizing hormone |
| LH-R | luteinizing hormone receptor |
| LH-RH | luteinizing hormone-releasing hormone |
| MA | megestrol acetate |
| MAL | luteal Hhrmone |
| mL | milliliters |
| NaCl | sodium chloride |
| P4 | progesterone |
| PGE2 | prostaglandin E2 |
| PBS | phosphate-buffered saline |
| pZP | porcine zona pellucida |
| SIZP | soluble isolated zona pellucida |
| s.c. | subcutaneous |
| STAR | steroidogenic acute regulatory protein |
| TT-KK-pZP3 | promiscuous T cell epitope of tetanus toxoid conjugated with pZP3 |
| ZP | zona pellucida |
References
- Vansandt, L.M.; Meinsohn, M.C.; Godin, P.; Nagykery, N.; Sicher, N.; Kano, M.; Kashiwagi, A.; Chauvin, M.; Saatcioglu, H.D.; Barnes, J.L.; et al. Durable contraception in the female domestic cat using viral-vectored delivery of a feline anti-Müllerian hormone transgene. Nat. Commun. 2023, 14, 3140. [Google Scholar] [CrossRef] [PubMed]
- Saxena, B.B.; Clavio, A.; Singh, M.; Rathnam, P.; Bukharovich, E.Y.; Reimers, T.J.; Saxena, A.; Perkins, S. Effect of immunization with bovine luteinizing hormone receptor on ovarian function in cats. Am. J. Vet. Res. 2003, 64, 292–298. [Google Scholar] [CrossRef]
- Mendoza Roldan, J.A.; Otranto, D. Zoonotic parasites associated with predation by dogs and cats. Parasites Vectors 2023, 16, 55. [Google Scholar] [CrossRef]
- Sánchez, L.M.E.; Ortiz, R.C.R.; Berumen, F.L.R.; Piña, F.J.G.; Delgado, R.M.R. Importancia del manejo de la población canina en situación de calle en México: Perspectivas y desafíos. CIBA Rev. Iberoam. Cienc. Biol. Agropecu. 2023, 12, 24. [Google Scholar]
- Bryce, C.M. Dogs as Pets and Pests: Global Patterns of Canine Abundance, Activity, and Health. Integr. Comp. Biol. 2021, 61, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Orduña-Villaseñor, M.; Valenzuela-Galván, D.; Schondube, J.E. Tus mejores amigos pueden ser tus peores enemigos: Impacto de los gatos y perros domésticos en países megadiversos. Rev. Mex. Biodivers. 2023, 94, e944850. [Google Scholar] [CrossRef]
- Peña-Corona, S.I.; Gomez-Vazquez, J.P.; López-Flores, E.A.; Vargas Estrada, D.; Arvizu-Tovar, L.O.; Pérez-Rivero, J.J.; Juárez Rodríguez, I.; Sierra Resendiz, A.; Soberanis-Ramos, O. Use of an Extrapolation Method To Estimate the Population of Cats and Dogs Living at Homes in Mexico in 2022; Veterinaria México OA: Ciudad de México, Mexico, 2022; p. 9. [Google Scholar]
- Pelaez, M.; Echevarría, L.; Soler-Tovar, D.; Falcón, N. Métodos de contracepción en el control poblacional de perros: Un punto de vista de los médicos veterinarios de clínica de animales de compañía. Salud Tecnol. Vet. 2018, 6, 55. [Google Scholar] [CrossRef]
- Salamanca, C.A.; Polo, L.J.; Vargas, J. Sobrepoblación canina y felina: Tendencias y nuevas perspectivas. Rev. Fac. Med. Vet. Zootec. 2011, 58, 45–53. [Google Scholar]
- Kustritz, R. Effects of surgical sterilization on canine and feline health and on society. Reprod. Domest. Anim. Zuchthyg. 2012, 47, 214–222. [Google Scholar] [CrossRef]
- Oliveira-Martins, M.; Portugal, M.; Cardoso, L.; Martins-Bessa, A. The Impact of Pediatric Neutering in Dogs and Cats-A Retrospective Study. Animals 2023, 13, 2487. [Google Scholar] [CrossRef]
- McKenzie, B. Evaluating the benefits and risks of neutering dogs and cats. CABI Rev. 2010, 5, 1–18. [Google Scholar] [CrossRef]
