
Mitochondrial Variation of Bottlenose Dolphins (Tursiops truncatus) from the Canary Islands Suggests a Key Population for Conservation with High Connectivity within the North-East Atlantic Ocean
 ,
,  , , , and
, , , and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
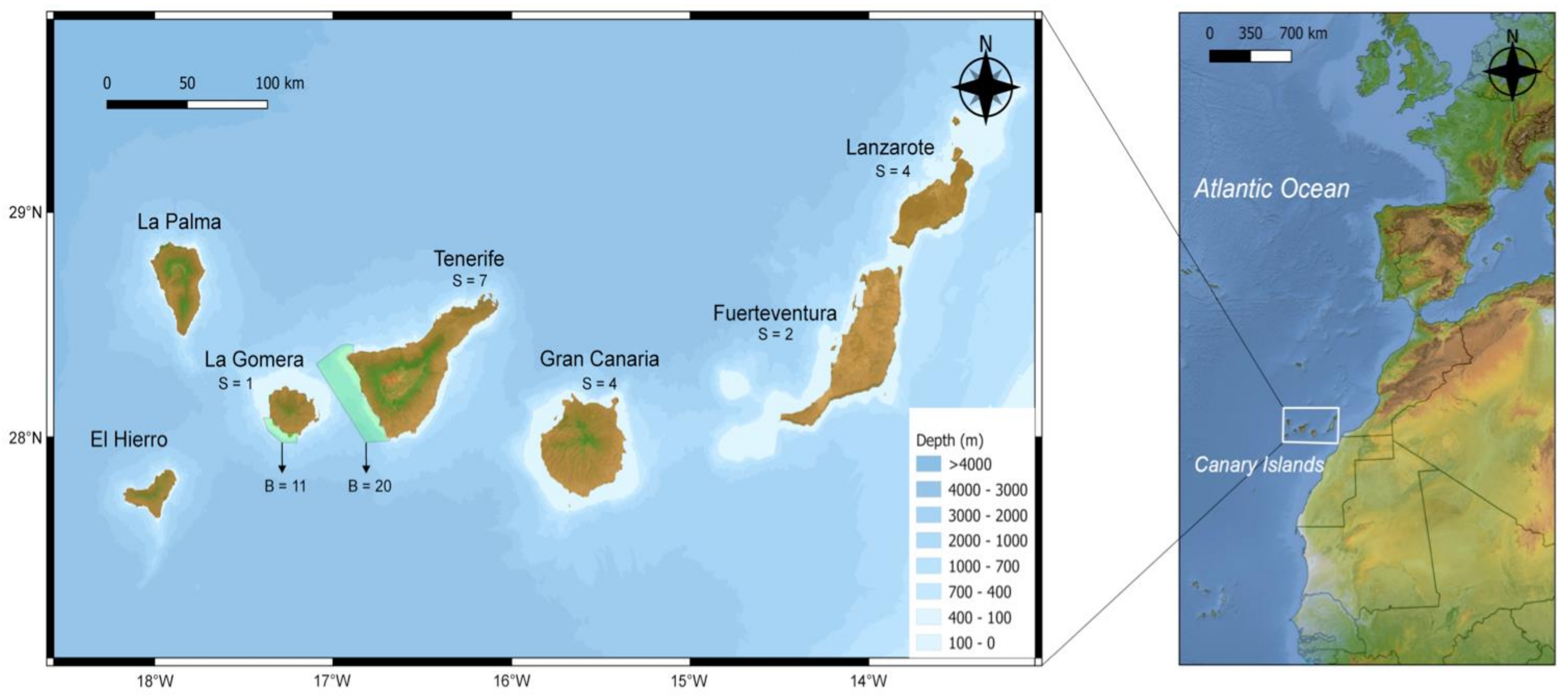
2.1. Sample Collection
2.2. Genetic Analysis: Population Structure and Diversity in the Canary Islands
2.3. Genetic Analysis: Population Structure and Diversity in the NEAO
3. Results
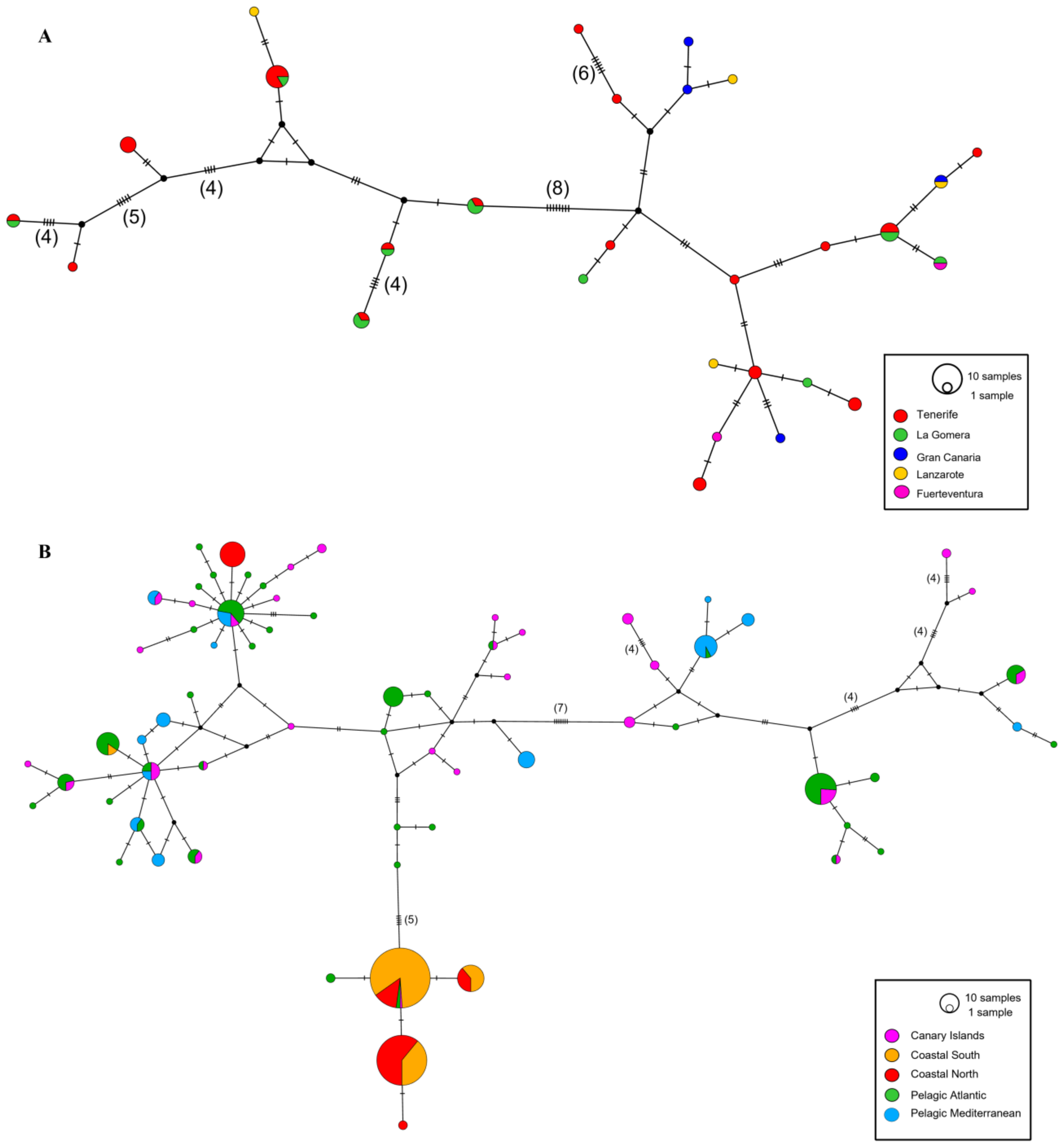
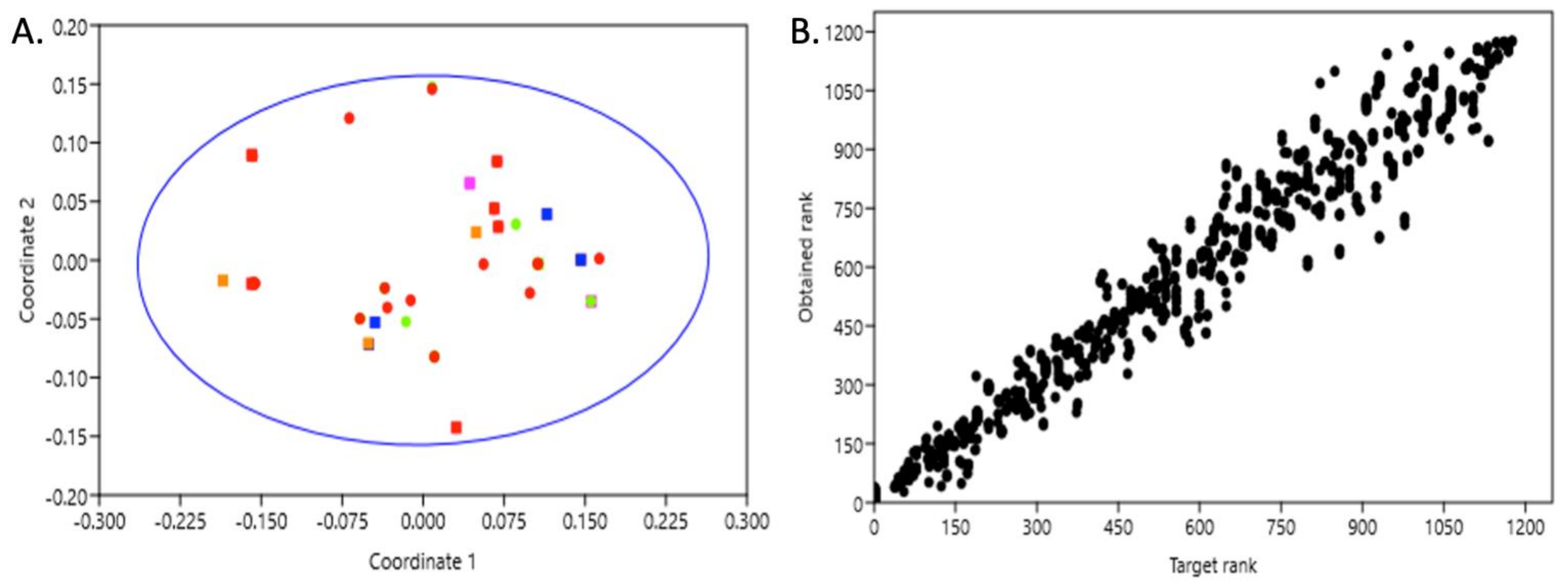
3.1. Population Structure and Genetic Diversity in the Canary Islands
3.2. Population Structure and Genetic Diversity in the Canary Islands
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wells, R.S.; Scott, M.D. Common Bottlenose Dolphin: Tursiops truncatus. In Encyclopedia of Marine Mammals, 2nd ed.; Perrin, W.F., Würsig, B., Thewissen, J.G.M., Eds.; Academic Press: London, UK, 2009; pp. 249–255. ISBN 978-0-12-373553-9. [Google Scholar]
- Wells, R.S.; Rhinehart, H.L.; Hansen, L.J.; Sweeney, J.C.; Townsend, F.I.; Stone, R.; Casper, D.R.; Scott, M.D.; Hohn, A.A.; Rowles, T.K. Bottlenose Dolphins as Marine Ecosystem Sentinels: Developing a Health Monitoring System. EcoHealth 2004, 1, 246–254. [Google Scholar] [CrossRef]
- Torres, L.G.; Read, A.J.; Halpin, P. Fine-Scale Habitat Modeling of a Top Marine Predator: Do Prey Data Improve Predictive Capacity. Ecol. Appl. 2008, 18, 1702–1717. [Google Scholar] [CrossRef]
- Ballance, L.T. Habitat Use Patterns and Ranges of the Bottlenose Dolphin in the Gulf of California, Mexico. Mar. Mammal Sci. 1992, 8, 262–274. [Google Scholar] [CrossRef]