- Spain, C.V.; Scarlett, J.M.; Houpt, K.A. Long-term risks and benefits of early-age gonadectomy in dogs. J. Am. Vet. Med. Assoc. 2004, 224, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Benavides Melo, C.J.; Astaíza Martínez, J.M.; Rojas, M.L. Complicaciones por esterilización quirúrgica mediante ovariohisterectomía en perras: Revisión sistemática. Rev. Med. Vet. 2018, 1, 83–93. [Google Scholar] [CrossRef]
- Uribe Sarmiento, F.F.; Prada Delgado, Y.F.; Rodríguez Barajas, B.S.; Bayona Sánchez, J.A. Métodos de Esterilización en Caninos Y Felinos; Revisión de Literatura; Universidad Cooperativa de Colombia: Medellín, Colombia, 2018. [Google Scholar]
- Trujillo Santillán, N.S. Estimación de la Población de Perros Y Gatos Vagabundos Dentro Del Cantón Riobamba; Universidad de Cuenca: Cuenca, Ecuador, 2022. [Google Scholar]
- Zumpano, R.; Tortosa, A.; Degregorio, O.J. Estimación del impacto de la esterilización en el índice de crecimiento de la población de caninos. Rev. Investig. Vet. Del. Perú 2011, 22, 336–341. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. bmj 2021, 372, n71. [Google Scholar] [CrossRef]
- Eade, J.A.; Roberston, I.D.; James, C.M. Contraceptive potential of porcine and feline zona pellucida A, B and C subunits in domestic cats. Reproduction 2009, 137, 913–922. [Google Scholar] [CrossRef]
- Fischer, A.; Benka, V.A.; Briggs, J.R.; Driancourt, M.-A.; Maki, J.; Mora, D.S.; Morris, K.N.; Myers, K.A.; Rhodes, L.; Vansandt, L.M. Effectiveness of GonaCon as an immunocontraceptive in colony-housed cats. J. Feline Med. Surg. 2018, 20, 786–792. [Google Scholar] [CrossRef]
- Gorman, S.P.; Levy, J.K.; Hampton, A.L.; Collante, W.R.; Harris, A.L.; Brown, R.G. Evaluation of a porcine zona pellucida vaccine for the immunocontraception of domestic kittens (Felis catus). Theriogenology 2002, 58, 135–149. [Google Scholar] [CrossRef]
- Ivanovo, M.; Petrov, M.; Klissourska, D.; Mollova, M. Contraceptive potential of porcine zona pellucida in cats. Theriogenology 1995, 43, 969–981. [Google Scholar] [CrossRef]
- Levy, J.K.; Friary, J.A.; Miller, L.A.; Tucker, S.J.; Fagerstone, K.A. Long-term fertility control in female cats with GonaCon™, a GnRH immunocontraceptive. Theriogenology 2011, 76, 1517–1525. [Google Scholar] [CrossRef]
- Levy, J.K.; Mansour, M.; Crawford, P.C.; Pohajdak, B.; Brown, R.G. Survey of zona pellucida antigens for immunocontraception of cats. Theriogenology 2005, 63, 1334–1341. [Google Scholar] [CrossRef] [PubMed]
- Robbins, S.; Jelinski, M.; Stotish, R. Assessment of the immunological and biological efficacy of two different doses of a recombinant GnRH vaccine in domestic male and female cats (Felis catus). J. Reprod. Immunol. 2004, 64, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, C.; Volpato, R.; Destro, F.; Trevisol, E.; Sousa, N.R.; Guaitolini, C.; Derussi, A.; Rascado, T.; Lopes, M.D. Ovarian activity reversibility after the use of deslorelin acetate as a short-term contraceptive in domestic queens. Theriogenology 2012, 78, 817–822. [Google Scholar] [CrossRef]
- Furthner, E.; Roos, J.; Niewiadomska, Z.; Maenhoudt, C.; Fontbonne, A. Contraceptive implants used by cat breeders in France: A study of 140 purebred cats. J. Feline Med. Surg. 2020, 22, 984–992. [Google Scholar] [CrossRef]
- Gimenez, F.; Stornelli, M.C.; Tittarelli, C.M.; Savignone, C.A.; Dorna, I.; de la Sota, R.; Stornelli, M.A. Suppression of estrus in cats with melatonin implants. Theriogenology 2009, 72, 493–499. [Google Scholar] [CrossRef]
- Lopez Merlo, M.; Faya, M.; Blanco, P.G.; Carranza, A.; Barbeito, C.; Gobello, C. Failure of a single dose of medroxyprogesterone acetate to induce uterine infertility in postnatally treated domestic cats. Theriogenology 2016, 85, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Goericke-Pesch, S.; Georgiev, P.; Atanasov, A.; Albouy, M.; Navarro, C.; Wehrend, A. Treatment of queens in estrus and after estrus with a GnRH-agonist implant containing 4.7 mg deslorelin; hormonal response, duration of efficacy, and reversibility. Theriogenology 2013, 79, 640–646. [Google Scholar] [CrossRef]
- Graham, L.; Swanson, W.; Wildt, D.; Brown, J. Influence of oral melatonin on natural and gonadotropin-induced ovarian function in the domestic cat. Theriogenology 2004, 61, 1061–1076. [Google Scholar] [CrossRef]
- Rubion, S.; Driancourt, M. Controlled delivery of a GnRH agonist by a silastic implant (Gonazon) results in long-term contraception in queens. Reprod. Domest. Anim. 2009, 44, 79–82. [Google Scholar] [CrossRef]
- Toydemir, T.; Kılıçarslan, M.; Olgaç, V. Effects of the GnRH analogue deslorelin implants on reproduction in female domestic cats. Theriogenology 2012, 77, 662–674. [Google Scholar] [CrossRef]
- Bender, S.C.; Bergman, D.L.; Wenning, K.M.; Miller, L.A.; Slate, D.; Jackson, F.R.; Rupprecht, C.E. No adverse effects of simultaneous vaccination with the immunocontraceptive GonaCon™ and a commercial rabies vaccine on rabies virus neutralizing antibody production in dogs. Vaccine 2009, 27, 7210–7213. [Google Scholar] [CrossRef]
- Esmailnejad, A.; Nikahval, B.; Mogheiseh, A.; Karampour, R.; Karami, S. The detection of canine anti-sperm antibody following parenteral immunization of bitches against homogenized whole sperm. Basic Clin. Androl. 2020, 30, 1. [Google Scholar] [CrossRef]
- Gupta, S.K.; Toor, S.; Minhas, V.; Chaudhary, P.; Raman, M.; Anoop, S.; Panda, A.K. Contraceptive efficacy of recombinant porcine zona proteins and fusion protein encompassing canine ZP3 fragment and GnRH in female beagle dogs. Am. J. Reprod. Immunol. 2022, 87, e13536. [Google Scholar] [CrossRef]
- Liu, Y.; Tian, Y.; Zhao, X.; Jiang, S.; Li, F.; Zhang, Y.; Zhang, X.; Li, Y.; Zhou, J.; Fang, F. Immunization of dogs with recombinant GnRH-1 suppresses the development of reproductive function. Theriogenology 2015, 83, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Mahi-Brown, C.A.; Yanagimachi, R.; Hoffman, J.; Huang, T., Jr. Fertility control in the bitch by active immunization with porcine zonae pellucidae: Use of different adjuvants and patterns of estradiol and progesterone levels in estrous cycles. Biol. Reprod. 1985, 32, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Mahi-Brown, C.A.; Huang, T.T., Jr.; Yanagimachi, R. Infertility in bitches induced by active immunization with porcine zonae pellucidae. J. Exp. Zool. 1982, 222, 89–95. [Google Scholar] [CrossRef]