- Dos Santos, M.E.; Louro, S.; Couchinho, M.; Brito, C. Whistles of Bottlenose Dolphins (Tursiops truncatus) in the Sado Estuary, Portugal: Characteristics, Production Rates, and Long-Term Contour Stability. Aquat. Mamm. 2005, 31, 453–462. [Google Scholar] [CrossRef]
- Wells, R.; Natoli, A.; Braulik, G. Tursiops truncatus (Errata Version Published in 2019). In The IUCN Red List of Threatened Species; IUCN: Cambridge, UK, 2019. [Google Scholar] [CrossRef]
- Natoli, A.; Genov, T.; Kerem, D.; Gonzalvo, J.; Lauriano, G.; Holcer, D.; Labach, H.; Marsili, L.; Mazzariol, S.; Moura, A.E.; et al. Tursiops truncatus (Mediterranean Subpopulation) (Errata Version Published in 2022). In The IUCN Red List of Threatened Species; IUCN: Cambridge, UK, 2021. [Google Scholar]
- Currey, R.J.C.; Dawson, S.M.; Slooten, E. An Approach for Regional Threat Assessment under IUCN Red List Criteria That Is Robust to Uncertainty: The Fiordland Bottlenose Dolphins Are Critically Endangered. Biol. Conserv. 2009, 142, 1570–1579. [Google Scholar] [CrossRef]
- Certain, G.; Ridoux, V.; van Canneyt, O.; Bretagnolle, V. Delphinid Spatial Distribution and Abundance Estimates over the Shelf of the Bay of Biscay. ICES J. Mar. Sci. 2008, 65, 656–666. [Google Scholar] [CrossRef]
- Hammond, P.S.; Macleod, K.; Berggren, P.; Borchers, D.L.; Burt, L.; Cañadas, A.; Desportes, G.; Donovan, G.P.; Gilles, A.; Gillespie, D.; et al. Cetacean Abundance and Distribution in European Atlantic Shelf Waters to Inform Conservation and Management. Biol. Conserv. 2013, 164, 107–122. [Google Scholar] [CrossRef]
- Walker, W.A. Geographical Variation in Morphology and Biology of Bottlenose Dolphins (Tursiops) in the Eastern North Pacific; NOAA Administrative Report LJ-81-0003c.; National Marine Fisheries Service, Southwest Fisheries Center: La Jolla, CA, USA, 1981. [Google Scholar]
- Mead, J.G.; Potter, C.W. Recognizing Two Populations off the Bottlenose Dolphin (Tursiops truncatus) of the Atlantic Coast of North America—Morphologic and Ecologic Considerations. In International Marine Biology Research Institute: IBI Reports; International Marine Biology Research Institute: Kamogawa, Japan, 1995; pp. 31–44. [Google Scholar]
- Bearzi, M.; Saylan, C.A.; Hwang, A. Ecology and Comparison of Coastal and Offshore Bottlenose Dolphins (Tursiops truncatus) in California. Mar. Freshw. Res. 2009, 60, 584–593. [Google Scholar] [CrossRef]
- Perrin, W.F.; Thieleking, J.L.; Walker, W.A.; Archer, F.I.; Robertson, K.M. Common Bottlenose Dolphins (Tursiops truncatus) in California Waters: Cranial Differentiation of Coastal and Offshore Ecotypes. Mar. Mammal Sci. 2011, 27, 769–792. [Google Scholar] [CrossRef]
- Oudejans, M.G.; Visser, F.; Englund, A.; Rogan, E.; Ingram, S.N. Evidence for Distinct Coastal and Offshore Communities of Bottlenose Dolphins in the North East Atlantic. PLoS ONE 2015, 10, e0122668. [Google Scholar] [CrossRef]
- Hoelzel, A.R.; Potter, C.W.; Best, P.B. Genetic Differentiation between Parapatric ‘Nearshore’ and ‘Offshore’ Populations of the Bottlenose Dolphin. Proc. R. Soc. Lond. B Biol. Sci. 1998, 265, 1177–1183. [Google Scholar] [CrossRef]
- Natoli, A.; Birkun, A.; Aguilar, A.; Lopez, A.; Hoelzel, A.R. Habitat Structure and the Dispersal of Male and Female Bottlenose Dolphins (Tursiops truncatus). Proc. R. Soc. B Biol. Sci. 2005, 272, 1217–1226. [Google Scholar] [CrossRef]
- Quérouil, S.; Silva, M.A.; Freitas, L.; Prieto, R.; Magalhães, S.; Dinis, A.; Alves, F.; Matos, J.A.; Mendonça, D.; Hammond, P.S.; et al. High Gene Flow in Oceanic Bottlenose Dolphins (Tursiops truncatus) of the North Atlantic. Conserv. Genet. 2007, 8, 1405–1419. [Google Scholar] [CrossRef]