- Saxena, B.; Clavio, A.; Singh, M.; Rathnam, P.; Bukharovich, Y.; Reimers, T., Jr.; Saxena, A.; Perkins, S. Modulation of ovarian function in female dogs immunized with bovine luteinizing hormone receptor. Reprod. Domest. Anim. 2002, 37, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Pino, F.; Gutiérrez-Cedillo, V.; Canales-Vargas, E.J.; Gress-Ortega, L.R.; Miller, L.A.; Rupprecht, C.E.; Bender, S.C.; García-Reyna, P.; Ocampo-López, J.; Slate, D. Concomitant administration of GonaCon™ and rabies vaccine in female dogs (Canis familiaris) in Mexico. Vaccine 2013, 31, 4442–4447. [Google Scholar] [CrossRef]
- Beijerink, N.; Bhatti, S.; Okkens, A.; Dieleman, S.; Duchateau, L.; Kooistra, H. Pulsatile plasma profiles of FSH and LH before and during medroxyprogesterone acetate treatment in the bitch. Theriogenology 2008, 70, 179–185. [Google Scholar] [CrossRef]
- Brändli, S.; Palm, J.; Kowalewski, M.P.; Reichler, I.M. Long-term effect of repeated deslorelin acetate treatment in bitches for reproduction control. Theriogenology 2021, 173, 73–82. [Google Scholar]
- Corrada, Y.; Hermo, G.; Johnson, C.; Trigg, T.; Gobello, C. Short-term progestin treatments prevent estrous induction by a GnRH agonist implant in anestrous bitches. Theriogenology 2006, 65, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Gontier, A.; Youala, M.; Fontaine, C.; Raibon, E.; Fournel, S.; Briantais, P.; Rigaut, D. Efficacy and safety of 4.7 mg deslorelin acetate implants in suppressing oestrus cycle in prepubertal female dogs. Animals 2022, 12, 3504. [Google Scholar] [CrossRef]
- Jurczak, A.; Janowski, T.; Zdunczyk, S.; Failing, K.; Schuler, G.; Hoffmann, B. Attempts to downregulate ovarian function in the bitch by applying a GnRH agonist implant in combination with a 3ß-hydroxysteroid-dehydrogenase blocker; a pilot study. Theriogenology 2020, 145, 176–180. [Google Scholar] [CrossRef]
- Kaya, D.; Gram, A.; Kowalewski, M.P.; Schäfer-Somi, S.; Kuru, M.; Boos, A.; Aslan, S. Expression of GnRH receptor in the canine corpus luteum, and luteal function following deslorelin acetate-induced puberty delay. Reprod. Domest. Anim. 2017, 52, 1104–1112. [Google Scholar] [CrossRef]
- Kaya, D.; Schäfer-Somi, S.; Kurt, B.; Kuru, M.; Kaya, S.; Kaçar, C.; Aksoy, Ö.; Aslan, S. Clinical use of deslorelin implants for the long-term contraception in prepubertal bitches: Effects on epiphyseal closure, body development, and time to puberty. Theriogenology 2015, 83, 1147–1153. [Google Scholar] [CrossRef]
- LACOSTE, D.; DUBÉ, D.; TRUDEL, C.; BÉLANGER, A.; LABRIE, F. Normal gonadal functions and fertility after 23 months of treatment of prepubertal male and female dogs with the GnRh agonist [D-Trp6, des-Gly-NH210] GnRH ethylamide. J. Androl. 1989, 10, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Marino, G.; Rizzo, S.; Quartuccio, M.; Macri, F.; Pagano, G.; Taormina, A.; Cristarella, S.; Zanghi, A. Deslorelin Implants in Pre-pubertal Female Dogs: Short-and Long-Term Effects on the Genital Tract. Reprod. Domest. Anim. 2014, 49, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Peña-Corona, S.; León, P.; Mendieta, E.; Villanueva, M.; Salame, A.; Vargas, D.; Mora, G.; Serrano, H.; Villa-Godoy, A. Effect of a single application of coumestrol and/or dimethyl sulfoxide, on sex hormone levels and vaginal cytology of anestrus bitches. Vet. México 2019, 6, 1–15. [Google Scholar] [CrossRef]