- Fruet, P.F.; Secchi, E.R.; Daura-Jorge, F.; Vermeulen, E.; Flores, P.A.C.; Simões-Lopes, P.C.; Genoves, R.C.; Laporta, P.; Di Tullio, J.C.; Freitas, T.R.O.; et al. Remarkably Low Genetic Diversity and Strong Population Structure in Common Bottlenose Dolphins (Tursiops truncatus) from Coastal Waters of the Southwestern Atlantic Ocean. Conserv. Genet. 2014, 15, 879–895. [Google Scholar] [CrossRef]
- Louis, M.; Viricel, A.; Lucas, T.; Peltier, H.; Alfonsi, E.; Berrow, S.; Brownlow, A.; Covelo, P.; Dabin, W.; Deaville, R.; et al. Habitat-Driven Population Structure of Bottlenose Dolphins, Tursiops truncatus, in the North-East Atlantic. Mol. Ecol. 2014, 23, 857–874. [Google Scholar] [CrossRef]
- Louis, M.; Fontaine, M.C.; Spitz, J.; Schlund, E.; Dabin, W.; Deaville, R.; Caurant, F.; Cherel, Y.; Guinet, C.; Simon-Bouhet, B. Ecological Opportunities and Specializations Shaped Genetic Divergence in a Highly Mobile Marine Top Predator. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141558. [Google Scholar] [CrossRef] [PubMed]
- Gaspari, S.; Scheinin, A.; Holcer, D.; Fortuna, C.; Natali, C.; Genov, T.; Frantzis, A.; Chelazzi, G.; Moura, A.E. Drivers of Population Structure of the Bottlenose Dolphin (Tursiops truncatus) in the Eastern Mediterranean Sea. Evol. Biol. 2015, 42, 177–190. [Google Scholar] [CrossRef]
- Nykänen, M.; Louis, M.; Dillane, E.; Alfonsi, E.; Berrow, S.; O’Brien, J.; Brownlow, A.; Covelo, P.; Dabin, W.; Deaville, R.; et al. Fine-Scale Population Structure and Connectivity of Bottlenose Dolphins, Tursiops truncatus, in European Waters and Implications for Conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 197–211. [Google Scholar] [CrossRef]
- Wiszniewski, J.; Allen, S.J.; Möller, L.M. Social Cohesion in a Hierarchically Structured Embayment Population of Indo-Pacific Bottlenose Dolphins. Anim. Behav. 2009, 77, 1449–1457. [Google Scholar] [CrossRef]
- Mirimin, L.; Miller, R.; Dillane, E.; Berrow, S.D.; Ingram, S.; Cross, T.F.; Rogan, E. Fine-Scale Population Genetic Structuring of Bottlenose Dolphins in Irish Coastal Waters. Anim. Conserv. 2011, 14, 342–353. [Google Scholar] [CrossRef]
- Allen, S.J.; Bryant, K.A.; Kraus, R.H.S.; Loneragan, N.R.; Kopps, A.M.; Brown, A.M.; Gerber, L.; Krützen, M. Genetic Isolation between Coastal and Fishery-impacted, Offshore Bottlenose Dolphin (Tursiops spp.) Populations. Mol. Ecol. 2016, 25, 2735–2753. [Google Scholar] [CrossRef] [PubMed]
- Fernández, R.; Santos, M.B.; Pierce, G.J.; Llavona, Á.; López, A.; Silva, M.A.; Ferreira, M.; Carrillo, M.; Cermeño, P.; Lens, S.; et al. Fine-Scale Genetic Structure of Bottlenose Dolphins, Tursiops truncatus, in Atlantic Coastal Waters of the Iberian Peninsula. Hydrobiologia 2011, 670, 111–125. [Google Scholar] [CrossRef]
- Nichols, C.; Herman, J.; Gaggiotti, O.E.; Dobney, K.M.; Parsons, K.; Hoelzel, A.R. Genetic Isolation of a Now Extinct Population of Bottlenose Dolphins (Tursiops truncatus). Proc. R. Soc. B Biol. Sci. 2007, 274, 1611–1616. [Google Scholar] [CrossRef]
- Crespo, E.A.; Notarbartolo di Sciara, G.; Reeves, R.R.; Smith, B.D. Dolphins, Whales and Porpoises: 2002–2010 Conservation Action Plan for the World’s Cetaceans; IUCN: Gland, UK, 2003; ISBN 978-2-8317-0656-6. [Google Scholar]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. Introduction to Conservation Genetics. Available online: https://www.cambridge.org/highereducation/books/introduction-to-conservation-genetics/696B4E558C93F7FBF9C33D6358EA7425 (accessed on 19 December 2023).