- Peña-Corona, S.; León-Ortiz, P.; Villanueva-Martínez, M.; Mendoza-Rodríguez, C.; Martínez-Maya, J.; Serrano, H.; Vargas, D.; Villa-Godoy, A. Administration of Coumestrol and/or Dimethyl Sulfoxide Affects Vaginal Epithelium and Sex Hormones in Female Dog. Indian J. Anim. Res. 2022, 6, 1–15. [Google Scholar] [CrossRef]
- Romagnoli, S.; Stelletta, C.; Milani, C.; Gelli, D.; Falomo, M.E.; Mollo, A. Clinical use of deslorelin for the control of reproduction in the bitch. Reprod. Domest. Anim. 2009, 44, 36–39. [Google Scholar] [CrossRef]
- Rubion, S.; Desmoulins, P.; Rivière-Godet, E.; Kinziger, M.; Salavert, F.; Rutten, F.; Flochlay-Sigognault, A.; Driancourt, M. Treatment with a subcutaneous GnRH agonist containing controlled release device reversibly prevents puberty in bitches. Theriogenology 2006, 66, 1651–1654. [Google Scholar] [CrossRef]
- Sahara, K.; Tsutsui, S.; Naito, Y.; Fujikura, K. Prevention of estrus in bitches by subcutaneous implantation of chlormadinone acetate. J. Vet. Med. Sci. 1993, 55, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Sawada, T.; Tamada, H.; Inaba, T.; Mori, J. Prevention of estrus in the bitch with chlormadinone acetate administered orally. J. Vet. Med. Sci. 1992, 54, 595–596. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schäfer-Somi, S.; Kaya, D.; Sözmen, M.; Kaya, S.; Aslan, S. Pre-pubertal treatment with a Gn RH agonist in bitches—Effect on the uterus and hormone receptor expression. Reprod. Domest. Anim. 2018, 53, 103–109. [Google Scholar] [CrossRef]
- Sung, M.; Armour, A.; Wright, P. The influence of exogenous progestin on the occurrence of proestrous or estrous signs, plasma concentrations of luteinizing hormone and estradiol in deslorelin (GnRH agonist) treated anestrous bitches. Theriogenology 2006, 66, 1513–1517. [Google Scholar] [CrossRef]
- Valiente, C.; Diaz, J.; Rosa, D.; Mattioli, G.; Romero, G.G.; Gobello, C. Effect of a GnRH antagonist on GnRH agonist–implanted anestrous bitches. Theriogenology 2009, 72, 926–929. [Google Scholar] [CrossRef] [PubMed]
- Valiente, C.; Romero, G.G.; Corrada, Y.; De la Sota, P.; Hermo, G.; Gobello, C. Interruption of the canine estrous cycle with a low and a high dose of the GnRH antagonist, acyline. Theriogenology 2009, 71, 408–411. [Google Scholar] [CrossRef]
- Concannon, P.; Weinstein, P.; Whaley, S.; Frank, D. Suppression of luteal function in dogs by luteinizing hormone antiserum and by bromocriptine. Reproduction 1987, 81, 175–180. [Google Scholar] [CrossRef]
- De Gier, J.; Wolthers, C.; Galac, S.; Okkens, A.; Kooistra, H. Effects of the 3β-hydroxysteroid dehydrogenase inhibitor trilostane on luteal progesterone production in the dog. Theriogenology 2011, 75, 1271–1279. [Google Scholar] [CrossRef]
- Janowski, T.; Fingerhut, J.; Kowalewski, M.P.; Zduńczyk, S.; Domosławska, A.; Jurczak, A.; Boos, A.; Schuler, G.; Hoffmann, B. In vivo investigations on luteotropic activity of prostaglandins during early diestrus in nonpregnant bitches. Theriogenology 2014, 82, 915–920. [Google Scholar] [CrossRef]
- Kowalewski, M.; Ihle, S.; Siemieniuch, M.; Gram, A.; Boos, A.; Zduńczyk, S.; Fingerhut, J.; Hoffmann, B.; Schuler, G.; Jurczak, A. Formation of the early canine CL and the role of prostaglandin E2 (PGE2) in regulation of its function: An in vivo approach. Theriogenology 2015, 83, 1038–1047. [Google Scholar] [CrossRef]