- Hare, M.P.; Nunney, L.; Schwartz, M.K.; Ruzzante, D.E.; Burford, M.; Waples, R.S.; Ruegg, K.; Palstra, F. Understanding and Estimating Effective Population Size for Practical Application in Marine Species Management. Conserv. Biol. 2011, 25, 438–449. [Google Scholar] [CrossRef]
- Troll, V.R.; Carracedo, J.C. Chapter 1—The Canary Islands: An Introduction. In The Geology of the Canary Islands; Troll, V.R., Carracedo, J.C., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 1–41. ISBN 978-0-12-809663-5. [Google Scholar]
- Correia, A.M.; Gil, Á.; Valente, R.F.; Rosso, M.; Sousa-Pinto, I.; Pierce, G.J. Distribution of Cetacean Species at a Large Scale—Connecting Continents with the Macaronesian Archipelagos in the Eastern North Atlantic. Divers. Distrib. 2020, 26, 1234–1247. [Google Scholar] [CrossRef]
- Tobeña, M.; Escánez, A.; Rodríguez, Y.; López, C.; Ritter, F.; Aguilar, N. Inter-Island Movements of Common Bottlenose Dolphins Tursiops truncatus among the Canary Islands: Online Catalogues and Implications for Conservation and Management. Afr. J. Mar. Sci. 2014, 36, 137–141. [Google Scholar] [CrossRef]
- Dinis, A.; Molina, C.; Tobeña, M.; Sambolino, A.; Hartman, K.; Fernandez, M.; Magalhães, S.; dos Santos, R.P.; Ritter, F.; Martín, V.; et al. Large-Scale Movements of Common Bottlenose Dolphins in the Atlantic: Dolphins with an International Courtyard. PeerJ 2021, 9, e11069. [Google Scholar] [CrossRef]
- Rosel, P.E. PCR-based sex determination in Odontocete cetaceans. Conserv. Gen. 2003, 4, 647–649. [Google Scholar] [CrossRef]
- Dalebout, M.L.; Robertson, K.M.; Frantzis, A.; Engelhaupt, D.; Mignucci-Giannoni, A.A.; Rosario-Delestre, R.J.; Baker, C.S. Worldwide Structure of mtDNA Diversity among Cuvier’s Beaked Whales (Ziphius cavirostris): Implications for Threatened Populations. Mol. Ecol. 2005, 14, 3353–3371. [Google Scholar] [CrossRef]
- Miralles, L.; Lens, S.; Rodríguez-Folgar, A.; Carrillo, M.; Martín, V.; Mikkelsen, B.; Garcia-Vazquez, E. Interspecific Introgression in Cetaceans: DNA Markers Reveal Post-F1 Status of a Pilot Whale. PLoS ONE 2013, 8, e69511. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Oxford University Press: Oxford, UK, 1999; Volume 41, pp. 95–98. [Google Scholar]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (Version 3.0): An Integrated Software Package for Population Genetics Data Analysis. Evol. Bioinform. 2005, 1. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-Joining Networks for Inferring Intraspecific Phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.W.; Bryant, D. Popart: Full-Feature Software for Haplotype Network Construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.; Ryan, P. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Tamura, K. Estimation of the Number of Nucleotide Substitutions When There Are Strong Transition-Transversion and G+C-Content Biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; Stevens, M.; Wagner, H. Vegan: Community Ecology Package. R Package Version. 2.0-10; CRAN: Vienna, Austria, 2013. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Wells, R.S.; Rhinehart, H.L.; Cunningham, P.; Whaley, J.; Baran, M.; Koberna, C.; Costa, D.P. Long Distance Offshore Movements of Bottlenose Dolphins. Mar. Mammal Sci. 1999, 15, 1098–1114. [Google Scholar] [CrossRef]
- Genov, T.; Železnik, J.; Bruno, C.; Ascheri, D.; Fontanesi, E.; Blasi, M.F. The Longest Recorded Movement of an Inshore Common Bottlenose Dolphin (Tursiops truncatus). Mamm. Biol. 2022, 102, 1469–1481. [Google Scholar] [CrossRef]
- Bhatia, G.; Patterson, N.; Sankararaman, S.; Price, A.L. Estimating and Interpreting FST: The Impact of Rare Variants. Genome Res. 2013, 23, 1514–1521. [Google Scholar] [CrossRef]
- Holsinger, K.E.; Weir, B.S. Genetics in Geographically Structured Populations: Defining, Estimating and Interpreting FST. Nat. Rev. Genet. 2009, 10, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, S. Estructura Genética de las Poblaciones de Cetáceos del Archipiélago Canario: Secuenciación de la Región Control y los Genes COI y NADH5 del ADN Mitocondrial [Genetic Structure of the Cetacean Populations from the Canarian Archipelago: Sequencing of the Control Region and the COI and NADH5 Genes of the Mitochondrial DNA]. Ph.D. Dissertation, Universidad de Las Palmas de Gran Canaria, Las Palmas, Spain, 2002. [Google Scholar]
- Herrera, I.; Carrillo, M.; Cosme de Esteban, M.; Haroun, R. Distribution of cetaceans in the canary islands (Northeast Atlantic ocean): Implications for the natura 2000 network and future conservation measures. Front. Mar. Sci. 2021, 8, 669790. [Google Scholar] [CrossRef]
- Do Amaral, K.B.; Barragán-Barrera, D.C.; Mesa-Gutiérrez, R.A.; Farías-Curtidor, N.; Caballero Gaitán, S.J.; Méndez-Fernandez, P.; Oliveira Santos, M.C.; Rinaldi, C.; Rinaldi, R.; Siciliano, S.; et al. Seascape genetics of the Atlantic Spotted Dolphin (Stenella frontalis) based on mitochondrial DNA. J. Hered. 2021, 112, 646–662. [Google Scholar] [CrossRef]
- Hooker, S.K.; Cañadas, A.; Hyrenbach, K.D.; Corrigan, C.; Polovina, J.J.; Reeves, R.R. Making Protected Area Networks Effective for Marine Top Predators. Endanger. Species Res. 2011, 13, 203–218. [Google Scholar] [CrossRef]
- Allendorf, F.W.; Hohenlohe, P.A.; Luikart, G. Genomics and the Future of Conservation Genetics. Nat. Rev. Genet. 2010, 11, 697–709. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Populations | n | S | Nh | h | Hd (SD) | π (SD) | k |
|---|---|---|---|---|---|---|---|
| Tenerife | 27 | 35 | 17 | 11 | 0.952 (0.025) | 0.0166 (0.009) | 10.571 |
| La Gomera | 12 | 31 | 9 | 2 | 0.955 (0.047) | 0.0153 (0.008) | 9.713 |
| Lanzarote | 4 | 24 | 4 | 3 | 1.000 (0.177) | 0.0206 (0.014) | 13.121 |
| Gran Canaria | 4 | 16 | 4 | 3 | 1.000 (0.177) | 0.0149 (0.010) | 9.491 |
| Fuerteventura | 2 | 9 | 2 | 1 | 1.000 (0.500) | 0.0128 (0.014) | 8.122 |