- Onclin, K.; Verstegen, J. In vivo investigation of luteal function in dogs: Effects of cabergoline, a dopamine agonist, and prolactin on progesterone secretion during mid-pregnancy and-diestrus. Domest. Anim. Endocrinol. 1997, 14, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.T.; Gram, A.; Nowaczyk, R.; Boos, A.; Hoffmann, B.; Janowski, T.; Kowalewski, M.P. Prostaglandin-mediated effects in early canine corpus luteum: In vivo effects on vascular and immune factors. Reprod. Biol. 2019, 19, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Albers-Wolthers, C.; De Gier, J.; Rutten, V.; van Kooten, P.; Leegwater, P.; Schaefers-Okkens, A.; Kooistra, H. The effects of kisspeptin agonist canine KP-10 and kisspeptin antagonist p271 on plasma LH concentrations during different stages of the estrous cycle and anestrus in the bitch. Theriogenology 2016, 86, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Albers-Wolthers, C.; De Gier, J.; Walen, M.; Van Kooten, P.; Lambalk, C.; Leegwater, P.; Roelen, B.; Schaefers-Okkens, A.; Rutten, V.; Millar, R. In vitro and in vivo effects of kisspeptin antagonists p234, p271, p354, and p356 on GPR54 activation. PLoS ONE 2017, 12, e0179156. [Google Scholar] [CrossRef]
- Albers-Wolthers, K.H.; De Gier, J.; Kooistra, H.S.; Rutten, V.P.; Van Kooten, P.J.; De Graaf, J.J.; Leegwater, P.A.; Millar, R.P.; Schaefers-Okkens, A.C. Identification of a novel kisspeptin with high gonadotrophin stimulatory activity in the dog. Neuroendocrinology 2014, 99, 178–189. [Google Scholar] [CrossRef]
- Terse, P.S.; Peggins, J.; Seminara, S.B. Safety Evaluation of KP-10 (Metastin 45–54) Following once Daily Intravenous Administration for 14 Days in Dog. Int. J. Toxicol. 2021, 40, 337–343. [Google Scholar] [CrossRef]
- Mogheiseh, A.; Khafi, M.S.A.; Ahmadi, N.; Farkhani, S.R.; Bandariyan, E. Ultrasonographic and histopathologic changes following injection of neutral zinc gluconate in dog’s ovaries. Comp. Clin. Pathol. 2017, 26, 1093–1098. [Google Scholar] [CrossRef]
- Rajabi, A.; Mogheiseh, A.; Nazifi, S.; Ahrari-Khafi, M.; Dehghanian, A.; Vesal, N.; Bigham-Sadegh, A. Effect of direct therapeutic ultrasound exposure of ovaries on histopathology, inflammatory response, and oxidative stress in dogs. BMC Vet. Res. 2023, 19, 88. [Google Scholar] [CrossRef]
- Schaut, R.; Jackman, J.; Brewer, M.; Mendoza, K.; Jones, D.; Narasimhan, B. A polyanhydride-based implant. Single dose vaccine Platf. long-term immunity. Vaccine 2018, 36, 1024–1025. [Google Scholar] [CrossRef]
- Schaut, R.G.; Brewer, M.T.; Hostetter, J.M.; Mendoza, K.; Vela-Ramirez, J.E.; Kelly, S.M.; Jackman, J.K.; Dell’Anna, G.; Howard, J.M.; Narasimhan, B. A single dose polyanhydride-based vaccine platform promotes and maintains anti-GnRH antibody titers. Vaccine 2018, 36, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Samoylov, A.; Napier, I.; Morrison, N.; Cochran, A.; Schemera, B.; Wright, J.; Cattley, R.; Samoylova, T. DNA Vaccine Targeting Gonadotropin-Releasing Hormone Receptor and Its Application in Animal Contraception. Mol. Biotechnol. 2019, 61, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Sandam, N.P.; Prakash, D.; Thimmareddy, P. Immunocontraceptive potential of a GnRH receptor-based fusion recombinant protein. J. Genet. Eng. Biotechnol. 2021, 19, 63. [Google Scholar] [CrossRef] [PubMed]
- Vasetska, A. Non-surgical methods of regulation reproductive function and contraception males of domestic animals. Ukr. J. Vet. Agric. Sci. 2020, 3, 44–50. [Google Scholar] [CrossRef]
- Samoylova, T.I.; Braden, T.D.; Spencer, J.A.; Bartol, F.F. Immunocontraception: Filamentous Bacteriophage as a Platform for Vaccine Development. Curr. Med. Chem. 2017, 24, 3907–3920. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.K.; Jones, R.L.; Kraneburg, C.J.; Cochran, A.M.; Samoylov, A.M.; Wright, J.C.; Hutchinson, C.; Picut, C.; Cattley, R.C.; Martin, D.R.; et al. Phage constructs targeting gonadotropin-releasing hormone for fertility control: Evaluation in cats. J. Feline. Med. Surg. 2020, 22, 685–695. [Google Scholar] [CrossRef]
- Risso, A.; Corrada, Y.; Barbeito, C.; Díaz, J.D.; Gobello, C. Long-term-release GnRH agonists postpone puberty in domestic cats. Reprod. Domest. Anim. 2012, 47, 936–938. [Google Scholar] [CrossRef]
- Trigg, T.; Wright, P.; Armour, A.; Williamson, P.; Junaidi, A.; Martin, G.; Doyle, A.; Walsh, J. Use of a GnRH analogue implant to produce reversible long-term suppression of reproductive function in male and female domestic dogs. J. Reprod. Fertil. Suppl. 2001, 57, 255–261. [Google Scholar]
- Gobello, C. New GnRH analogs in canine reproduction. Anim. Reprod. Sci. 2007, 100, 1–13. [Google Scholar] [CrossRef]
- Tamada, H.; Kawate, N.; Inaba, T.; Sawada, T. Long-term prevention of estrus in the bitch and queen using chlormadinone acetate. Can. Vet. J. 2003, 44, 416. [Google Scholar]
- Fraser, B.; Wilkins, A.; Whiting, S.; Liang, M.; Rebourcet, D.; Nixon, B.; Aitken, R.J. Development of peptides for targeting cell ablation agents concurrently to the Sertoli and Leydig cell populations of the testes: An approach to non-surgical sterilization. PLoS ONE 2024, 19, e0292198. [Google Scholar] [CrossRef] [PubMed]
- Faya, M.; Marchetti, C.; Priotto, M.; Grisolía, M.; D’Francisco, F.; Gobello, C. Postponement of canine puberty by neonatal administration of a long term release GnRH superagonist. Theriogenology 2018, 118, 190–195. [Google Scholar] [CrossRef] [PubMed]
- De Roux, N.; Genin, E.; Carel, J.-C.; Matsuda, F.; Chaussain, J.-L.; Milgrom, E. Hypogonadotropic hypogonadism due to loss of function of the KiSS1-derived peptide receptor GPR54. Proc. Natl. Acad. Sci. USA 2003, 100, 10972–10976. [Google Scholar] [CrossRef]
- Seminara, S.B.; Messager, S.; Chatzidaki, E.E.; Thresher, R.R.; Acierno, J.S., Jr.; Shagoury, J.K.; Bo-Abbas, Y.; Kuohung, W.; Schwinof, K.M.; Hendrick, A.G. The GPR54 gene as a regulator of puberty. N. Engl. J. Med. 2003, 349, 1614–1627. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Hernández, J.M.; Martin, G.B.; Becerril-Pérez, C.M.; Pro-Martínez, A.; Cortez-Romero, C.; Gallegos-Sánchez, J. Kisspeptin stimulates the pulsatile secretion of luteinizing hormone (LH) during postpartum anestrus in ewes undergoing continuous and restricted suckling. Animals 2021, 11, 2656. [Google Scholar] [CrossRef]
- Colledge, W. Transgenic mouse models to study Gpr54/kisspeptin physiology. Peptides 2009, 30, 34–41. [Google Scholar] [CrossRef]