| Total | 49 | 43 | 28 | / | 0.969 (0.011) | 0.0165 (0.009) | 10.509 |
| Populations | n | S | Nh | h | Hd (SD) | π (SD) | k |
|---|---|---|---|---|---|---|---|
| Canary Islands | 49 | 43 | 28 | 17 | 0.969 (0.011) | 0.0165 (0.009) | 10.503 |
| Coastal south | 115 | 12 | 4 | 0 | 0.499 (0.044) | 0.0014 (0.001) | 0.889 |
| Coastal north | 76 | 13 | 5 | 2 | 0.667 (0.042) | 0.0063 (0.003) | 4.028 |
| Pelagic Atlantic | 101 | 41 | 38 | 25 | 0.929 (0.013) | 0.0155 (0.007) | 9.881 |
| Pelagic Mediterranean | 51 | 28 | 15 | 8 | 0.902 (0.022) | 0.0137 (0.007) | 8.680 |
| Total | 392 | 56 | 70 | / | 0.905 (0.009) | 0.0140 (0.007) | 8.894 |
| Populations | Canary Islands | Coastal South | Coastal North | Pelagic Atlantic | Pelagic Mediterranean |
|---|---|---|---|---|---|
| Canary Islands | - | 0.291 ** | 0.191 ** | 0.015 | 0.057 ** |
| Coastal south | 0.635 ** | - | 0.252 ** | 0.279 ** | 0.328 ** |
| Coastal north | 0.401 ** | 0.233 ** | - | 0.195 ** | 0.222 ** |
| Pelagic Atlantic | 0.004 | 0.541 ** | 0.349 ** | - | 0.071 ** |
| Pelagic Mediterranean | 0.040 | 0.671 ** | 0.446 ** | 0.056 ** | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Lobo, D.A.; Monteoliva, A.P.; Fernandez, A.; Arbelo, M.; de la Fuente, J.; Pérez-Gil, M.; Varo-Cruz, N.; Servidio, A.; Pérez-Gil, E.; Borrell, Y.J.; et al. Mitochondrial Variation of Bottlenose Dolphins (Tursiops truncatus) from the Canary Islands Suggests a Key Population for Conservation with High Connectivity within the North-East Atlantic Ocean. Animals 2024, 14, 901. https://doi.org/10.3390/ani14060901
Gómez-Lobo DA, Monteoliva AP, Fernandez A, Arbelo M, de la Fuente J, Pérez-Gil M, Varo-Cruz N, Servidio A, Pérez-Gil E, Borrell YJ, et al. Mitochondrial Variation of Bottlenose Dolphins (Tursiops truncatus) from the Canary Islands Suggests a Key Population for Conservation with High Connectivity within the North-East Atlantic Ocean. Animals. 2024; 14(6):901. https://doi.org/10.3390/ani14060901
Chicago/Turabian StyleGómez-Lobo, Daniel A., Agustín P. Monteoliva, Antonio Fernandez, Manuel Arbelo, Jesús de la Fuente, Mónica Pérez-Gil, Nuria Varo-Cruz, Antonella Servidio, Enrique Pérez-Gil, Yaisel J. Borrell, and et al. 2024. "Mitochondrial Variation of Bottlenose Dolphins (Tursiops truncatus) from the Canary Islands Suggests a Key Population for Conservation with High Connectivity within the North-East Atlantic Ocean" Animals 14, no. 6: 901. https://doi.org/10.3390/ani14060901
APA StyleGómez-Lobo, D. A., Monteoliva, A. P., Fernandez, A., Arbelo, M., de la Fuente, J., Pérez-Gil, M., Varo-Cruz, N., Servidio, A., Pérez-Gil, E., Borrell, Y. J., & Miralles, L. (2024). Mitochondrial Variation of Bottlenose Dolphins (Tursiops truncatus) from the Canary Islands Suggests a Key Population for Conservation with High Connectivity within the North-East Atlantic Ocean. Animals, 14(6), 901. https://doi.org/10.3390/ani14060901