- Kotani, M.; Detheux, M.; Vandenbogaerde, A.; Communi, D.; Vanderwinden, J.-M.; Le Poul, E.; Brézillon, S.; Tyldesley, R.; Suarez-Huerta, N.; Vandeput, F. The metastasis suppressor gene KiSS-1 encodes kisspeptins, the natural ligands of the orphan G protein-coupled receptor GPR54. J. Biol. Chem. 2001, 276, 34631–34636. [Google Scholar] [CrossRef]
- Clarke, I.J. Control of GnRH secretion: One step back. Front. Neuroendocrinol. 2011, 32, 367–375. [Google Scholar] [CrossRef]
- Pineda, R.; Garcia-Galiano, D.; Roseweir, A.; Romero, M.; Sanchez-Garrido, M.; Ruiz-Pino, F.; Morgan, K.; Pinilla, L.; Millar, R.; Tena-Sempere, M. Critical roles of kisspeptins in female puberty and preovulatory gonadotropin surges as revealed by a novel antagonist. Endocrinology 2010, 151, 722–730. [Google Scholar] [CrossRef]
- Fan-Mei, Z.; Yi, D.; Teketay, W.; Hai-Jing, J.; Sohail, A.; Gui-Qiong, L.; Xun-Ping, J. Recent advances in immunocastration in sheep and goat and its animal welfare benefits: A review. J. Integr. Agric. 2022, 21, 299–309. [Google Scholar] [CrossRef]
- Kano, M.; Sosulski, A.E.; Zhang, L.; Saatcioglu, H.D.; Wang, D.; Nagykery, N.; Sabatini, M.E.; Gao, G.; Donahoe, P.K.; Pépin, D. AMH/MIS as a contraceptive that protects the ovarian reserve during chemotherapy. Proc. Natl. Acad. Sci. USA 2017, 114, E1688–E1697. [Google Scholar] [CrossRef] [PubMed]
- Bolghanabadi, M.; Sedigh, H.S.; Mirshokraei, P.; Rajabioun, M. Simultaneously administration of cabergoline and PMSG reduces the duration of estrus induction in anestrous bitches. Vet. Res. Forum 2023, 14, 665–671. [Google Scholar]
- Dissen, G.A.; Lomniczi, A.; Boudreau, R.L.; Chen, Y.H.; Davidson, B.L.; Ojeda, S.R. Targeted gene silencing to induce permanent sterility. Reprod. Domest. Anim. 2012, 47, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Naso, M.F.; Tomkowicz, B.; Perry III, W.L.; Strohl, W.R. Adeno-associated virus (AAV) as a vector for gene therapy. BioDrugs 2017, 31, 317–334. [Google Scholar] [CrossRef]
- Avilez Lopez, E.J.; Cuadra Palacios, J.D. Comparación de Dos Técnicas Quirúrgicas, para Ovariohisterectomía Felina en Clínica Veterinaria Mimos. Bachelor’s Thesis, Universidad Nacional Agraria, La Molina, Peru, 2020. [Google Scholar]
- Iglesias, B.A.L.; Argüello, V.F.; Rodríguez, J.G.; Hernandez, A.A. Técnica de ovariectomía con mínima invasión en caninos en México. Jóvenes Cienc. 2022, 14, 1–11. [Google Scholar] [CrossRef]
- Ealo, I.M.; Berna, I.L.; Eugenia, M. Esterilización en Hembras de la Especie Canina: Ovariectomía vs. Ovariohisterectomía. Bachelor’s Thesis, Universidad Zaragoza Facultad de Veterinaria, Zaragoza, Spain, 2020. [Google Scholar]
- Masache, J.L.; Brito, M.C.; Sagbay, C.F.; Webster, P.G.; Garnica, F.P.; Mínguez, C. Ovariectomía en perras: Comparación entre el abordaje medial o lateral. Rev. Investig. Vet. Del. Perú 2016, 27, 309–315. [Google Scholar] [CrossRef]
- Little, S.E. Female reproduction. Cat 2011, 1195–1227. [Google Scholar]
- Wigham, E.E.; Moxon, R.S.; England, G.C.; Wood, J.L.; Morters, M.K. Seasonality in oestrus and litter size in an assistance dog breeding colony in the United Kingdom. Vet. Rec. 2017, 181, 371. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).